
Investigating Bergamot Essential Oil (BEO) Properties: Cytoprotection in Neuronal Cells Exposed to Heavy Metals and Antibacterial Activities


 ,
,  ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. GC-MS Analysis
2.3. Cell Culture
2.4. Evaluation of Cytotoxicity Using MTT Assay
2.5. Evaluation of Cell Viability by Means of Calcein-AM Assay
2.6. Lactate Dehydrogenase (LDH) Release
2.7. Measurement of Intracellular ROS Production
2.8. Antibacterial Studies
2.9. Statistical Analysis
3. Results
3.1. GC-MS Analysis
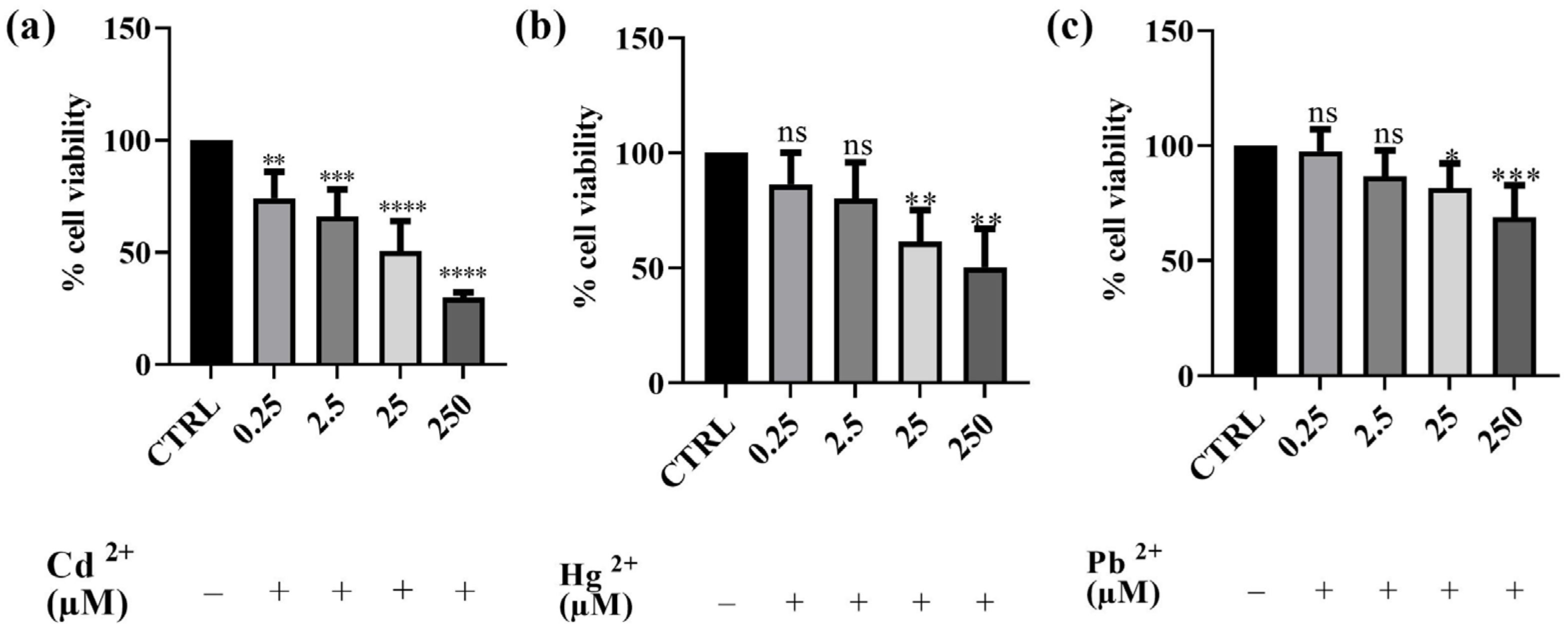
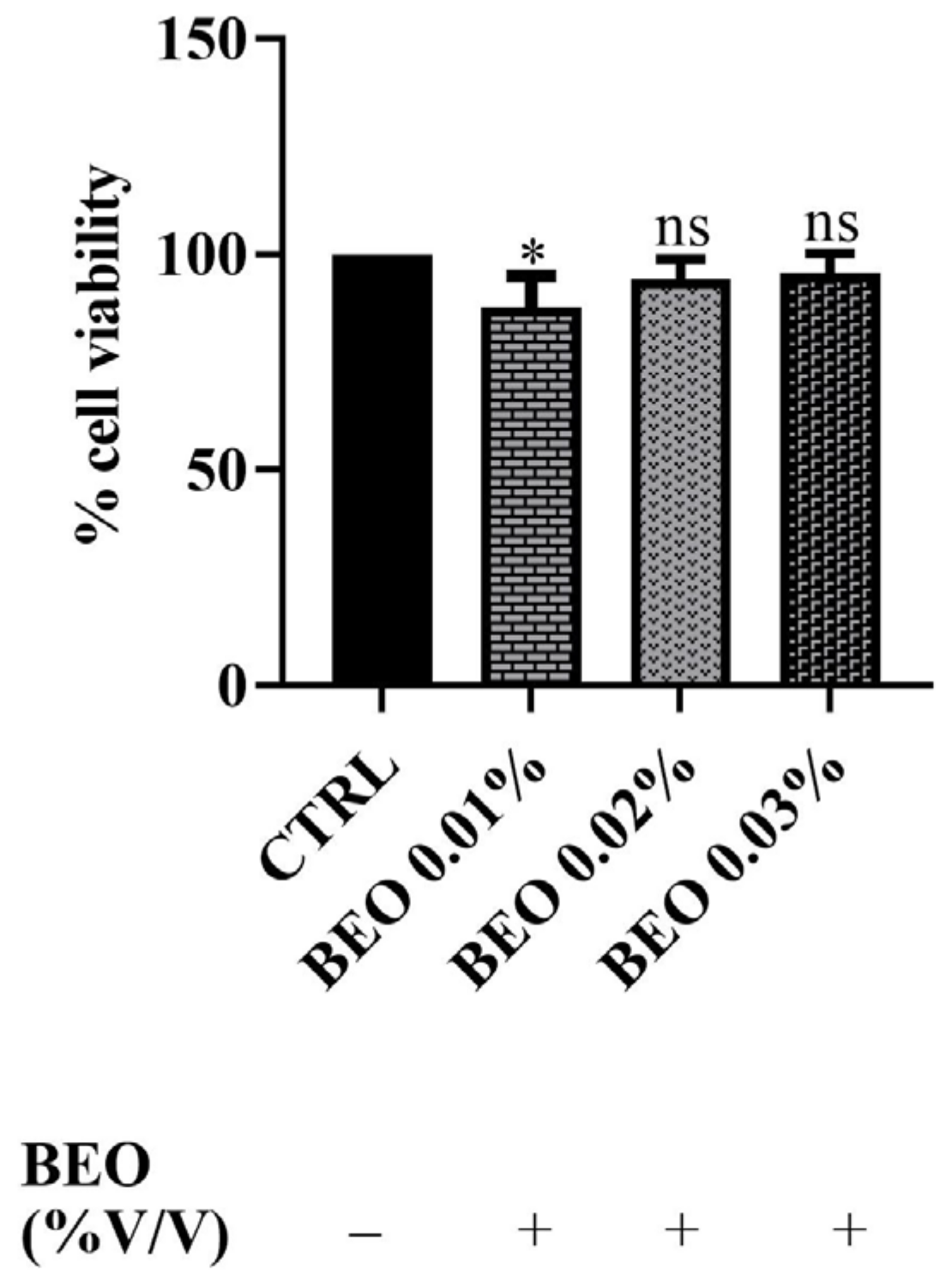
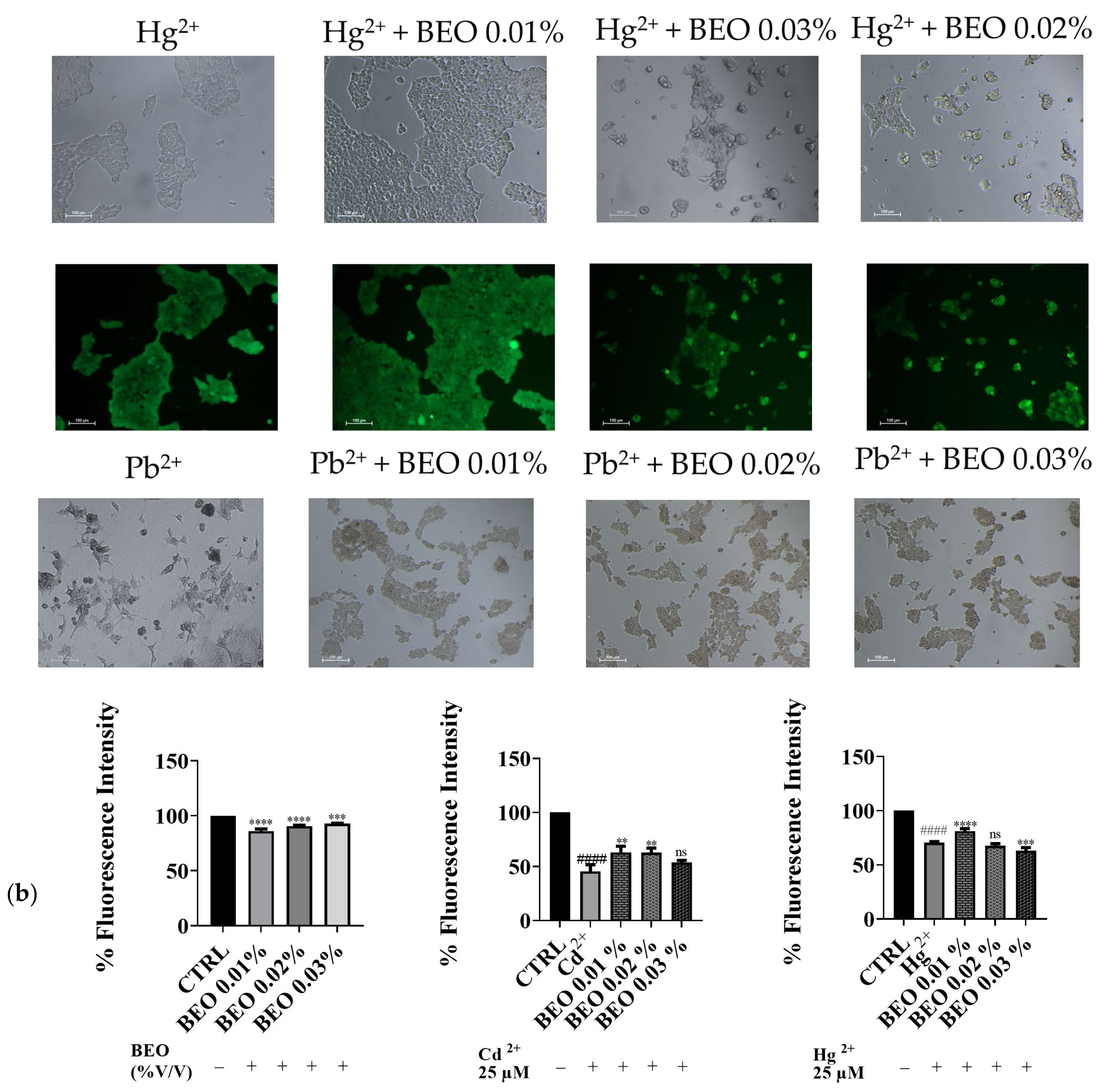
3.2. Effects of Cd2+, Hg2+, Pb2+, and BEO on Cell Viability
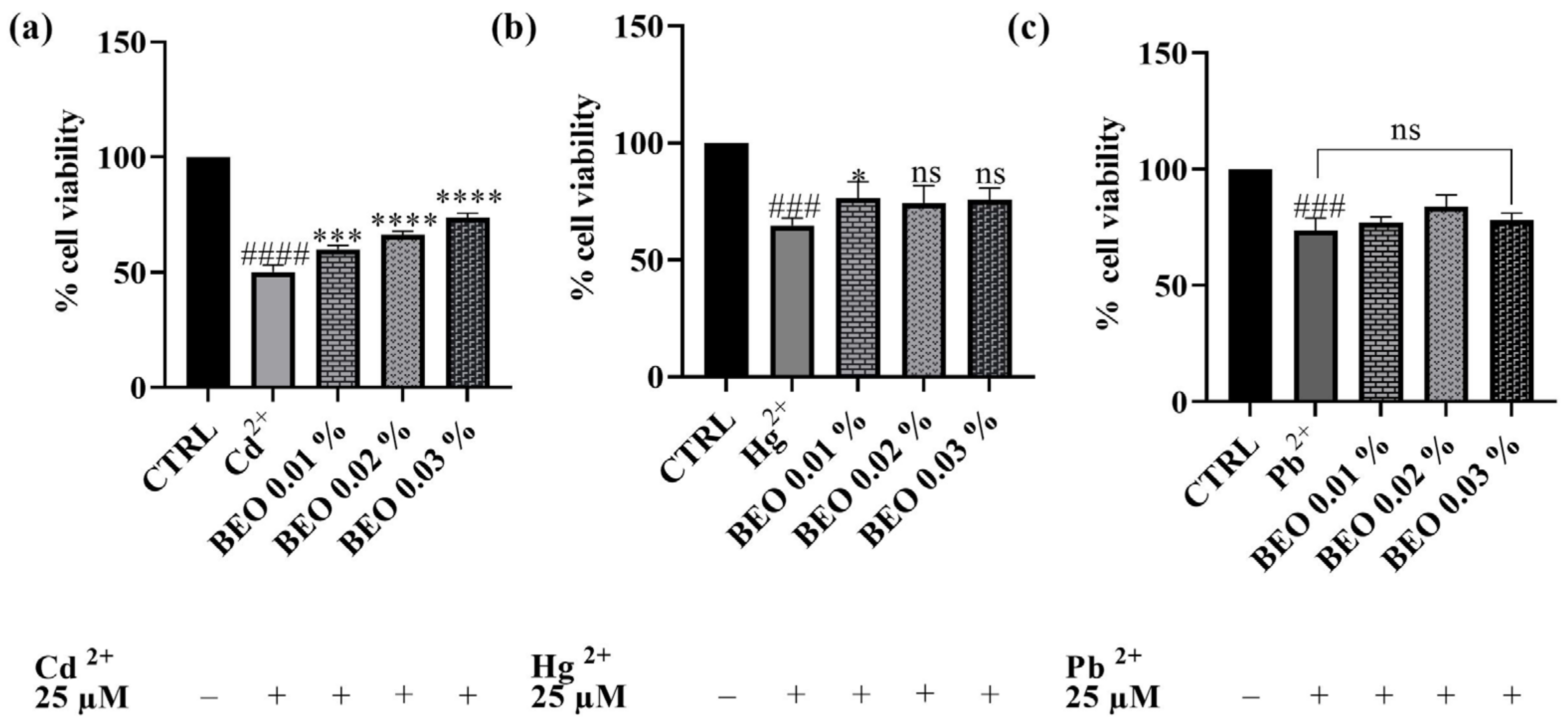
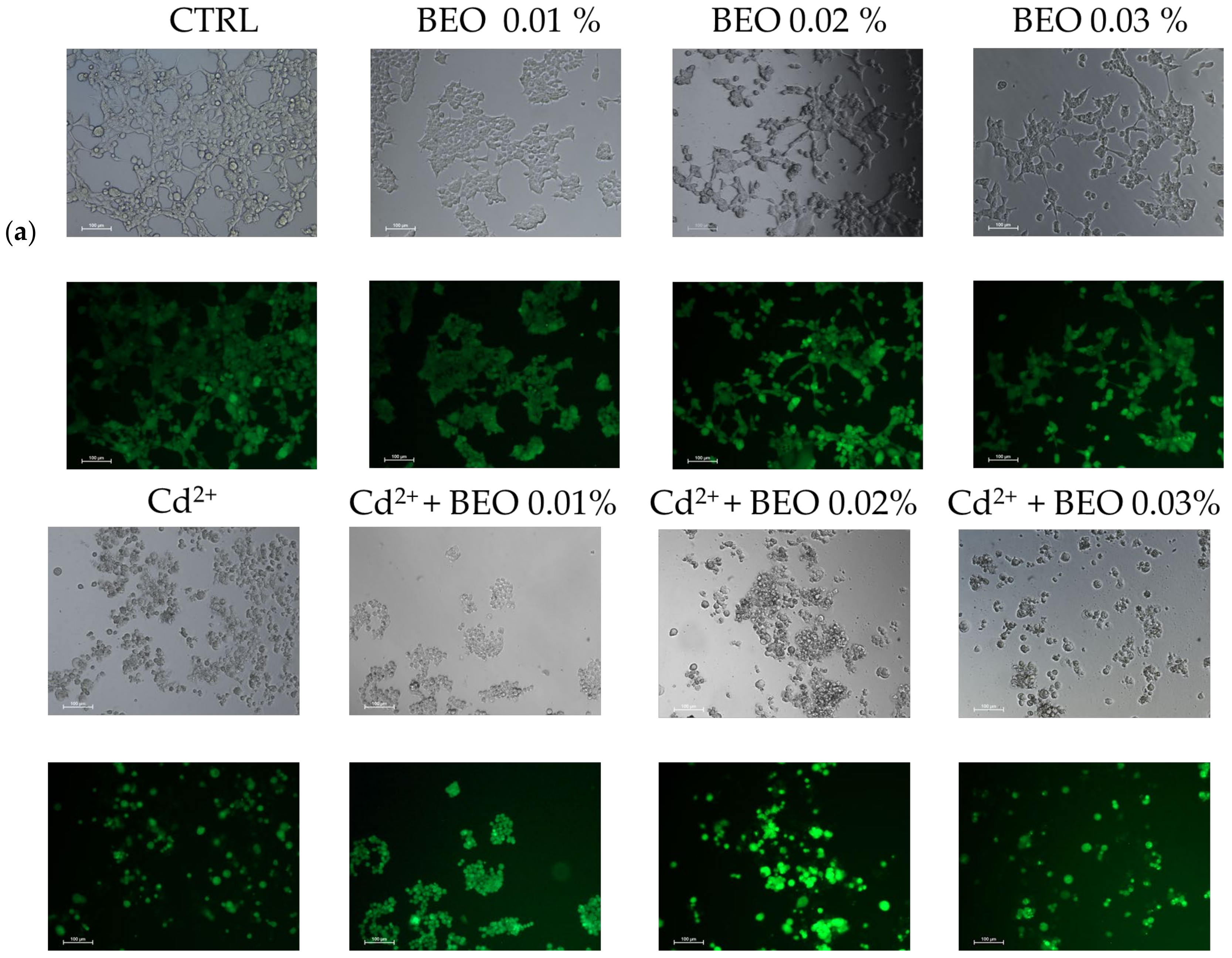
3.3. Effect of BEO on Cd2+-, Hg2+-, and Pb2+-Induced Cytotoxicity
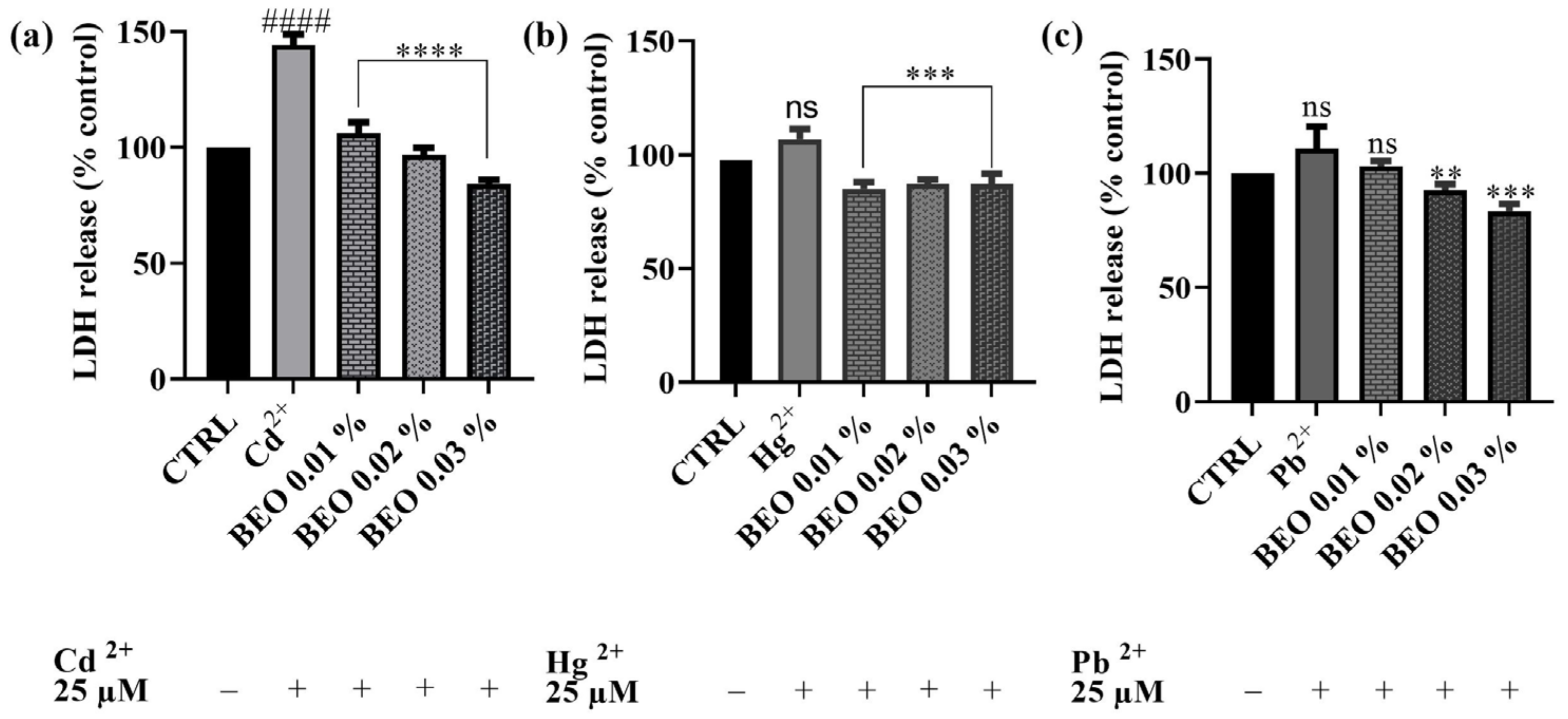
3.4. Effects of BEO on Cd2+, Hg2+, and Pb2+-Induced LDH Release
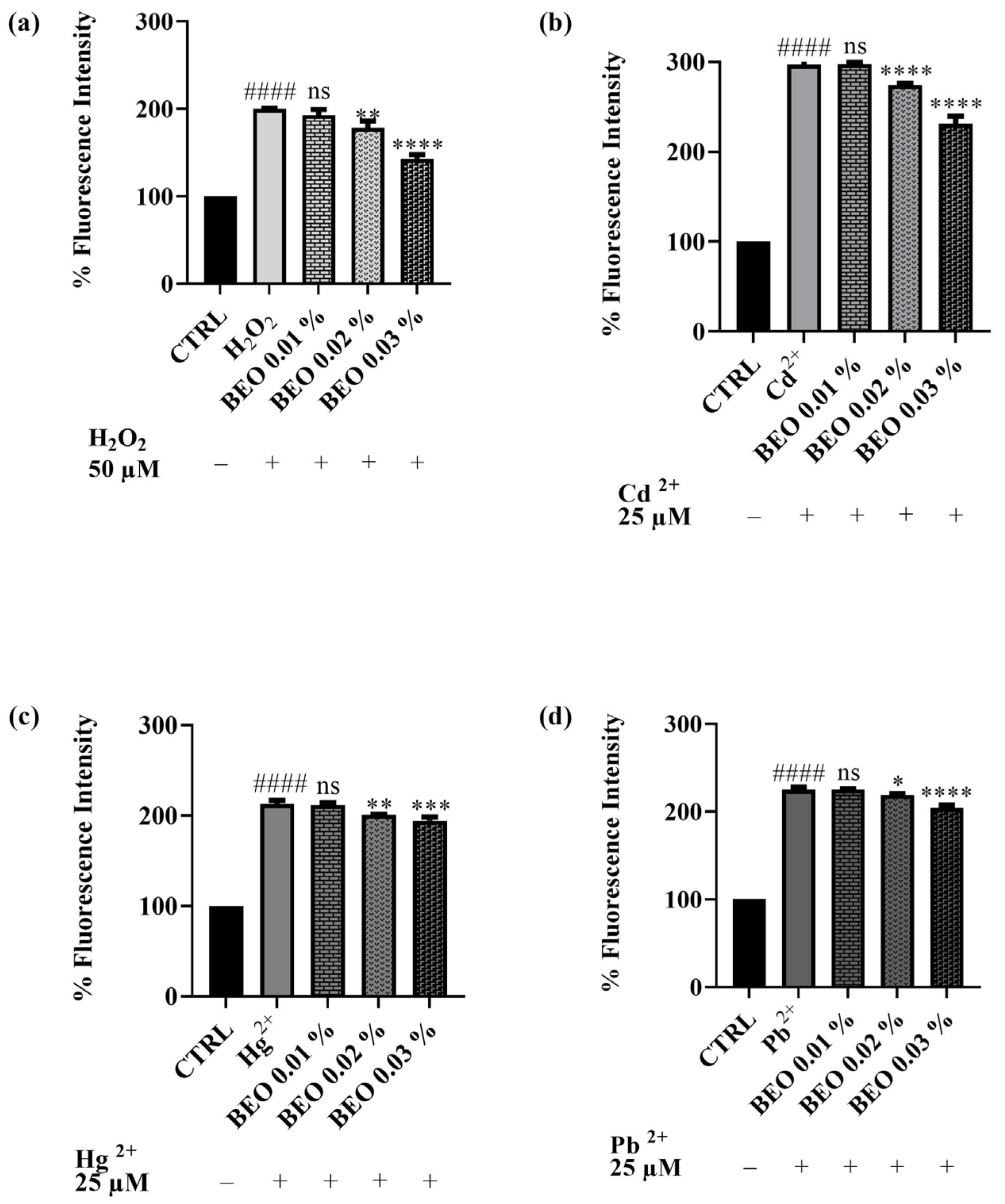
3.5. Scavenging Effects of BEO Against Cd2-, Hg2+-, Pb2+-, and H2O2 -Induced Oxidative Stress
3.6. Antibacterial Effects of BEO
4. Discussion
4.1. BEO Chemical Composition
4.2. Effects of Cd2+, Hg2+, Pb2+, and BEO on Cell Viability and the Cytoprotective Role of BEO Against Heavy Metals-Induced Cytotoxicity
4.3. Effects of BEO on Cd2+, Hg2+, and Pb2+-Induced LDH Release and Oxidative Stress
4.4. Antibacterial Effects of BEO
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dosoky, N.S.; Setzer, W.N. Biological Activities and Safety of Citrus spp. Essential Oils. Int. J. Mol. Sci. 2018, 19, 1966. [Google Scholar] [CrossRef]
- Gioffrè, G.; Ursino, D.; Labate, M.L.C.; Giuffrè, A.M. The peel essential oil composition of bergamot fruit (Citrus bergamia, Risso) of Reggio Calabria (Italy): A review. Emir. J. Food Agric. 2020, 32, 835–845. [Google Scholar]
- Testai, L.; Calderone, V. Nutraceutical Value of Citrus Flavanones and Their Implications in Cardiovascular Disease. Nutrients 2017, 9, 502. [Google Scholar] [CrossRef] [PubMed]
- Cirmi, S.; Bisignano, C.; Mandalari, G.; Navarra, M. Anti-infective potential of Citrus bergamia Risso et Poiteau (bergamot) derivatives: A systematic review. Phytother. Res. PTR 2016, 30, 1404–1411. [Google Scholar] [PubMed]
- Cirmi, S.; Ferlazzo, N.; Lombardo, G.E.; Ventura-Spagnolo, E.; Gangemi, S.; Calapai, G.; Navarra, M. Neurodegenerative Diseases: Might Citrus Flavonoids Play a Protective Role? Molecules 2016, 21, 1312. [Google Scholar] [CrossRef]
- Rombolà, L.; Amantea, D.; Russo, R.; Adornetto, A.; Berliocchi, L.; Tridico, L.; Corasaniti, M.; Sakurada, S.; Sakurada, T.; Bagetta, G.; et al. Rational basis for the use of bergamot essential oil in complementary medicine to treat chronic pain. Mini-Rev. Med. Chem. 2016, 16, 721–728. [Google Scholar] [PubMed]
