
Impact of Hydroxytyrosol-Rich Extract Supplementation in a High-Fat Diet on Gilthead Sea Bream (Sparus aurata) Lipid Metabolism

 , , , , ,
, , , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Trial
2.2. Somatic Parameters and Lipid Composition
2.3. Plasma Metabolites
2.4. Histology
2.5. Gene Expression
2.5.1. RNA Extraction and cDNA Synthesis
2.5.2. Real-Time Quantitative PCR (qPCR)
2.6. Protein Homogenates and Western Blotting
2.7. Lipid Peroxidation
2.8. Preadipocyte Primary Culture and Treatments
2.8.1. Cell Viability
2.8.2. Lipid Accumulation
2.9. Statistical Analysis
3. Results
3.1. Somatic Parameters, Lipid Composition and Plasma Metabolites
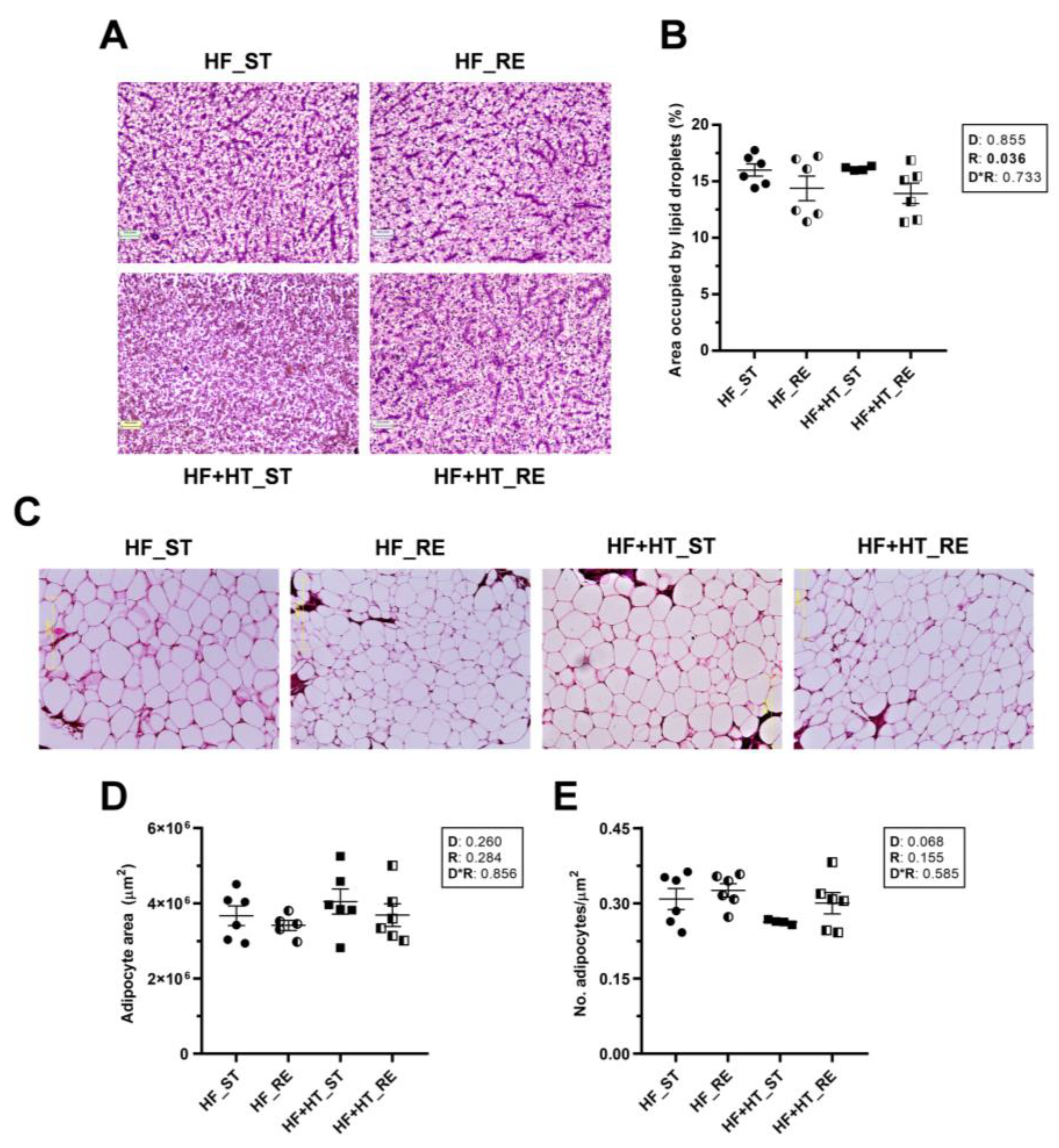
3.2. Histological Changes in Liver and Adipose Tissue
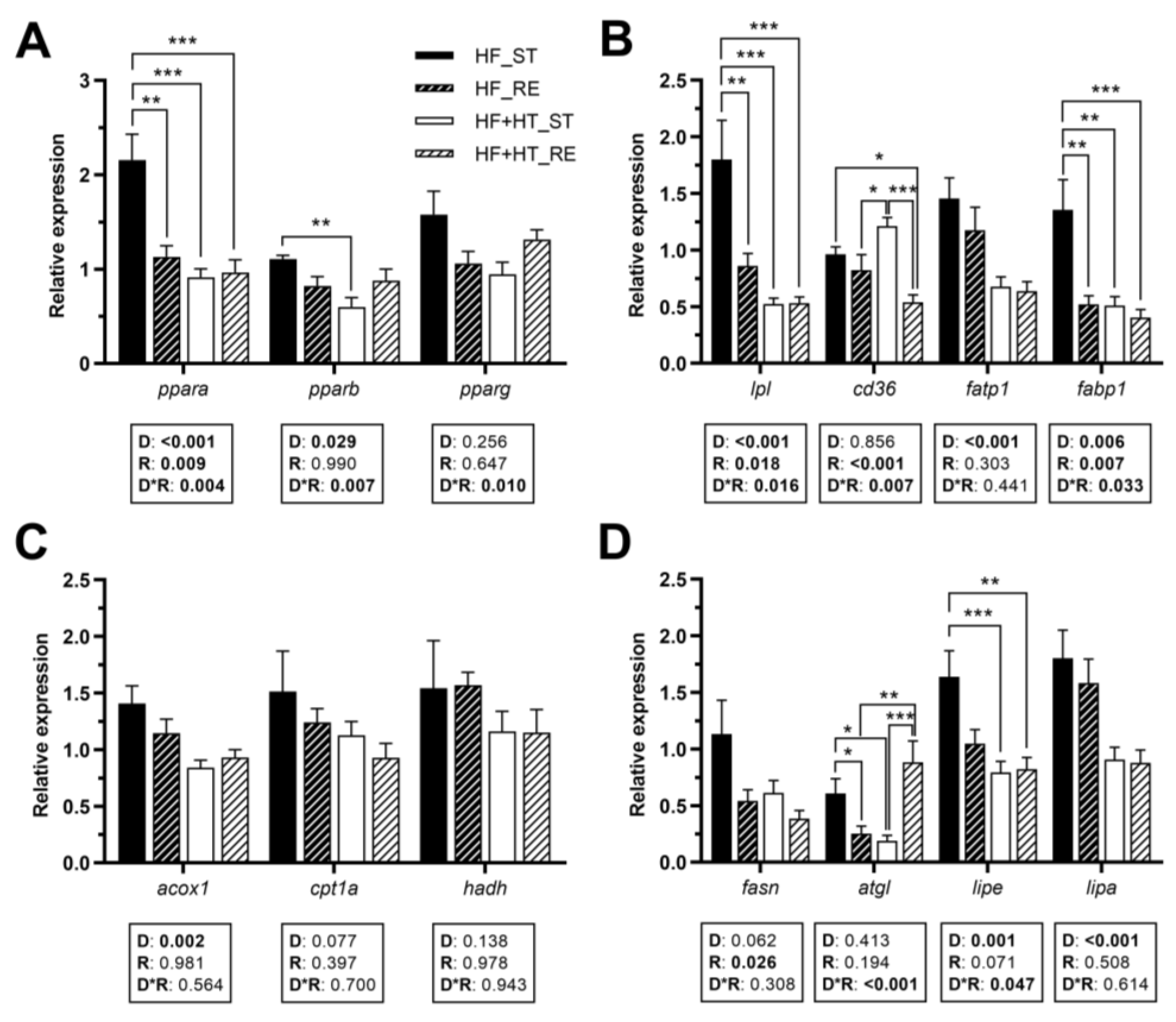
3.3. Lipid Metabolism in the Liver
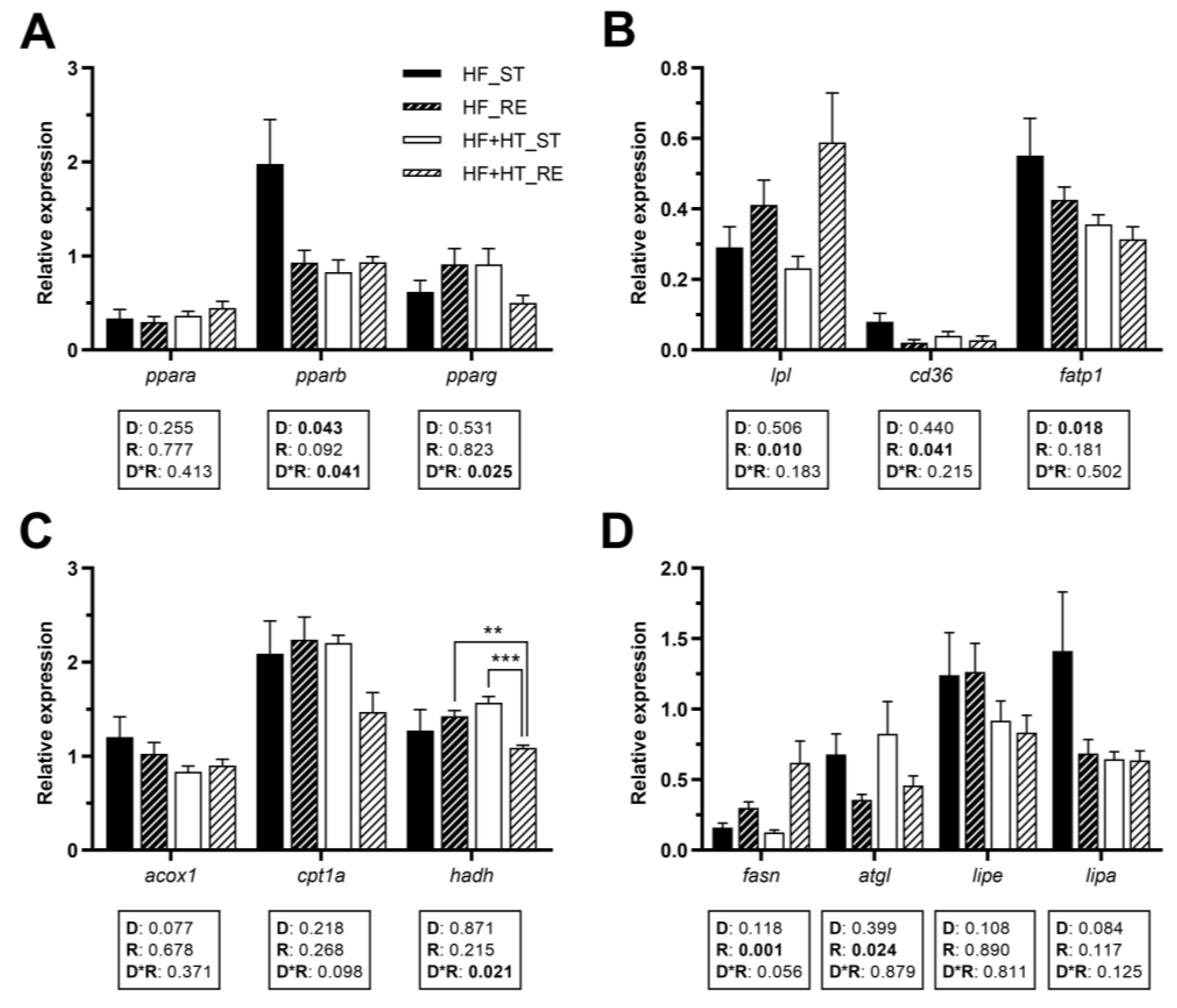
3.4. Lipid Metabolism in Adipose Tissue
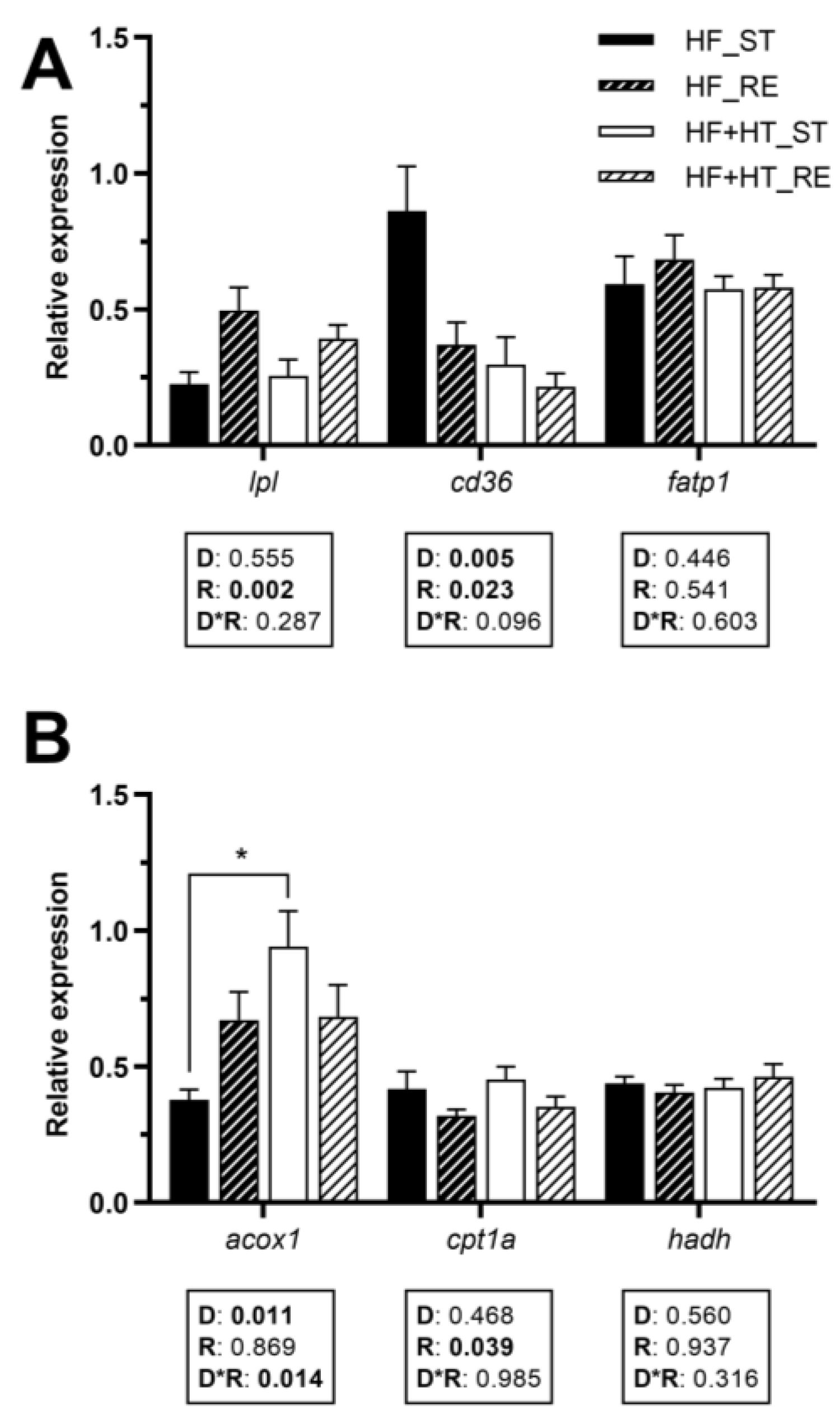
3.5. Lipid Metabolism in White Muscle
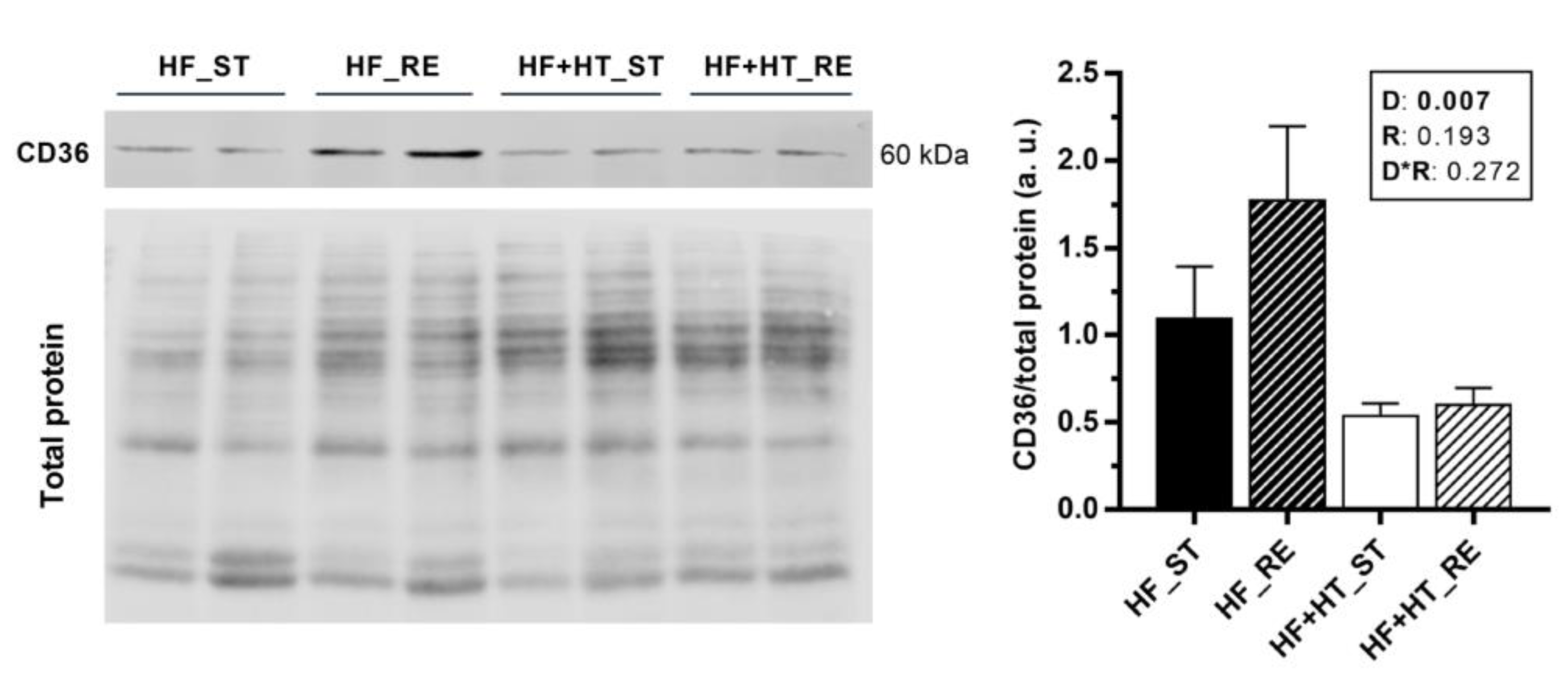
3.6. CD36 Protein Expression and Lipid Peroxidation in Liver and White Muscle
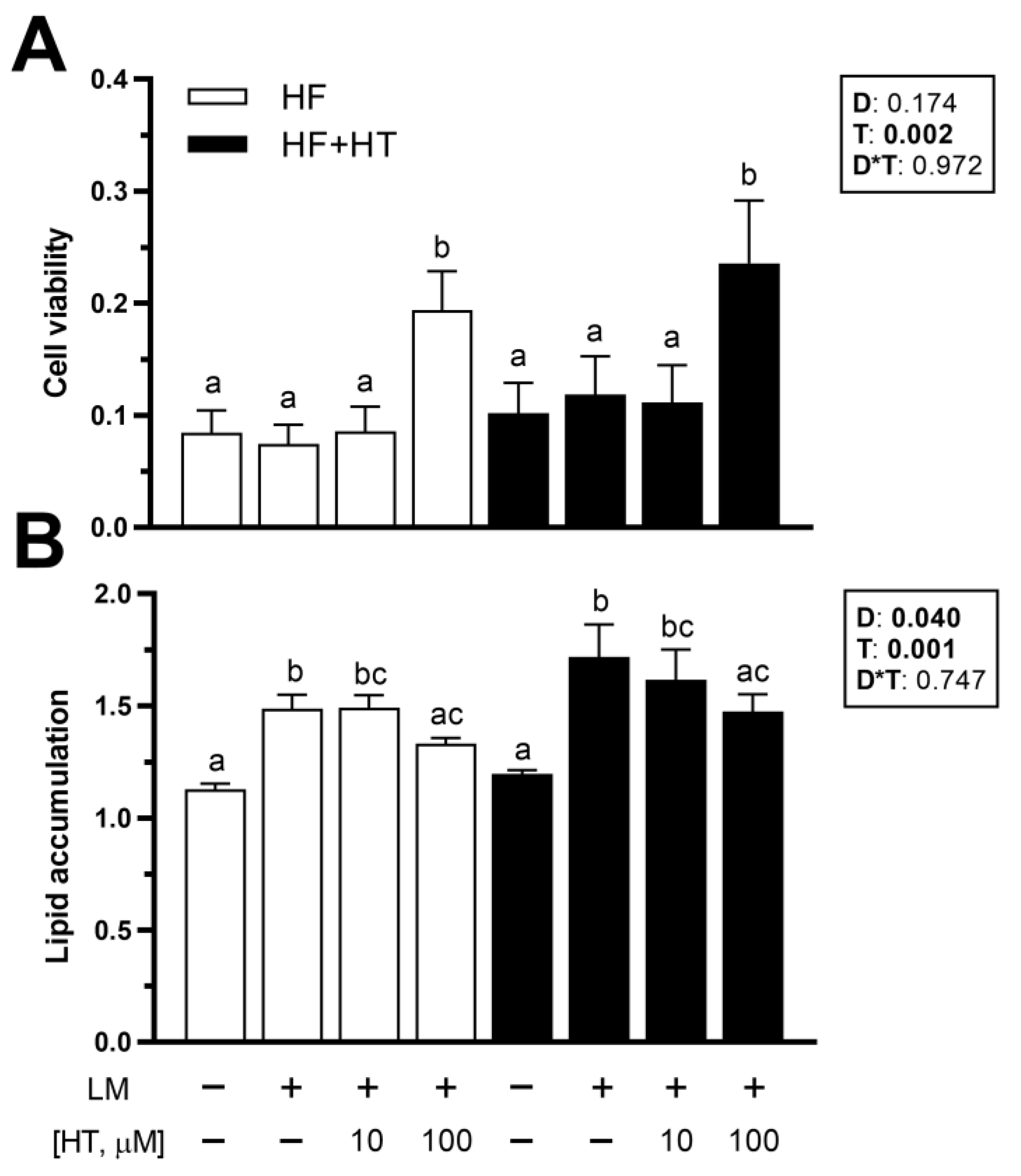
3.7. Viability and Lipid Accumulation in Primary Cultured Preadipocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turchini, G.M.; Francis, D.S.; Du, Z.Y.; Olsen, R.E.; Ringø, E.; Tocher, D.R. The lipids. In Fish Nutrition; Academic Press: Cambridge, MA, USA, 2022; Volume 3, pp. 303–467. ISBN 9780128195871. [Google Scholar]
- Sabzi, E.; Mohammadiazarm, H.; Salati, A.P. Effect of dietary L-carnitine and lipid levels on growth performance, blood biochemical parameters and antioxidant status in juvenile common carp (Cyprinus carpio). Aquaculture 2017, 480, 89–93. [Google Scholar] [CrossRef]
- Tang, T.; Hu, Y.; Peng, M.; Chu, W.; Hu, Y.; Zhong, L. Effects of high-fat diet on growth performance, lipid accumulation and lipid metabolism-related MicroRNA/gene expression in the liver of grass carp (Ctenopharyngodon idella). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 234, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Naiel, M.A.E.; Negm, S.S.; Ghazanfar, S.; Shukry, M.; Abdelnour, S.A. The risk assessment of high-fat diet in farmed fish and its mitigation approaches: A review. J. Anim. Physiol. Anim. Nutr. 2022, 107, 948–969. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.-Y.; Clouet, P.; Zheng, W.-H.; Degrace, P.; Tian, L.-X.; Liu, Y.-J. Biochemical hepatic alterations and body lipid composition in the herbivorous grass carp (Ctenopharyngodon idella) fed high-fat diets. Br. J. Nutr. 2006, 95, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Liu, Y.; Tian, L.; Wang, J.; Wang, Y.; Liang, G. Effect of dietary lipid level on growth, feed utilization and body composition by juvenile grass carp (Ctenopharyngodon idella). Aquac. Nutr. 2005, 11, 139–146. [Google Scholar] [CrossRef]
