Protective Mechanism of Rosa roxburghii Tratt Fermentation Broth against Ultraviolet-A-Induced Photoaging of Human Embryonic Skin Fibroblasts

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. RRT Fermentation
2.3. Measurement of Total Sugar, Reducing Sugar, Flavonoids, Polyphenols, and Protein Content
2.4. In Vitro Antioxidant Activity Analysis
2.5. Cell Culture
2.6. Sample Preparation for Cell Experiments
2.7. Assay of Cell Viability
2.8. ROS Content Detection
2.9. Enzyme Activity Assay
2.10. Total Antioxidant Capacity and Lipid Peroxidation Level
2.11. Enzyme-Linked Immunosorbent Assay (ELISA)
2.12. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.13. RNA Sequencing (RNA-seq)
2.14. Data Analysis
3. Results
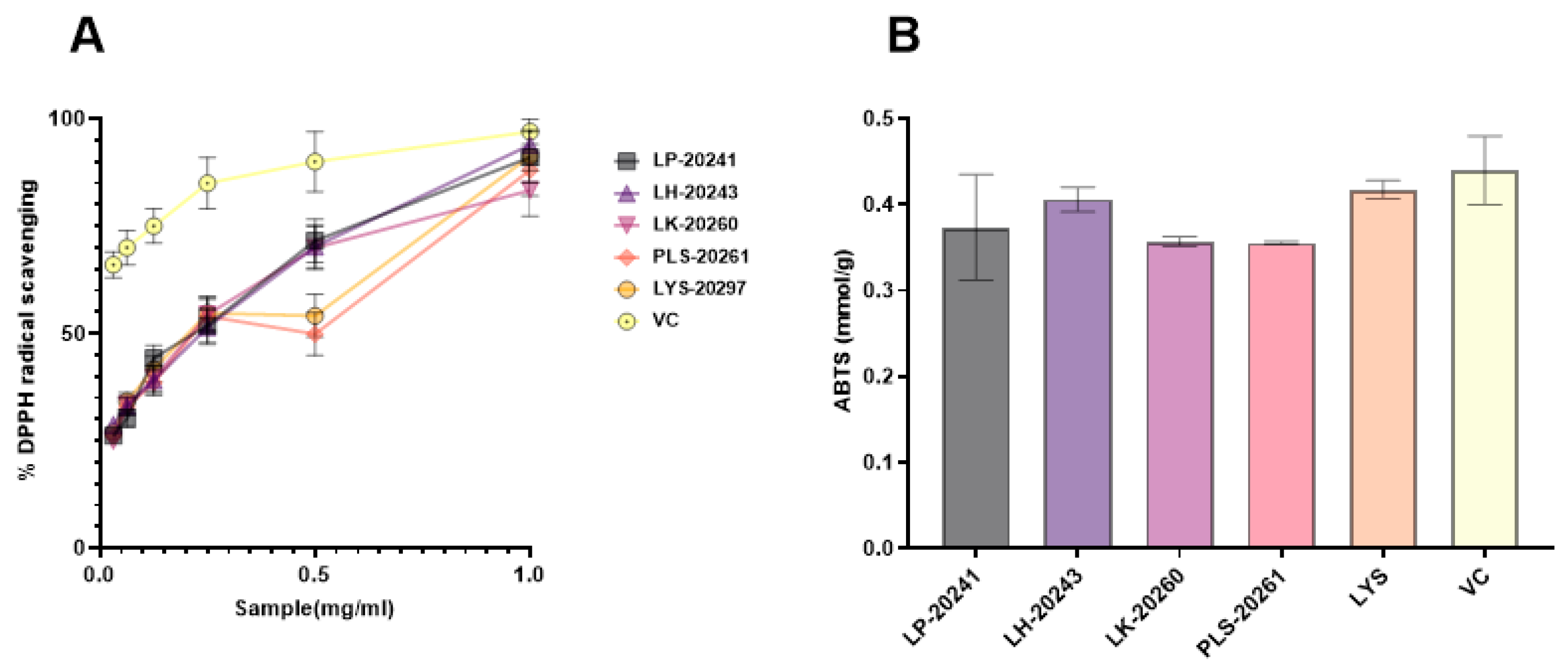
3.1. Active Substance Content
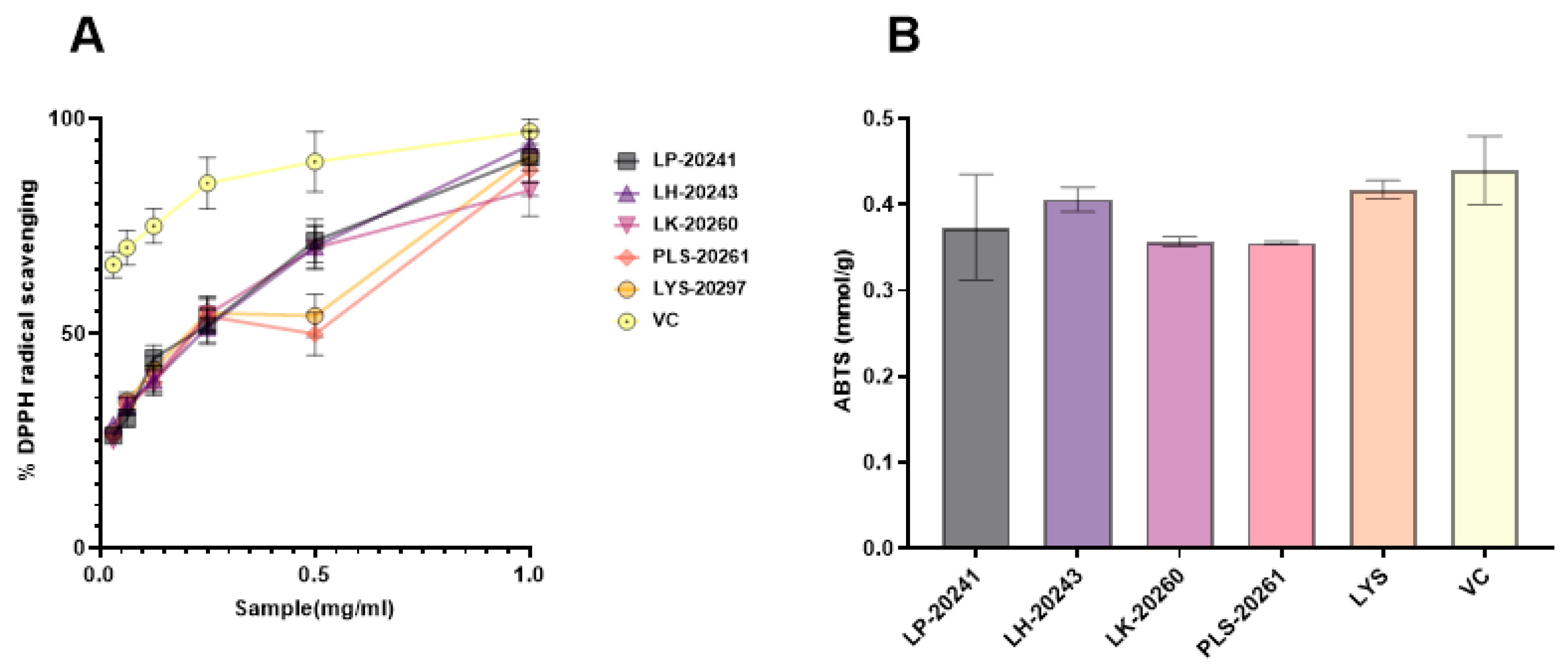
3.2. Antioxidant Efficacy In Vitro
3.3. Cell Viability and UVA Model Establishment
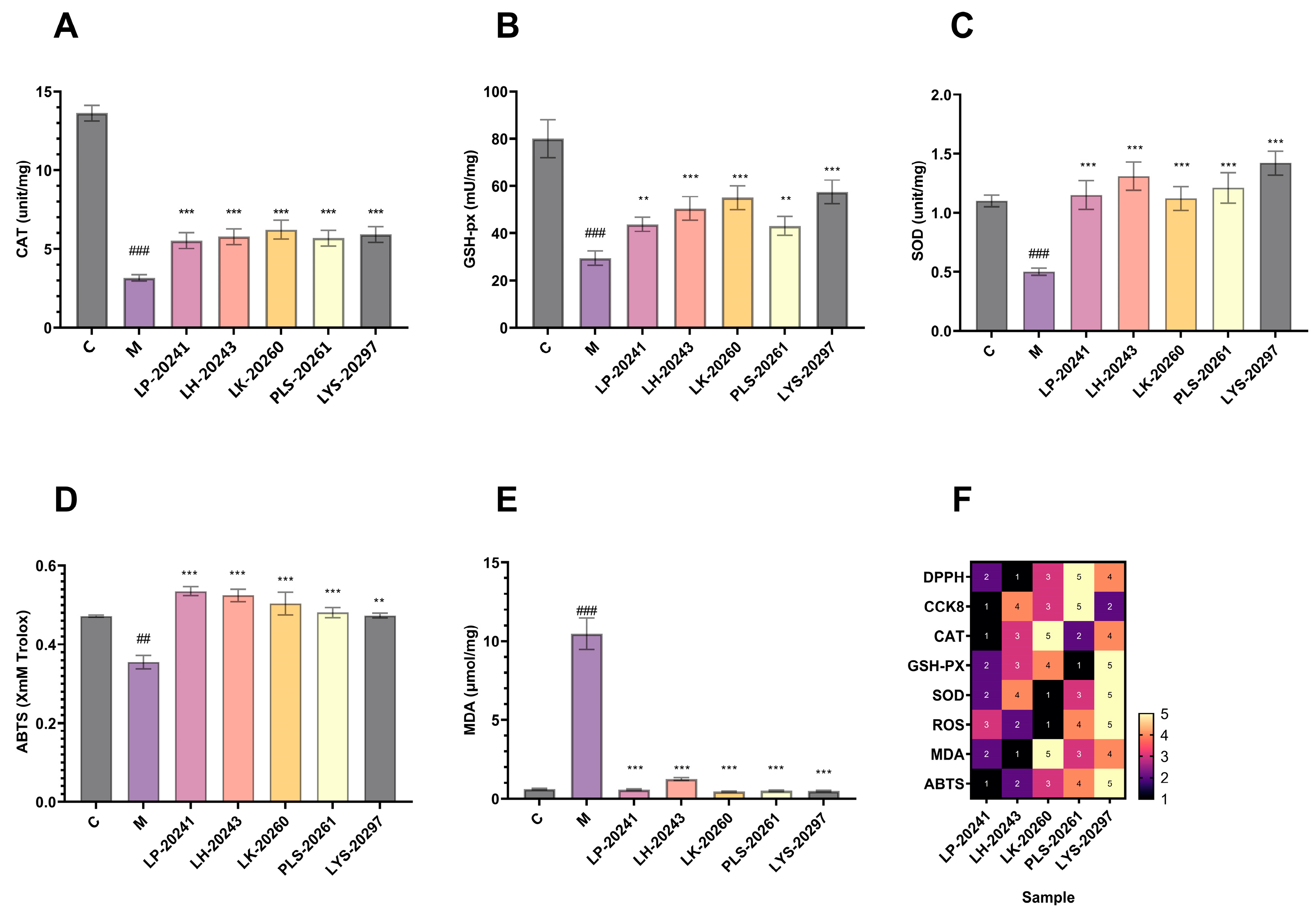
3.4. Intracellular Antioxidant Levels
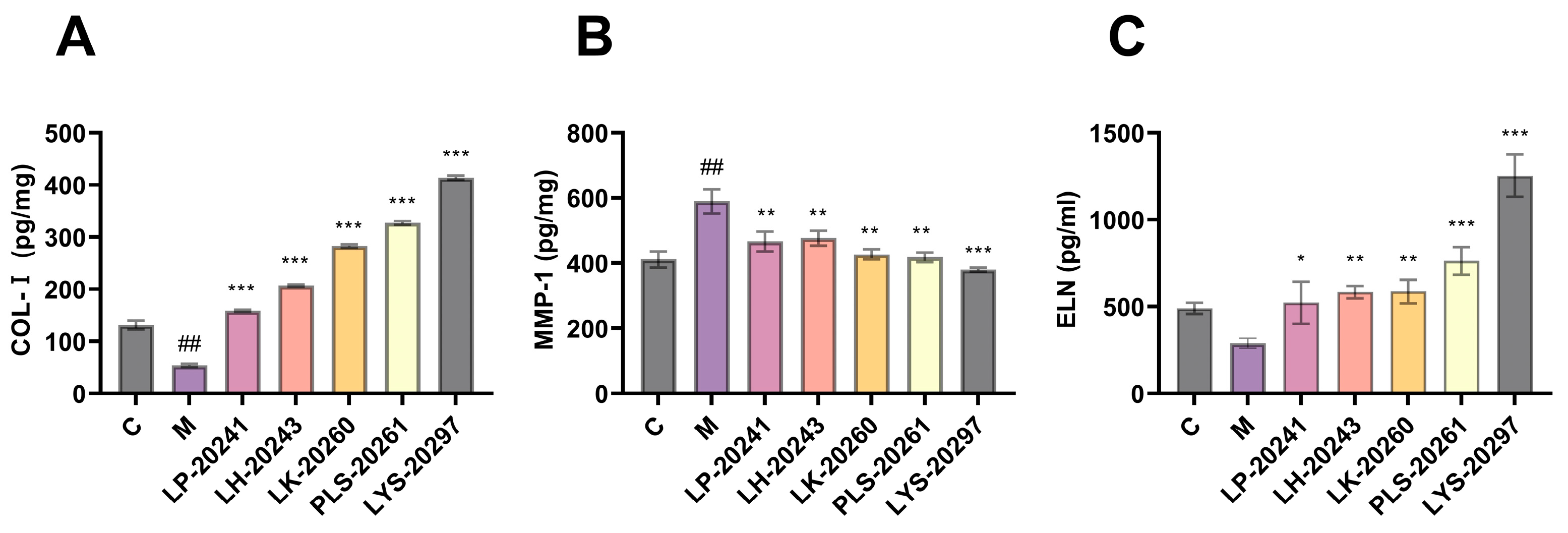
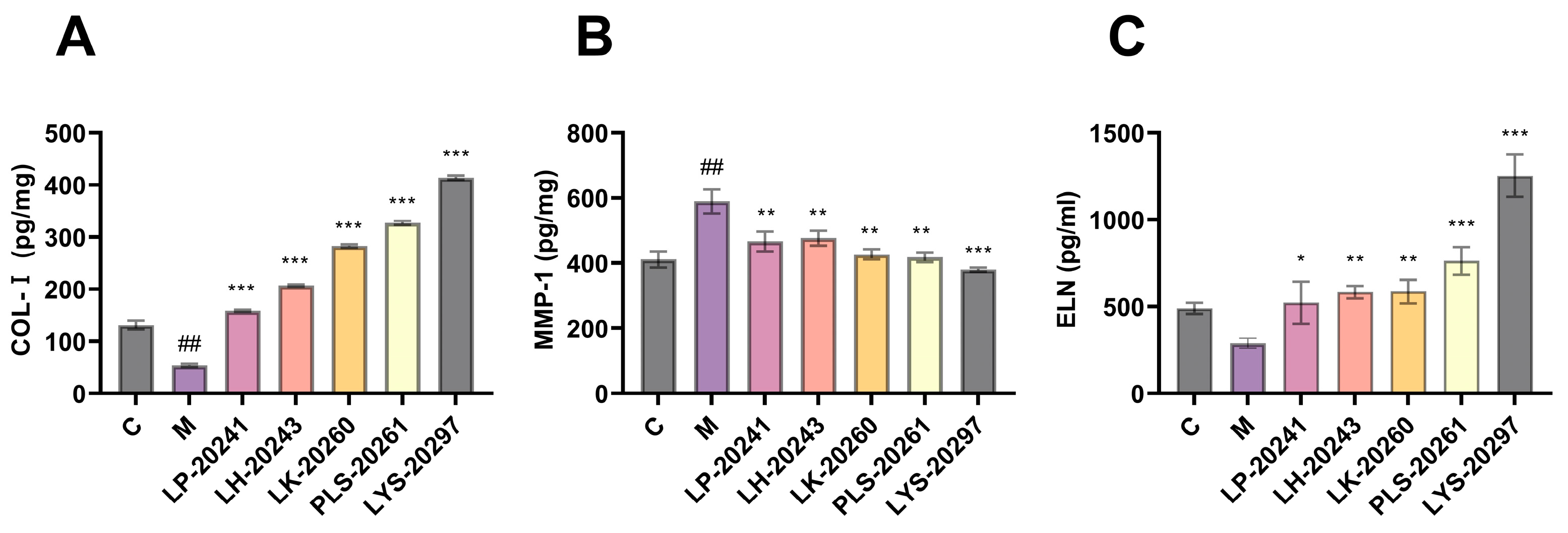
3.5. Detection of Photoaging-Related Protein Content
3.6. The Expression Levels of Collagen Synthesis- (TGF-β) and Degradation (MMP)-Related Genes Were Detected
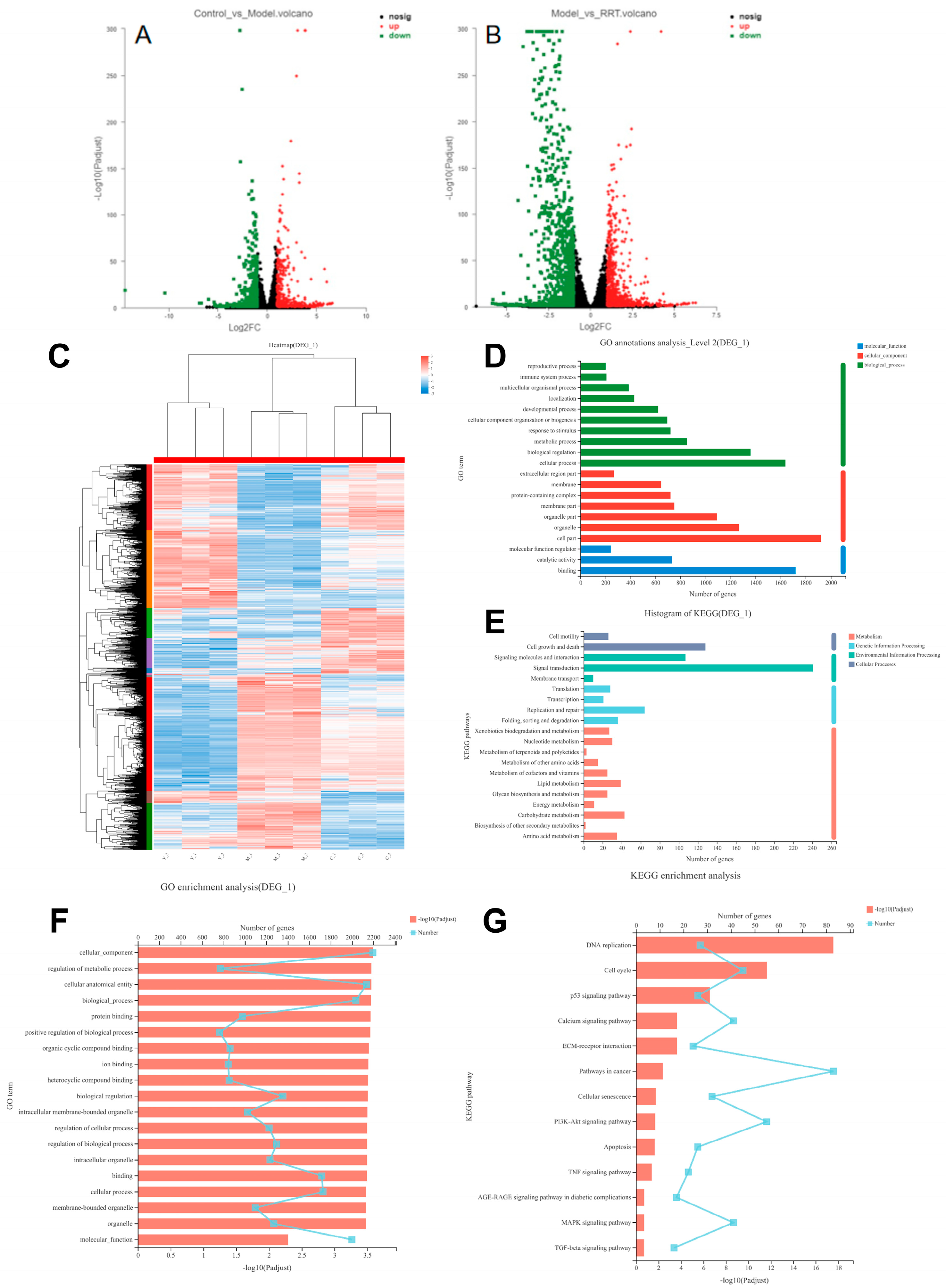
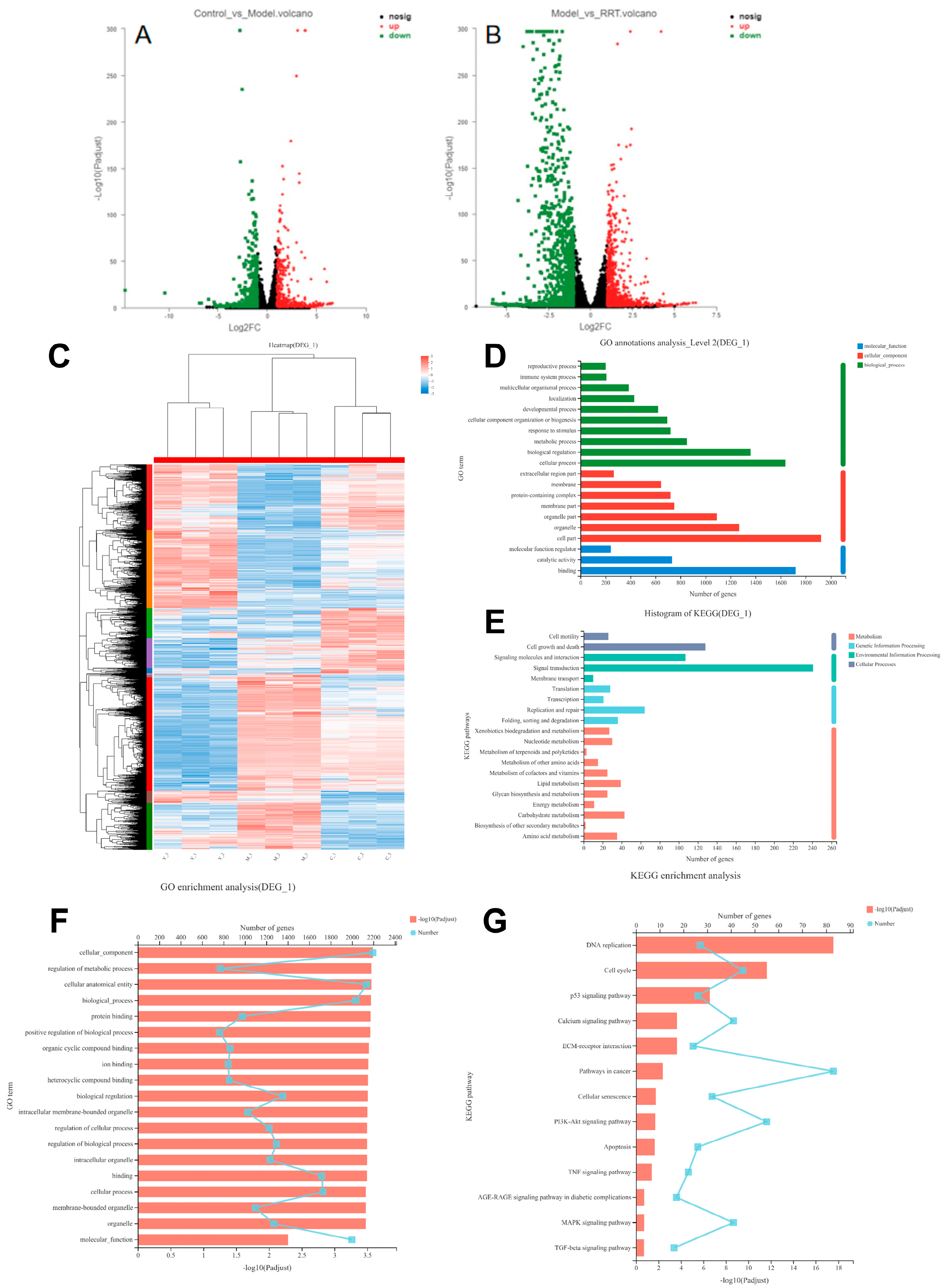
3.7. Transcriptome Sequence Analyses
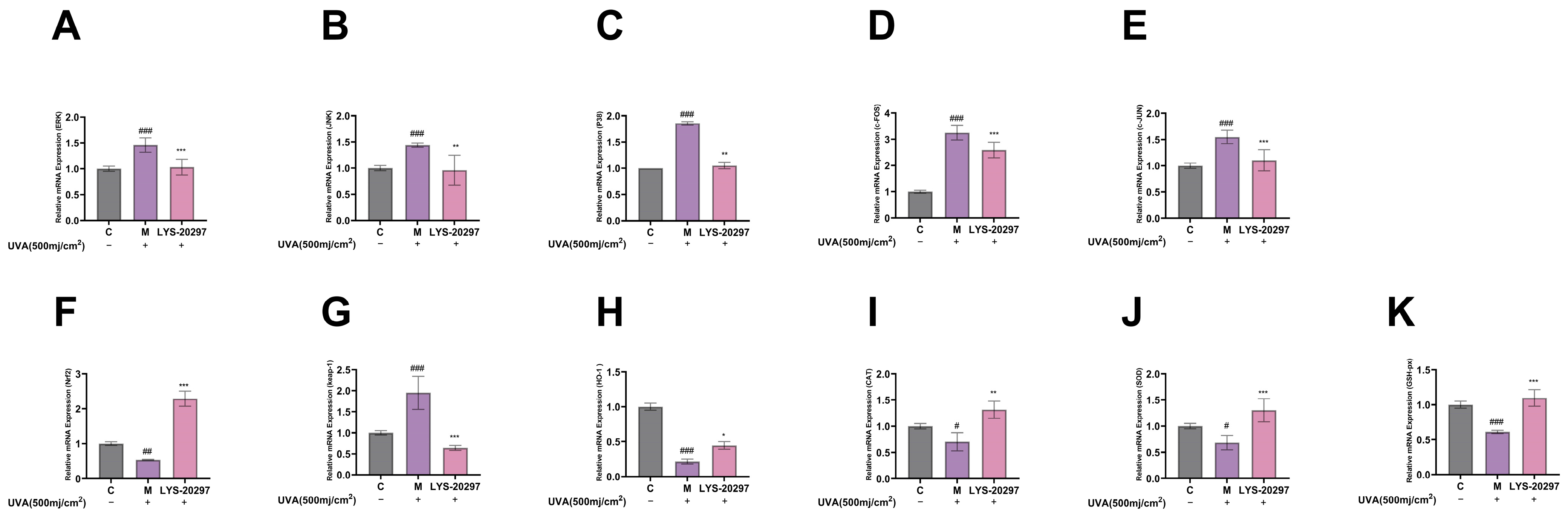
3.8. The Expression Levels of Oxidative Stress- (MAPK/AP-1) and Antioxidant Enzyme (Nrf2/Keap-1)-Related Genes Were Detected
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, L.; Wei, T.; Zheng, L.; Jiang, F.; Ma, W.; Lu, M.; Wu, X.; An, H. Recent Advances on Main Active Ingredients, Pharmacological Activities of Rosa Roxbughii and Its Development and Utilization. Foods 2023, 12, 1051. [Google Scholar] [CrossRef]
