

Phytochemical Composition and Biological Properties of Macleania rupestris Fruit Extract: Insights into Its Antimicrobial and Antioxidant Activity

 ,
,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Physico-Chemical Analysis
2.2. Plant Extract
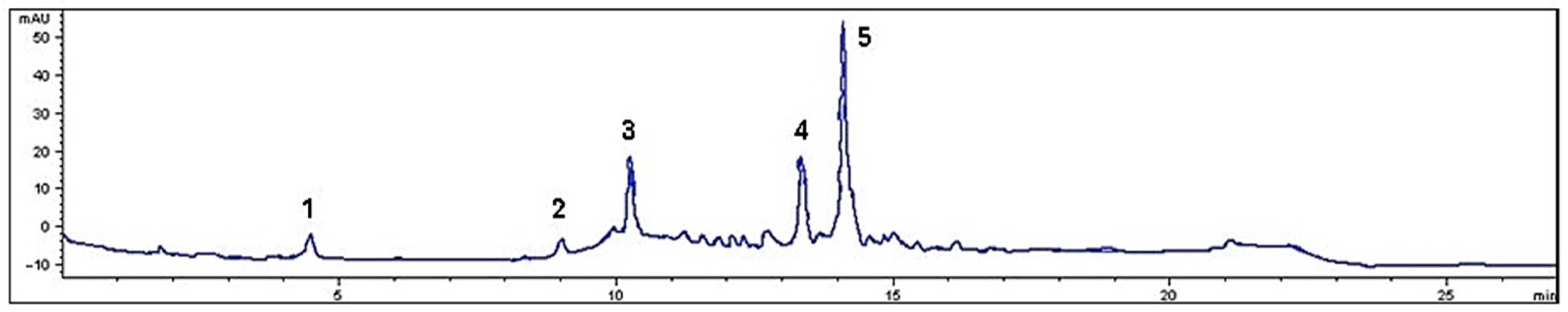
2.3. Phenolic Compound Identification
2.4. Organic Acid Identification
2.5. Total Anthocyanins
2.6. Antibacterial Activity Assay
2.7. Antifungal Activity Assay
2.8. Antibiofilm Activity
2.9. Antioxidant Activity
2.10. Statistical Analysis
3. Results and Discussion
3.1. Physico-Chemical Analysis
3.2. Minimum Inhibitory Concentration of Multidrug-Resistant Bacteria
3.3. Evaluation of Anti-Fungal Activity
3.4. Evaluation of Biofilm Inhibition Activity
3.5. Evaluation of Antioxidant Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luteyn, J. The plant family ericaceae (“blueberries”) in ecuador: Ecology, diversity, economic importance, and conservation. REMCB 2021, 42, 79–98. [Google Scholar] [CrossRef]
- Andrade, J.M.; Lucero Mosquera, H.; Armijos, C. Ethnobotany of indigenous saraguros: Medicinal plants used by community healers “hampiyachakkuna” in the san lucas parish, southern ecuador. BioMed Res. Int. 2017, 2017, 9343724. [Google Scholar] [CrossRef]
- Cardozo Gutiérrez, R.H.; Córdoba Cárdenas, S.L.; González Corredor, J.D.; Guzmán Castañeda, J.R.; Lancheros Redondo, H.O.; Mesa Castellanos, L.I.; Pacheco Salamanca, R.A.; Pérez Martínez, B.A.; Ramos Rodríguez, F.A.; Torres Cárcamo, M.E.; et al. Uva Camarona Macleania Rupestris (Kunth) A.C. Smith. In Especies Útiles en la Región Andina de Colombia Tomo I; Imprenta Nacional de Colombia: Bogotá, Colombia, 2009; pp. 81–93. ISBN 978-958-97749-2-2. [Google Scholar]
- Tene, V.; Malagón, O.; Finzi, P.V.; Vidari, G.; Armijos, C.; Zaragoza, T. An ethnobotanical survey of medicinal plants used in Loja and Zamora-Chinchipe, Ecuador. J. Ethnopharmacol. 2007, 111, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Dastmalchi, K.; Flores, G.; Wu, S.-B.; Pedraza-Peñalosa, P.; Long, C.; Kennelly, E.J. Antioxidant and metabolite profiling of North American and neotropical blueberries using LC-TOF-MS and multivariate analyses. J. Agric. Food Chem. 2013, 61, 3548–3559. [Google Scholar] [CrossRef]
- Jara Bermeo, A.; Quiroz Durazno, T.; Cazar Ramírez, M.E.; Valdes, M.E. Caracterización nutricional y actividad antioxidante de Macleania rupestris (Joyapa) y Vaccinium floribundum Kunth (Mortiño). Univ. Del Azuay 2013, 5, 38–48. [Google Scholar]
- Suquillo Bermeo, M.B.; Vargas López, J.H. Evaluation of the antioxidant potential of spray-dried microencapsulated extract and its formulation of Macleania rupestris (Kunth) A.C. Sm. Int. J. Appl. Eng. Technol. 2023, 5, 1–12. [Google Scholar]
- Ispiryan, A.; Atkociuniene, V.; Makstutiene, N.; Sarkinas, A.; Salaseviciene, A.; Urbonaviciene, D.; Viskelis, J.; Pakeltiene, R.; Raudone, L. Correlation between Antimicrobial Activity Values and Total Phenolic Content/Antioxidant Activity in Rubus idaeus L. Plants 2024, 13, 504. [Google Scholar] [CrossRef]