- Corasaniti, M.T.; Maiuolo, J.; Maida, S.; Fratto, V.; Navarra, M.; Russo, R.; Amantea, D.; Morrone, L.A.; Bagetta, G. Cell signaling pathways in the mechanisms of neuroprotection afforded by bergamot essential oil against NMDA-induced cell death in vitro. Br. J. Pharmacol. 2007, 151, 518–529. [Google Scholar]
- Lin, X.; Cao, S.; Sun, J.; Lu, D.; Zhong, B.; Chun, J. The Chemical Compositions, and Antibacterial and Antioxidant Activities of Four Types of Citrus Essential Oils. Molecules 2021, 26, 3412. [Google Scholar] [CrossRef]
- Rehman, Q.; Rehman, K.; Akash, M.S.H. Heavy Metals and Neurological Disorders: From Exposure to Preventive Interventions. In Emerging Contaminants and Associated Treatment Technologies; Springer International Publishing: Cham, Switzerland, 2021; pp. 69–87. [Google Scholar]
- Bakulski, K.M.; Seo, Y.A.; Hickman, R.C.; Brandt, D.; Vadari, H.S.; Hu, H.; Park, S.K. Heavy Metals Exposure and Alzheimer’s Disease and Related Dementias. J. Alzheimers Dis. 2020, 76, 1215–1242. [Google Scholar]
- Kabir, T.; Uddin, S.; Zaman, S.; Begum, Y.; Ashraf, G.M.; Bin-Jumah, M.N.; Bungau, S.G.; Mousa, S.A.; Abdel-Daim, M.M. Molecular Mechanisms of Metal Toxicity in the Pathogenesis of Alzheimer’s Disease. Mol. Neurobiol. 2021, 58, 1–20. [Google Scholar]
- Meleleo, D.; Sblano, C.; Storelli, M.M.; Mallamaci, R. Evidence of cadmium and mercury involvement in the Aβ42 aggregation process. Biophys. Chem. 2020, 266, 106453. [Google Scholar]
- Garza-Lombo, C.; Posadas, Y.; Quintanar, L.; Gonsebatt, M.E.; Franco, R. Neurotoxicity linked to dysfunctional metal ion homeostasis and xenobiotic metal exposure: Redox Signaling and Oxidative Stress. Antioxid. Redox Signal. 2018, 28, 1669–1703. [Google Scholar] [CrossRef] [PubMed]
- Branca, J.J.V.; Fiorillo, C.; Carrino, D.; Paternostro, F.; Taddei, N.; Gulisano, M.; Pacini, A.; Becatti, M. Cadmium-induced oxidative stress: Focus on the central nervous system. Antioxidants 2020, 9, 492. [Google Scholar] [CrossRef] [PubMed]
- Mallamaci, R.; Barbarossa, A.; Carrieri, A.; Meleleo, D.; Carocci, A. Evaluation of the Potential Cytoprotective Effect of Melatonin in Comparison with Vitamin E and Trolox against Cd2+-Induced Toxicity in SH-SY5Y, HCT 116, and HepG2 Cell Lines. Int. J. Mol. Sci. 2024, 25, 8055. [Google Scholar] [CrossRef]
- Hunaiti, A.A.; Soud, M. Effect of lead concentration on the level of glutathione, glutathione S-transferase, reductase and peroxidase in human blood. Sci. Total Environ. 2000, 248, 45–50. [Google Scholar] [PubMed]
- Birla, H.; Minocha, T.; Kumar, G.; Misra, A.; Singh, S.K. Role of oxidative stress and metal toxicity in the progression of Alzheimer’s disease. Curr. Neuropharmacol. 2020, 18, 552–562. [Google Scholar]
- Zhang, H.; Wang, J.; Zhang, K.; Shi, J.; Gao, Y.; Zheng, J.; Li, H. Association between heavy metals exposure and persistent infections: The mediating role of immune function. Public Health Front. 2024, 12, 1367644. [Google Scholar]