- Li, H.; Fang, W.; Wang, Z.; Chen, Y. Physicochemical, Biological Properties, and Flavour Profile of Rosa Roxburghii Tratt, Pyracantha Fortuneana, and Rosa Laevigata Michx Fruits: A Comprehensive Review. Food Chem. 2022, 366, 130509. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, G.; Wang, X.; Qin, L.; Xu, C.; She, X.; He, Y.; Tan, D. Chemical Constituents and Bioactivities of Rosa Roxburghii: A Systematic Review. Food Sci. Technol. 2022, 42, e72722. [Google Scholar] [CrossRef]
- Wang, L.-T.; Lv, M.-J.; An, J.-Y.; Fan, X.-H.; Dong, M.-Z.; Zhang, S.-D.; Wang, J.-D.; Wang, Y.-Q.; Cai, Z.-H.; Fu, Y.-J. Botanical Characteristics, Phytochemistry and Related Biological Activities of Rosa Roxburghii Tratt Fruit, and Its Potential Use in Functional Foods: A Review. Food Funct. 2021, 12, 1432–1451. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jiang, L.; Liu, Y.; Zou, S.; Lu, M.; An, H. Metabolomics Combined with Transcriptomics Analysis Revealed the Amino Acids, Phenolic Acids, and Flavonol Derivatives Biosynthesis Network in Developing Rosa Roxburghii Fruit. Foods 2022, 11, 1639. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhang, B.; Fu, X.; Huang, Q.; Li, C.; Liu, G.; Hai Liu, R. Recent Advances in Polysaccharides from Rose Roxburghii Tratt Fruits: Isolation, Structural Characterization, and Bioactivities. Food Funct. 2022, 13, 12561–12571. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, C.; Huang, Q.; Fu, X. Polysaccharide from Rosa Roxburghii Tratt Fruit Attenuates Hyperglycemia and Hyperlipidemia and Regulates Colon Microbiota in Diabetic Db/Db Mice. J. Agric. Food Chem. 2020, 68, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Li, C.; Chen, Q.; Xie, X.; Fu, X.; Chen, C.; Huang, Q.; Huang, Z.; Dong, H. Identification of Polyphenols from Rosa Roxburghii Tratt Pomace and Evaluation of in Vitro and in Vivo Antioxidant Activity. Food Chem. 2022, 377, 131922. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.; Barbosa, J.; Silva, J.; Gomes, A.M.; Pintado, M.; Silva, C.L.M.; Morais, A.M.M.B.; Teixeira, P. A Feasibility Study of Lactobacillus Plantarum in Fruit Powders after Processing and Storage. Int. J. Food Sci. Tech. 2016, 51, 381–388. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of Fermentation on the Antioxidant Activity in Plant-Based Foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of Lactobacillus Strains on Phenolic Profile, Color Attributes and Antioxidant Activities of Lactic-Acid-Fermented Mulberry Juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Fang, J.; Sun, Q.; Wang, Z.; Song, Z.; Geng, J.; Wang, C.; Li, M.; Wang, D. Enhancement of Human Epidermal Cell Defense against UVB Damage by Fermentation of Passiflora Edulis Sims Peel with Saccharomyces Cerevisiae. Nutrients 2023, 15, 501. [Google Scholar] [CrossRef]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of Blueberry and Blackberry Juices Using Lactobacillus Plantarum, Streptococcus Thermophilus and Bifidobacterium Bifidum: Growth of Probiotics, Metabolism of Phenolics, Antioxidant Capacity in Vitro and Sensory Evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, T.; Dai, Y.; Jiang, G.; Peng, Y.; Wang, J.; Song, Y.; Ding, Z. Effects of Probiotics on Antioxidant Activity, Flavor Compounds and Sensory Evaluation of Rosa Roxburghii Tratt. LWT 2023, 179, 114664. [Google Scholar] [CrossRef]
- Latimer, J.A.; Lloyd, J.J.; Diffey, B.L.; Matts, P.J.; Birch-Machin, M.A. Determination of the Action Spectrum of UVR-Induced Mitochondrial DNA Damage in Human Skin Cells. J. Investig. Dermatol. 2015, 135, 2512–2518. [Google Scholar] [CrossRef]
- Gao, W.; Wang, Y.; Hwang, E.; Lin, P.; Bae, J.; Seo, S.A.; Yan, Z.; Yi, T.-H. Rubus Idaeus L. (Red Raspberry) Blocks UVB-Induced MMP Production and Promotes Type I Procollagen Synthesis via Inhibition of MAPK/AP-1, NF-Κβ and Stimulation of TGF-β/Smad, Nrf2 in Normal Human Dermal Fibroblasts. J. Photochem. Photobiol. B Biol. 2018, 185, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Wu, T.; Liu, T.; Yang, H.; Ding, X.; Chen, Y.; Mu, Y. Vicenin-2 Ameliorates Oxidative Damage and Photoaging via Modulation of MAPKs and MMPs Signaling in UVB Radiation Exposed Human Skin