- Nadimpalli, M.L.; Chan, C.W.; Doron, S. Antibiotic resistance: A call to action to prevent the next epidemic of inequality. Nat. Med. 2021, 27, 187–188. [Google Scholar] [CrossRef]
- Lewis, K.; Lee, R.E.; Brötz-Oesterhelt, H.; Hiller, S.; Rodnina, M.V.; Schneider, T.; Weingarth, M.; Wohlgemuth, I. Sophisticated natural products as antibiotics. Nature 2024, 632, 39–49. [Google Scholar] [CrossRef]
- Liu, S.; Sun, L.; Zhang, P.; Niu, C. Recent Advances in Grayanane Diterpenes: Isolation, Structural Diversity, and Bioactivities from Ericaceae Family (2018–2024). Molecules 2024, 29, 1649. [Google Scholar] [CrossRef]
- Ștefănescu, B.E.; Szabo, K.; Mocan, A.; Crişan, G. Phenolic Compounds from Five Ericaceae Species Leaves and Their Related Bioavailability and Health Benefits. Molecules 2019, 24, 2046. [Google Scholar] [CrossRef] [PubMed]
- Armijos, C.; Ramírez, J.; Vidari, G. Poorly investigated ecuadorian medicinal plants. Plants 2022, 11, 1590. [Google Scholar] [CrossRef]
- Coyago-Cruz, E.; Valenzuela, D.; Guachamin, A.; Méndez, G.; Heredia-Moya, J.; Vera, E. Bioactive Compound Profiling and Antioxidant Activity of Phytelephas tenuicaulis and Other Amazonian Fruits. Foods 2024, 13, 2151. [Google Scholar] [CrossRef] [PubMed]
- Coyago-Cruz, E.; Guachamin, A.; Villacís, M.; Rivera, J.; Neto, M.; Méndez, G.; Heredia-Moya, J.; Vera, E. Evaluation of bioactive compounds and antioxidant activity in 51 minor tropical fruits of ecuador. Foods 2023, 12, 4439. [Google Scholar] [CrossRef]
- Barba-Ostria, C.; Carrera-Pacheco, S.E.; Gonzalez-Pastor, R.; Zuñiga, J.; Mayorga-Ramos, A.; Tejera, E.; Guamán, L. Exploring the Multifaceted Biological Activities of Anthocyanins Isolated from Two Andean Berries. Foods 2024, 13, 2625. [Google Scholar] [CrossRef]
- Coyago-Cruz, E.; Guachamin, A.; Méndez, G.; Moya, M.; Martínez, A.; Viera, W.; Heredia-Moya, J.; Beltrán, E.; Vera, E.; Villacís, M. Functional and Antioxidant Evaluation of Two Ecotypes of Control and Grafted Tree Tomato (Solanum betaceum) at Different Altitudes. Foods 2023, 12, 3494. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Teran, R.; Guevara, R.; Mora, J.; Dobronski, L.; Barreiro-Costa, O.; Beske, T.; Pérez-Barrera, J.; Araya-Maturana, R.; Rojas-Silva, P.; Poveda, A.; et al. Characterization of Antimicrobial, Antioxidant, and Leishmanicidal Activities of Schiff Base Derivatives of 4-Aminoantipyrine. Molecules 2019, 24, 2696. [Google Scholar] [CrossRef]
- CLSI Clinical and Laboratory Standards Institute (CLSI). Method for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 12th ed.; CLSI Standard M07: Wayne, PA, USA, 2024; pp. 21–68. ISBN 978-1-68440-226-7. [Google Scholar]