- Li, F.; Hearn, M.; Bennett, L.E. The role of microbial infection in the pathogenesis of Alzheimer’s disease and the opportunity for protection by anti-microbial peptides. Crit. Rev. Microbiol. 2021, 47, 240–253. [Google Scholar]
- Qneibi, M.; Bdir, S.; Maayeh, C.; Bdair, M.; Sandouka, D.; Basit, D.; Hallak, M. A Comprehensive Review of Essential Oils and Their Pharmacological Activities in Neurological Disorders: Exploring Neuroprotective Potential. Neurochem. Res. 2024, 49, 258–289. [Google Scholar]
- Prashar, A.; Locke, I.C.; Evans, C.S. Cytotoxicity of lavender oil and its major components to human skin cells. Cell Prolif. 2004, 37, 221–229. [Google Scholar]
- Fadel, B.A.; Elwakil, B.H.; Fawzy, E.E.; Shaaban, M.M.; Olama, Z.A. Nanoemulsion of Lavandula angustifolia Essential Oil/Gold Nanoparticles: Antibacterial Effect against Multidrug-Resistant Wound-Causing Bacteria. Molecules 2023, 28, 6988. [Google Scholar] [CrossRef]
- Khan, R.A.; Mallick, N.; Feroz, Z. Anti-inflammatory effects of Citrus sinensis L., Citrus paradisi L. and their combinations. Pak. J. Pharm. Sci. 2016, 29, 843–852. [Google Scholar] [PubMed]
- Adorisio, S.; Muscari, I.; Fierabracci, A.; Thi Thuy, T.; Marchetti, M.C.; Ayroldi, E.; Delfino, D.V. Biological effects of bergamot and its potential therapeutic use as an anti-inflammatory, antioxidant, and anticancer agent. Pharm. Biol. 2023, 61, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Acar, Ü.; Kesbiç, O.S.; İnanan, B.E.; Yılmaz, S. Effects of dietary Bergamot (Citrus bergamia) peel oil on growth, haematology and immune response of European sea bass (Dicentrarchus labrax) juveniles. Aquac. Res. 2019, 50, 3305–3312. [Google Scholar]
- Yuan, C.; Shin, M.; Park, Y.; Choi, B.; Jang, S.; Lim, C.; Yun, H.S.; Lee, I.S.; Won, S.Y.; Cho, K.S. Linalool Alleviates Abeta42-Induced Neurodegeneration via Suppressing ROS Production and Inflammation in Fly and Rat Models of Alzheimer’s Disease. Oxidative Med. Cell. Longev. 2021, 2021, 8887716. [Google Scholar] [CrossRef]
- Shin, M.; Liu, Q.F.; Choi, B.; Shin, C.; Lee, B.; Yuan, C.; Song, Y.J.; Yun, H.S.; Lee, I.-S.; Koo, B.-S.; et al. Neuroprotective effects of limonene (+) against Aβ42-induced neurotoxicity in a Drosophila model of Alzheimer’s disease. Biol. Pharm. Bull. 2019, 43, 409–417. [Google Scholar]
- Quirino, A.; Giorgi, V.; Palma, E.A.-O.X.; Marascio, N.A.-O.; Morelli, P.A.-O.; Maletta, A.A.-O.X.; Divenuto, F.; De Angelis, G.; Tancrè, V.; Nucera, S.; et al. Citrus bergamia: Kinetics of Antimicrobial Activity on Clinical Isolates. Antibiotics 2022, 11, 361. [Google Scholar] [CrossRef]
- Rosato, A.; Sblano, S.; Salvagno, L.; Carocci, A.; Clodoveo, M.L.; Corbo, F.; Fracchiolla, G. Anti-Biofilm Inhibitory Synergistic Effects of Combinations of Essential Oils and Antibiotics. Antibiotics 2020, 9, 637. [Google Scholar] [CrossRef]
- Kaja, S.; Payne, A.J.; Naumchuk, Y.; Koulen, P. Quantification of lactate dehydrogenase for cell viability testing using cell lines and primary cultured astrocytes. Curr. Protoc. Toxicol. 2017, 72, 2–26. [Google Scholar]