Cells. J. Photochem. Photobiol. B Biol. 2019, 190, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, H. Protective Effect of Gelatin Polypeptides from Pacific Cod (Gadus Macrocephalus) against UV Irradiation-Induced Damages by Inhibiting Inflammation and Improving Transforming Growth Factor-β/Smad Signaling Pathway. J. Photochem. Photobiol. B Biol. 2016, 162, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; He, T.; Kang, S.; Voorhees, J.J.; Fisher, G.J. Solar Ultraviolet Irradiation Reduces Collagen in Photoaged Human Skin by Blocking Transforming Growth Factor-β Type II Receptor/Smad Signaling. Am. J. Pathol. 2004, 165, 741–751. [Google Scholar] [CrossRef]
- Bang, J.S.; Choung, S.-Y. Inhibitory Effect of Oyster Hydrolysate on Wrinkle Formation against UVB Irradiation in Human Dermal Fibroblast via MAPK/AP-1 and TGFβ/Smad Pathway. J. Photochem. Photobiol. B Biol. 2020, 209, 111946. [Google Scholar] [CrossRef]
- Kwon, K.-R.; Alam, M.B.; Park, J.-H.; Kim, T.-H.; Lee, S.-H. Attenuation of UVB-Induced Photo-Aging by Polyphenolic-Rich Spatholobus Suberectus Stem Extract Via Modulation of MAPK/AP-1/MMPs Signaling in Human Keratinocytes. Nutrients 2019, 11, 1341. [Google Scholar] [CrossRef]
- Lu, J.; Hou, H.; Fan, Y.; Yang, T.; Li, B. Identification of MMP-1 Inhibitory Peptides from Cod Skin Gelatin Hydrolysates and the Inhibition Mechanism by MAPK Signaling Pathway. J. Funct. Foods 2017, 33, 251–260. [Google Scholar] [CrossRef]
- Alafiatayo, A.A.; Lai, K.-S.; Ahmad, S.; Mahmood, M.; Shaharuddin, N.A. RNA-Seq Analysis Revealed Genes Associated with UV-Induced Cell Necrosis through MAPK/TNF-α Pathways in Human Dermal Fibroblast Cells as an Inducer of Premature Photoaging. Genomics 2020, 112, 484–493. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Chang, C.-T.; Gowrisankar, Y.V.; Chen, X.-Z.; Lin, H.-C.; Yen, H.-R.; Yang, H.-L. Zerumbone Exhibits Antiphotoaging and Dermatoprotective Properties in Ultraviolet A-Irradiated Human Skin Fibroblast Cells via the Activation of Nrf2/ARE Defensive Pathway. Oxidative Med. Cell. Longev. 2019, 2019, 1–14. [Google Scholar] [CrossRef]
