

Combining Abiotic Stresses as a Low-Cost Strategy for Increasing the Phenolic Content in Apple Agro-Industrial By-Products




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Assay 1: Experimental Design for Biofortification Process Using Wounding-Induced Stress
2.3. Assay 2: Experimental Design for Combined Biofortification Process Using Wounding and UV-A Radiation Stress
2.4. Enzymatic Activity Determinations
2.4.1. Phenylalanine Ammonia-Lyase Enzyme (PAL)
2.4.2. Polyphenol Oxidase Enzyme (PPO)
2.4.3. Protein Content Determination
2.5. Phenolic Compounds Determination
2.5.1. Total Phenolic Content
2.5.2. Phenolic Compound Profile
2.6. Statical Analysis
3. Results
3.1. Assay 1 Biofortification Process Using Wounding-Induced Stress
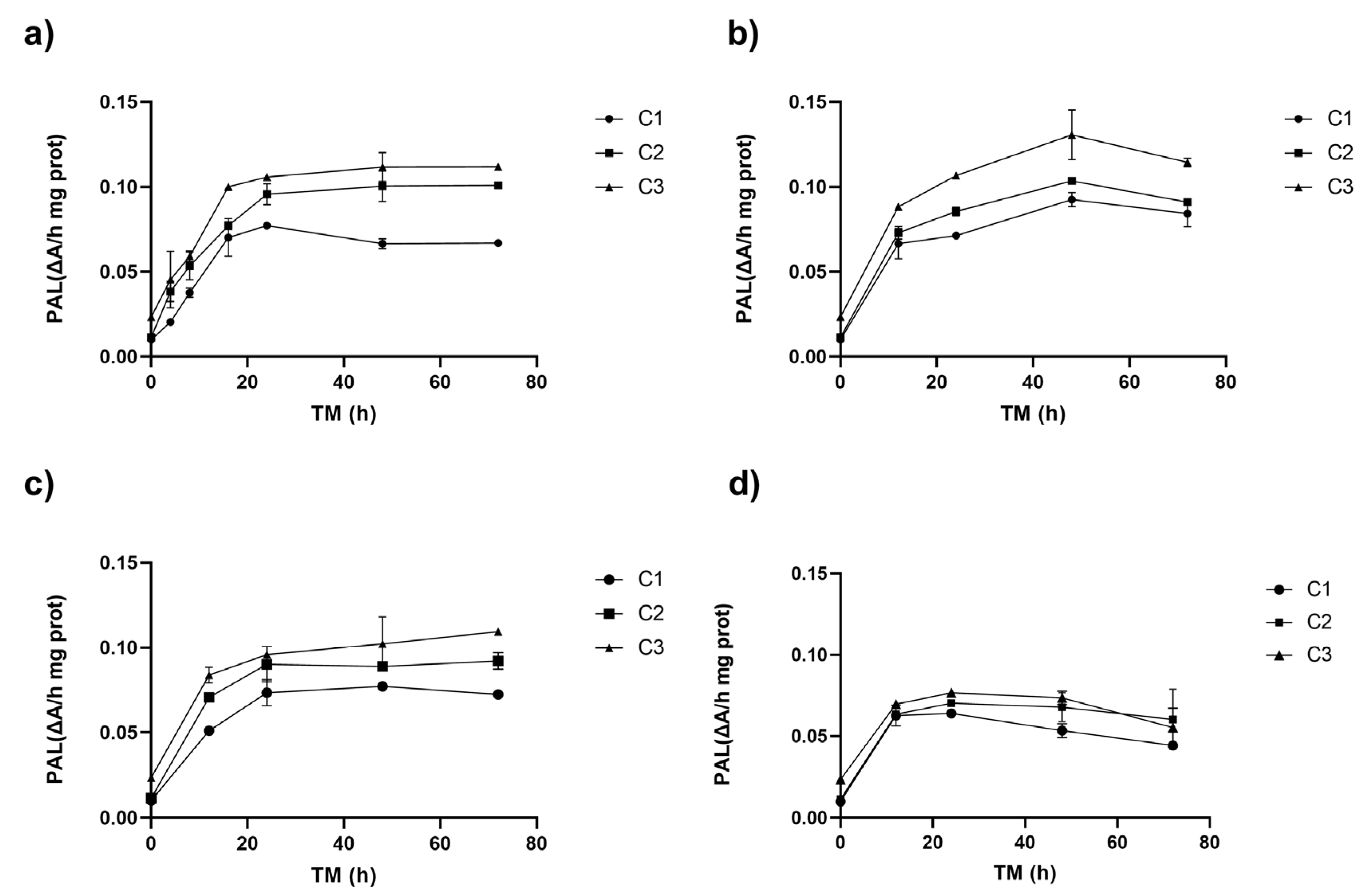
3.1.1. Effect on the Phenylalanine Ammonia-Lyase Activity
3.1.2. Effect on the Polyphenol Oxidase Activity
