
Evaluation of Hypoglycemic Polyphenolic Compounds in Blueberry Extract: Functional Effects and Mechanisms




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Analysis of Polyphenol
2.3. Cell Culture and Treatments
2.4. MTT Assay
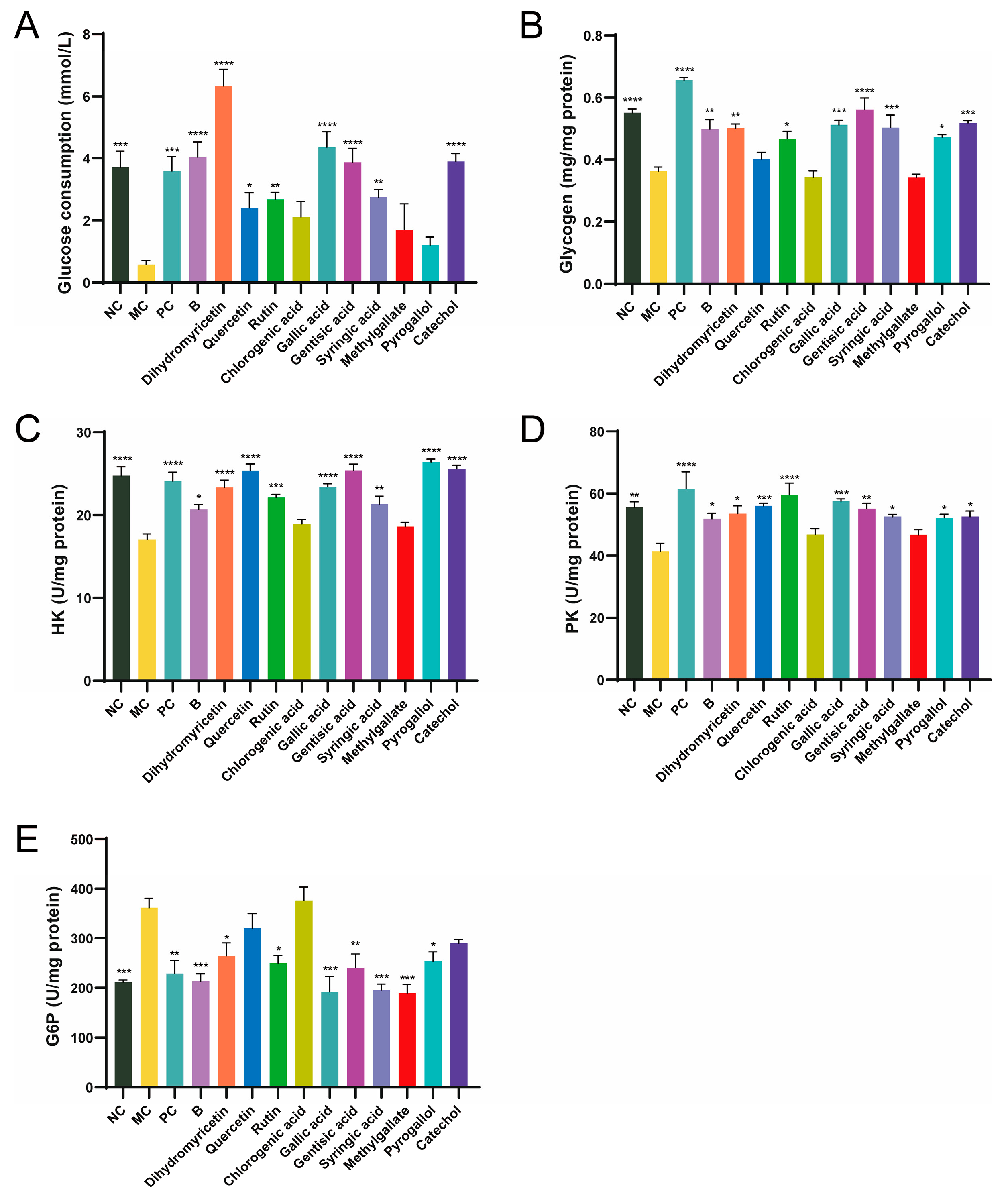