- Chen, F.; Tang, Y.; Sun, Y.; Veeraraghavan, V.P.; Mohan, S.K.; Cui, C. 6-Shogaol, a Active Constiuents of Ginger Prevents UVB Radiation Mediated Inflammation and Oxidative Stress through Modulating NrF2 Signaling in Human Epidermal Keratinocytes (HaCaT Cells). J. Photochem. Photobiol. B Biol. 2019, 197, 111518. [Google Scholar] [CrossRef]
- Wu, C.; Li, T.; Qi, J.; Jiang, T.; Xu, H.; Lei, H. Effects of Lactic Acid Fermentation-Based Biotransformation on Phenolic Profiles, Antioxidant Capacity and Flavor Volatiles of Apple Juice. LWT 2020, 122, 109064. [Google Scholar] [CrossRef]
- Li, W.; Ji, J.; Rui, X.; Yu, J.; Tang, W.; Chen, X.; Jiang, M.; Dong, M. Production of Exopolysaccharides by Lactobacillus Helveticus MB2-1 and Its Functional Characteristics in Vitro. LWT Food Sci. Technol. 2014, 59, 732–739. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.-J.; Hu, J.-L.; Nie, Q.-X.; Chen, H.-H.; Xiong, T.; Nie, S.-P.; Xie, M.-Y. Polysaccharide from Fermented Momordica Charantia L. with Lactobacillus Plantarum NCU116 Ameliorates Type 2 Diabetes in Rats. Carbohydr. Polym. 2018, 201, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Hong, R.; Zhang, R.; Yi, Y.; Dong, L.; Liu, L.; Jia, X.; Ma, Y.; Zhang, M. Physicochemical and Biological Properties of Longan Pulp Polysaccharides Modified by Lactobacillus Fermentum Fermentation. Int. J. Biol. Macromol. 2019, 125, 232–237. [Google Scholar] [CrossRef]
- Deng, R.; Zeng, H. Pasteurization Used in the Rosa Roxburghii Pulp. Food Ind. 2017, 38, 124–127. [Google Scholar]
- Liu, P.; Yu, D.; Wang, C.; Zhao, D.; Zhang, J.; Li, M. Optimization of Microbial Fermentation Extraction Process of Panax Notoginseng and Evaluation of Its Anti-Inflammatory Effect. Food Res. Dev. 2019, 40, 71–78+83. [Google Scholar] [CrossRef]
- Yu, L.; Xiang, P.; Ye, L.; Liu, J. Determination and Analysis of Reducing Sugar in Four Kinds of Fruits. Guangdong Chem. Ind. 2021, 48, 243–244+248. [Google Scholar]