- Arikan, S. Current status of antifungal susceptibility testing methods. Med. Mycol. 2007, 45, 569–587. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; CLSI supplement M38M51S, Ed.; Clinical and Laboratory Standards Institute: Wayne, PE, USA, 2022; ISBN 978-1-68440-161-1. [Google Scholar]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005, 1, 1B.1.1–1B.1.18. [Google Scholar] [CrossRef]
- Coyago-Cruz, E.; Salazar, I.; Guachamin, A.; Alomoto, M.; Cerna, M.; Mendez, G.; Heredia-Moya, J.; Vera, E. Bioactive Compounds, Antioxidant, and Antimicrobial Activity of Seeds and Mucilage of Non-Traditional Cocoas. Antioxidants 2025, 14, 299. [Google Scholar] [CrossRef]
- Curillo, D.; Cevallos-Cevallos, J.M.; Chica, E.; Peña, D. Characterization of the microbial community associated with the roots of joyapa (Macleania rupestris) and the effect of fungal isolates on seedling development. Revis. Bionatura 2023, 8, 1–7. [Google Scholar] [CrossRef]
- Abbasi, K.S.; Qayyum, A.; Jahangir, M.; Sohail, A.; Nisa, S. Comparative analysis of citrus fruits for nutraceutical properties. Food Sci. Technol. 2020, 40, 153–157. [Google Scholar]
- Alam, S.; Bibi, S.; Ali, M. Fungal diversity and physicochemical changes in fresh fruits during storage at ambient room conditions. Appl. Microbiol. Theory Technol. 2020, 1, 18–31. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Fiorentini, D.; Cappadone, C.; Farruggia, G.; Prata, C. Magnesium: Biochemistry, nutrition, detection, and social impact of diseases linked to its deficiency. Nutrients 2021, 13, 1136. [Google Scholar] [CrossRef]
- European Food Safety Autority. Overview on dietary reference values for the EU population as derived by the EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). In Summary of Dietary Reference Values—Version 4 (September 2017); EFSA: Parma, Italy, 2017. [Google Scholar]
- Bae, J.; Kim, N.; Shin, Y.; Kim, S.-Y.; Kim, Y.-J. Activity of catechins and their applications. Biomed. Dermatol. 2020, 4, 8. [Google Scholar] [CrossRef]
- Coyago-Cruz, E.; Corell, M.; Moriana, A.; Hernanz, D.; Benítez-González, A.M.; Stinco, C.M.; Meléndez-Martínez, A.J. Antioxidants (carotenoids and phenolics) profile of cherry tomatoes as influenced by deficit irrigation, ripening and cluster. Food Chem. 2018, 240, 870–884. [Google Scholar] [CrossRef] [PubMed]
- Dobrinas, S.; Soceanu, A. Determination of total phenolic content from plant extracts used in cosmetic purpose. J. Sci. Arts 2021, 21, 247–260. [Google Scholar] [CrossRef]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; El-Ansary, D.O.; Al-Mana, F.; Mahmoud, E.A. Polyphenol Content and Biological Activities of Ruta graveolens L. and Artemisia abrotanum L. in Northern Saudi Arabia. Processes 2020, 8, 531. [Google Scholar] [CrossRef]
- Hirai, I.; Okuno, M.; Katsuma, R.; Arita, N.; Tachibana, M.; Yamamoto, Y. Characterisation of anti-Staphylococcus aureus activity of quercetin. Int. J. Food Sci. Technol. 2010, 45, 1250–1254. [Google Scholar] [CrossRef]