2.5. Glucose Consumption Assay
2.6. Glycogen, Hexokinase (HK), Pyruvate Kinase (PK), Glucose-6-Phosphatase (G6P)
2.7. Animal Experiment Design
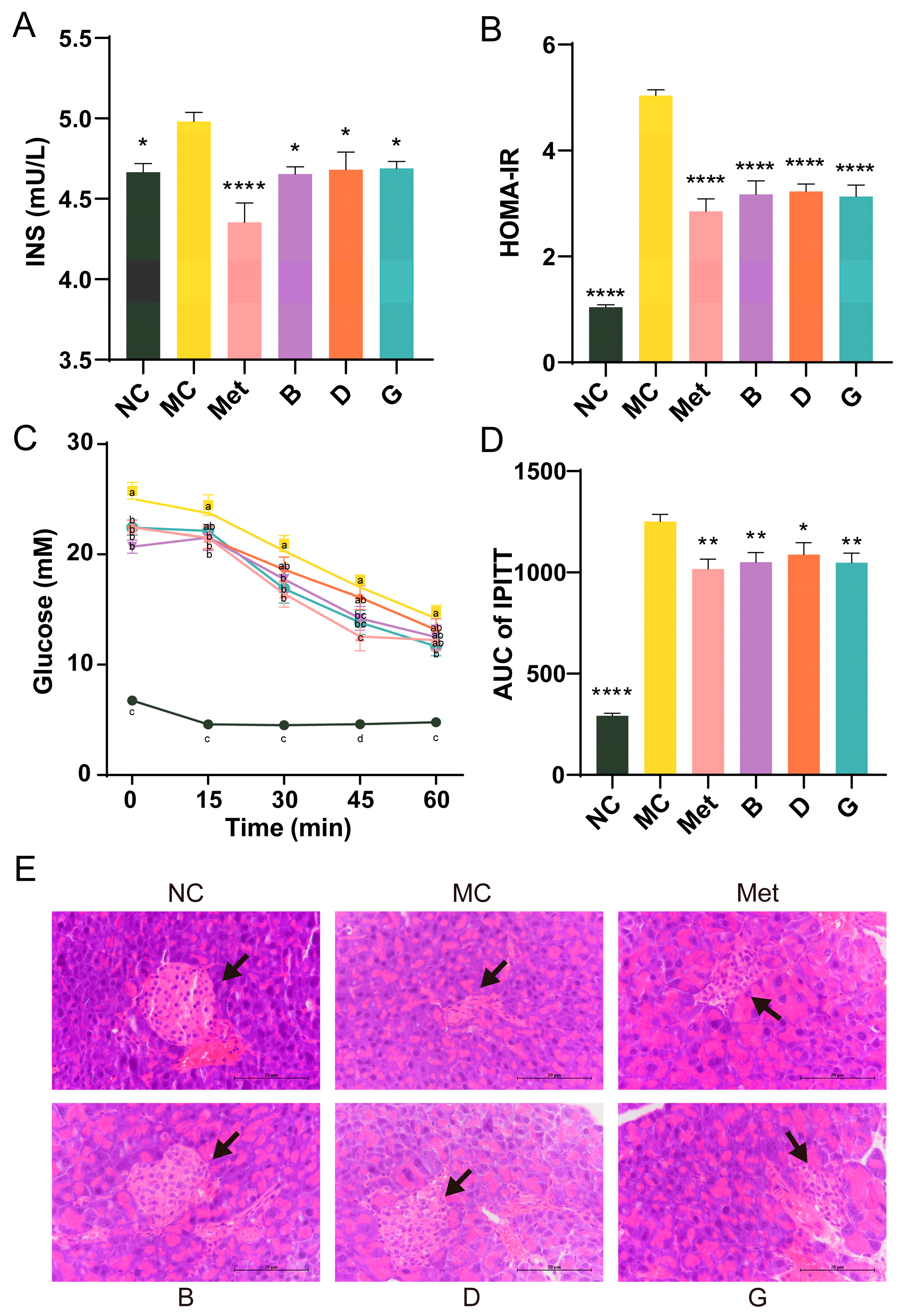
2.8. OGTT and IPITT
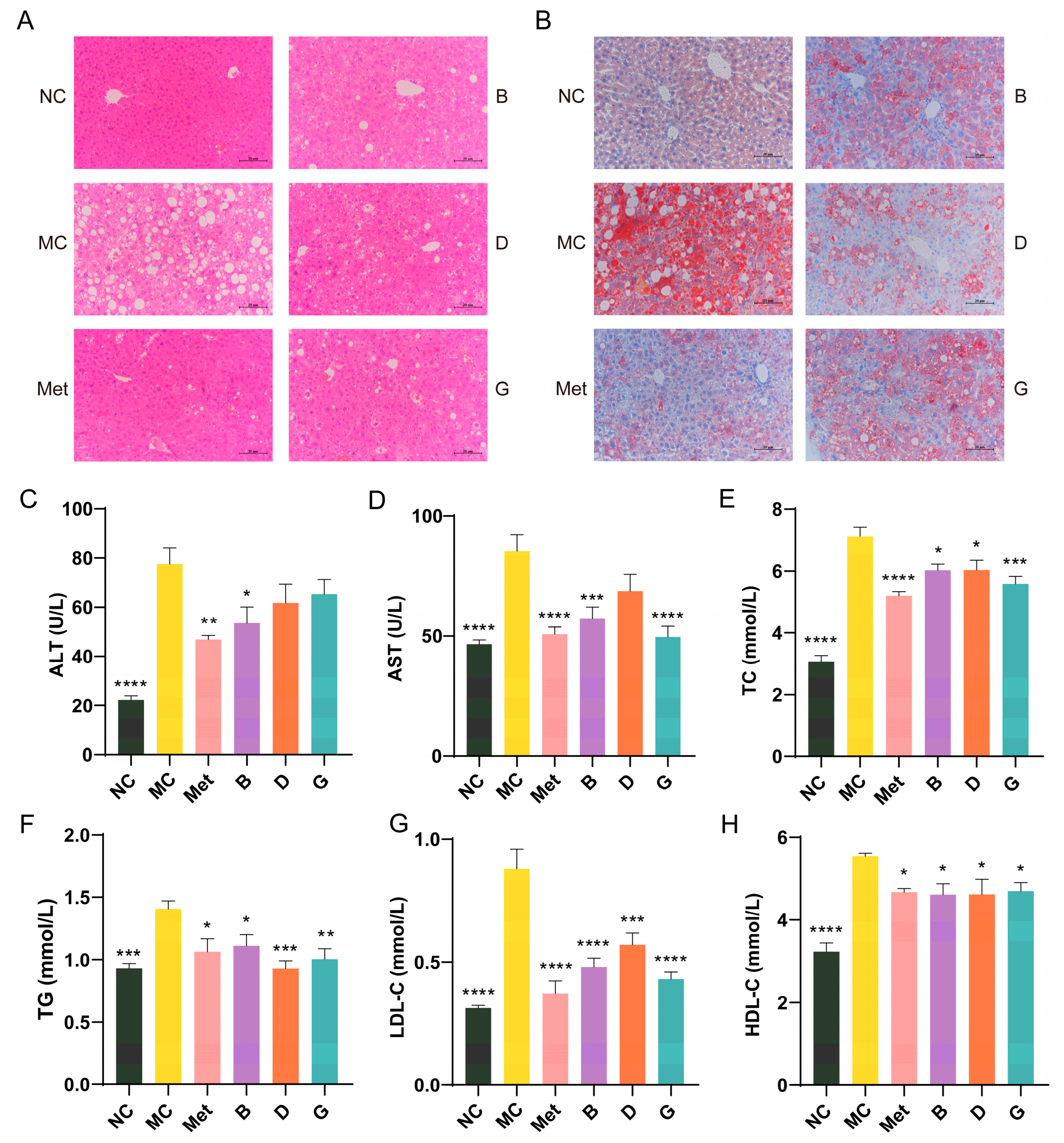
2.9. Serum Biochemical Indexes
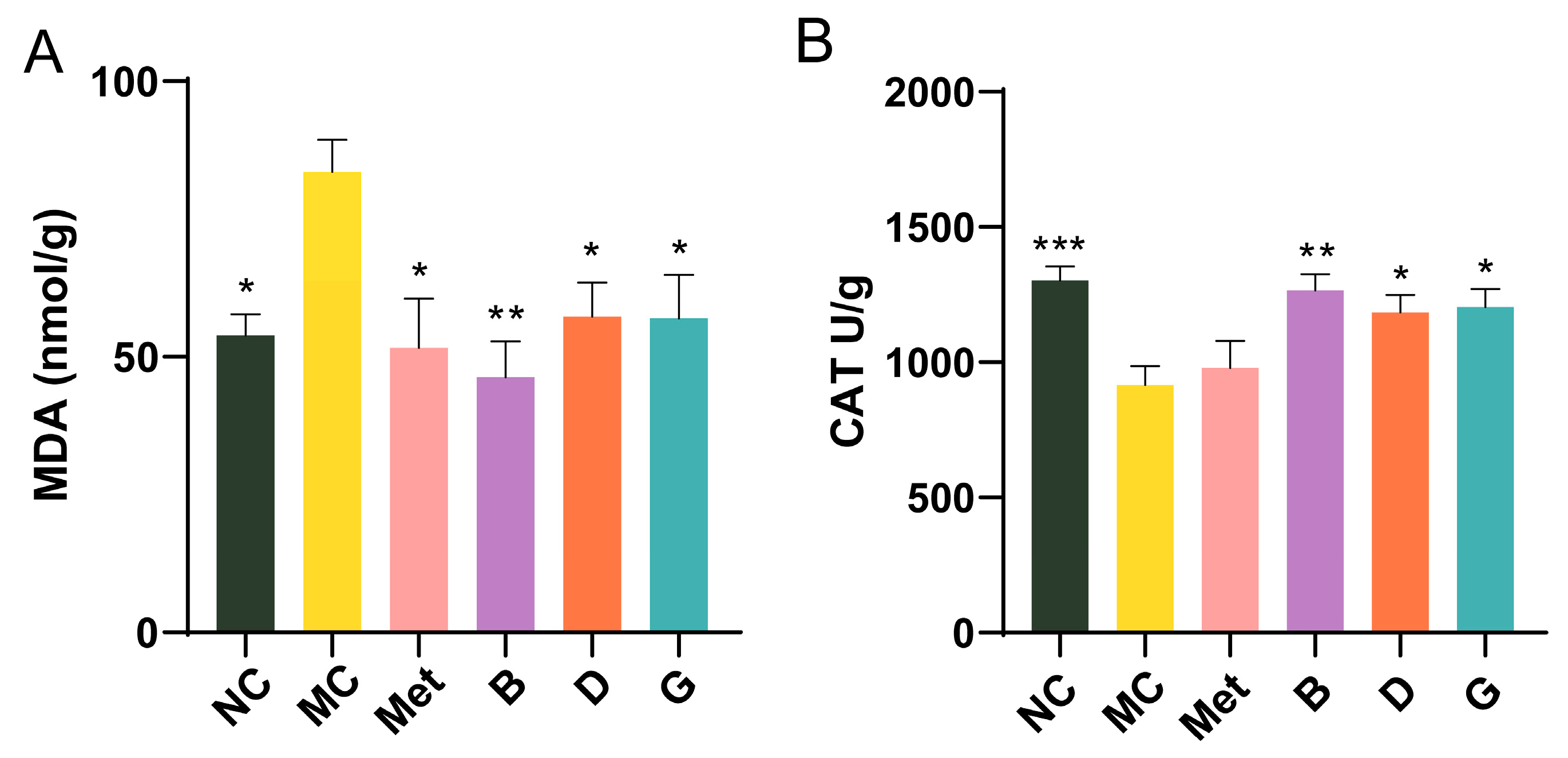