- You, S.; Wang, C.; Zhang, J.; Li, M.; Zhao, D.; An, Q. Preliminary Purification and Anti-Aging Activity Evaluation of Total Flavonoids Prepared from Sea-Buckthorn Seed Meal by Fermentation. China Surfactant Deterg. Cosmet. 2019, 49, 661–667. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar] [CrossRef]
- Zhao, D.; Yan, M.; Xu, H.; Liang, H.; Zhang, J.; Li, M.; Wang, C. Antioxidant and Antiaging Activity of Fermented Coix Seed Polysaccharides on Caenorhabditis Elegans. Nutrients 2023, 15, 2474. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Shi, X.; Zhang, J.; Li, L.; Di, F.; Li, M.; Wang, C.; An, Q.; Zhao, D. Role of PI3K-AKT Pathway in Ultraviolet Ray and Hydrogen Peroxide-Induced Oxidative Damage and Its Repair by Grain Ferments. Foods 2023, 12, 806. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, P.J.; Challis, G.L. Discovery of Microbial Natural Products by Activation of Silent Biosynthetic Gene Clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.; Hover, B.M.; Brady, S.F. Culture-Independent Discovery of Natural Products from Soil Metagenomes. J. Ind. Microbiol. Biotechnol. 2016, 43, 129–141. [Google Scholar] [CrossRef]
- Wiemann, P.; Keller, N.P. Strategies for Mining Fungal Natural Products. J. Ind. Microbiol. Biotechnol. 2014, 41, 301–313. [Google Scholar] [CrossRef]
- Peng, W.; Guo, X.; Xu, X.; Zou, D.; Zou, H.; Yang, X. Advances in Polysaccharide Production Based on the Co-Culture of Microbes. Polymers 2023, 15, 2847. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, M.; Jiang, J. Mitochondrial Dysfunction in Neurodegenerative Diseases and Drug Targets via Apoptotic Signaling. Mitochondrion 2019, 49, 35–45. [Google Scholar] [CrossRef]
- Birch-Machin, M.A.; Bowman, A. Oxidative Stress and Ageing. Br. J. Dermatol. 2016, 175, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, D.; Fu, H.; Zhao, D.; Zhang, J.; Li, M.; Wang, C. Protective Effects of Extracellular Proteins of Saccharomycopsis Fibuligera on UVA-Damaged Human Skin Fibroblasts. J. Funct. Foods 2022, 88, 104897. [Google Scholar] [CrossRef]