- Phalanisong, P.; Vichitphan, K.; Han, J.; Vichitphan, S. High Antioxidant and Phenolic Contents Related to Antibacterial Activity against Gastrointestinal Pathogenic Bacteria of Some Thai Medicinal Plants. PJ 2018, 10, 341–348. [Google Scholar] [CrossRef]
- da Silva, C.M.A.; Costa, B.M.S.; Silva, A.G.; Souza, E.B.; Silva, M.V.; Correia, M.T.; SantÂ’Anna, S.A.P.; de Menezes, L.V.L. Antimicrobial activity of several Brazilian medicinal plants against phytopathogenic bacteria. Afr. J. Microbiol. Res. 2016, 10, 578–583. [Google Scholar]
- Raybaudi-Massilia, R.M.; Mosqueda-Melgar, J.; Martín-Belloso, O. Antimicrobial activity of malic acid against Listeria monocytogenes, Salmonella Enteritidis and Escherichia coli O157:H7 in apple, pear and melon juices. Food Control 2009, 20, 105–112. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular Basis of Bacterial Outer Membrane Permeability Revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Yamasaki, S.; Wang, L.-Y.; Hirata, T.; Hayashi-Nishino, M.; Nishino, K. Multidrug efflux pumps contribute to Escherichia coli biofilm maintenance. Int. J. Antimicrob. Agents 2015, 45, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Jothika, M.; Padmavathy, K.; Prabu, D. Phytochemicals as a Potential Alternative for Combating Multidrug-Resistant Bacterial Infections. In Plant Derived Bioactive Compounds in Human Health and Disease; CRC Press: Boca Raton, FL, USA, 2024; pp. 33–48. ISBN 9781003486237. [Google Scholar]
- Cheng, Q.; Dickwella Widanage, M.C.; Yarava, J.R.; Ankur, A.; Latgé, J.-P.; Wang, P.; Wang, T. Molecular architecture of chitin and chitosan-dominated cell walls in zygomycetous fungal pathogens by solid-state NMR. Nat. Commun. 2024, 15, 8295. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A. Chitin and glucan, the yin and yang of the fungal cell wall, implications for antifungal drug discovery and therapy. Adv. Appl. Microbiol. 2013, 83, 145–172. [Google Scholar] [CrossRef]
- Yang, M.; Abdullah; Ahmad, N.; Hussain, M.; Lu, X.; Xu, J.; Zhong, H.; Guan, R. A review of recent advances on cyanidin-3-glucoside: The biotransformation, absorption, bioactivity and applications of nano-encapsulation. Food Funct. 2023, 14, 6320–6345. [Google Scholar] [CrossRef]
- Patra, S.; Makhal, P.N.; Jaryal, S.; More, N.; Kaki, V.R. Anthocyanins: Plant-based flavonoid pigments with diverse biological activities. Int. J. Plant Based Pharm. 2022, 2, 118–127. [Google Scholar] [CrossRef]
- Oliveira, H.; Correia, P.; Bessa, L.J.; Guimarães, M.; Gameiro, P.; Freitas, V.d.; Mateus, N.; Cruz, L.; Fernandes, I. Cyanidin-3-glucoside Lipophilic Conjugates for Topical Application: Tuning the Antimicrobial Activities with Fatty Acid Chain Length. Processes 2021, 9, 340. [Google Scholar] [CrossRef]
- Jasińska, K.; Fabiszewska, A.; Białecka-Florjańczyk, E.; Zieniuk, B. Mini-Review on the Enzymatic Lipophilization of Phenolics Present in Plant Extracts with the Special Emphasis on Anthocyanins. Antioxidants 2022, 11, 1528. [Google Scholar] [CrossRef] [PubMed]
