
Cyanide Insensitive Oxidase Confers Hydrogen Sulfide and Nitric Oxide Tolerance to Pseudomonas aeruginosa Aerobic Respiration

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains, Growth Conditions, and Membrane Preparations
2.3. Oxygraphic Measurements
2.4. RNA Extraction and RT-qPCR Analyses
3. Results
3.1. Effect of Reducing Systems on Respiration of P. aeruginosa Membranes
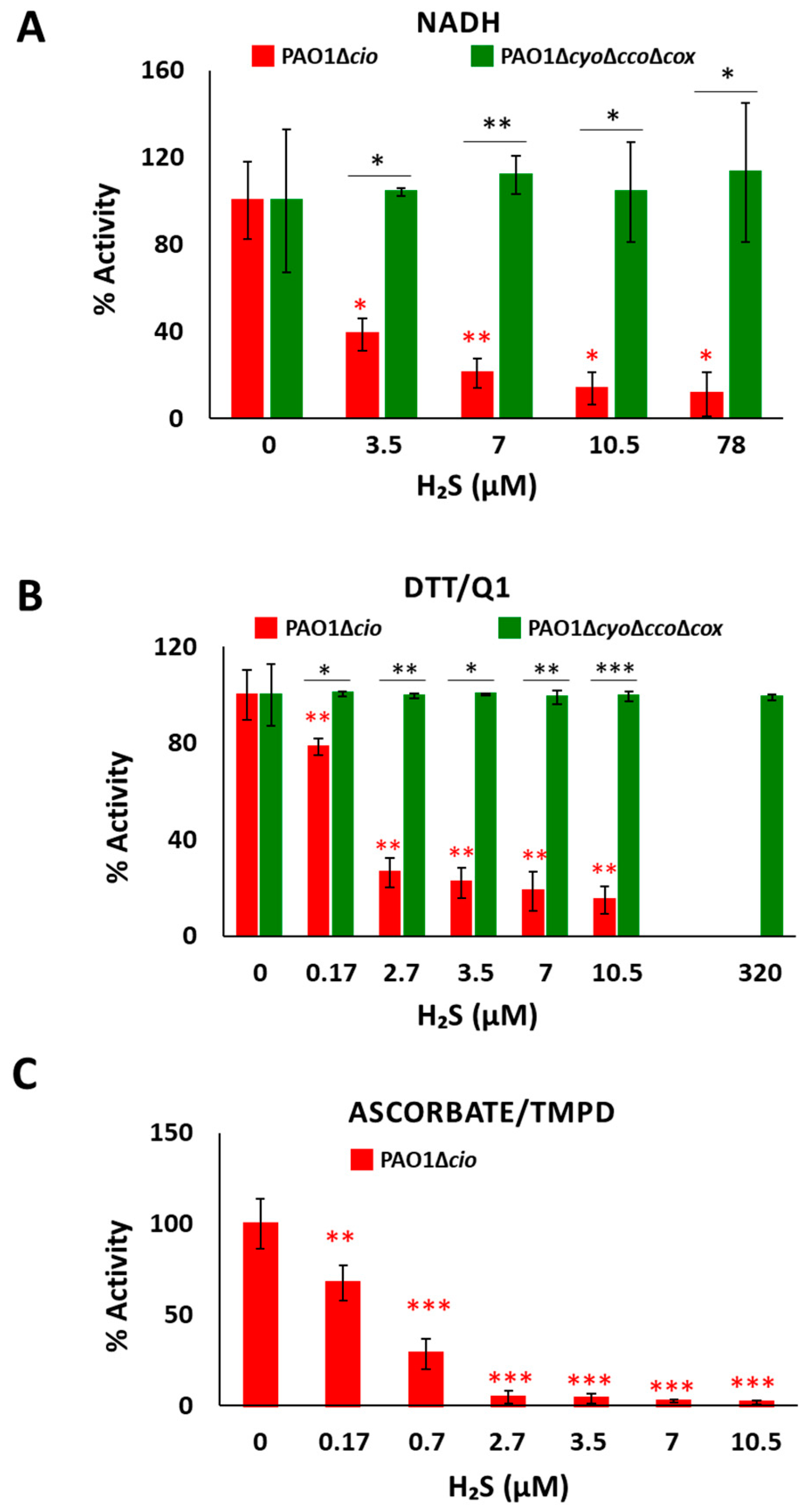
3.2. Effect of Sulfide on Respiration of P. aeruginosa Membrane Preparations
3.3. Effect of Sulfide on P. aeruginosa Cell Growth
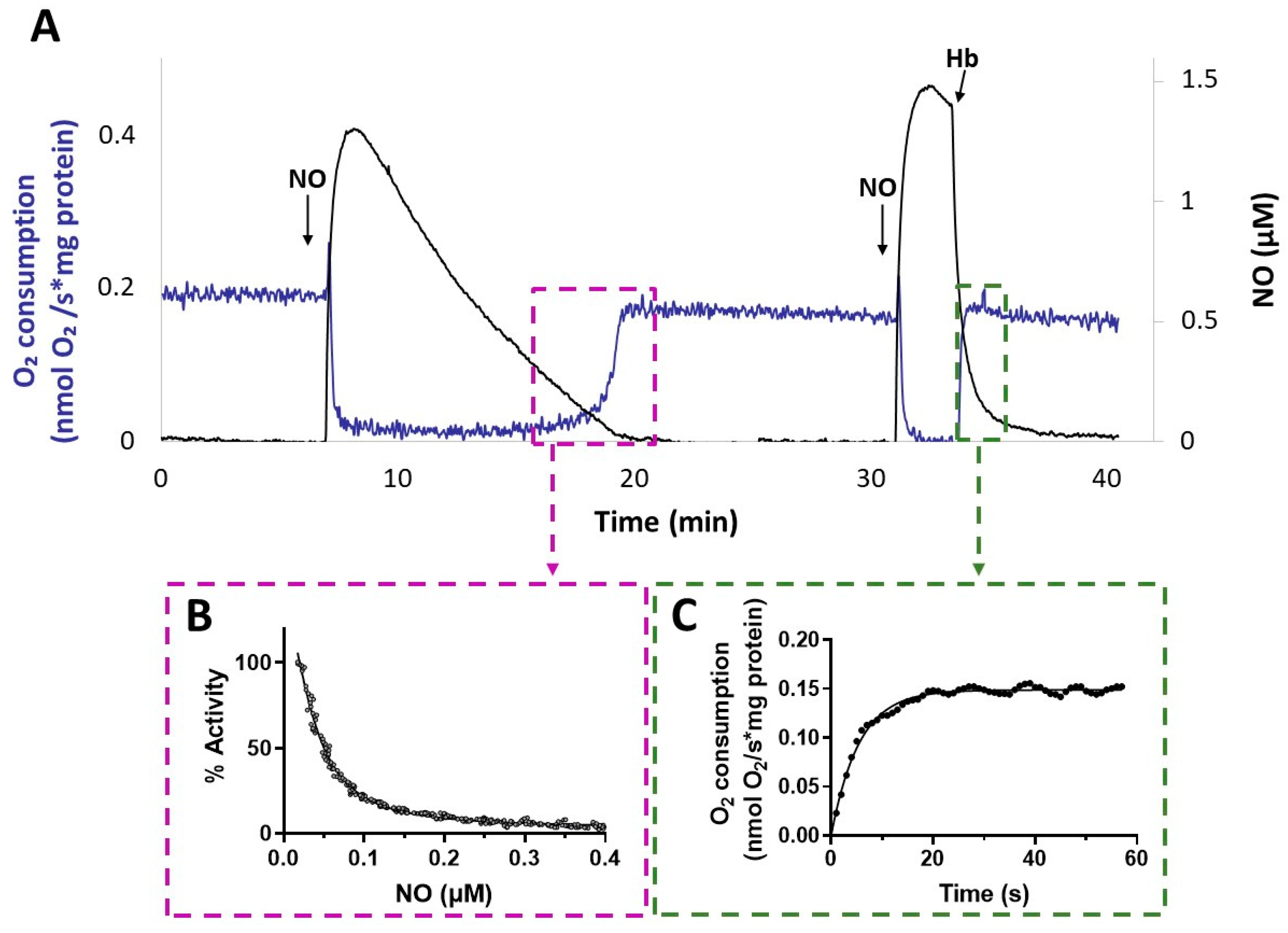
3.4. Effect of NO on Respiration of P. aeruginosa Membranes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szabo, C. A Timeline of Hydrogen Sulfide (H2S) Research: From Environmental Toxin to Biological Mediator. Biochem. Pharmacol. 2018, 149, 5–19. [Google Scholar] [CrossRef]
- Deshmukh, R.; Harwansh, R.K.; Bandyopadhyay, N.; Bandopadhyay, S.; Kumar, P. Pharmacology of Gasotransmitters (Nitric Oxide and Carbon Monoxide) and Their Action. In Frontiers in Pharmacology of Neurotransmitters; Springer: Singapore, 2020; pp. 579–617. [Google Scholar]
- Lo Faro, M.L.; Fox, B.; Whatmore, J.L.; Winyard, P.G.; Whiteman, M. Hydrogen Sulfide and Nitric Oxide Interactions in Inflammation. Nitric Oxide 2014, 41, 38–47. [Google Scholar] [CrossRef]
- Mancuso, C.; Navarra, P.; Preziosi, P. Roles of Nitric Oxide, Carbon Monoxide, and Hydrogen Sulfide in the Regulation of the Hypothalamic–Pituitary–Adrenal Axis. J. Neurochem. 2010, 113, 563–575. [Google Scholar] [CrossRef]