- Jung, H.-Y.; Shin, J.-C.; Park, S.-M.; Kim, N.-R.; Kwak, W.; Choi, B.-H. Pinus Densiflora Extract Protects Human Skin Fibroblasts against UVB-Induced Photoaging by Inhibiting the Expression of MMPs and Increasing Type I Procollagen Expression. Toxicol. Rep. 2014, 1, 658–666. [Google Scholar] [CrossRef]
- Park, B.; Hwang, E.; Seo, S.A.; Zhang, M.; Park, S.-Y.; Yi, T.-H. Dietary Rosa Damascena Protects against UVB-Induced Skin Aging by Improving Collagen Synthesis via MMPs Reduction through Alterations of c-Jun and c-Fos and TGF-Β1 Stimulation Mediated Smad2/3 and Smad7. J. Funct. Foods 2017, 36, 480–489. [Google Scholar] [CrossRef]
- Hirota, A.; Kawachi, Y.; Yamamoto, M.; Koga, T.; Hamada, K.; Otsuka, F. Acceleration of UVB-Induced Photoageing in Nrf2 Gene-Deficient Mice: Acceleration of Photoageing in Nrf2-Null Mice. Exp. Dermatol. 2011, 20, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Thulabandu, V.; Chen, D.; Atit, R.P. Dermal Fibroblast in Cutaneous Development and Healing. WIREs Dev. Biol. 2018, 7, e307. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | pH | Conductivities (μS/cm) | Viscosities (mPa·S) |
|---|---|---|---|
| LP-20241 | 4.12 | 1766 | 17.4 |
| LH-20243 | 4.01 | 1654 | 17.4 |
| LK-20260 | 3.99 | 1712 | 16.9 |
| PLS-20261 | 4.36 | 1822 | 19.1 |
| LYS-20297 | 4.27 | 1824 | 16.0 |
| Concentration (mg/mL) | LP-20241 | LH-20243 | LK-20260 | PLS-20261 | LYS-20297 |
|---|---|---|---|---|---|
| Total sugars | 85.78 ± 0.005 *** | 72.83 ± 0.006 *** | 69.44 ± 0.002 *** | 73.45 ± 0.005 *** | 130.92 ± 0.009 |
| Reducing sugars | 9.21 ± 0.083 *** | 7.76 ± 0.139 *** | 7.51 ± 0.131 *** | 6.59 ± 0.037 *** | 10.50 ± 0.083 |
| Flavonoids | 1.40 ± 0.026 *** | 3.14 ± 0.035 *** | 1.10 ± 0.016 *** | 1.40 ± 0.018 *** | 1.81 ± 0.035 |
| Polyphenols | 8.75 ± 0.001 *** | 11.20 ± 0.004 ** | 11.66 ± 0.002 *** | 8.46 ± 0.002 *** | 10.46 ± 0.001 |