- Lu, K.-L.; Wang, L.-N.; Zhang, D.-D.; Liu, W.-B.; Xu, W.-N. Berberine attenuates oxidative stress and hepatocytes apoptosis via protecting mitochondria in blunt snout bream Megalobrama amblycephala fed high-fat diets. Fish Physiol. Biochem. 2017, 43, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Soengas, J.L.; Cerdá-Reverter, J.M.; Delgado, M.J. Central regulation of food intake in fish: An evolutionary perspective. J. Mol. Endocrinol. 2018, 60, R171–R199. [Google Scholar] [CrossRef] [PubMed]
- Karković Marković, A.; Torić, J.; Barbarić, M.; Jakobušić Brala, C. Hydroxytyrosol, tyrosol and derivatives and their potential effects on human health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef]
- Silva, A.F.R.; Resende, D.; Monteiro, M.; Coimbra, M.A.; Silva, A.M.S.; Cardoso, S.M. Application of hydroxytyrosol in the functional foods field: From ingredient to dietary supplements. Antioxidants 2020, 9, 1246. [Google Scholar] [CrossRef]
- Bertelli, M.; Karim, A.; Paolacci, S.; Manara, E.; Kurti, D.; Dhuli, K.; Bushati, V.; Miertus, J.; Pangallo, D.; Baglivo, M.; et al. Hydroxytyrosol: A natural compound with promising pharmacological activities. J. Biotechnol. 2020, 309, 29–33. [Google Scholar] [CrossRef]
- Fki, I.; Sayadi, S.; Mahmoudi, A.; Daoued, I.; Marrekchi, R.; Ghorbel, H. Comparative study on beneficial effects of hydroxytyrosol- and oleuropein-rich olive leaf extracts on high-fat diet-induced lipid metabolism disturbance and liver injury in rats. Biomed Res. Int. 2020, 2020, 1315202. [Google Scholar] [CrossRef] [PubMed]
- Illesca, P.; Valenzuela, R.; Espinosa, A.; Echeverría, F.; Soto-Alarcon, S.; Ortiz, M.; Videla, L.A. Hydroxytyrosol supplementation ameliorates the metabolic disturbances in white adipose tissue from mice fed a high-fat diet through recovery of transcription factors Nrf2, SREBP-1c, PPAR-γ and NF-κB. Biomed. Pharmacother. 2019, 109, 2472–2481. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, N.; Ma2, Y.; Wen, D. Hydroxytyrosol improves obesity and insulin resistance by modulating gut microbiota in high-fat diet-induced obese mice. Front. Microbiol. 2019, 10, 390. [Google Scholar] [CrossRef]
- Poudyal, H.; Campbell, F.; Brown, L. Olive leaf extract attenuates cardiac, hepatic, and metabolic changes in high carbohydrate-, high fat-fed rats. J. Nutr. 2010, 140, 946–953. [Google Scholar] [CrossRef]