- El-Houri, R.B.; Kotowska, D.; Olsen, L.C.B.; Bhattacharya, S.; Christensen, L.P.; Grevsen, K.; Oksbjerg, N.; Færgeman, N.; Kristiansen, K.; Christensen, K.B. Screening for bioactive metabolites in plant extracts modulating glucose uptake and fat accumulation. Evid. Based Complement. Alternat. Med. 2014, 2014, 156398. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.F.G.; Sales, J.A.; da Rocha, M.G.; Galdino, L.M.; de Aguiar, L.; Pereira-Neto, W.d.A.; de Aguiar Cordeiro, R.; Castelo-Branco, D.d.S.C.M.; Sidrim, J.J.C.; Brilhante, R.S.N. Antifungal effects of the flavonoids kaempferol and quercetin: A possible alternative for the control of fungal biofilms. Biofouling 2019, 35, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Childers, D.S.; Usher, J. Is metabolic generalism the Breakfast of Champions for pathogenic Candida species? PLoS Pathog. 2024, 20, e1012752. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Ghasemian, A. An overview on anti-biofilm properties of quercetin against bacterial pathogens. World J. Microbiol. Biotechnol. 2019, 35, 143. [Google Scholar] [CrossRef]
- Vazquez-Armenta, F.J.; Bernal-Mercado, A.T.; Tapia-Rodriguez, M.R.; Gonzalez-Aguilar, G.A.; Lopez-Zavala, A.A.; Martinez-Tellez, M.A.; Hernandez-Oñate, M.A.; Ayala-Zavala, J.F. Quercetin reduces adhesion and inhibits biofilm development by Listeria monocytogenes by reducing the amount of extracellular proteins. Food Control 2018, 90, 266–273. [Google Scholar] [CrossRef]
- Rodríguez-López, P.; Rodríguez-Herrera, J.J.; Vázquez-Sánchez, D.; López Cabo, M. Current Knowledge on Listeria monocytogenes Biofilms in Food-Related Environments: Incidence, Resistance to Biocides, Ecology and Biocontrol. Foods 2018, 7, 85. [Google Scholar] [CrossRef]
- Brauge, T.; Sadovskaya, I.; Faille, C.; Benezech, T.; Maes, E.; Guerardel, Y.; Midelet-Bourdin, G. Teichoic acid is the major polysaccharide present in the Listeria monocytogenes biofilm matrix. FEMS Microbiol. Lett. 2016, 363, fnv229. [Google Scholar] [CrossRef]
- Lee, J.-H.; Park, J.-H.; Cho, H.S.; Joo, S.W.; Cho, M.H.; Lee, J. Anti-biofilm activities of quercetin and tannic acid against Staphylococcus aureus. Biofouling 2013, 29, 491–499. [Google Scholar] [CrossRef]
- Ren, Y.; Zhu, R.; You, X.; Li, D.; Guo, M.; Fei, B.; Liu, Y.; Yang, X.; Liu, X.; Li, Y. Quercetin: A promising virulence inhibitor of Pseudomonas aeruginosa LasB in vitro. Appl. Microbiol. Biotechnol. 2024, 108, 57. [Google Scholar] [CrossRef]
- Ouyang, J.; Sun, F.; Feng, W.; Sun, Y.; Qiu, X.; Xiong, L.; Liu, Y.; Chen, Y. Quercetin is an effective inhibitor of quorum sensing, biofilm formation and virulence factors in Pseudomonas aeruginosa. J. Appl. Microbiol. 2016, 120, 966–974. [Google Scholar] [CrossRef]
- Kaur, J.; Kaur, R. p-Coumaric Acid: A Naturally Occurring Chemical with Potential Therapeutic Applications. Curr. Org. Chem. 2022, 26, 1333–1349. [Google Scholar] [CrossRef]