- Farrugia, G.; Szurszewski, J.H. Carbon Monoxide, Hydrogen Sulfide, and Nitric Oxide as Signaling Molecules in the Gastrointestinal Tract. Gastroenterology 2014, 147, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R.; Donald, J.A. Nervous Control of Circulation--the Role of Gasotransmitters, NO, CO, and H2S. Acta Histochem. 2009, 111, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Wareham, L.K.; Southam, H.M.; Poole, R.K. Do Nitric Oxide, Carbon Monoxide and Hydrogen Sulfide Really Qualify as “gasotransmitters” in Bacteria? Biochem. Soc. Trans. 2018, 46, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Jesse, H.E.; Poole, R.K.; Tinajero-Trejo, M. Gasotransmitters, Poisons, and Antimicrobials: It’s a Gas, Gas, Gas! F1000Prime Rep. 2013, 5, 28. [Google Scholar] [CrossRef]
- Toliver-Kinsky, T.; Cui, W.; Törö, G.; Lee, S.J.; Shatalin, K.; Nudler, E.; Szaboa, C. H2S, a Bacterial Defense Mechanism against the Host Immune Response. Infect. Immun. 2018, 87, e00272-18. [Google Scholar] [CrossRef] [PubMed]
- Luhachack, L.; Nudler, E. Bacterial Gasotransmitters: An Innate Defense against Antibiotics. Curr. Opin. Microbiol. 2014, 21, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.; Seregina, T.; Nagornykh, M.; Luhachack, L.G.; Korolkova, N.; Lopes, L.E.; Kotova, V.; Zavilgelsky, G.; Shakulov, R.; Shatalin, K.; et al. Mechanism of H2S-Mediated Protection against Oxidative Stress in Escherichia coli. Proc. Natl. Acad. Sci. USA 2017, 114, 6022–6027. [Google Scholar] [CrossRef]
- Bryan, N.S.; Lefer, D.J. Update on Gaseous Signaling Molecules Nitric Oxide and Hydrogen Sulfide: Strategies to Capture Their Functional Activity for Human Therapeutics. Mol. Pharmacol. 2019, 96, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.S.; Miranda, V.; Saraiva, L.M. Hydrogen Sulfide and Carbon Monoxide Tolerance in Bacteria. Antioxidants 2021, 10, 729. [Google Scholar] [CrossRef] [PubMed]