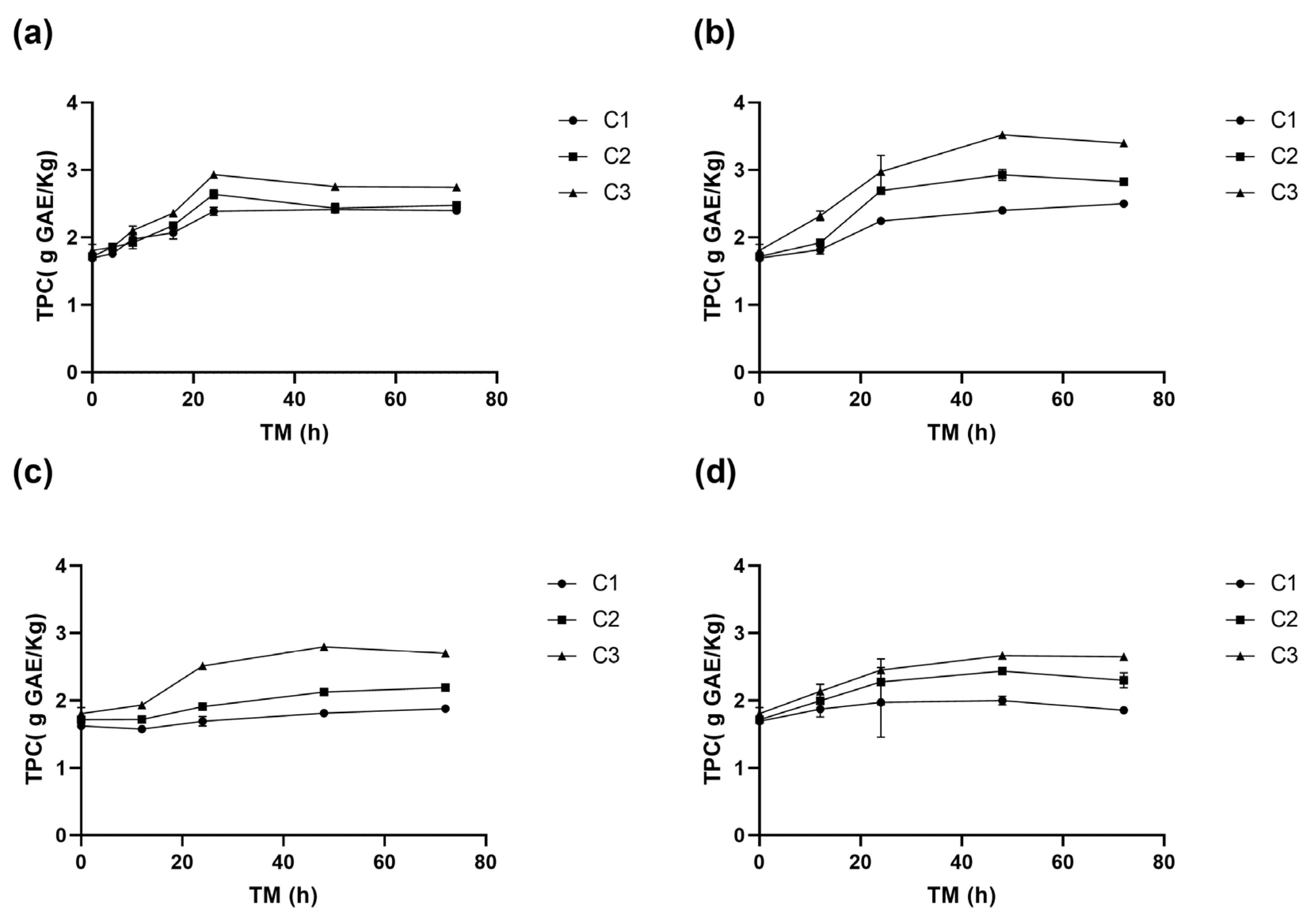
3.1.3. Effect on the Total Phenolic Content
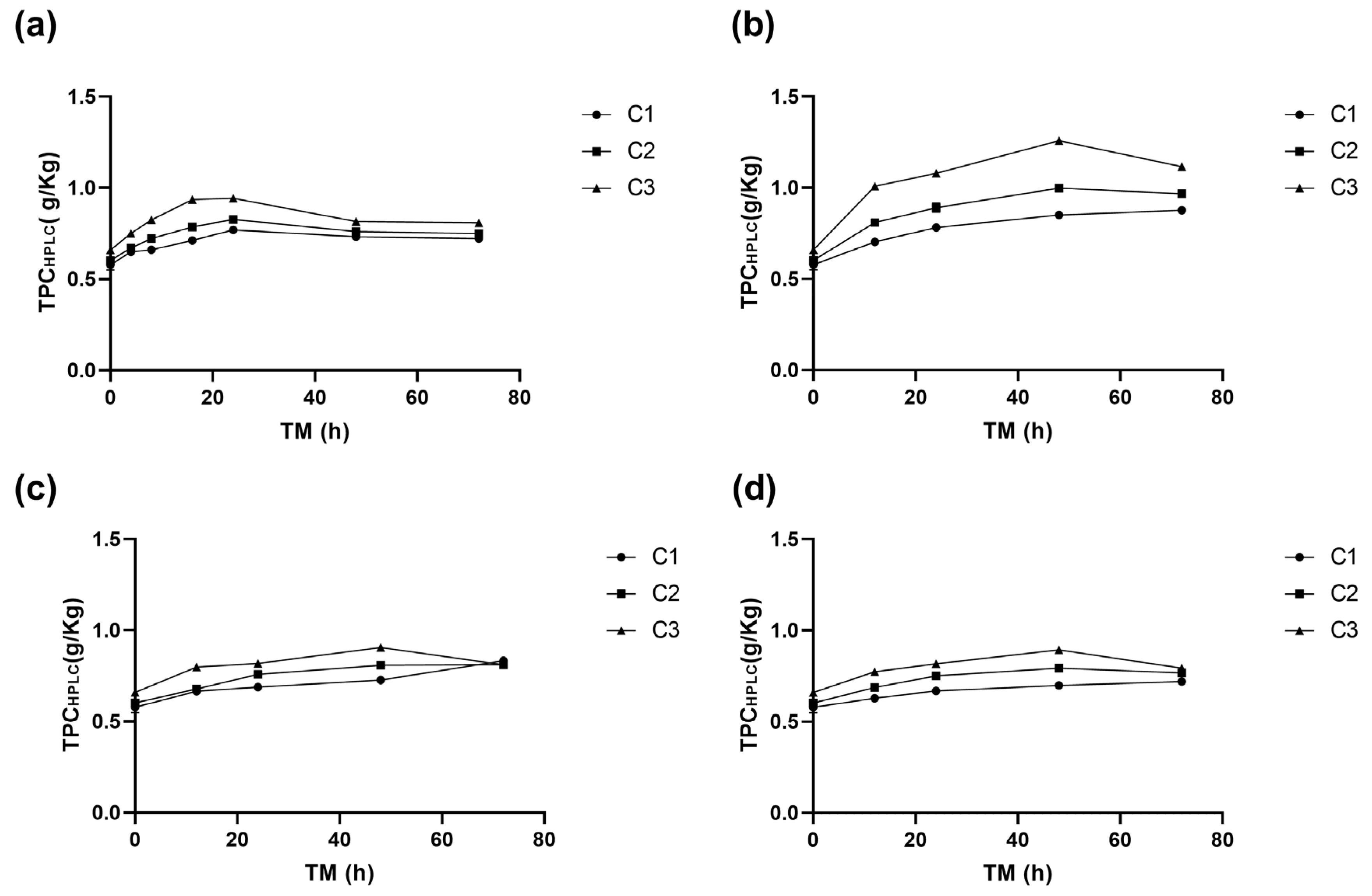
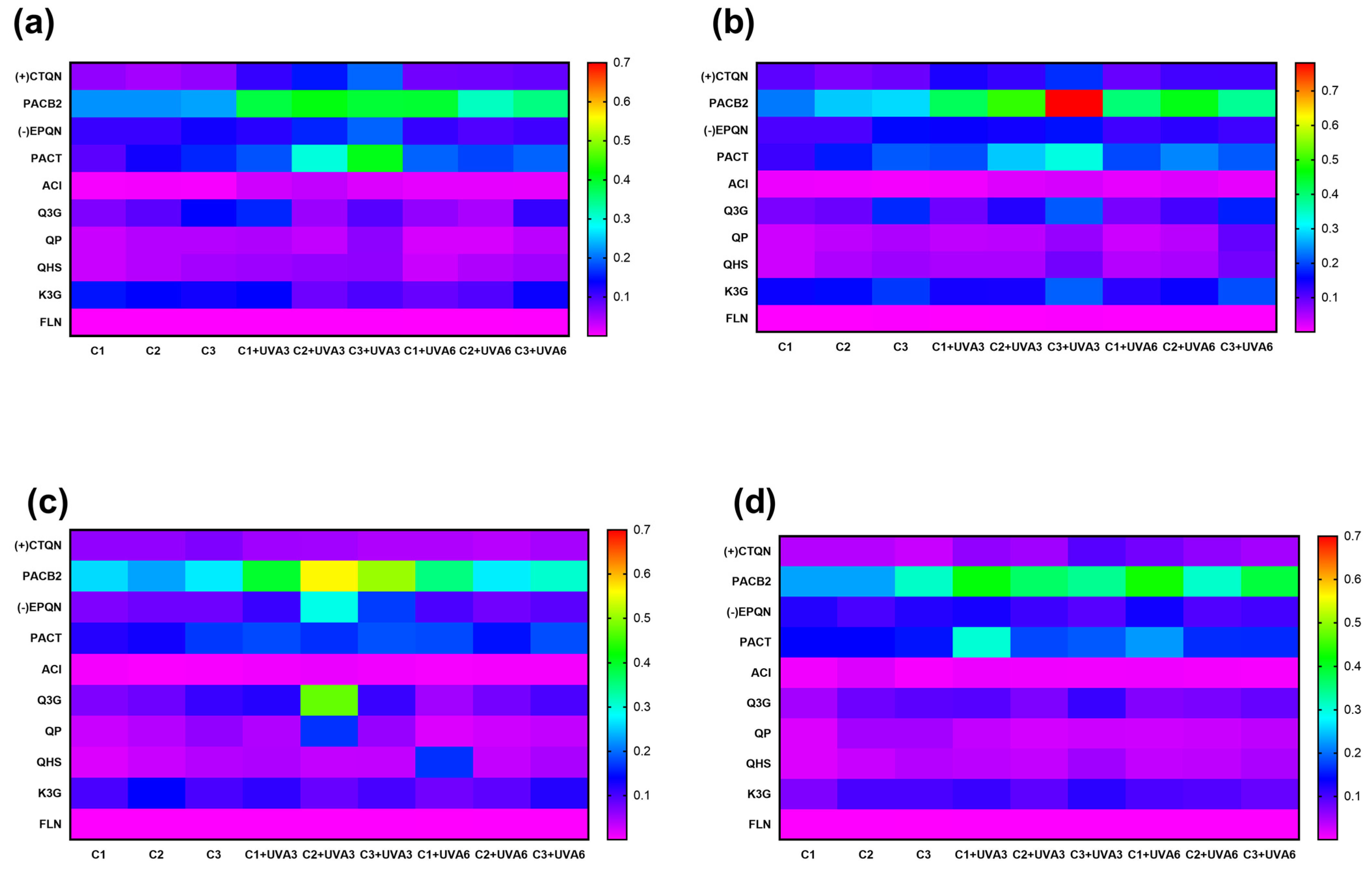
3.1.4. Effect on the Phenolic Compound Profile
3.1.5. Optimum Conditions of RM Biofortification Process Using Wounding-Induced Stress (Assay 1)
3.2. Assay 2: Combined Biofortification Process Using Wounding and UV-A Radiation Stress
3.2.1. Effect on PAL Enzyme Activity
3.2.2. Effect on PPO Enzyme Activity
3.2.3. Effect on Total Phenolic Compounds
3.2.4. Effect on Individual Phenolic Compounds
3.2.5. Optimum Experimental Variables for the Combined Biofortification Process Using Wounding and UV-A Radiation Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO—Food and Agriculture Organization Food and Agriculture Organization of the United Nations. (2023). FAOSTAT: Statistical Database. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 2 October 2024).