- Barbarossa, A.; Ceramella, J.; Carocci, A.; Iacopetta, D.; Rosato, A.; Limongelli, F.; Carrieri, A.; Bonofiglio, D.; Sinicropi, M.S. Benzothiazole-Phthalimide Hybrids as Anti-Breast Cancer and Antimicrobial Agents. Antibiotics 2023, 12, 1651. [Google Scholar] [CrossRef]
- Barbarossa, A.; Sblano, S.; Rosato, A.; Carrieri, A.; Corbo, F.; Clodoveo, M.L.; Fracchiolla, G.; Carocci, A. Synergistic Action of Cinnamomum verum Essential Oil with Sertraline. Antibiotics 2022, 11, 1617. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.P.; Patel, J.B.; Burnhman, C.-A.; ZImmer, B.L. Clinical and Laboratory Standards Institute Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically Standard, Approval CDM-A. M07 Methods Dilution Antimicrob. Susceptibility Tests Bact. Grow Aerob. 2018; p. 91. Available online: https://kaldur.landspitali.is/focal/gaedahandbaekur/gnhsykla.nsf/5e27f2e5a88c898e00256500003c98c2/4c17737f339875620025756e003852bb/$FILE/M07Ed11E%20Methods%20for%20Dilution%20Antimicrobial%20Susceptibility%20Tests%20for%20Bacteria%20That%20Grow%20Aerobically.pdf (accessed on 25 February 2025).
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corp.: Carol Stream, IL, USA, 2007. [Google Scholar]
- NIST. Mass Spectral Library (NIST/EPA/NIH), 17; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2017. [Google Scholar]
- FFNSC. Flavors and Fragrances of Natural and Synthetic Compounds; Mass Spectral Database, 2; Shimadzu Corps: Kyoto, Japan, 2012. [Google Scholar]
- Yu, G.; Wu, L.; Su, Q.; Ji, X.; Zhou, J.; Wu, S.; Tang, Y.; Li, H. Neurotoxic effects of heavy metal pollutants in the environment: Focusing on epigenetic mechanisms. Environ. Pollut. 2024, 345, 123563. [Google Scholar]
- Xu, L.; He, D.; Bai, Y. Microglia-mediated inflammation and neurodegenerative disease. Mol. Neurobiol. 2016, 53, 6709–6715. [Google Scholar] [PubMed]
- Stojanović, N.M.; Ranđelović, P.J.; Simonović, M.; Radić, M.; Todorović, S.; Corrigan, M.; Harkin, A.; Boylan, F. Essential Oil Constituents as Anti-Inflammatory and Neuroprotective Agents: An Insight through Microglia Modulation. Int. J. Mol. Sci. 2024, 25, 5168. [Google Scholar] [CrossRef]
- Li, B.; Xia, M.; Zorec, R.; Parpura, V.; Verkhratsky, A. Astrocytes in Heavy Metal Neurotoxicity and Neurodegeneration. Brain Res. 2021, 1752, 147234. [Google Scholar] [CrossRef]
- Miyazawa, M.; Shindo, M.; Shimada, T. Metabolism of (+)- and (−)-Limonenes to Respective Carveols and Perillyl Alcohols by CYP2C9 and CYP2C19 in Human Liver Microsomes. Drug Metab. Dispos. 2002, 30, 602–607. [Google Scholar] [PubMed]
- Maugeri, A.; Lombardo, G.E.; Musumeci, L.; Russo, C.; Gangemi, S.; Calapai, G.; Cirmi, S.; Navarra, M. Bergamottin and 5-Geranyloxy-7-methoxycoumarin Cooperate in the Cytotoxic Effect of Citrus bergamia (Bergamot) Essential Oil in Human Neuroblastoma SH-SY5Y Cell Line. Toxins 2021, 13, 275. [Google Scholar] [CrossRef] [PubMed]