| Proteins | 46.02 ± 0.026 * | 45.00 ± 0.001 ** | 46.02 ± 0.012 * | 44.33 ± 0.008 ** | 49.06 ± 0.017 |
| Gene Name | Gene Description | C vs. M | M vs. R |
|---|---|---|---|
| Hmox1 | Heme oxygenase 1 | DOWN | UP |
| Jun | Jun proto-oncogene Symbol; Acc: | UP | DOWN |
| Fos | FBJ osteosarcoma oncogene | UP | DOWN |
| Egfr | Epidermal growth factor receptor | DOWN | UP |
| Col4a6 | Collagen, Type IV, Alpha 6 | DOWN | UP |
| Col4a1 | Collagen, Type IV, Alpha 1 | DOWN | UP |
| Col4a2 | Collagen, Type IV, Alpha 2 | DOWN | UP |
| Mmp2 | Matrix metallopeptidase 2 | UP | DOWN |
| Tgfb2 | Transforming growth factor, Beta 2 | DOWN | UP |
| Bcl2 | B cell leukemia/lymphoma 2 | DOWN | UP |
| Mmp14 | Matrix metallopeptidase 14 | UP | DOWN |
| Mmp9 | Matrix metallopeptidase 9 | UP | DOWN |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, M.; Fu, H.; Mo, Q.; Wang, S.; Wang, C.; Wang, D.; Zhang, J.; Li, M. Protective Mechanism of Rosa roxburghii Tratt Fermentation Broth against Ultraviolet-A-Induced Photoaging of Human Embryonic Skin Fibroblasts. Antioxidants 2024, 13, 382. https://doi.org/10.3390/antiox13030382
Yuan M, Fu H, Mo Q, Wang S, Wang C, Wang D, Zhang J, Li M. Protective Mechanism of Rosa roxburghii Tratt Fermentation Broth against Ultraviolet-A-Induced Photoaging of Human Embryonic Skin Fibroblasts. Antioxidants. 2024; 13(3):382. https://doi.org/10.3390/antiox13030382
Chicago/Turabian StyleYuan, Minglu, Hao Fu, Qiuting Mo, Shiwei Wang, Changtao Wang, Dongdong Wang, Jiachan Zhang, and Meng Li. 2024. "Protective Mechanism of Rosa roxburghii Tratt Fermentation Broth against Ultraviolet-A-Induced Photoaging of Human Embryonic Skin Fibroblasts" Antioxidants 13, no. 3: 382. https://doi.org/10.3390/antiox13030382
APA StyleYuan, M., Fu, H., Mo, Q., Wang, S., Wang, C., Wang, D., Zhang, J., & Li, M. (2024). Protective Mechanism of Rosa roxburghii Tratt Fermentation Broth against Ultraviolet-A-Induced Photoaging of Human Embryonic Skin Fibroblasts. Antioxidants, 13(3), 382. https://doi.org/10.3390/antiox13030382