- Rodríguez-Arzuaga, M.; Salsi, M.S.; Piagentini, A.M. Storage Quality of Fresh-Cut Apples Treated with Yerba Mate (Ilex paraguariensis). J. Food Sci. Technol. 2020, 58, 186–196. [Google Scholar] [CrossRef]
- Scarano, P.; Sciarrillo, R.; Tartaglia, M.; Zuzolo, D.; Guarino, C. Circular Economy and Secondary Raw Materials from Fruits as Sustainable Source for Recovery and Reuse. A Review. Trends Food Sci. Technol. 2022, 122, 157–170. [Google Scholar] [CrossRef]
- Banerjee, J.; Singh, R.; Vijayaraghavan, R.; MacFarlane, D.; Patti, A.F.; Arora, A. Bioactives from Fruit Processing Wastes: Green Approaches to Valuable Chemicals. Food Chem. 2017, 225, 10–22. [Google Scholar] [CrossRef]
- Gazala, K.; Masoodi, F.A.; Masarat, H.D.; Rayees, B.; Shoib, M.W. Extraction and Characterisation of Pectin from Two Apple Juice Concentrate Processing Plants. Int. Food Res. J. 2017, 24, 594–599. [Google Scholar]
- Coman, V.; Teleky, B.E.; Mitrea, L.; Martău, G.A.; Szabo, K.; Călinoiu, L.F.; Vodnar, D.C. Bioactive Potential of Fruit and Vegetable Wastes. Adv. Food Nutr. Res. 2020, 91, 157–225. [Google Scholar] [CrossRef]
- Fecka, I.; Bednarska, K.; Włodarczyk, M. Fragaria × ananassa Cv. Senga Sengana Leaf: An Agricultural Waste with Antiglycation Potential and High Content of Ellagitannins, Flavonols, and 2-Pyrone-4,6-Dicarboxylic Acid. Molecules 2022, 27, 5293. [Google Scholar] [CrossRef]
- Galanakis, C.M. Food Waste Recovery: Processing Technologies and Industrial Techniques; Elsevier: Amsterdam, The Netherlands, 2015; ISBN 9780128004197. [Google Scholar]
- Ambroselli, D.; Masciulli, F.; Romano, E.; Catanzaro, G.; Besharat, Z.M.; Massari, M.C.; Ferretti, E.; Migliaccio, S.; Izzo, L.; Ritieni, A.; et al. New Advances in Metabolic Syndrome, from Prevention to Treatment: The Role of Diet and Food. Nutrients 2023, 15, 640. [Google Scholar] [CrossRef]
- Tarola, A.M.; Girelli, A.M.; D’Ascenzo, F. Bioactive Polyphenol Profiles and Antioxidant Activity in Italian Apples Varieties. Ital. J. Food Sci. 2019, 31, 243–252. [Google Scholar]
- Isah, T. Stress and Defense Responses in Plant Secondary Metabolites Production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef]
- Rabelo, M.C.; Bang, W.Y.; Nair, V.; Alves, R.E.; Jacobo-Velázquez, D.A.; Sreedharan, S.; de Miranda, M.R.A.; Cisneros-Zevallos, L. UVC Light Modulates Vitamin C and Phenolic Biosynthesis in Acerola Fruit: Role of Increased Mitochondria Activity and ROS Production. Sci. Rep. 2020, 10, 21972. [Google Scholar] [CrossRef]
- Yang, L.; Fanourakis, D.; Tsaniklidis, G.; Li, K.; Yang, Q.; Li, T. Contrary to Red, Blue Monochromatic Light Improves the Bioactive Compound Content in Broccoli Sprouts. Agronomy 2021, 11, 2139. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Xu, Y.; Yang, X.; Ji, Y.; Feng, K.; Sarengaowa. Metabolomics and Physiological Analyses Validates Previous Findings on the Mechanism of Response to Wounding Stress of Different Intensities in Broccoli. Food Res. Int. 2021, 140, 110058. [Google Scholar] [CrossRef]
- Edyta, Ł.; Iwona, C. Plant Responses to Wounding. In Biological Diversity—From Cell to Ecosystem; Wiley: Hoboken, NJ, USA, 2012; pp. 73–85. ISBN 9788362069286. [Google Scholar]
- Zapata, R.; Martínez-Zamora, L.; Cano-Lamadrid, M.; Artés-Hernández, F. Wounding Citrus Peel By-Products as Abiotic Stress to Induce the Synthesis of Phenolic Compounds? Horticulturae 2024, 10, 885. [Google Scholar] [CrossRef]
- Kang, H.M.; Saltveit, M.E. Antioxidant Capacity of Lettuce Leaf Tissue Increases after Wounding. J. Agric. Food Chem. 2002, 50, 7536–7541. [Google Scholar] [CrossRef]
- Becerra-Moreno, A.; Redondo-Gil, M.; Benavides, J.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Combined Effect of Water Loss and Wounding Stress on Gene Activation of Metabolic Pathways Associated with Phenolic Biosynthesis in Carrot. Front. Plant Sci. 2015, 6, 837. [Google Scholar] [CrossRef]
- Darré, M.; Vicente, A.R.; Cisneros-Zevallos, L.; Artés-Hernández, F. Postharvest Ultraviolet Radiation in Fruit and Vegetables: Applications and Factors Modulating Its Efficacy on Bioactive Compounds and Microbial Growth. Foods 2022, 11, 653. [Google Scholar] [CrossRef]
- Villamil-Galindo, E.; Antunes-Ricardo, M.; Piagentini, A.M.; Jacobo-Velázquez, D.A. Adding Value to Strawberry Agro-Industrial by-Products through Ultraviolet A-Induced Biofortification of Antioxidant and Anti-Inflammatory Phenolic Compounds. Front. Nutr. 2022, 9, 1080147. [Google Scholar] [CrossRef]
- Van de Velde, F.; Fenoglio, C.; Piagentini, A.M.; Pirovani, M.E. Modeling the Impact of the Type of Cutting and Storage Temperature on the Bioactive Compound Content, Phenylpropanoid Metabolism Enzymes and Quality Attributes of Fresh-Cut Strawberries. Food Bioprocess Technol. 2018, 11, 96–109. [Google Scholar] [CrossRef]