- Valenzuela, R.; Echeverria, F.; Ortiz, M.; Rincón-Cervera, M.Á.; Espinosa, A.; Hernandez-Rodas, M.C.; Illesca, P.; Valenzuela, A.; Videla, L.A. Hydroxytyrosol prevents reduction in liver activity of Δ-5 and Δ-6 desaturases, oxidative stress, and depletion in long chain polyunsaturated fatty acid content in different tissues of high-fat diet fed mice. Lipids Health Dis. 2017, 16, 64. [Google Scholar] [CrossRef]
- Drira, R.; Chen, S.; Sakamoto, K. Oleuropein and hydroxytyrosol inhibit adipocyte differentiation in 3 T3-L1 cells. Life Sci. 2011, 89, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Stefanon, B.; Colitti, M. Hydroxytyrosol, an ingredient of olive oil, reduces triglyceride accumulation and promotes lipolysis in human primary visceral adipocytes during differentiation. Exp. Biol. Med. 2016, 241, 1796–1802. [Google Scholar] [CrossRef]
- Dong, Y.Z.; Li, L.; Espe, M.; Lu, K.L.; Rahimnejad, S. Hydroxytyrosol attenuates hepatic fat accumulation via activating mitochondrial biogenesis and autophagy through the AMPK pathway. J. Agric. Food Chem. 2020, 68, 9377–9386. [Google Scholar] [CrossRef]
- Dong, Y.; Xia, T.; Yu, M.; Wang, L.; Song, K.; Zhang, C.; Lu, K. Hydroxytyrosol attenuates high-fat-diet-induced oxidative stress, apoptosis and inflammation of blunt snout bream (Megalobrama amblycephala) through its regulation of mitochondrial homeostasis. Fishes 2022, 7, 78. [Google Scholar] [CrossRef]
- Lutfi, E.; Babin, P.J.; Gutiérrez, J.; Capilla, E.; Navarro, I. Caffeic acid and hydroxytyrosol have anti-obesogenic properties in zebrafish and rainbow trout models. PLoS ONE 2017, 12, e0178833. [Google Scholar] [CrossRef]
- Balbuena-Pecino, S.; Montblanch, M.; García-Meilán, I.; Fontanillas, R.; Gallardo, Á.; Gutiérrez, J.; Navarro, I.; Capilla, E. Hydroxytyrosol-rich extract from olive juice as an additive in gilthead sea bream juveniles fed a high-fat diet: Regulation of somatic growth. Front. Physiol. 2022, 13, 966175. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Balbuena-Pecino, S.; Riera-Heredia, N.; Vélez, E.J.; Gutiérrez, J.; Navarro, I.; Riera-Codina, M.; Capilla, E. Temperature affects musculoskeletal development and muscle lipid metabolism of gilthead sea bream (Sparus aurata). Front. Endocrinol. 2019, 10, 439265. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Salmerón, C.; Acerete, L.; Gutiérrez, J.; Navarro, I.; Capilla, E. Characterization and endocrine regulation of proliferation and differentiation of primary cultured preadipocytes from gilthead sea bream (Sparus aurata). Domest. Anim. Endocrinol. 2013, 45, 1–10. [Google Scholar] [CrossRef]
- Balbuena-Pecino, S.; Lutfi, E.; Riera-Heredia, N.; Gasch-Navalón, E.; Vélez, E.J.; Gutiérrez, J.; Capilla, E.; Navarro, I. Genistein induces adipogenic and autophagic effects in rainbow trout (Oncorhynchus mykiss) adipose tissue: In vitro and in vivo models. Int. J. Mol. Sci. 2020, 21, 5884. [Google Scholar] [CrossRef]