- Mondello, L.; Verzera, A.; Previti, P.; Crispo, F.; Dugo, G. Multidimensional capillary GC−GC for the analysis of complex samples. 5. Enantiomeric distribution of monoterpene hydrocarbons, monoterpene alcohols, and linalyl acetate of bergamot (Citrus bergamia Risso et Poiteau). Oils. J. Agric. Food Chem. 1998, 46, 4275–4282. [Google Scholar]
- Furneri, P.M.; Mondello, L.; Mandalari, G.; Paolino, D.; Dugo, P.; Garozzo, A.; Bisignano, G. In vitro antimycoplasmal activity of Citrus bergamia essential oil and its major components. Eur. J. Med. Chem. 2012, 52, 66–69. [Google Scholar]
- Rasheed, H.A.; Rehman, A.; Chen, X.; Aziz, T.; Al-Asmari, F.; Alhomrani, M.; Cui, H.; Lin, L. Unveiling the anti-listerial effect of Citrus bergamia essential oil: Mechanism of membrane disruption and anti-hemolytic activity. Food Biosci. 2024, 61, 104742. [Google Scholar]
- Bozova, B.; Gölükcü, M.; Giuffrè, A.M. The effect of different hydrodistillation times on the composition and yield of bergamot (Citrus bergamia Risso) peel essential oil and a comparison of the cold-pressing method. Flavour Fragr. J. 2024, 39, 263–270. [Google Scholar]
- Arruebarrena, M.A.; Hawe, C.T.; Lee, Y.M.; Branco, R.C. Mechanisms of Cadmium Neurotoxicity. Int. J. Mol. Sci. 2023, 24, 16558. [Google Scholar] [CrossRef]
- Xu, M.-Y.; Wang, P.; Sun, Y.-J.; Yang, L.; Wu, Y.-J. Joint toxicity of chlorpyrifos and cadmium on the oxidative stress and mitochondrial damage in neuronal cells. Food Chem. Toxicol. 2017, 103, 246–252. [Google Scholar]
- Mallamaci, R.; Barbarossa, A.; Carocci, A.; Meleleo, D. Evaluation of the Potential Protective Effect of Ellagic Acid against Heavy Metal (Cadmium, Mercury, and Lead) Toxicity in SH-SY5Y Neuroblastoma Cells. Foods 2024, 13, 419. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Lopez, M.E.; Augusto-Oliveira, M.; Lopes-Araújo, A.; Santos-Sacramento, L.; Souza-Monteiro, J.R.; da Rocha, F.F.; Arrifano, G.d.P. Chapter Eight-Mercury neurotoxicity in gold miners. In Advances in Neurotoxicology; Lucchini, R.G., Aschner, M., Costa, L.G., Eds.; Academic Press: Cambridge, MA, USA, 2022; Volume 7, pp. 283–314. [Google Scholar]
- Virgolini, M.B.; Aschner, M. Chapter 5—Molecular mechanisms of lead neurotoxicity. In Advances in Neurotoxicology; Aschner, M., Costa, L.G., Eds.; Academic Press: Cambridge, MA, USA, 2021; Volume 5, pp. 159–213. [Google Scholar]
- Berliocchi, L.; Ciociaro, A.; Russo, R.; Cassiano, M.G.V.; Blandini, F.; Rotiroti, D.; Morrone, L.A.; Corasaniti, M.T. Toxic profile of bergamot essential oil on survival and proliferation of SH-SY5Y neuroblastoma cells. Food Chem. Toxicol. 2011, 49, 2780–2792. [Google Scholar]
- Teixeira, J.; Basit, F.; Willems, P.H.G.M.; Wagenaars, J.A.; van de Westerlo, E.; Amorim, R.; Cagide, F.; Benfeito, S.; Oliveira, C.; Borges, F.; et al. Mitochondria-Targeted Phenolic Antioxidants Induce ROS-Protective Pathways in Primary Human Skin Fibroblasts. Free. Radic. Biol. Med. 2021, 163, 314–324. [Google Scholar]