- Lou, Z.; Tang, Y.; Song, X.; Wang, H. Metabolomics-Based Screening of Biofilm-Inhibitory Compounds against Pseudomonas aeruginosa from Burdock Leaf. Molecules 2015, 20, 16266–16277. [Google Scholar] [CrossRef] [PubMed]
- Ojha, D.; Patil, K.N. p-Coumaric acid inhibits the Listeria monocytogenes RecA protein functions and SOS response: An antimicrobial target. Biochem. Biophys. Res. Commun. 2019, 517, 655–661. [Google Scholar] [CrossRef]
- Zawiła, T.; Swolana, D.; Zawiła, M.; Wojtyczka, R.D. Synergistic Interactions between Selected β-Lactam Antibiotics and Cinnamic Acid and Its Chosen Derivatives. Antibiotics 2024, 13, 710. [Google Scholar] [CrossRef] [PubMed]
- Myszka, K.; Schmidt, M.T.; Białas, W.; Olkowicz, M.; Leja, K.; Czaczyk, K. Role of gallic and p-coumaric acids in the AHL-dependent expression of flgA gene and in the process of biofilm formation in food-associated Pseudomonas fluorescens KM120. J. Sci. Food Agric. 2016, 96, 4037–4047. [Google Scholar] [CrossRef]
- Coban, H.B. Organic acids as antimicrobial food agents: Applications and microbial productions. Bioprocess Biosyst. Eng. 2020, 43, 569–591. [Google Scholar] [CrossRef]
- Sundberg, C.; Jönsson, H. Process inhibition due to organic acids in fed-batch composting of food waste–influence of starting culture. Biodegradation 2005, 16, 205–213. [Google Scholar] [CrossRef]
- Canibe, N.; Steien, S.H.; Overland, M.; Jensen, B.B. Effect of K-diformate in starter diets on acidity, microbiota, and the amount of organic acids in the digestive tract of piglets, and on gastric alterations. J. Anim. Sci. 2001, 79, 2123. [Google Scholar] [CrossRef]
- Singla, R.; Goel, H.; Ganguli, A. Novel synergistic approach to exploit the bactericidal efficacy of commercial disinfectants on the biofilms of Salmonella enterica serovar Typhimurium. J. Biosci. Bioeng. 2014, 118, 34–40. [Google Scholar] [CrossRef]
- Over, K.F.; Hettiarachchy, N.; Johnson, M.G.; Davis, B. Effect of organic acids and plant extracts on Escherichia coli O157:H7, Listeria monocytogenes, and Salmonella Typhimurium in broth culture model and chicken meat systems. J. Food Sci. 2009, 74, M515–M521. [Google Scholar] [CrossRef]
- Mikaelyan, A.R.; Babayan, B.G.; Grigoryan, A.L.; Grigoryan, A.M.; Asatryan, N.L.; Melkumyan, M.A. Tartaric acid new derivatives as prospective and safe alternative to antimicrobials for food products packing. FFHD 2024, 14, 33. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Z.; Chen, D.; Yu, Z.; Huang, L.; Yu, H.; Yao, W.; Xie, Y. Inactivation effect of Staphylococcus aureus and application on fresh-cut pineapples by plasma-activated tartaric acid. Food Biosci. 2023, 54, 102789. [Google Scholar] [CrossRef]
- Itam, A.; Wati, M.S.; Agustin, V.; Sabri, N.; Jumanah, R.A.; Efdi, M. Comparative Study of Phytochemical, Antioxidant, and Cytotoxic Activities and Phenolic Content of Syzygium aqueum (Burm. f. Alston f.) Extracts Growing in West Sumatera Indonesia. Sci. World J. 2021, 2021, 5537597. [Google Scholar] [CrossRef] [PubMed]