- Basto-Silva, C.; Balbuena-Pecino, S.; Oliva-Teles, A.; Riera-Heredia, N.; Navarro, I.; Guerreiro, I.; Capilla, E. Gilthead seabream (Sparus aurata) in vitro adipogenesis and its endocrine regulation by leptin, ghrelin, and insulin. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 249, 110772. [Google Scholar] [CrossRef]
- Li, L.Y.; Limbu, S.M.; Ma, Q.; Chen, L.Q.; Zhang, M.L.; Du, Z.Y. The metabolic regulation of dietary L-carnitine in aquaculture nutrition: Present status and future research strategies. Rev. Aquac. 2019, 11, 1228–1257. [Google Scholar] [CrossRef]
- Rueda-Jasso, R.; Conceição, L.E.C.; Dias, J.; De Coen, W.; Gomes, E.; Rees, J.F.; Soares, F.; Dinis, M.T.; Sorgeloos, P. Effect of dietary non-protein energy levels on condition and oxidative status of Senegalese sole (Solea senegalensis) juveniles. Aquaculture 2004, 231, 417–433. [Google Scholar] [CrossRef]
- Weil, C.; Lefèvre, F.; Bugeon, J. Characteristics and metabolism of different adipose tissues in fish. Rev. Fish Biol. Fish. 2013, 23, 157–173. [Google Scholar] [CrossRef]
- Sheridan, M.A. Lipid dynamics in fish: Aspects of absorption, transportation, deposition and mobilization. Comp. Biochem. Physiol. Part B Biochem. 1988, 90, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, M.A. Regulation of lipid metabolism in poikilothermic vertebrates. Comp. Biochem. Physiol. Part B Biochem. 1994, 107, 495–508. [Google Scholar] [CrossRef]
- Cao, K.; Xu, J.; Zou, X.; Li, Y.; Chen, C.; Zheng, A.; Li, H.; Li, H.; Szeto, I.M.Y.; Shi, Y.; et al. Hydroxytyrosol prevents diet-induced metabolic syndrome and attenuates mitochondrial abnormalities in obese mice. Free Radic. Biol. Med. 2014, 67, 396–407. [Google Scholar] [CrossRef]
- Dong, Y.; Yu, M.; Wu, Y.; Xia, T.; Wang, L.; Song, K.; Zhang, C.; Lu, K.; Rahimnejad, S. Hydroxytyrosol Promotes the mitochondrial function through activating mitophagy. Antioxidants 2022, 11, 893. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Andree, K.B.; Sanahuja, I.; Holhorea, P.G.; Calduch-Giner, J.; Morais, S.; Pastor, J.J.; Pérez-Sánchez, J.; Gisbert, E. Bile salt dietary supplementation promotes growth and reduces body adiposity in gilthead seabream (Sparus aurata). Aquaculture 2023, 566, 739203. [Google Scholar] [CrossRef]
- Yilmaz, E.; Naz, M.; Akyurt, I. Effect of dietary olive pomace oil and L-carnitine on growth and chemical compostion of African catfish, Clarias gariepinus (Burchell, 1822). Isr. J. Aquac.-Bamidgeh 2004, 56, 14–21. [Google Scholar] [CrossRef]