- Kumar, K.J.S.; Vani, M.G.; Wang, S.Y. Limonene protects human skin keratinocytes against UVB-induced photodamage and photoaging by activating the Nrf2-dependent antioxidant defense system. Environ. Toxicol. 2022, 37, 2897–2909. [Google Scholar] [PubMed]
- Van der Werf, M.J.; Jongejan, H.; Franssen, M.C.R. Resolution of Limonene 1,2-Epoxide Diastereomers by Mercury(II) Ions. Tetrahedron Lett. 2001, 42, 5521–5524. [Google Scholar] [CrossRef]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar]
- Surh, Y.-J.; Kundu, J.K.; Na, H.-K. Nrf2 as a Master Redox Switch in Turning on the Cellular Signaling Involved in the Induction of Cytoprotective Genes by Some Chemopreventive Phytochemicals. Planta Medica 2008, 74, 1526–1539. [Google Scholar]
- Kim, H.J.; Herath, K.H.I.N.M.; Dinh, D.T.T.; Kim, H.S.; Jeon, Y.J.; Kim, H.J.; Jee, Y. Sargassum horneri ethanol extract containing polyphenols attenuates PM-induced oxidative stress via ROS scavenging and transition metal chelation. J. Funct. Foods 2021, 79, 104401. [Google Scholar]
- Berliocchi, L.; Russo, R.; Levato, A.; Fratto, V.; Bagetta, G.; Sakurada, S.; Sakurada, T.; Mercuri, N.B.; Corasaniti, M.T. (−)-Linalool attenuates allodynia in neuropathic pain induced by spinal nerve ligation in c57/bl6 mice. Int. Rev. Neurobiol. 2009, 85, 221–235. [Google Scholar] [PubMed]
- Verma, N.; Yadav, A.; Bal, S.; Gupta, R.; Aggarwal, N. In vitro Studies on Ameliorative Effects of Limonene on Cadmium-Induced Genotoxicity in Cultured Human Peripheral Blood Lymphocytes. Appl. Biochem. Biotechnol. 2019, 187, 1384–1397. [Google Scholar] [PubMed]
- Simons, T.J. Lead-calcium interactions in cellular lead toxicity. Neurotoxicology 1993, 14, 77–85. [Google Scholar]
- Brzóska, M.M.; Kozłowska, M.; Rogalska, J.; Gałażyn-Sidorczuk, M.; Roszczenko, A.; Smereczański, N.M. Enhanced Zinc Intake Protects against Oxidative Stress and Its Consequences in the Brain: A Study in an In vivo Rat Model of Cadmium Exposure. Nutrients 2021, 13, 478. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.C.; Peixe, T.S.; Mesas, A.E.; Paoliello, M.M. Lead Exposure and Oxidative Stress: A Systematic Review. Rev. Environ. Contam. Toxicol. 2016, 236, 193–238. [Google Scholar]
- Ajsuvakova, O.P.; Tinkova, A.; Aschner, M.; Rochae, J.B.T.; Michalkef, B.; Skalnayaa, M.G.; Skalny, A.V.; Butnariug, M.; Dadari, M.; Saracg, J.; et al. Sulfhydryl groups as targets of Hg toxicity. Coord. Chem. Rev. 2020, 15, 417. [Google Scholar]
- Meng, J.; Wang, W.-X.; Li, L.; Zhang, G. Tissue-Specific Molecular and Cellular Toxicity of Pb in the Oyster (Crassostrea gigas): MRNA Expression and Physiological Studies. Aquat. Toxicol. 2018, 198, 257–268. [Google Scholar]
- Mani, M.S.; Kunnathully, V.; Rao, C.; Kabekkodu, S.P.; Joshi, M.B.; D’Souza, H.S. Modifying effects of δ-Aminolevulinate dehydratase polymorphism on blood lead levels and ALAD activity. Toxicol. Lett. 2018, 295, 351–356. [Google Scholar]