- Villamil-Galindo, E.; Piagentini, A.M. Kinetic Modeling of Valuable Phenolic Compounds Extraction from Strawberry and Apple Agro-industrial By-products. J. Food Process Eng. 2024, 47, e14573. [Google Scholar] [CrossRef]
- Villamil-Galindo, E.; Piagentini, A. Green Solvents for the Recovery of Phenolic Compounds from Strawberry (Fragaria x ananassa Duch) and Apple (Malus domestica) Agro-Industrial Bio-Wastes. Rev. Fac. Cienc. Agrar. UNCuyo 2024, 56, 149–160. [Google Scholar] [CrossRef]
- Reyes, L.F.; Cisneros-Zevallos, L. Wounding Stress Increases the Phenolic Content and Antioxidant Capacity of Purple-Flesh Potatoes (Solanum tuberosum L.). J. Agric. Food Chem. 2003, 51, 5296–5300. [Google Scholar] [CrossRef]
- Ibanez, F.; Bang, W.Y.; Lombardini, L.; Cisneros-Zevallos, L. Solving the Controversy of Healthier Organic Fruit: Leaf Wounding Triggers Distant Gene Expression Response of Polyphenol Biosynthesis in Strawberry Fruit (Fragaria x ananassa). Sci. Rep. 2019, 9, 19239. [Google Scholar] [CrossRef]
- Babaoglu Aydaş, S.; Ozturk, S.; Aslim, B. Phenylalanine Ammonia Lyase (PAL) Enzyme Activity and Antioxidant Properties of Some Cyanobacteria Isolates. Food Chem. 2013, 136, 164–169. [Google Scholar] [CrossRef]
- Xu, Z.; Luo, H.; Ji, A.; Zhang, X.; Song, J.; Chen, S. Global Identification of the Full-Length Transcripts and Alternative Splicing Related to Phenolic Acid Biosynthetic Genes in Salvia Miltiorrhiza. Front. Plant Sci. 2016, 7, 100. [Google Scholar] [CrossRef]
- Kong, J.-Q. Phenylalanine Ammonia-Lyase, a Key Component Used for Phenylpropanoids Production by Metabolic Engineering. RSC Adv. 2015, 5, 62587–62603. [Google Scholar] [CrossRef]
- Hu, W.; Guan, Y.; Wang, Y.; Yuan, N. Effect of Wounding Intensity on Edible Quality by Regulating Physiological and ROS Metabolism in Fresh-Cut Pumpkins. Horticulturae 2023, 9, 512. [Google Scholar] [CrossRef]
- Singh, B.; Suri, K.; Shevkani, K.; Kaur, A.; Kaur, A.; Singh, N. Enzymatic Browning of Fruit and Vegetables: A Review. In Enzymes in Food Technology: Improvements and Innovations; Springer: Cham, Switzerland, 2018; pp. 73–78. ISBN 9789811319334. [Google Scholar]
- Araji, S.; Grammer, T.A.; Gertzen, R.; Anderson, S.D.; Mikulic-Petkovsek, M.; Veberic, R.; Phu, M.L.; Solar, A.; Leslie, C.A.; Dandekar, A.M.; et al. Novel Roles for the Polyphenol Oxidase Enzyme in Secondary Metabolism and the Regulation of Cell Death in Walnut. Plant Physiol. 2014, 164, 1191–1203. [Google Scholar] [CrossRef]
- Serra, S.; Anthony, B.; Boscolo Sesillo, F.; Masia, A.; Musacchi, S. Determination of Post-Harvest Biochemical Composition, Enzymatic Activities, and Oxidative Browning in 14 Apple Cultivars. Foods 2021, 10, 186. [Google Scholar] [CrossRef]
- Torres-Contreras, A.M.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as Biofactories: Stress-Induced Production of Chlorogenic Acid Isomers in Potato Tubers as Affected by Wounding Intensity and Storage Time. Ind. Crops Prod. 2014, 62, 61–66. [Google Scholar] [CrossRef]
- Ortega-Hernández, E.; Nair, V.; Serrano-Sandoval, S.N.; Welti-Chanes, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Wounding and UVB Light Synergistically Induce the Postharvest Biosynthesis of Indicaxanthin and Betanin in Red Prickly Pears. Postharvest Biol. Technol. 2020, 167, 111247. [Google Scholar] [CrossRef]
- Santana-Gálvez, J.; Santacruz, A.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Postharvest Wounding Stress in Horticultural Crops as a Tool for Designing Novel Functional Foods and Beverages with Enhanced Nutraceutical Content: Carrot Juice as a Case Study. J. Food Sci. 2019, 84, 1151–1161. [Google Scholar] [CrossRef]
- Guyot, S.; Le Bourvellec, C.; Marnet, N.; Drilleau, J.F. Procyanidins Are the Most Abundant Polyphenols in Dessert Apples at Maturity. LWT Food Sci. Technol. 2002, 35, 289–291. [Google Scholar] [CrossRef]
- Hu, W.; Guan, Y.; Ji, Y.; Yang, X. Effect of Cutting Styles on Quality, Antioxidant Activity, Membrane Lipid Peroxidation, and Browning in Fresh-Cut Potatoes. Food Biosci. 2021, 44, 101435. [Google Scholar] [CrossRef]
- Kalinowska, M.; Bielawska, A.; Lewandowska-Siwkiewicz, H.; Priebe, W.; Lewandowski, W. Apples: Content of Phenolic Compounds vs. Variety, Part of Apple and Cultivation Model, Extraction of Phenolic Compounds, Biological Properties. Plant Physiol. Biochem. 2014, 84, 169–188. [Google Scholar] [CrossRef]
- Kim, I.; Ku, K.H.; Jeong, M.C.; Kim, S.S.; Mitchell, A.E.; Lee, J. A Comparison of the Chemical Composition and Antioxidant Activity of Several New Early- to Mid-Season Apple Cultivars for a Warmer Climate with Traditional Cultivars. J. Sci. Food Agric. 2019, 99, 4712–4724. [Google Scholar] [CrossRef]