- Grigorakis, K. Compositional and organoleptic quality of farmed and wild gilthead sea bream (Sparus aurata) and sea bass (Dicentrarchus labrax) and factors affecting it: A review. Aquaculture 2007, 272, 55–75. [Google Scholar] [CrossRef]
- Henderson, G.C. Plasma free fatty acid concentration as a modifiable risk factor for metabolic disease. Nutrients 2021, 13, 2590. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Baba, E.; Acar, Ü.; Yılmaz, S.; Zemheri, F.; Ergün, S. Dietary olive leaf (Olea europea L.) extract alters some immune gene expression levels and disease resistance to Yersinia ruckeri infection in rainbow trout Oncorhynchus mykiss. Fish Shellfish Immunol. 2018, 79, 28–33. [Google Scholar] [CrossRef]
- Lu, K.L.; Xu, W.N.; Wang, L.N.; Zhang, D.D.; Zhang, C.N.; Liu, W. Bin Hepatic β-oxidation and regulation of carnitine palmitoyltransferase (CPT) I in blunt snout bream Megalobrama amblycephala fed a high fat diet. PLoS ONE 2014, 9, e93135. [Google Scholar] [CrossRef]
- Lu, K.L.; Xu, W.N.; Li, X.F.; Liu, W.B.; Wang, L.N.; Zhang, C.N. Hepatic triacylglycerol secretion, lipid transport and tissue lipid uptake in blunt snout bream (Megalobrama amblycephala) fed high-fat diet. Aquaculture 2013, 408–409, 160–168. [Google Scholar] [CrossRef]
- Newberry, E.P.; Xie, Y.; Kennedy, S.M.; Luo, J.; Davidson, N.O. Protection against Western diet-induced obesity and hepatic steatosis in liver fatty acid-binding protein knockout mice. Hepatology 2006, 44, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Doege, H.; Grimm, D.; Falcon, A.; Tsang, B.; Storm, T.A.; Xu, H.; Ortegon, A.M.; Kazantzis, M.; Kay, M.A.; Stahl, A. Silencing of hepatic fatty acid transporter protein 5 in vivo reverses diet-induced non-alcoholic fatty liver disease and improves hyperglycemia. J. Biol. Chem. 2008, 283, 22186–22192. [Google Scholar] [CrossRef]
- Falcon, A.; Doege, H.; Fluitt, A.; Tsang, B.; Watson, N.; Kay, M.A.; Stahl, A. FATP2 is a hepatic fatty acid transporter and peroxisomal very long-chain acyl-CoA synthetase. Am. J. Physiol.-Endocrinol. Metab. 2010, 299, 384–393. [Google Scholar] [CrossRef]
- Sánchez-Gurmaches, J.; Østbye, T.K.; Navarro, I.; Torgersen, J.; Hevrøy, E.M.; Ruyter, B.; Torstensen, B.E. In vivo and in vitro insulin and fasting control of the transmembrane fatty acid transport proteins in Atlantic salmon (Salmo salar). Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 301, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Liu, W.; Gao, W.; Wu, F.; Yu, L.; Lu, X.; Yang, C.G.; Jiang, M.; Wen, H. Molecular cloning and gene/protein expression of FAT/CD36 from grass carp (Ctenopharyngodon idella) and the regulation of its expression by dietary energy. Fish Physiol. Biochem. 2017, 43, 875–888. [Google Scholar] [CrossRef]