- Shafaei, N.; Barkhordar, S.M.A.; Rahmani, F.; Nabi, S.; Idliki, R.B.; Alimirzaei, M.; Karimi, E.; Oskoueian, E. Protective Effects of Anethum graveolens Seed’s Oil Nanoemulsion Against Cadmium-Induced Oxidative Stress in Mice. Biol. Trace Elem. Res. 2020, 198, 583–591. [Google Scholar]
- Baschieri, A.; Ajvazi, M.D.; Tonfack, J.L.F.; Valgimigli, L.; Amorati, R. Explaining the antioxidant activity of some common non-phenolic components of essential oils. Food Chem. 2017, 232, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Okoye, C.N.; Stevens, D.; Kamunde, C. Modulation of mitochondrial site-specific hydrogen peroxide efflux by exogenous stressors. Free Radic. Biol. Med. 2021, 164, 439–456. [Google Scholar] [CrossRef]
- Fidalgo, F.; Freitas, R.; Ferreira, R.; Pessoa, A.M.; Teixeira, J. Solanum nigrum L. antioxidant defence system isozymes are regulated transcriptionally and posttranslationally in Cd-induced stress. Environ. Exp. Bot. 2011, 72, 312–319. [Google Scholar] [CrossRef]
- Bassolé, I.H.N.; Lamien-Meda, A.; Bayala, B.; Tirogo, S.; Franz, C.; Novak, J.; Nebié, R.C.; Dicko, M.H. Composition and Antimicrobial Activities of Lippia multiflora Moldenke, Mentha × piperita L. and Ocimum basilicum L. Essential Oils and Their Major Monoterpene Alcohols Alone and in Combination. Molecules 2010, 15, 7825–7839. [Google Scholar] [CrossRef]
- Iseppi, R.; Condò, C.; Messi, P. Synergistic Inhibition of Methicillin-Resistant Staphylococcus aureus (MRSA) by Melaleuca alternifolia Chell (Tea Tree) and Eucalyptus globulus Labill. Essential Oils in Association with Oxacillin. Antibiotics 2023, 12, 846. [Google Scholar]
- Gong, H.; Hu, X.; Liao, M.; Fa, K.; Ciumac, D.; Clifton, L.A.; Sani, M.A.; King, S.M.; Maestro, A.; Separovic, F.; et al. Structural Disruptions of the Outer Membranes of Gram-Negative Bacteria by Rationally Designed Amphiphilic Antimicrobial Peptides. ACS Appl. Mater. Interfaces 2021, 13, 16062–16074. [Google Scholar] [CrossRef] [PubMed]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar]
- Sharma, K.; Guleria, S.; Razdan, V.K.; Babu, V. Synergistic antioxidant and antimicrobial activities of essential oils of some selected medicinal plants in combination and with synthetic compounds. Ind. Crop Prod. 2020, 154, 112569. [Google Scholar] [CrossRef]
- Barbarossa, A.; Rosato, A.; Carrieri, A.; Tardugno, R.; Corbo, F.; Clodoveo, M.L.; Fracchiolla, G.; Carocci, A. Antifungal Biofilm Inhibitory Effects of Combinations of Diclofenac and Essential Oils. Antibiotics 2023, 12, 1673. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compound a | Class b | RI c | RI Lit. d | Area % ± SD e | ID f |
|---|---|---|---|---|---|---|
| 1 | α-thujene | MH | 924 | 924 | 0.22 ± 0.01 | Std,RI,MS |
| 2 | α-pinene | MH | 930 | 932 | 1.08 ± 0.00 | Std,RI,MS |