- Rosales, T.K.O.; Fabi, J.P. Valorization of Polyphenolic Compounds from Food Industry By-Products for Application in Polysaccharide-Based Nanoparticles. Front. Nutr. 2023, 10, 1144677. [Google Scholar] [CrossRef]
- Escarpa, A.; González, M.C. High-Performance Liquid Chromatography with Diode-Array Detection for the Determination of Phenolic Compounds in Peel and Pulp from Different Apple Varieties. J. Chromatogr. A 1998, 823, 331–337. [Google Scholar] [CrossRef]
- Diemer, E.; Morad, C.; Mouterde, L.; Grimi, N.; Ioannou, I. Postharvest Caffeoylquinic Acid Accumulation in Forced Chicory Roots: Insights into the Role of Temperature, Water Loss, and Biological Defense Mechanisms. ACS Food Sci. Technol. 2024, 4, 470–478. [Google Scholar] [CrossRef]
- Rammohan, A.; Reddy, J.S.; Sravya, G.; Rao, C.N.; Zyryanov, G.V. Chalcone Synthesis, Properties and Medicinal Applications: A Review. Environ. Chem. Lett. 2020, 18, 433–458. [Google Scholar] [CrossRef]
- Song, Y.; Ma, B.; Guo, Q.; Zhou, L.; Lv, C.; Liu, X.; Wang, J.; Zhou, X.; Zhang, C. UV-B Induces the Expression of Flavonoid Biosynthetic Pathways in Blueberry (Vaccinium corymbosum) Calli. Front. Plant Sci. 2022, 13, 1079087. [Google Scholar] [CrossRef]
- Lante, A.; Tinello, F.; Nicoletto, M. UV-A Light Treatment for Controlling Enzymatic Browning of Fresh-Cut Fruits. Innov. Food Sci. Emerg. Technol. 2016, 34, 141–147. [Google Scholar] [CrossRef]
- Ortega-Hernández, E.; Welti-Chanes, J.; Jacobo-Velázquez, D.A. Effects of UVB Light, Wounding Stress, and Storage Time on the Accumulation of Betalains, Phenolic Compounds, and Ascorbic Acid in Red Prickly Pear (Opuntia Ficus-Indica Cv. Rojo Vigor). Food Bioprocess Technol. 2018, 11, 2265–2274. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Han, C.; Ji, N.; Jin, P.; Zheng, Y. UV-C Treatment Maintains Quality and Enhances Antioxidant Capacity of Fresh-Cut Strawberries. Postharvest Biol. Technol. 2019, 156, 110945. [Google Scholar] [CrossRef]
- Rai, N.; Morales, L.O.; Aphalo, P.J. Perception of Solar UV Radiation by Plants: Photoreceptors and Mechanisms. Plant Physiol. 2021, 186, 1382–1396. [Google Scholar] [CrossRef]
- Guo, J.; Wang, M.H. Ultraviolet A-Specific Induction of Anthocyanin Biosynthesis and PAL Expression in Tomato (Solanum lycopersicum L.). Plant Growth Regul. 2010, 62, 1–8. [Google Scholar] [CrossRef]
- Khanal, R.C.; Howard, L.R.; Brownmiller, C.R.; Prior, R.L. Influence of Extrusion Processing on Procyanidin Composition and Total Anthocyanin Contents of Blueberry Pomace. J. Food Sci. 2009, 74, H52–H58. [Google Scholar] [CrossRef]
- Rue, E.; Rush, M.; Van Breemen, R. Procyanidins: A Comprehensive Review Encompassing Structure Elucidation via Mass Spectrometry. Phytochem 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Kimmel, E.M.; Jerome, M.; Holderness, J.; Snyder, D.; Kemoli, S.; Jutila, M.A.; Hedges, J.F. Oligomeric Procyanidins Stimulate Innate Antiviral Immunity in Dengue Virus Infected Human PBMCs. Antivir. Res. 2011, 90, 80–86. [Google Scholar] [CrossRef]
- Rodrigues, J.P.B.; Liberal, Â.; Petropoulos, S.A.; Ferreira, I.C.F.R.; Oliveira, M.B.P.P.; Fernandes, Â.; Barros, L. Agri-Food Surplus, Waste and Loss as Sustainable Biobased Ingredients: A Review. Molecules 2022, 27, 5200. [Google Scholar] [CrossRef]
- Martínez-Zamora, L.; Castillejo, N.; Artés-Hernández, F. Postharvest UV-B and Photoperiod with Blue + Red LEDs as Strategies to Stimulate Carotenogenesis in Bell Peppers. Appl. Sci. 2021, 11, 3736. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ct | TM (h) | (+)CTQN | PACB2 | (-)EPQN | PACT | ACl | Q3G | QP | QHS | K3G | FLN |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C1 | 0 | 0.02 ± 0.002 Ci | 0.24 ± 0.005 Aab | 0.10 ± 0.003 Ce | 0.08 ± 0.02 Af | 0.004 ± 0.001 Cg | 0.04 ± 0.003 Ci | 0.01 ± 0.001 Bb | 0.02 ± 0.0003 De | 0.06 ± 0.01 Cg | 0.0002 ± 0.00003 De |
| 12 | 0.06 ± 0.001 Bgh | 0.20 ± 0.003 Cde | 0.10 ± 0.002 BCde | 0.07 ± 0.01 Afgi | 0.004 ± 0.0001 BCfg | 0.07 ± 0.0003 Bef | 0.03 ± 0.0004 Aab | 0.04 ± 0.0004 Acd | 0.10 ± 0.002 Bdef | 0.0006 ± 0.000001 Ccde | |
| 24 | 0.06 ± 0.002 Bfg | 0.21 ± 0.002 BCde | 0.11 ± 0.003 ABc | 0.08 ± 0.003 Afg | 0.005 ± 0.00001 Bef | 0.07 ± 0.001 BABe | 0.03 ± 0.001 Aab | 0.04 ± 0.0001 Abcd | 0.10 ± 0.01 Bb-f | 0.0009 ± 0.0001 Bbc | |
| 48 | 0.06 ± 0.001 Bcde | 0.22 ± 0.0001 Bcd | 0.11 ± 0.01 Ac | 0.09 ± 0.007 Af | 0.005 ± 0.00002 Bef | 0.07 ± 0.0005 ABe | 0.03 ± 0.0002 Aab | 0.03 ± 0.001 Bd | 0.15 ± 0.002 Aa | 0.001 ± 0.00002 ABab | |
| 72 | 0.07 ± 0.001 Aef | 0.21 ± 0.003 Cde | 0.11 ± 0.01 Ac | 0.05 ± 0.01 Ai | 0.013 ± 0.0003 Aa | 0.08 ± 0.0003 Ade | 0.03 ± 0.0001 Aab | 0.03 ± 0.0001 Cde | 0.15 ± 0.01 Aa | 0.0011 ± 0.00002 Aab | |
| C2 | 0 | 0.01 ± 0.002 Bi | 0.21 ± 0.01 ABde | 0.08 ± 0.001 Df | 0.09 ± 0.002 Cef | 0.002 ± 0.00002 Ci | 0.05 ± 0.002 Bgi | 0.04 ± 0.005 Aa | 0.03 ± 0.01 Ade | 0.09 ± 0.02 Cfg | 0.0003 ± 0.00005 Be |