- Mustarichie, R. The antioxidant activity and phytochemical screening of ethanol extract, fractions of water, ethyl acetate and n-hexane from mistletoe tea (Scurrula atropurpurea bl. dans). Asian J. Pharm. Clin. Res. 2017, 10, 343. [Google Scholar] [CrossRef]
- Sricharoen, P.; Techawongstein, S.; Chanthai, S. A high correlation indicating for an evaluation of antioxidant activity and total phenolics content of various chilli varieties. J. Food Sci. Technol. 2015, 52, 8077–8085. [Google Scholar] [CrossRef]
- Pavlović, R.D.; Lakusić, B.; Doslov-Kokorus, Z.; Kovacević, N. Arbutin content and antioxidant activity of some Ericaceae species. Pharmazie 2009, 64, 656–659. [Google Scholar]
- Kim, Y.-H.; Bang, C.-Y.; Won, E.-K.; Kim, J.-P.; Choung, S.-Y. Antioxidant activities of Vaccinium uliginosum L. extract and its active components. J. Med. Food 2009, 12, 885–892. [Google Scholar] [CrossRef]
- Dastmalchi, K.; Flores, G.; Petrova, V.; Pedraza-Peñalosa, P.; Kennelly, E.J. Edible neotropical blueberries: Antioxidant and compositional fingerprint analysis. J. Agric. Food Chem. 2011, 59, 3020–3026. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Morais, J.B.S.; Severo, J.S.; Santos, L.R.D.; de Sousa Melo, S.R.; de Oliveira Santos, R.; de Oliveira, A.R.S.; Cruz, K.J.C.; do Nascimento Marreiro, D. Role of Magnesium in Oxidative Stress in Individuals with Obesity. Biol. Trace Elem. Res. 2017, 176, 20–26. [Google Scholar] [CrossRef]
- Ghosh, N.; Chakraborty, T.; Mallick, S.; Mana, S.; Singha, D.; Ghosh, B.; Roy, S. Synthesis, characterization and study of antioxidant activity of quercetin-magnesium complex. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 151, 807–813. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, R.; Della Porta, M.; Piuri, G.; Maier, J.A. Magnesium: A defense line to mitigate inflammation and oxidative stress in adipose tissue. Antioxidants 2024, 13, 893. [Google Scholar] [CrossRef] [PubMed]
- Rumpf, J.; Burger, R.; Schulze, M. Statistical evaluation of DPPH, ABTS, FRAP, and Folin-Ciocalteu assays to assess the antioxidant capacity of lignins. Int. J. Biol. Macromol. 2023, 233, 123470. [Google Scholar] [CrossRef]
- Schaich, K.M.; Tian, X.; Xie, J. Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays. J. Funct. Foods 2015, 14, 111–125. [Google Scholar] [CrossRef]
- Zhang, D.; Chu, L.; Liu, Y.; Wang, A.; Ji, B.; Wu, W.; Zhou, F.; Wei, Y.; Cheng, Q.; Cai, S.; et al. Analysis of the antioxidant capacities of flavonoids under different spectrophotometric assays using cyclic voltammetry and density functional theory. J. Agric. Food Chem. 2011, 59, 10277–10285. [Google Scholar] [CrossRef]
- Chaves, N.; Santiago, A.; Alías, J.C. Quantification of the antioxidant activity of plant extracts: Analysis of sensitivity and hierarchization based on the method used. Antioxidants 2020, 9, 76. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Parameters | Value |
|---|---|
| Weight (g) | 2.1 ± 0.4 |