2.10. Oxidative Stress in Liver
2.11. Histopathological Observation of Liver
2.12. Analyses of mRNA Expression
2.13. Western Blot Assay
2.14. Statistics
3. Results
3.1. Quantitation of Polyphenols in BPE
3.2. In Vitro Evaluation of BPE Polyphenols Exhibiting Hypoglycemic Activity
3.3. Effects of BPE on Blood Glucose Levels in Diabetic Mice
3.4. Effect of BPE on Insulin Resistance in Diabetic Mice
3.5. Effects of BPE on Blood Lipid Levels in Diabetic Mice
3.6. Effect of BPE on the Level of Oxidative Stress in the Liver
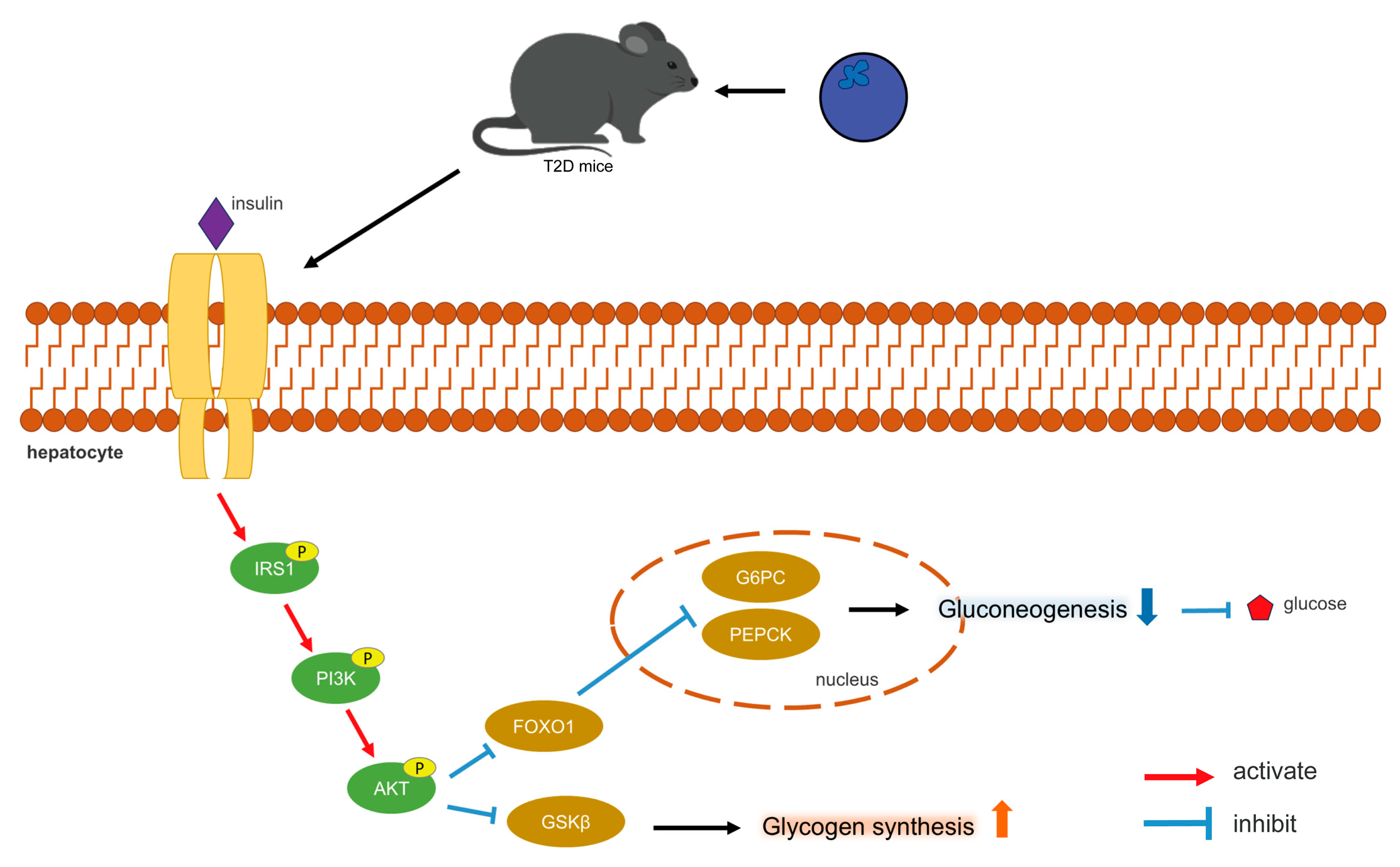
3.7. The Effect of BPE on the Expression of Key Genes of Glucose Metabolism and Lipid Metabolism in Diabetic Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas 10th Edition. 2021. Available online: https://diabetesatlas.org/atlas/tenth-edition/ (accessed on 13 March 2024).