| 12 | 0.05 ± 0.002 Bef | 0.20 ± 0.003 Be | 0.11 ± 0.0003 Bcd | 0.12 ± 0.001 Bde | 0.003 ± 0.00002 Cgi | 0.09 ± 0.005 Acd | 0.04 ± 0.0001 Aab | 0.03 ± 0.001 Ad | 0.09 ± 0.008 Cfg | 0.0009 ± 0.0003 ABbcd | |
| 24 | 0.05 ± 0.002 Bbcd | 0.21 ± 0.005 ABde | 0.11 ± 0.001 ABc | 0.13 ± 0.001 Aa-d | 0.005 ± 0.0006 Bde | 0.09 ± 0.001 Acd | 0.04 ± 0.0003 Aab | 0.03 ± 0.002 Ad | 0.11 ± 0.001 ABa-e | 0.0011 ± 0.0001 Aab | |
| 48 | 0.05 ± 0.001 Bb | 0.22 ± 0.002 Abcd | 0.11 ± 0.001 Ac | 0.13 ± 0.001 Abcd | 0.006 ± 0.0005 Bd | 0.09 ± 0.0005 Ac | 0.04 ± 0.0006 Aa | 0.04 ± 0.0003 Acd | 0.14 ± 0.001 Aabc | 0.0011 ± 0.00003 Aab | |
| 72 | 0.07 ± 0.003 Adef | 0.22 ± 0.0001 A | 0.10 ± 0.001 Ce | 0.05 ± 0.001 Dgi | 0.012 ± 0.0004 Ab | 0.09 ± 0.0004 Ac | 0.04 ± 0.00002 Aa | 0.03 ± 0.002 Ad | 0.15 ± 0.0003 Aab | 0.0011 ± 0.00001 A | |
| C3 | 0 | 0.03 ± 0.002 Ch | 0.17 ± 0.003 Df | 0.10 ± 0.003 De | 0.13 ± 0.001 Bcd | 0.002 ± 0.00003 Di | 0.05 ± 0.001 Dfg | 0.05 ± 0.0005 Aa | 0.04 ± 0.002 Cbcd | 0.09 ± 0.01 Cef | 0.0004 ± 0.00003 Cde |
| 12 | 0.05 ± 0.003 Bb | 0.23 ± 0.005 ABabc | 0.12 ± 0.001 Bb | 0.15 ± 0.002 Aabc | 0.003 ± 0.0001 Cgi | 0.10 ± 0.0006 Cc | 0.02 ± 0.03 Aab | 0.03 ± 0.0007 Dde | 0.11 ± 0.007 BCc-f | 0.0009 ± 0.00004 Bab | |
| 24 | 0.06 ± 0.001 ABa | 0.24 ± 0.001 ABa | 0.13 ± 0.002 ABa | 0.16 ± 0.002 Aab | 0.004 ± 0.0002 Bfg | 0.13 ± 0.001 Bb | 0.05 ± 0.0001 Aa | 0.05 ± 0.0007 Aa | 0.12 ± 0.002 ABa-d | 0.0011 ± 0.0001 ABab | |
| 48 | 0.06 ± 0.001 Aa | 0.23 ± 0.001 Babc | 0.13 ± 0.001 Aa | 0.16 ± 0.001 Ba | 0.004 ± 0.0002 Bfg | 0.14 ± 0.0001 ABab | 0.04 ± 0.001 Aa | 0.05 ± 0.0004 Babc | 0.13 ± 0.005 Abc | 0.0012 ± 0.00001 ABab | |
| 72 | 0.06 ± 0.001 Abc | 0.21 ± 0.001 Cde | 0.11 ± 0.01 Cc | 0.05 ± 0.01 Ci | 0.009 ± 0.0001 Ac | 0.15 ± 0.01 Aa | 0.05 ± 0.0005 Aa | 0.05 ± 0.0009 ABab | 0.14 ± 0.001 Aab | 0.0014 ± 0.00013 Aa |
| Ct | TM (h) | (+)CTQN | PACB2 | (-)EPQN | PACT | ACl | Q3G | QP | QHS | K3G | FLN |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C1 | 0 | 0.02 ± 0.001 Cgi | 0.24 ± 0.004 Acde | 0.10 ± 0.003 Aefg | 0.08 ± 0.01 Ai | 0.004 ± 0.001 Cgi | 0.04 ± 0.002 Dl | 0.01 ± 0.001 Ci | 0.02 ± 0.0003 De | 0.06 ± 0.01 Cg | 0.0002 ± 0.00002 Eg |
| 12 | 0.05 ± 0.003 Ce | 0.22 ± 0.004 Bfg | 0.11 ± 0.001 Adef | 0.09 ± 0.005 Ai | 0.006 ± 0.0001 Bef | 0.08 ± 0.0003 Bg | 0.03 ± 0.0003 Bg | 0.02 ± 0.001 Cde | 0.1 ± 0.0001 Bef | 0.002 ± 0.00007 Bef | |
| 24 | 0.08 ± 0.0001 Bd | 0.22 ± 0.005 ABefg | 0.10 ± 0.006 Ag | 0.10 ± 0.004 Afgi | 0.007 ± 0.0002 Bef | 0.07 ± 0.0016 Ci | 0.03 ± 0.00001 Afg | 0.03 ± 0.001 Bcd | 0.14 ± 0.002 Abc | 0.0025 ± 0.00003 Abc | |
| 48 | 0.10 ± 0.003 Aab | 0.23 ± 0.002 ABd-g | 0.11 ± 0.005 Ad-g | 0.12 ± 0.001 Aef | 0.011 ± 0.001 Abc | 0.08 ± 0.0007 Bg | 0.03 ± 0.001 Bg | 0.03 ± 0.001 Bcd | 0.15 ± 0.0004 Abc | 0.001 ± 0.00002 Dbc | |
| 72 | 0.10 ± 0.002 Aab | 0.23 ± 0.009 ABd-g | 0.11 ± 0.0007 Ad-g | 0.12 ± 0.004 Aefg | 0.012 ± 0.0001 Ab | 0.09 ± 0.002 Af | 0.03 ± 0.001 Afg | 0.04 ± 0.001 Abc | 0.14 ± 0.002 Abc | 0.0015 ± 0.00003 Cbc | |
| C2 | 0 | 0.01 ± 0.001 Di | 0.21 ± 0.01 Bfg | 0.08 ± 0.001 Ci | 0.09 ± 0.001 Cgi | 0.002 ± 0.001 Di | 0.05 ± 0.002 Dk | 0.04 ± 0.005 ABde | 0.03 ± 0.01 Acde | 0.09 ± 0.02 Bfg | 0.0003 ± 0.00005 Efg |
| 12 | 0.06 ± 0.001 Ce | 0.22 ± 0.001 Bfg | 0.12 ± 0.0003 Ad | 0.14 ± 0.003 Bde | 0.006 ± 0.0003 Cef | 0.10 ± 0.001 Ad | 0.04 ± 0.001 Bef | 0.04 ± 0.0006 Abc | 0.09 ± 0.001 Bfg | 0.0014 ± 0.0006 Bfg | |
| 24 | 0.06 ± 0.001 Cfe | 0.23 ± 0.005 Bdef | 0.12 ± 0.0005 Ad | 0.1 ± 0.006 Abc | 0.008 ± 0.001 BCde | 0.09 ± 0.001 Cf | 0.05 ± 0.001 Abc | 0.04 ± 0.0002 Abcd | 0.12 ± 0.01 ABc-f | 0.0026 ± 0.00001 Ac-f | |
| 48 | 0.08 ± 0.01 Bcd | 0.28 ± 0.005 Aab | 0.11 ± 0.01 ABde | 0.17 ± 0.0004 Abc | 0.009 ± 0.002 Bcd | 0.09 ± 0.0002 BCef | 0.04 ± 0.0006 ABcde | 0.05 ± 0.002 Aab | 0.16 ± 0.001 Ab | 0.001 ± 0.00002 Cb | |
| 72 | 0.11 ± 0.002 Aa | 0.28 ± 0.008 Aab | 0.10 ± 0.00004 Befg | 0.16 ± 0.003 Ac | 0.015 ± 0.003 Aa | 0.10 ± 0.002 ABde | 0.04 ± 0.0005 Bef | 0.05 ± 0.0007 Aab | 0.11 ± 0.001 Bdef | 0.0009 ± 0.0004 Ddef | |
| C3 | 0 | 0.03 ± 0.001 Dg | 0.17 ± 0.003 Di | 0.10 ± 0.002 Dfg | 0.13 ± 0.001 De | 0.002 ± 0.00003 Ci | 0.05 ± 0.001 Dj | 0.05 ± 0.0004 Cbcd | 0.04 ± 0.001 Dbcd | 0.09 ± 0.006 Cfg | 0.0004 ± 0.00004 Dfg |
| 12 | 0.06 ± 0.001 Ce | 0.26 ± 0.001 Bbc | 0.17 ± 0.001 Aa | 0.17 ± 0.002 Bbc | 0.006 ± 0.0003 Bef | 0.12 ± 0.0003 Cc | 0.05 ± 0.005 Cbcd | 0.05 ± 0.0001 Cab | 0.11 ± 0.001 BCc-f | 0.0021 ± 0.00001 Bc-f | |
| 24 | 0.08 ± 0.002 Bdf | 0.24 ± 0.004 Ccd | 0.14 ± 0.0008 Cb | 0.15 ± 0.002 Ccd | 0.006 ± 0.00001 Bfg | 0.16 ± 0.002 Bb | 0.10 ± 0.0005 Aa | 0.06 ± 0.001 Ba | 0.14 ± 0.001 Bbcd | 0.0036 ± 0.00001 Abcd | |