- Shatalin, K.; Shatalina, E.; Mironov, A.; Nudler, E. H2S: A Universal Defense against Antibiotics in Bacteria. Science 2011, 334, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Gusarov, I.; Shatalin, K.; Starodubtseva, M.; Nudler, E. Endogenous Nitric Oxide Protects Bacteria against a Wide Spectrum of Antibiotics. Science 2009, 325, 1380–1384. [Google Scholar] [CrossRef]
- Walsh, B.J.C.; Giedroc, D.P. H2S and Reactive Sulfur Signaling at the Host-Bacterial Pathogen Interface. J. Biol. Chem. 2020, 295, 13150–13168. [Google Scholar] [CrossRef] [PubMed]
- Pal, V.K.; Bandyopadhyay, P.; Singh, A. Hydrogen Sulfide in Physiology and Pathogenesis of Bacteria and Viruses. IUBMB Life 2018, 70, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Shen, J.; Edmonds, K.A.; Luebke, J.L.; Hickey, A.K.; Palmer, L.D.; Chang, F.-M.J.; Bruce, K.A.; Kehl-Fie, T.E.; Skaar, E.P.; et al. Sulfide Homeostasis and Nitroxyl Intersect via Formation of Reactive Sulfur Species in Staphylococcus aureus. mSphere 2017, 2, e00082-17. [Google Scholar] [CrossRef] [PubMed]
- Kaila, V.R.I.; Wikström, M. Architecture of Bacterial Respiratory Chains. Nat. Rev. Microbiol. 2021, 19, 319–330. [Google Scholar] [CrossRef]
- Hards, K.; Cook, G.M. Targeting Bacterial Energetics to Produce New Antimicrobials. Drug Resist. Updat. 2018, 36, 1–12. [Google Scholar] [CrossRef]
- Folkesson, A.; Jelsbak, L.; Yang, L.; Johansen, H.K.; Ciofu, O.; Hoiby, N.; Molin, S. Adaptation of Pseudomonas aeruginosa to the Cystic Fibrosis Airway: An Evolutionary Perspective. Nat. Rev. Microbiol. 2012, 10, 841–851. [Google Scholar] [CrossRef]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa Evolutionary Adaptation and Diversification in Cystic Fibrosis Chronic Lung Infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 27 February 2017).