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2021. Diabetes Care 2021, 44, S15–S33. [Google Scholar] [CrossRef] [PubMed]
- Zimmet, P.; Alberti, K.G.M.M.; Shaw, J. Global and Societal Implications of the Diabetes Epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.A.; Creager, M.A. Vascular Complications of Diabetes. Circ. Res. 2016, 118, 1771–1785. [Google Scholar] [CrossRef] [PubMed]
- Sattar, N.; McGuire, D.K. Pathways to Cardiorenal Complications in Type 2 Diabetes Mellitus: A Need to Rethink. Circulation 2018, 138, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Demir, S.; Nawroth, P.P.; Herzig, S.; Ekim Üstünel, B. Emerging Targets in Type 2 Diabetes and Diabetic Complications. Adv. Sci. 2021, 8, 2100275. [Google Scholar] [CrossRef]
- Tahrani, A.A.; Barnett, A.H.; Bailey, C.J. Pharmacology and Therapeutic Implications of Current Drugs for Type 2 Diabetes Mellitus. Nat. Rev. Endocrinol. 2016, 12, 566–592. [Google Scholar] [CrossRef]
- Upadhyay, J.; Polyzos, S.A.; Perakakis, N.; Thakkar, B.; Paschou, S.A.; Katsiki, N.; Underwood, P.; Park, K.H.; Seufert, J.; Kang, E.S.; et al. Pharmacotherapy of Type 2 Diabetes: An Update. Metabolism 2018, 78, 13–42. [Google Scholar] [CrossRef]
- Grunberger, G. Should Side Effects Influence the Selection of Antidiabetic Therapies in Type 2 Diabetes? Curr. Diab. Rep. 2017, 17, 21. [Google Scholar] [CrossRef]
- DeMarsilis, A.; Reddy, N.; Boutari, C.; Filippaios, A.; Sternthal, E.; Katsiki, N.; Mantzoros, C. Pharmacotherapy of Type 2 Diabetes: An Update and Future Directions. Metabolism 2022, 137, 155332. [Google Scholar] [CrossRef]
- Zarich, S.W. Antidiabetic Agents and Cardiovascular Risk in Type 2 Diabetes. Nat. Rev. Endocrinol. 2009, 5, 500–506. [Google Scholar] [CrossRef]
- Wang, H.; Guo, X.; Hu, X.; Li, T.; Fu, X.; Liu, R.H. Comparison of Phytochemical Profiles, Antioxidant and Cellular Antioxidant Activities of Different Varieties of Blueberry (Vaccinium Spp.). Food Chem. 2017, 217, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Dróżdż, P.; Šėžienė, V.; Pyrzynska, K. Phytochemical Properties and Antioxidant Activities of Extracts from Wild Blueberries and Lingonberries. Plant Foods Hum. Nutr. 2017, 72, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Sobolev, A.P.; Ciampa, A.; Ingallina, C.; Mannina, L.; Capitani, D.; Ernesti, I.; Maggi, E.; Businaro, R.; Del Ben, M.; Engel, P.; et al. Blueberry-Based Meals for Obese Patients with Metabolic Syndrome: A Multidisciplinary Metabolomic Pilot Study. Metabolites 2019, 9, 138. [Google Scholar] [CrossRef]
- Duan, Y.; Tarafdar, A.; Chaurasia, D.; Singh, A.; Bhargava, P.C.; Yang, J.; Li, Z.; Ni, X.; Tian, Y.; Li, H.; et al. Blueberry Fruit Valorization and Valuable Constituents: A Review. Int. J. Food Microbiol. 2022, 381, 109890. [Google Scholar] [CrossRef]