- Glatz, J.F.C.; Nabben, M.; Heather, L.C.; Bonen, A.; Luiken, J.J.F.P. Regulation of the subcellular trafficking of CD36, a major determinant of cardiac fatty acid utilization. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1461–1471. [Google Scholar] [CrossRef]
- Garbacz, W.G.; Lu, P.; Miller, T.M.; Poloyac, S.M.; Eyre, N.S.; Mayrhofer, G.; Xu, M.; Ren, S.; Xie, W. Hepatic overexpression of CD36 improves glycogen homeostasis and attenuates high-fat diet-induced hepatic steatosis and insulin resistance. Mol. Cell. Biol. 2016, 36, 2715–2727. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M. PPAR-gamma: Adipogenic regulator and thiazolidinedione receptor. Diabetes 1998, 47, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Southgate, R.J.; Holmes, A.G.; Febbraio, M.A. Suppression of plasma free fatty acids upregulates peroxisome proliferator-activated receptor (PPAR) α and δ and PPAR coactivator 1α in human skeletal muscle, but not lipid regulatory genes. J. Mol. Endocrinol. 2004, 33, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Costello, J.; Godinho, L.F.; Islinger, M. Peroxisome-mitochondria interplay and disease. J. Inherit. Metab. Dis. 2015, 38, 681–702. [Google Scholar] [CrossRef]
- Sánchez-Gurmaches, J.; Cruz-Garcia, L.; Gutiérrez, J.; Navarro, I. Adiponectin effects and gene expression in rainbow trout: An in vivo and in vitro approach. J. Exp. Biol. 2012, 215, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- Torstensen, B.E.; Nanton, D.A.; Olsvik, P.A.; Sundvold, H.; Stubhaug, I. Gene expression of fatty acid-binding proteins, fatty acid transport proteins (cd36 and FATP) and β-oxidation-related genes in Atlantic salmon (Salmo salar L.) fed fish oil or vegetable oil. Aquac. Nutr. 2009, 15, 440–451. [Google Scholar] [CrossRef]
- Hamilton, J.A. New insights into the roles of proteins and lipids in membrane transport of fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Chaiyapechara, S.; Casten, M.T.; Hardy, R.W.; Dong, F.M. Fish performance, fillet characteristics, and health assessment index of rainbow trout (Oncorhynchus mykiss) fed diets containing adequate and high concentrations of lipid and vitamin E. Aquaculture 2003, 219, 715–738. [Google Scholar] [CrossRef]
- Dias, J.; Corraze, G.; Arzel, J.; Alvarez, M.; Bautista, J.; Lopez-Bote, C.; Kaushik, S. Effets du rapport protéine/énergie des régimes alimentaires chez la truite et le bar en élévage. Perspectives de contrôle nutritionnel des dépôts lipidiques. Cybium 1999, 23 (Suppl. S1), 127–137. [Google Scholar]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Wang, B.; Van Veldhoven, P.P.; Brees, C.; Rubio, N.; Nordgren, M.; Apanasets, O.; Kunze, M.; Baes, M.; Agostinis, P.; Fransen, M. Mitochondria are targets for peroxisome-derived oxidative stress in cultured mammalian cells. Free Radic. Biol. Med. 2013, 65, 882–894. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Zheng, A.; Yang, L.; Liu, J.; Chen, C.; Tang, Y.; Zou, X.; Li, Y.; Long, J.; et al. Mitochondrial dysfunction-associated OPA1 cleavage contributes to muscle degeneration: Preventative effect of hydroxytyrosol acetate. Cell Death Dis. 2014, 5, e1521. [Google Scholar] [CrossRef]
- Du, Z.Y.; Clouet, P.; Huang, L.M.; Degrace, P.; Zheng, W.H.; He, J.G.; Tian, L.X.; Liu, Y.J. Utilization of different dietary lipid sources at high level in herbivorous grass carp (Ctenopharyngodon idella): Mechanism related to hepatic fatty acid oxidation. Aquac. Nutr. 2008, 14, 77–92. [Google Scholar] [CrossRef]
- Lu, K.L.; Xu, W.N.; Liu, W.B.; Wang, L.N.; Zhang, C.N.; Li, X.F. Association of mitochondrial dysfunction with oxidative stress and immune suppression in blunt snout bream Megalobrama amblycephala fed a high-fat diet. J. Aquat. Anim. Health 2014, 26, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Lutfi, E.; Riera-Heredia, N.; Córdoba, M.; Porte, C.; Gutiérrez, J.; Capilla, E.; Navarro, I. Tributyltin and triphenyltin exposure promotes in vitro adipogenic differentiation but alters the adipocyte phenotype in rainbow trout. Aquat. Toxicol. 2017, 188, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Tutino, V.; Orlando, A.; Russo, F.; Notarnicola, M. Hydroxytyrosol inhibits cannabinoid CB1 receptor gene expression in 3T3-L1 preadipocyte cell line. J. Cell. Physiol. 2016, 231, 483–489. [Google Scholar] [CrossRef]
- Warnke, I.; Goralczyk, R.; Fuhrer, E.; Schwager, J. Dietary constituents reduce lipid accumulation in murine C3H10 T1/2 adipocytes: A novel fluorescent method to quantify fat droplets. Nutr. Metab. 2011, 8, 30. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HF | HF+HT | |
|---|---|---|
| Ingredients (%) | ||
| Corn gluten | 3.80 | 3.80 |
| Wheat gluten | 20.00 | 20.00 |
| Fava beans | 8.00 | 8.00 |
| Soya concentrate | 25.00 | 25.00 |
| Fish oil | 9.98 | 9.98 |
| Fish meal | 15.00 | 15.00 |
| Rapeseed oil | 10.14 | 10.14 |
| Yttrium premix | 0.10 | 0.10 |
| Phosphate | 1.04 | 1.04 |
| Vitamin mineral premix | 0.44 | 0.44 |
| Wheat | 6.50 | 4.85 |
| HIDROX® | 0.00 | 1.66 |
| Composition (%) | ||
| Dry matter | 93.0 | 93.0 |
| Moisture | 7.0 | 7.0 |
| Crude protein | 46.8 | 46.7 |
| Crude fat | 24.0 | 24.2 |
| Ash | 5.4 | 5.6 |
| Crude fiber | 1.9 | 1.8 |
| Starch | 8.8 | 7.8 |
| HF_ST | HF_RE | HF+HT_ST | HF+HT_RE | D | R | D*R | |
|---|---|---|---|---|---|---|---|
| MFI | 0.86 ± 0.10 | 0.68 ± 0.07 | 1.01 ± 0.16 | 0.72 ± 0.13 | 0.435 | 0.086 | 0.654 |