- Borisov, V.B.; Siletsky, S.A.; Paiardini, A.; Hoogewijs, D.; Forte, E.; Giuffrè, A.; Poole, R.K. Bacterial Oxidases of the Cytochrome bd Family: Redox Enzymes of Unique Structure, Function, and Utility as Drug Targets. Antioxid. Redox Signal 2021, 34, 1280–1318. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, L.; Pitt, M.; Williams, H.D. The CioAB Genes from Pseudomonas aeruginosa Code for a Novel Cyanide-Insensitive Terminal Oxidase Related to the Cytochrome bd Quinol Oxidases. Mol. Microbiol. 1997, 24, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Arai, H. Regulation and Function of Versatile Aerobic and Anaerobic Respiratory Metabolism in Pseudomonas aeruginosa. Front. Microbiol. 2011, 2, 103. [Google Scholar] [CrossRef]
- Alvarez-Ortega, C.; Harwood, C.S. Responses of Pseudomonas aeruginosa to Low Oxygen Indicate That Growth in the Cystic Fibrosis Lung Is by Aerobic Respiration. Mol. Microbiol. 2007, 65, 153–165. [Google Scholar] [CrossRef]
- Arai, H.; Kawakami, T.; Osamura, T.; Hirai, T.; Sakai, Y.; Ishii, M. Enzymatic Characterization and in vivo Function of Five Terminal Oxidases in Pseudomonas aeruginosa. J. Bacteriol. 2014, 196, 4206–4215. [Google Scholar] [CrossRef]
- Zlosnik, J.E.A.; Tavankar, G.R.; Bundy, J.G.; Mossialos, D.; O’Toole, R.; Williams, H.D. Investigation of the Physiological Relationship between the Cyanide-Insensitive Oxidase and Cyanide Production in Pseudomonas aeruginosa. Microbiology 2006, 152, 1407–1415. [Google Scholar] [CrossRef]
- Cunningham, L.; Williams, H.D. Isolation and Characterization of Mutants Defective in the Cyanide-Insensitive Respiratory Pathway of Pseudomonas aeruginosa. J. Bacteriol. 1995, 177, 432–438. [Google Scholar] [CrossRef][Green Version]
- Cooper, M.; Tavankar, G.R.; Williams, H.D. Regulation of Expression of the Cyanide-Insensitive Terminal Oxidase in Pseudomonas aeruginosa. Microbiology 2003, 149, 1275–1284. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hirai, T.; Osamura, T.; Ishii, M.; Arai, H. Expression of Multiple cbb3 Cytochrome c Oxidase Isoforms by Combinations of Multiple Isosubunits in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2016, 113, 12815–12819. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, M.; Achard, M.E.S.; Idris, A.; Totsika, M.; Phan, M.D.; Peters, K.M.; Sarkar, S.; Ribeiro, C.A.; Holyoake, L.V.; Ladakis, D.; et al. The Cytochrome bd-I Respiratory Oxidase Augments Survival of Multidrug-Resistant Escherichia coli during Infection. Sci. Rep. 2016, 6, 35285. [Google Scholar] [CrossRef]
- Seregina, T.A.; Lobanov, K.V.; Shakulov, R.S.; Mironov, A.S. Inactivation of terminal oxidase bd-I leads to supersensitivity of E. coli to quinolone and beta-lactam antibiotics. Mol. Biol. (Mosk) 2022, 56, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Borisov, V.B.; Forte, E.; Siletsky, S.A.; Sarti, P.; Giuffrè, A. Cytochrome bd from Escherichia coli Catalyzes Peroxynitrite Decomposition. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 182–188. [Google Scholar] [CrossRef]
- Borisov, V.B.; Forte, E. Bioenergetics and Reactive Nitrogen Species in Bacteria. Int. J. Mol. Sci. 2022, 23, 7321. [Google Scholar] [CrossRef]