- Esfandiar, Z.; Hosseini-Esfahani, F.; Mirmiran, P.; Yuzbashian, E.; Azizi, F. The Association of Dietary Polyphenol Intake with the Risk of Type 2 Diabetes: Tehran Lipid and Glucose Study. Diabetes Metab. Syndr. Obes. 2020, 13, 1643–1652. [Google Scholar] [CrossRef]
- Guo, J.; Han, X.; Tan, H.; Huang, W.; You, Y.; Zhan, J. Blueberry Extract Improves Obesity through Regulation of the Gut Microbiota and Bile Acids via Pathways Involving FXR and TGR5. iScience 2019, 19, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Palma, X.; Thomas-Valdés, S.; Cruz, G. Acute Consumption of Blueberries and Short-Term Blueberry Supplementation Improve Glucose Management and Insulin Levels in Sedentary Subjects. Nutrients 2021, 13, 1458. [Google Scholar] [CrossRef] [PubMed]
- Rocha, D.M.U.P.; Caldas, A.P.S.; da Silva, B.P.; Hermsdorff, H.H.M.; de Cassia Goncalves Alfenas, R. Effects of Blueberry and Cranberry Consumption on Type 2 Diabetes Glycemic Control: A Systematic Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1816–1828. [Google Scholar] [CrossRef]
- Stote, K.S.; Wilson, M.M.; Hallenbeck, D.; Thomas, K.; Rourke, J.M.; Sweeney, M.I.; Gottschall-Pass, K.T.; Gosmanov, A.R. Effect of Blueberry Consumption on Cardiometabolic Health Parameters in Men with Type 2 Diabetes: An 8-Week, Double-Blind, Randomized, Placebo-Controlled Trial. Curr. Dev. Nutr. 2020, 4, nzaa030. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, Y.; Yang, M.; Cao, J.; Khan, A.; Cheng, G. UHPLC-ESI-HRMS/MS Analysis on Phenolic Compositions of Different E Se Tea Extracts and Their Antioxidant and Cytoprotective Activities. Food Chem. 2020, 318, 126512. [Google Scholar] [CrossRef]
- Chen, K.; Chen, X.; Xue, H.; Zhang, P.; Fang, W.; Chen, X.; Ling, W. Coenzyme Q10 Attenuates High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease through Activation of the AMPK Pathway. Food Funct. 2019, 10, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Luo, J.; Chai, Y.; Guo, Y.; Tianzhi, Y.; Bao, Y. Hypoglycemic Effect of Taraxacum Officinale Root Extract and Its Synergism with Radix Astragali Extract. Food Sci. Nutr. 2021, 9, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Kupriyanova, Y.; Zaharia, O.P.; Bobrov, P.; Karusheva, Y.; Burkart, V.; Szendroedi, J.; Hwang, J.H.; Roden, M.; Roden, M.; Al-Hasani, H.; et al. Early Changes in Hepatic Energy Metabolism and Lipid Content in Recent-Onset Type 1 and 2 Diabetes Mellitus. J. Hepatol. 2021, 74, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qian, F.; Zuo, Y.; Yuan, J.; Chen, S.; Wu, S.; Wang, A. U-Shaped Relationship of High-Density Lipoprotein Cholesterol and Incidence of Total, Ischemic and Hemorrhagic Stroke: A Prospective Cohort Study. Stroke 2022, 53, 1624–1632. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Han, G.; Liu, X.; Chen, B.; Peng, K.; Shi, Y.; Zhang, M.; Yang, Y.; Cui, J.; Song, L.; et al. Association of High-Density Lipoprotein Cholesterol with All-Cause and Cause-Specific Mortality in a Chinese Population of 3.3 Million Adults: A Prospective Cohort Study. Lancet Reg. Health West. Pac. 2024, 42, 100874. [Google Scholar] [CrossRef]