| Liver lipid (%) | 17.08 ± 1.54 | 16.64 ± 0.66 | 13.57 ± 1.51 | 16.73 ± 1.23 | 0.202 | 0.307 | 0.178 |
| WM lipid (%) | 6.48 ± 0.52 a | 7.89 ± 0.91 ab | 10.26 ± 1.14 b | 7.01 ± 0.90 ab | 0.115 | 0.311 | 0.013 |
| HF_ST | HF_RE | HF+HT_ST | HF+HT_RE | D | R | D*R | |
|---|---|---|---|---|---|---|---|
| Glucose (mg/dL) | 96.78 ± 3.09 | 91.99 ± 3.08 | 97.41 ± 3.04 | 97.54 ± 1.68 | 0.269 | 0.404 | 0.378 |
| FFA (mg/dL) | 6.42 ± 0.47 a | 4.45 ± 0.42 b | 4.28 ± 0.23 b | 4.98 ± 0.38 b | 0.047 | 0.109 | 0.002 |
| HDL (mg/dL) | 289.81 ± 13.48 | 224.10 ± 16.33 | 264.41 ± 7.27 | 237.63 ± 11.74 | 0.633 | <0.001 | 0.124 |
| LDL/VLDL (mg/dL) | 81.33 ± 13.98 | 48.93 ± 5.44 | 87.46 ± 7.78 | 59.08 ± 5.10 | 0.390 | 0.003 | 0.831 |
| TG (mg/dL) | 335.06 ± 21.06 | 276.00 ± 14.09 | 325.17 ± 28.63 | 293.17 ± 18.12 | 0.871 | 0.049 | 0.546 |
| HF_ST | HF_RE | HF+HT_ST | HF+HT_RE | D | R | D*R | |
|---|---|---|---|---|---|---|---|
| Liver LPO | 0.994 ± 0.09 | 1.039 ± 0.17 | 0.927 ± 0.12 | 0.814 ± 0.10 | 0.230 | 0.779 | 0.513 |
| WM LPO | 0.050 ± 0.005 | 0.039 ± 0.005 | 0.061 ± 0.008 | 0.068 ± 0.007 | 0.004 | 0.786 | 0.164 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balbuena-Pecino, S.; Montblanch, M.; Rosell-Moll, E.; González-Fernández, V.; García-Meilán, I.; Fontanillas, R.; Gallardo, Á.; Gutiérrez, J.; Capilla, E.; Navarro, I. Impact of Hydroxytyrosol-Rich Extract Supplementation in a High-Fat Diet on Gilthead Sea Bream (Sparus aurata) Lipid Metabolism. Antioxidants 2024, 13, 403. https://doi.org/10.3390/antiox13040403
Balbuena-Pecino S, Montblanch M, Rosell-Moll E, González-Fernández V, García-Meilán I, Fontanillas R, Gallardo Á, Gutiérrez J, Capilla E, Navarro I. Impact of Hydroxytyrosol-Rich Extract Supplementation in a High-Fat Diet on Gilthead Sea Bream (Sparus aurata) Lipid Metabolism. Antioxidants. 2024; 13(4):403. https://doi.org/10.3390/antiox13040403
Chicago/Turabian StyleBalbuena-Pecino, Sara, Manel Montblanch, Enrique Rosell-Moll, Verónica González-Fernández, Irene García-Meilán, Ramon Fontanillas, Ángeles Gallardo, Joaquim Gutiérrez, Encarnación Capilla, and Isabel Navarro. 2024. "Impact of Hydroxytyrosol-Rich Extract Supplementation in a High-Fat Diet on Gilthead Sea Bream (Sparus aurata) Lipid Metabolism" Antioxidants 13, no. 4: 403. https://doi.org/10.3390/antiox13040403
APA StyleBalbuena-Pecino, S., Montblanch, M., Rosell-Moll, E., González-Fernández, V., García-Meilán, I., Fontanillas, R., Gallardo, Á., Gutiérrez, J., Capilla, E., & Navarro, I. (2024). Impact of Hydroxytyrosol-Rich Extract Supplementation in a High-Fat Diet on Gilthead Sea Bream (Sparus aurata) Lipid Metabolism. Antioxidants, 13(4), 403. https://doi.org/10.3390/antiox13040403