- Borisov, V.B.; Forte, E.; Konstantinov, A.A.; Poole, R.K.; Sarti, P.; Giuffrè, A. Interaction of the Bacterial Terminal Oxidase Cytochrome bd with Nitric Oxide. FEBS Lett. 2004, 576, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.G.; Shepherd, M.; Nicholls, P.; Dobbin, P.S.; Dodsworth, K.S.; Poole, R.K.; Cooper, C.E. Cytochrome bd Confers Nitric Oxide Resistance to Escherichia coli. Nat. Chem. Biol. 2009, 5, 94–96. [Google Scholar] [CrossRef]
- Borisov, V.B.; Forte, E.; Davletshin, A.; Mastronicola, D.; Sarti, P.; Giuffrè, A. Cytochrome bd Oxidase from Escherichia coli Displays High Catalase Activity: An Additional Defense against Oxidative Stress. FEBS Lett. 2013, 587, 2214–2218. [Google Scholar] [CrossRef]
- Borisov, V.B.; Nastasi, M.R.; Forte, E. Cytochrome bd as Antioxidant Redox Enzyme. Mol. Biol. 2023, 57, 1077–1084. [Google Scholar] [CrossRef]
- Al-Attar, S.; Yu, Y.; Pinkse, M.; Hoeser, J.; Friedrich, T.; Bald, D.; De Vries, S. Cytochrome bd Displays Significant Quinol Peroxidase Activity. Sci. Rep. 2016, 6, 27631. [Google Scholar] [CrossRef]
- Forte, E.; Borisov, V.B.; Falabella, M.; Colaço, H.G.; Tinajero-Trejo, M.; Poole, R.K.; Vicente, J.B.; Sarti, P.; Giuffrè, A. The Terminal Oxidase Cytochrome bd Promotes Sulfide-Resistant Bacterial Respiration and Growth. Sci. Rep. 2016, 6, 23788. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; Falcone, M.; Molin, S.; Johansen, H.K. High-Resolution in Situ Transcriptomics of Pseudomonas aeruginosa Unveils Genotype Independent Patho-Phenotypes in Cystic Fibrosis Lungs. Nat. Commun. 2018, 9, 3459. [Google Scholar] [CrossRef]
- Cowley, E.S.; Kopf, S.H.; Lariviere, A.; Ziebis, W.; Newman, D.K. Pediatric Cystic Fibrosis Sputum Can Be Chemically Dynamic, Anoxic, and Extremely Reduced Due to Hydrogen Sulfide Formation. mBio 2015, 6, e00767-15. [Google Scholar] [CrossRef]
- Nagy, P.; Pálinkás, Z.; Nagy, A.; Budai, B.; Tóth, I.; Vasas, A. Chemical Aspects of Hydrogen Sulfide Measurements in Physiological Samples. Biochim. Biophys. Acta 2014, 1840, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Nashef, A.S.; Osuga, D.T.; Feeney, R.E. Determination of Hydrogen Sulfide with 5,5′-Dithiobis-(2-Nitrobenzoic Acid), N-Ethylmaleimide, and Parachloromercuribenzoate. Anal. Biochem. 1977, 79, 394–405. [Google Scholar] [CrossRef]
- Stubauer, G.; Giuffrè, A.; Brunori, M.; Sarti, P. Cytochrome c Oxidase Does Not Catalyze the Anaerobic Reduction of NO. Biochem. Biophys. Res. Commun. 1998, 245, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, S.; Visca, P.; Frangipani, E. Gallium-Protoporphyrin IX Inhibits Pseudomonas aeruginosa Growth by Targeting Cytochromes. Front. Cell Infect. Microbiol. 2017, 7, 12. [Google Scholar] [CrossRef]
- Frangipani, E.; Slaveykova, V.I.; Reimmann, C.; Haas, D. Adaptation of Aerobically Growing Pseudomonas aeruginosa to Copper Starvation. J. Bacteriol. 2008, 190, 6706–6717. [Google Scholar] [CrossRef]
- Sarti, P.; Giuffré, A.; Forte, E.; Mastronicola, D.; Barone, M.C.; Brunori, M. Nitric Oxide and Cytochrome c Oxidase: Mechanisms of Inhibition and NO Degradation. Biochem. Biophys. Res. Commun. 2000, 274, 183–187. [Google Scholar] [CrossRef]
- Letizia, M.; Mellini, M.; Fortuna, A.; Visca, P.; Imperi, F.; Leoni, L.; Rampioni, G. PqsE Expands and Differentially Modulates the RhlR Quorum Sensing Regulon in Pseudomonas aeruginosa. Microbiol. Spectr. 2022, 10, e0096122. [Google Scholar] [CrossRef]