| Equatorial diameter (mm) | 14.3 ± 1.4 |
| Longitudinal diameter (mm) | 14.5 ± 1.2 |
| pH | 3.7 ± 0.6 |
| Soluble solids (°Brix) | 10.3 ± 0.6 |
| Total titratable acidity (%) | 0.8 ± 0.1 |
| Moisture (%) | 82.6 ± 1.3 |
| Ethereal extract (g/100 g) * | 0.7 ± 0.0 |
| Calorific value (kcal) * | 69.0 ± 0.1 |
| Ash (%) (200 mg/100 g) | 0.2 ± 0.0 |
| Fe (mg/100 g DW 1) * | 8.4 ± 0.6 |
| Na (mg/100 g DW 1) * | 74.5 ± 9.4 |
| K (mg/100 g DW 1) * | 241.6 ± 54.7 |
| Mg (mg/100 g DW 1) * | 576.5 ± 6.1 |
| Fibre (g/100 g) * | 4.3 ± 0.0 |
| Total carbohydrate (g/100 g) * | 15.1 ± 0.2 |
| Protein (g/100 g) | 1.0 ± 0.1 |
| Parameters | Value | ||
|---|---|---|---|
| Phenolics (mg/100 g DW 1) | |||
| Gallic acid | 14.9 | ± | 0.0 |
| Catechin | 18.5 | ± | 5.9 |
| p-Coumaric acid | 511.1 | ± | 11.0 |
| Syringic acid | 151.9 | ± | 6.0 |
| Quercetin | 157.5 | ± | 7.2 |
| Total phenolics | 853.9 | ± | 18.3 |
| Organic acids (mg/100 g DW 1) | |||
| Malic acid | 189.4 | ± | 18.5 |
| Tartaric acid | 306.0 | ± | 13.4 |
| Citric acid | 77.8 | ± | 4.6 |
| Total organic acid | 573.2 | ± | 36.6 |
| Total anthocyanins (mg C-3-gl/100 g DW 1) | 21.5 | ± | 2.5 |
| Bacteria Strain | MIC (mg/mL) |
|---|---|
| K. pneumoniae | NA 1 |
| E. coli | 5.00 |
| E. faecalis | 1.25 |
| S. epidermidis | 5.00 |
| E. faecium | 1.25 |
| S. enterica serovariedad Kentucky | NA 1 |
| P. aeruginosa | NA 1 |
| Compound | DPPH (mmol TE/100 g DW) | ABTS mmol TE/100 g (DW) |
|---|---|---|
| Macleania rupestris | 1.62 ± 0.09 | 3.28 ± 0.06 |
| IC50 Trolox (mM) | 0.23 ± 0.04 | 0.43 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayorga-Ramos, A.; Zúñiga-Miranda, J.; Coyago-Cruz, E.; Heredia-Moya, J.; Guamán-Bautista, J.; Guamán, L.P. Phytochemical Composition and Biological Properties of Macleania rupestris Fruit Extract: Insights into Its Antimicrobial and Antioxidant Activity. Antioxidants 2025, 14, 394. https://doi.org/10.3390/antiox14040394
Mayorga-Ramos A, Zúñiga-Miranda J, Coyago-Cruz E, Heredia-Moya J, Guamán-Bautista J, Guamán LP. Phytochemical Composition and Biological Properties of Macleania rupestris Fruit Extract: Insights into Its Antimicrobial and Antioxidant Activity. Antioxidants. 2025; 14(4):394. https://doi.org/10.3390/antiox14040394
Chicago/Turabian StyleMayorga-Ramos, Arianna, Johana Zúñiga-Miranda, Elena Coyago-Cruz, Jorge Heredia-Moya, Jéssica Guamán-Bautista, and Linda P. Guamán. 2025. "Phytochemical Composition and Biological Properties of Macleania rupestris Fruit Extract: Insights into Its Antimicrobial and Antioxidant Activity" Antioxidants 14, no. 4: 394. https://doi.org/10.3390/antiox14040394
APA StyleMayorga-Ramos, A., Zúñiga-Miranda, J., Coyago-Cruz, E., Heredia-Moya, J., Guamán-Bautista, J., & Guamán, L. P. (2025). Phytochemical Composition and Biological Properties of Macleania rupestris Fruit Extract: Insights into Its Antimicrobial and Antioxidant Activity. Antioxidants, 14(4), 394. https://doi.org/10.3390/antiox14040394