| 48 | 0.09 ± 0.005 Abc | 0.29 ± 0.005 Aa | 0.16 ± 0.004 Bb | 0.21 ± 0.005 Aa | 0.007 ± 0.0003 Bdef | 0.18 ± 0.001 Aa | 0.05 ± 0.002 Bb | 0.06 ± 0.0002 Aa | 0.19 ± 0.0004 Aa | 0.0018 ± 0.00003 Ba | |
| 72 | 0.10 ± 0.003 Aab | 0.29 ± 0.001 Aa | 0.15 ± 0.002 Cbc | 0.19 ± 0.008 Bb | 0.012 ± 0.0001 Ab | 0.16 ± 0.001 Bb | 0.04 ± 0.001 Dde | 0.05 ± 0.003 Cab | 0.13 ± 0.02 Bb-e | 0.0014 ± 0.00002 Cb-e |
| Ct | TM (h) | (+)CTQN | PACB2 | (-)EPQN | PACT | ACl | Q3G | QP | QHS | K3G | FLN |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C1 | 0 | 0.02 ± 0.001 Di | 0.24 ± 0.004 BCef | 0.10 ± 0.003 Aab | 0.08 ± 0.01 Bj | 0.004 ± 0.001 Bef | 0.04 ± 0.002 Cj | 0.01 ± 0.001 Bk | 0.02 ± 0.0003 Bg | 0.06 ± 0.01 Ch | 0.0002 ± 0.00002 Cf |
| 12 | 0.04 ± 0.001 Cg | 0.23 ± 0.006 Cf | 0.10 ± 0.002 Aa | 0.11 ± 0.01 AB | 0.005 ± 0.0003 ABcd | 0.05 ± 0.002 Bi | 0.03 ± 0.0005 Aj | 0.02 ± 0.0003 Bfg | 0.08 ± 0.001 BCefg | 0.0026 ± 0.00005 Ac | |
| 24 | 0.05 ± 0.002 Cf | 0.25 ± 0.004 BCde | 0.05 ± 0.004 Ch | 0.12 ± 0.006 Abe–h | 0.005 ± 0.0002 ABbc | 0.06 ± 0.002 Ag | 0.03 ± 0.0009 Ahi | 0.02 ± 0.001 Befg | 0.11 ± 0.008 ABbc | 0.0003 ± 0.00007 BCef | |
| 48 | 0.06 ± 0.004 Bde | 0.26 ± 0.001 Bcd | 0.07 ± 0.004 B | 0.12 ± 0.008 ABefg | 0.006 ± 0.0004 Aab | 0.07 ± 0.0005 Aef | 0.03 ± 0.001 Ahi | 0.02 ± 0.011 Bfg | 0.1 ± 0.0003 BCc-f | 0.0005 ± 0.00001 Bef | |
| 72 | 0.07 ± 0.002 Ab | 0.28 ± 0.005 Ab | 0.06 ± 0.0002 BCg | 0.15 ± 0.01 Abc | 0.008 ± 0.0007 Aa | 0.06 ± 0.00001 Afg | 0.03 ± 0.0005 Ahi | 0.03 ± 0.004 Acde | 0.15 ± 0.02 Aa | 0.0003 ± 0.00001 BCef | |
| C2 | 0 | 0.01 ± 0.001 Dj | 0.21 ± 0.01 ABg | 0.08 ± 0.001 Cde | 0.09 ± 0.001 Cij | 0.002 ± 0.001 Bg | 0.05 ± 0.002 Ci | 0.04 ± 0.005 Ade | 0.03 ± 0.013 Adef | 0.09 ± 0.02 Bd-g | 0.0003 ± 0.00005 Bef |
| 12 | 0.05 ± 0.001 Cf | 0.20 ± 0.004 Bg | 0.09 ± 0.001 Bc | 0.11 ± 0.006 Bghi | 0.002 ± 0.002 Bg | 0.06 ± 0.002 Bg | 0.04 ± 0.001 Afg | 0.04 ± 0.001 Aabc | 0.08 ± 0.001 Bfg | 0.0035 ± 0.0004 Ab | |
| 24 | 0.06 ± 0.001 Be | 0.21 ± 0.008 ABg | 0.07 ± 0.001 Df | 0.13 ± 0.004 ABdef | 0.003 ± 0.002 ABfg | 0.07 ± 0.002 ABde | 0.04 ± 0.001 Aef | 0.04 ± 0.001 Aab | 0.15 ± 0.001 Aa | 0.0007 ± 0.0003 Bde | |
| 48 | 0.06 ± 0.001 Bde | 0.23 ± 0.003 ABf | 0.08 ± 0.0008 Ccd | 0.13 ± 0.003 Acde | 0.003 ± 0.002 ABef | 0.08 ± 0.005 Ab | 0.04 ± 0.0003 Ade | 0.03 ± 0.0002 Abcd | 0.14 ± 0.001 Aa | 0.0005 ± 0.00003 Bef | |
| 72 | 0.09 ± 0.002 Aa | 0.24 ± 0.0004 Aef | 0.09 ± 0.001 Ab | 0.14 ± 0.004 Abcd | 0.004 ± 0.002 Ade | 0.07 ± 0.001 ABfg | 0.03 ± 0.002 Agh | 0.03 ± 0.0003 Abcd | 0.11 ± 0.001 ABbc | 0.0003 ± 0.0001 Bef | |
| C3 | 0 | 0.03 ± 0.001 Ch | 0.17 ± 0.003 Dh | 0.10 ± 0.002 Aab | 0.13 ± 0.004 BCd-g | 0.002 ± 0.00003 Bg | 0.05 ± 0.001 Ch | 0.05 ± 0.0004 Bbc | 0.04 ± 0.001 Aabc | 0.09 ± 0.006 BCd-g | 0.0004 ± 0.00004 Bef |
| 12 | 0.06 ± 0.001 Bde | 0.25 ± 0.003 Cde | 0.10 ± 0.001 Aa | 0.13 ± 0.003 BCcde | 0.003 ± 0.00003 ABefg | 0.07 ± 0.003 Bde | 0.04 ± 0.0007 Bcd | 0.04 ± 0.002 Aa | 0.10 ± 0.004 Bcde | 0.004 ± 0.0002 Aa | |
| 24 | 0.06 ± 0.0004 Bd | 0.26 ± 0.001 BCc | 0.07 ± 0.001 Cf | 0.15 ± 0.007 ABb | 0.003 ± 0.0002 ABef | 0.07 ± 0.0007 Bcd | 0.05 ± 0.001 Bbc | 0.03 ± 0.0004 Bbcd | 0.12 ± 0.001 Ab | 0.001 ± 0.0002 Bd | |
| 48 | 0.07 ± 0.004 ABc | 0.27 ± 0.004 Bb | 0.08 ± 0.001 Bde | 0.17 ± 0.007 Aa | 0.004 ± 0.001 ABef | 0.11 ± 0.003 Aa | 0.06 ± 0.002 Aa | 0.04 ± 0.001 Aab | 0.10 ± 0.002 Bbcd | 0.001 ± 0.0001 Bd | |
| 72 | 0.07 ± 0.0003 Ab | 0.34 ± 0.005 Aa | 0.07 ± 0.004 BCe | 0.10 ± 0.02 Chi | 0.004 ± 0.0002 Ade | 0.08 ± 0.0005 Bbc | 0.05 ± 0.0004 Bb | 0.02 ± 0.001 Cfg | 0.08 ± 0.001 Cgh | 0.0003 ± 0.0002 Bef |
| Ct | TM (h) | (+)CTQN | PACB2 | (-)EPQN | PACT | ACl | Q3G | QP | QHS | K3G | FLN |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C1 | 0 | 0.02 ± 0.001 Cg | 0.24 ± 0.004 Acde | 0.10 ± 0.003 Bc | 0.08 ± 0.01 Cg | 0.004 ± 0.0008 BCb | 0.04 ± 0.002 Be | 0.01 ± 0.001 Be | 0.02 ± 0.001 Cg | 0.06 ± 0.012 B | 0.0002 ± 0.0002 Bi |
| 12 | 0.04 ± 0.001 Aabc | 0.22 ± 0.02 ABdef | 0.12 ± 0.003 Adef | 0.10 ± 0.01 BCf | 0.006 ± 0.001 ABab | 0.04 ± 0.001 Bde | 0.02 ± 0.01 ABde | 0.02 ± 0.001 Cfg | 0.06 ± 0.001 AB | 0.0007 ± 0.0001 Ai | |
| 24 | 0.04 ± 0.002 ABbcd | 0.21 ± 0.01 ABef | 0.12 ± 0.004 Aef | 0.14 ± 0.001 Bcd | 0.007 ± 0.0001 Aab | 0.04 ± 0.001 Bde | 0.02 ± 0.001 ABd | 0.02 ± 0.001 Cfg | 0.07 ± 0.004 AB | 0.0003 ± 0.00005 ABhi | |
| 48 | 0.04 ± 0.005 Aab | 0.23 ± 0.01 ABcde | 0.12 ± 0.004 Acde | 0.14 ± 0.01 Bcd | 0.007 ± 0.003 Aab | 0.05 ± 0.01 Bde | 0.02 ± 0.001 ABd | 0.02 ± 0.002 Bef | 0.07 ± 0.01 AB | 0.0004 ± 0.0002 ABf-i | |
| 72 | 0.03 ± 0.0002 BCfg | 0.18 ± 0.02 Bgh | 0.05 ± 0.004 Cfg | 0.19 ± 0.04 Aa | 0.003 ± 0.0004 Cb | 0.09 ± 0.01 Aa | 0.05 ± 0.01 Aab | 0.04 ± 0.01 Ab | 0.09 ± 0.007 A | 0.0004 ± 0.00002 ABb-e | |