- Nicholls, P.; Marshall, D.C.; Cooper, C.E.; Wilson, M.T. Sulfide Inhibition of and Metabolism by Cytochrome c Oxidase. Biochem. Soc. Trans. 2013, 41, 1312–1316. [Google Scholar] [CrossRef]
- Kawakami, T.; Kuroki, M.; Ishii, M.; Igarashi, Y.; Arai, H. Differential Expression of Multiple Terminal Oxidases for Aerobic Respiration in Pseudomonas aeruginosa. Env. Environ. Microbiol. 2010, 12, 1399–1412. [Google Scholar] [CrossRef]
- Borisov, V.B.; Forte, E.; Sarti, P.; Brunori, M.; Konstantinov, A.A.; Giuffrè, A. Redox Control of Fast Ligand Dissociation from Escherichia coli Cytochrome bd. Biochem. Biophys. Res. Commun. 2007, 355, 97–102. [Google Scholar] [CrossRef]
- Le Laz, S.; Kpebe, A.; Bauzan, M.; Lignon, S.; Rousset, M.; Brugna, M. Expression of Terminal Oxidases under Nutrient-Starved Conditions in Shewanella oneidensis: Detection of the A-Type Cytochrome c Oxidase. Sci. Rep. 2016, 6, 19726. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Garcia-Robledo, E.; Lund, M.B.; Lehner, P.; Borisov, S.M.; Klimant, I.; Revsbech, N.P.; Schramm, A. Gene Expression of Terminal Oxidases in Two Marine Bacterial Strains Exposed to Nanomolar Oxygen Concentrations. FEMS Microbiol. Ecol. 2018, 94, fiy072. [Google Scholar] [CrossRef]
- Trojan, D.; Garcia-Robledo, E.; Meier, D.V.; Hausmann, B.; Revsbech, N.P.; Eichorst, S.A.; Woebken, D. Microaerobic Lifestyle at Nanomolar O2 Concentrations Mediated by Low-Affinity Terminal Oxidases in Abundant Soil Bacteria. mSystems 2021, 6, e00250-21. [Google Scholar] [CrossRef]
- Lau, G.W.; Hassett, D.J.; Ran, H.; Kong, F. The Role of Pyocyanin in Pseudomonas aeruginosa Infection. Trends Mol. Med. 2004, 10, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, L.E.P.; Price-Whelan, A.; Petersen, A.; Whiteley, M.; Newman, D.K. The Phenazine Pyocyanin Is a Terminal Signalling Factor in the Quorum Sensing Network of Pseudomonas aeruginosa. Mol. Microbiol. 2006, 61, 1308–1321. [Google Scholar] [CrossRef]
- Shatalin, K.; Nuthanakanti, A.; Kaushik, A.; Shishov, D.; Peselis, A.; Shamovsky, I.; Pani, B.; Lechpammer, M.; Vasilyev, N.; Shatalina, E.; et al. Inhibitors of Bacterial H2S Biogenesis Targeting Antibiotic Resistance and Tolerance. Science 2021, 372, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Saini, V.; Chinta, K.C.; Reddy, V.P.; Glasgow, J.N.; Stein, A.; Lamprecht, D.A.; Rahman, M.A.; Mackenzie, J.S.; Truebody, B.E.; Adamson, J.H.; et al. Hydrogen Sulfide Stimulates Mycobacterium tuberculosis Respiration, Growth and Pathogenesis. Nat. Commun. 2020, 11, 557. [Google Scholar] [CrossRef]
- Kunota, T.T.R.; Rahman, M.A.; Truebody, B.E.; Mackenzie, J.S.; Saini, V.; Lamprecht, D.A.; Adamson, J.H.; Sevalkar, R.R.; Lancaster, J.R.; Berney, M.; et al. Mycobacterium tuberculosis H2S Functions as a Sink to Modulate Central Metabolism, Bioenergetics, and Drug Susceptibility. Antioxidants 2021, 10, 1285. [Google Scholar] [CrossRef]
- Rozansky, R.; Weber, D.; Gurevitch, J. Production of Hydrogen Sulfide by Pseudomonas aeruginosa. Am. J. Clin. Pathol. 1950, 20, 1090–1091. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Camacho, M.I.; Chen, Y.; Bhat, A.H.; Chang, C.; Peluso, E.A.; Wu, C.; Das, A.; Ton-That, H. Genetic Determinants of Hydrogen Sulfide Biosynthesis in Fusobacterium nucleatum Are Required for Bacterial Fitness, Antibiotic Sensitivity, and Virulence. mBio 2022, 13, e01936-22. [Google Scholar] [CrossRef]