| 3 | Sabinene | MH | 970 | 969 | 0.77 ± 0.02 | Std,RI,MS |
| 4 | β-pinene | MH | 973 | 974 | 5.76 ± 0.04 | Std,RI,MS |
| 5 | Myrcene | MH | 989 | 988 | 0.69 ± 0.02 | Std,RI,MS |
| 6 | α-terpinene | MH | 1014 | 1014 | 0.05 ± 0.00 | Std,RI,MS |
| 7 | p-cymene | MH | 1022 | 1020 | 0.76 ± 0.00 | Std,RI,MS |
| 8 | Limonene | MH | 1026 | 1024 | 38.24 ± 0.32 | Std,RI,MS |
| 9 | γ-terpinene | MH | 1056 | 1054 | 6.58 ± 0.07 | Std,RI,MS |
| 10 | Terpinolene | MH | 1085 | 1086 | 0.15 ± 0.01 | Std,RI,MS |
| 11 | Linalool | MO | 1098 | 1095 | 14.45 ±0.09 | Std,RI,MS |
| 12 | α-terpineol | MO | 1187 | 1186 | 0.02 ± 0.00 | Std,RI,MS |
| 13 | linalool acetate | MO | 1256 | 1254 | 30.50 ± 0.16 | RI,MS |
| 14 | neryl acetate | MO | 1364 | 1359 | 0.17 ± 0.02 | RI,MS |
| 15 | geranyl acetate | MO | 1383 | 1379 | 0.16 ± 0.02 | RI,MS |
| 16 | α-(E)-bergamotene | SH | 1433 | 1432 | 0.07 ± 0.01 | RI,MS |
| 17 | (Z)-α-bisabolene | SH | 1507 | 1506 | 0.10 ± 0.02 | RI,MS |
| Total identified | 99.77 ± 0.03 | |||||
| Monoterpene hydrocarbons | 54.31 ± 0.37 | |||||
| Oxygenated monoterpenes | 45.29 ± 0.10 | |||||
| Sesquiterpene hydrocarbons | 0.17 ± 0.02 |
| MIC | ||||||||
|---|---|---|---|---|---|---|---|---|
| Microorganisms | ||||||||
| Gram-Positive | Gram-Negative | |||||||
| E. faecalis ATCC 29212 | S. aureus ATCC 29213 | S. aureus ATCC 25923 | S. aureus ATCC 43300 | E. coli ATCC 25922 | E. coli ATCC 35218 | K. pneumoniae ATCC 13883 | K. pneumoniae ATCC 70063 | |
| BEO | 2 | 0.5 | 0.5 | 1 | 1 | 2 | 4 | 4 |
| Levofloxacin | 2 | 0.5 | 0.5 | 0.5 | 0.06 | 0.12 | 8 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbarossa, A.; Mallamaci, R.; Spinozzi, E.; Maggi, F.; Sgobba, M.N.; Rosato, A.; Carocci, A.; Meleleo, D. Investigating Bergamot Essential Oil (BEO) Properties: Cytoprotection in Neuronal Cells Exposed to Heavy Metals and Antibacterial Activities. Antioxidants 2025, 14, 400. https://doi.org/10.3390/antiox14040400
Barbarossa A, Mallamaci R, Spinozzi E, Maggi F, Sgobba MN, Rosato A, Carocci A, Meleleo D. Investigating Bergamot Essential Oil (BEO) Properties: Cytoprotection in Neuronal Cells Exposed to Heavy Metals and Antibacterial Activities. Antioxidants. 2025; 14(4):400. https://doi.org/10.3390/antiox14040400
Chicago/Turabian StyleBarbarossa, Alexia, Rosanna Mallamaci, Eleonora Spinozzi, Filippo Maggi, Maria Noemi Sgobba, Antonio Rosato, Alessia Carocci, and Daniela Meleleo. 2025. "Investigating Bergamot Essential Oil (BEO) Properties: Cytoprotection in Neuronal Cells Exposed to Heavy Metals and Antibacterial Activities" Antioxidants 14, no. 4: 400. https://doi.org/10.3390/antiox14040400
APA StyleBarbarossa, A., Mallamaci, R., Spinozzi, E., Maggi, F., Sgobba, M. N., Rosato, A., Carocci, A., & Meleleo, D. (2025). Investigating Bergamot Essential Oil (BEO) Properties: Cytoprotection in Neuronal Cells Exposed to Heavy Metals and Antibacterial Activities. Antioxidants, 14(4), 400. https://doi.org/10.3390/antiox14040400