| C2 | 0 | 0.01 ± 0.001 Dh | 0.21 ± 0.01 Aef | 0.08 ± 0.001 BCef | 0.09 ± 0.02 Dfg | 0.002 ± 0.0007 Ab | 0.05 ± 0.002 Cde | 0.04 ± 0.005 Bbc | 0.03 ± 0.001 Aef | 0.09 ± 0.02 A | 0.0003 ± 0.00005 Bc-f |
| 12 | 0.03 ± 0.001 Cfg | 0.23 ± 0.02 Acd | 0.08 ± 0.002 ABCcd | 0.12 ± 0.004 Ce | 0.005 ± 0.001 Aab | 0.07 ± 0.002 BCc | 0.04 ± 0.02 Bc | 0.03 ± 0.001 Acde | 0.08 ± 0.002 A | 0.0007 ± 0.0002 Ad-g | |
| 24 | 0.04 ± 0.002 Bbcd | 0.24 ± 0.01 Acd | 0.09 ± 0.001 ABcd | 0.12 ± 0.004 Ce | 0.005 ± 0.002 Aab | 0.09 ± 0.008 Aa | 0.04 ± 0.001 Bbc | 0.03 ± 0.001 Ade | 0.10 ± 0.002 A | 0.0005 ± 0.00018 ABabc | |
| 48 | 0.04 ± 0.001 Aa | 0.23 ± 0.01 Acde | 0.10 ± 0.001 Acde | 0.14 ± 0.007 Bc | 0.002 ± 0.0004 Aab | 0.08 ± 0.008 ABb | 0.05 ± 0.001 Aa | 0.03 ± 0.01 Ade | 0.10 ± 0.02 A | 0.0005 ± 0.00008 ABa-d | |
| 72 | 0.03 ± 0.0003 Bcde | 0.20 ± 0.02 Afg | 0.07 ± 0.01 Cfg | 0.17 ± 0.004 Ab | 0.02 ± 0.0004 Aa | 0.09 ± 0.002 Aa | 0.04 ± 0.01 Bbc | 0.04 ± 0.004 Abcd | 0.10 ± 0.004 A | 0.0004 ± 0.00008 ABab | |
| C3 | 0 | 0.03 ± 0.001 Ade | 0.17 ± 0.003 Dh | 0.10 ± 0.002 Bg | 0.13 ± 0.01 Cde | 0.002 ± 0.0003 Bb | 0.05 ± 0.001 Bd | 0.05 ± 0.003 Aabc | 0.04 ± 0.001 Bbcd | 0.09 ± 0.006 BC | 0.0004 ± 0.00004 Ab-e |
| 12 | 0.03 ± 0.001 Ade | 0.27 ± 0.02 Bb | 0.09 ± 0.006 Bb | 0.12 ± 0.01 Ce | 0.003 ± 0.001 ABb | 0.07 ± 0.001 Bc | 0.04 ± 0.003 Abc | 0.04 ± 0.001 Bb | 0.10 ± 0.03 AB | 0.0006 ± 0.00009 Aabc | |
| 24 | 0.04 ± 0.002 A | 0.27 ± 0.01 Bb | 0.10 ± 0.003 ABb | 0.14 ± 0.005 BC | 0.004 ± 0.0005 A | 0.07 ± 0.003 Bbc | 0.04 ± 0.001 Abc | 0.04 ± 0.01 ABbc | 0.11 ± 0.001 A | 0.0006 ± 0.00004 Aa | |
| 48 | 0.03 ± 0.001 Ade | 0.23 ± 0.01 Aa | 0.12 ± 0.002 Aa | 0.15 ± 0.005 Bc | 0.004 ± 0.020 Aab | 0.09 ± 0.006 Aa | 0.05 ± 0.001 Aab | 0.04 ± 0.003 ABb | 0.10 ± 0.002 AB | 0.0003 ± 0.00008 Aa-d | |
| 72 | 0.03 ± 0.001 Aef | 0.22 ± 0.02 Ccde | 0.07 ± 0.004 Ccde | 0.20 ± 0.0081 Aa | 0.004 ± 0.020 ABb | 0.10 ± 0.005 Aa | 0.05 ± 0.001 Aabc | 0.05 ± 0.003 Aa | 0.08 ± 0.003 C | 0.0003 ± 0.0001 Ae-h |
| Response (%) | Ct | Initial | 20 °C | 15 °C | 10 °C | 5 °C | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| UVA3 | UVA6 | UVA3 | UVA6 | UVA3 | UVA6 | UVA3 | UVA6 | UVA3 | UVA6 | ||
| PAL | C1 | 110.80 aA* | 44.03 aAB | 149.21 aA* | −58.31 aC | 127.88 bA | 68.11 aA | 113.59 bA* | 8.80 aB | 108.22 bA* | −52.76 aC |
| C2 | 115.32 aB | 148.32 aA | 143.67 aA* | −19.47 aBC | 162.42 abA* | 102.56 aA | 114.33 bB* | 16.22 abB | 149.11 aA* | −60.84 aC | |
| C3 | 142.99 aB | 148.32 aA | 172.82 aAB* | −9.99 aAB | 193.34 aA* | 38.72 aA | 145.02 aAB* | −64.55 bBC | 139.68 aB* | −72.94 aC | |
| PPO | C1 | 6.40 aB | −10.21 aC | 469.98 aA | 112.67 aA | −0.16 cB | 63.97 bAB | 128.74 bB | −32.69 bC | −8.04 bB | 2.82 bBC |
| C2 | 57.81 aC* | 26.96 aB | 354.87 aA* | 138.75 aA | 24.66 bC | 49.94 bB | 130.30 aB | 69.74 aB | 56.94 aC | 29.24 bB | |
| C3 | 7.32 aB | 1.58 aC | 470.60 aA | 156.23 aA | 108.99 aB | 95.23 aB | 96.01 abB | 115.90 aAB | 53.42 aB | 92.54 aB | |
| TPC | C1 | 104.64 bA* | 21.89 bC | 94.19 bA* | 58.75 bA | 99.15 cA | 32.42 bBC | 118.86 bA | 45.87 aAB | 43.16 bB* | 19,680 aC |
| C2 | 76.10 bB* | −2.56 bC | 70.06 bBC* | 41.10 bA | 135.42 bA* | 20.97 bB | 93.49 aBA | −7.38 bC | 28.17 aC* | −6.36 aC | |
| C3 | 85.08 aB* | 12.46 aC | 99.84 aB | 76.38 aA | 174.37 aA* | 41.53 aB | 103.66 aB* | −17.36 bD | 21.28 aC* | −12.26 aD | |
| TPCHPLC | C1 | 19.74 bB | 66.76 aA | 50.97 bA | 21.67 a | 53.60 cA | 37.43 aA | 31.58 aAB | 22.84 aA | 46.65 cA | 35.35 aA |
| C2 | 33.64 abB | 33.98 aA | 44.69 abB* | 0.51 aA | 62.32 bA | 49.12 aA | 30.69 aB | −13.32 aA | 4.84 abC | −1.75 aA | |
| C3 | 80.77 aC | 72.59 aA | 67.05 aC* | 21.98 aBC* | 126.15 aD* | 57.36 aAB | 32.88 aC* | 5.53 aC | 12.93 aD | 7.94 aC | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villamil-Galindo, E.; Jacobo-Velázquez, D.A.; Piagentini, A.M. Combining Abiotic Stresses as a Low-Cost Strategy for Increasing the Phenolic Content in Apple Agro-Industrial By-Products. Antioxidants 2025, 14, 287. https://doi.org/10.3390/antiox14030287
Villamil-Galindo E, Jacobo-Velázquez DA, Piagentini AM. Combining Abiotic Stresses as a Low-Cost Strategy for Increasing the Phenolic Content in Apple Agro-Industrial By-Products. Antioxidants. 2025; 14(3):287. https://doi.org/10.3390/antiox14030287
Chicago/Turabian StyleVillamil-Galindo, Esteban, Daniel A. Jacobo-Velázquez, and Andrea Marcela Piagentini. 2025. "Combining Abiotic Stresses as a Low-Cost Strategy for Increasing the Phenolic Content in Apple Agro-Industrial By-Products" Antioxidants 14, no. 3: 287. https://doi.org/10.3390/antiox14030287
APA StyleVillamil-Galindo, E., Jacobo-Velázquez, D. A., & Piagentini, A. M. (2025). Combining Abiotic Stresses as a Low-Cost Strategy for Increasing the Phenolic Content in Apple Agro-Industrial By-Products. Antioxidants, 14(3), 287. https://doi.org/10.3390/antiox14030287