- Seregina, T.A.; Lobanov, K.V.; Shakulov, R.S.; Mironov, A.S. Enhancement of the Bactericidal Effect of Antibiotics by Inhibition of Enzymes Involved in Production of Hydrogen Sulfide in Bacteria. Mol. Biol. 2022, 56, 638–648. [Google Scholar] [CrossRef]
- Weikum, J.; Ritzmann, N.; Jelden, N.; Klöckner, A.; Herkersdorf, S.; Josten, M.; Sahl, H.-G.; Grein, F. Sulfide Protects Staphylococcus aureus from Aminoglycoside Antibiotics but Cannot Be Regarded as a General Defense Mechanism against Antibiotics. Antimicrob. Agents Chemother. 2018, 62, e00602-18. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Ong, K.X.; Surendran, S.T.; Sinha, A.; Lai, J.J.H.; Chen, J.; Liang, J.; Tay, L.K.S.; Cui, L.; Loo, H.L.; et al. Hydrogen Sulfide Sensitizes Acinetobacter baumannii to Killing by Antibiotics. Front. Microbiol. 2020, 11, 563226. [Google Scholar] [CrossRef] [PubMed]
- Caruso, L.; Mellini, M.; Catalano Gonzaga, O.; Astegno, A.; Forte, E.; Di Matteo, A.; Giuffrè, A.; Visca, P.; Imperi, F.; Leoni, L.; et al. Hydrogen Sulfide Production Does Not Affect Antibiotic Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2024, 6, e0007524. [Google Scholar] [CrossRef] [PubMed]
- Xuan, G.; Lü, C.; Xu, H.; Li, K.; Liu, H.; Xia, Y.; Xun, L. Sulfane Sulfur Regulates LasR-Mediated Quorum Sensing and Virulence in Pseudomonas aeruginosa PAO1. Antioxidants 2021, 10, 1498. [Google Scholar] [CrossRef] [PubMed]
- Beebout, C.J.; Eberly, A.R.; Werby, S.H.; Reasoner, S.A.; Brannon, J.R.; De, S.; Fitzgerald, M.J.; Huggins, M.M.; Clayton, D.B.; Cegelski, L.; et al. Respiratory Heterogeneity Shapes Biofilm Formation and Host Colonization in Uropathogenic Escherichia coli. mBio 2019, 10, e02400-18. [Google Scholar] [CrossRef] [PubMed]
- Jones-Carson, J.; Husain, M.; Liu, L.; Orlicky, D.J.; Vázquez-Torres, A. Cytochrome bd -Dependent Bioenergetics and Antinitrosative Defenses in Salmonella Pathogenesis. mBio 2016, 7, e02052-16. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.R.; Dunman, P.M.; Fang, F.C. The Nitrosative Stress Response of Staphylococcus aureus Is Required for Resistance to Innate Immunity. Mol. Microbiol. 2006, 61, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Sohaskey, C.D.; Kana, B.D.; Dawes, S.; North, R.J.; Mizrahi, V.; Gennaro, M.L. Changes in Energy Metabolism of Mycobacterium tuberculosis in Mouse Lung and under in vitro Conditions Affecting Aerobic Respiration. Proc. Natl. Acad. Sci. USA 2005, 102, 15629–15634. [Google Scholar] [CrossRef]
- Moore, C.M.; Nakano, M.M.; Wang, T.; Ye, R.W.; Helmann, J.D. Response of Bacillus subtilis to Nitric Oxide and the Nitrosating Agent Sodium Nitroprusside. J. Bacteriol. 2004, 186, 4655–4664. [Google Scholar] [CrossRef] [PubMed]
- Arjona, D.; Wikström, M.; Ädelroth, P. Nitric Oxide Is a Potent Inhibitor of the cbb3-Type Heme-Copper Oxidases. FEBS Lett. 2015, 589, 1214–1218. [Google Scholar] [CrossRef]
- Brown, G.C.; Cooper, C.E. Nanomolar Concentrations of Nitric Oxide Reversibly Inhibit Synaptosomal Respiration by Competing with Oxygen at Cytochrome Oxidase. FEBS Lett. 1994, 356, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, A.; Borisov, V.B.; Mastronicola, D.; Sarti, P.; Forte, E. Cytochrome bd Oxidase and Nitric Oxide: From Reaction Mechanisms to Bacterial Physiology. FEBS Lett. 2012, 586, 622–629. [Google Scholar] [CrossRef]
- Fang, F.C. Perspectives Series: Host/Pathogen Interactions. Mechanisms of Nitric Oxide-Related Antimicrobial Activity. J. Clin. Investig. 1997, 99, 2818–2825. [Google Scholar] [CrossRef] [PubMed]
- Cutruzzolà, F.; Frankenberg-Dinkel, N. Origin and Impact of Nitric Oxide in Pseudomonas aeruginosa Biofilms. J. Bacteriol. 2016, 198, 55–65. [Google Scholar] [CrossRef]
- Arai, H.; Hayashi, M.; Kuroi, A.; Ishii, M.; Igarashi, Y. Transcriptional Regulation of the Flavohemoglobin Gene for Aerobic Nitric Oxide Detoxification by the Second Nitric Oxide-Responsive Regulator of Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 3960–3968. [Google Scholar] [CrossRef]
- Kakishima, K.; Shiratsuchi, A.; Taoka, A.; Nakanishi, Y.; Fukumori, Y. Participation of Nitric Oxide Reductase in Survival of Pseudomonas aeruginosa in LPS-Activated Macrophages. Biochem. Biophys. Res. Commun. 2007, 355, 587–591. [Google Scholar] [CrossRef]
- Arai, H.; Kodama, T.; Igarashi, Y. Effect of Nitrogen Oxides on Expression of the nir and nor Genes for Denitrification in Pseudomonas aeruginosa. FEMS Microbiol. Lett. 1999, 170, 19–24. [Google Scholar] [CrossRef]
- Friedrich, T.; Wohlwend, D.; Borisov, V.B. Recent Advances in Structural Studies of Cytochrome bd and Its Potential Application as a Drug Target. Int. J. Mol. Sci. 2022, 23, 3166. [Google Scholar] [CrossRef]
- Harikishore, A.; Mathiyazakan, V.; Pethe, K.; Gruber, G. Novel targets and inhibitors of the Mycobacterium tuberculosis cytochrome bd oxidase to foster anti-tuberculosis drug discovery. Expert Opin. Drug Discov. 2023, 18, 917–927. [Google Scholar] [CrossRef]
- Henry, S.A.; Webster, C.M.; Shaw, L.N.; Torres, N.J.; Jobson, M.E.; Totzke, B.C.; Jackson, J.K.; McGreig, J.E.; Wass, M.N.; Robinson, G.K.; et al. Steroid drugs inhibit bacterial respiratory oxidases and are lethal toward methicillin-resistant Staphylococcus aureus. J. Infect. Dis. 2024, jiad540. [Google Scholar] [CrossRef] [PubMed]
- Saha, P.; Das, S.; Indurthi, H.K.; Kumar, R.; Roy, A.; Kalia, N.P.; Sharma, D.K. Cytochrome bd oxidase: An emerging anti-tubercular drug target. RSC Med. Chem. 2024, 15, 769–787. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nastasi, M.R.; Caruso, L.; Giordano, F.; Mellini, M.; Rampioni, G.; Giuffrè, A.; Forte, E. Cyanide Insensitive Oxidase Confers Hydrogen Sulfide and Nitric Oxide Tolerance to Pseudomonas aeruginosa Aerobic Respiration. Antioxidants 2024, 13, 383. https://doi.org/10.3390/antiox13030383
Nastasi MR, Caruso L, Giordano F, Mellini M, Rampioni G, Giuffrè A, Forte E. Cyanide Insensitive Oxidase Confers Hydrogen Sulfide and Nitric Oxide Tolerance to Pseudomonas aeruginosa Aerobic Respiration. Antioxidants. 2024; 13(3):383. https://doi.org/10.3390/antiox13030383
Chicago/Turabian StyleNastasi, Martina R., Lorenzo Caruso, Francesca Giordano, Marta Mellini, Giordano Rampioni, Alessandro Giuffrè, and Elena Forte. 2024. "Cyanide Insensitive Oxidase Confers Hydrogen Sulfide and Nitric Oxide Tolerance to Pseudomonas aeruginosa Aerobic Respiration" Antioxidants 13, no. 3: 383. https://doi.org/10.3390/antiox13030383
APA StyleNastasi, M. R., Caruso, L., Giordano, F., Mellini, M., Rampioni, G., Giuffrè, A., & Forte, E. (2024). Cyanide Insensitive Oxidase Confers Hydrogen Sulfide and Nitric Oxide Tolerance to Pseudomonas aeruginosa Aerobic Respiration. Antioxidants, 13(3), 383. https://doi.org/10.3390/antiox13030383