- Oliveira, P.S.; Gazal, M.; Flores, N.P.; Zimmer, A.R.; Chaves, V.C.; Reginatto, F.H.; Kaster, M.P.; Tavares, R.G.; Spanevello, R.M.; Lencina, C.L.; et al. Vaccinium Virgatum Fruit Extract as an Important Adjuvant in Biochemical and Behavioral Alterations Observed in Animal Model of Metabolic Syndrome. Biomed. Pharmacother. 2017, 88, 939–947. [Google Scholar] [CrossRef]
- Nunes, S.; Viana, S.D.; Preguiça, I.; Alves, A.; Fernandes, R.; Teodoro, J.S.; Matos, P.; Figueirinha, A.; Salgueiro, L.; André, A.; et al. Blueberry Counteracts Prediabetes in a Hypercaloric Diet-Induced Rat Model and Rescues Hepatic Mitochondrial Bioenergetics. Nutrients 2021, 13, 4192. [Google Scholar] [CrossRef]
- Huang, W.; Yao, L.; He, X.; Wang, L.; Li, M.; Yang, Y.; Wan, C. Hypoglycemic Activity and Constituents Analysis of Blueberry (Vaccinium corymbosum) Fruit Extracts. Diabetes Metab. Syndr. Obes. 2018, 11, 357–366. [Google Scholar] [CrossRef]
- James, D.E.; Stöckli, J.; Birnbaum, M.J. The Aetiology and Molecular Landscape of Insulin Resistance. Nat. Rev. Mol. Cell Biol. 2021, 22, 751–771. [Google Scholar] [CrossRef]
- Nurullina, G.; Valeeva, F. AB0409 Modified Homa-Ir Index in Patients with Rheumatic Diseases Receiving Glucocorticoid Therapy. Ann. Rheum. Dis. 2016, 75, 1046. [Google Scholar] [CrossRef]
- Birkenfeld, A.L.; Shulman, G.I. Nonalcoholic Fatty Liver Disease, Hepatic Insulin Resistance, and Type 2 Diabetes. Hepatology 2014, 59, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Jurado-Aguilar, J.; Wahli, W.; Palomer, X.; Vázquez-Carrera, M. Increased Hepatic Gluconeogenesis and Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. 2024, 35, 1062–1077. [Google Scholar] [CrossRef]
- Liu, S.; Huang, S.; Wu, X.; Feng, Y.; Shen, Y.; Zhao, Q.S.; Leng, Y. Activation of SIK1 by Phanginin A Inhibits Hepatic Gluconeogenesis by Increasing PDE4 Activity and Suppressing the CAMP Signaling Pathway. Mol. Metab. 2020, 41, 101045. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Han, H.S.; Jung, Y.S.; Harris, R.A.; Koo, S.H.; Choi, H.S. The SMILE Transcriptional Corepressor Inhibits CAMP Response Element–Binding Protein (CREB)–Mediated Transactivation of Gluconeogenic Genes. J. Biol. Chem. 2018, 293, 13125–13133. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, M.; Inoue, S.; Horio, F.; Tsuda, T. Dietary Anthocyanin-Rich Bilberry Extract Ameliorates Hyperglycemia and Insulin Sensitivity via Activation of AMP-Activated Protein Kinase in Diabetic Mice. J. Nutr. 2010, 140, 527–533. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Guo, J.; Su, Z.Q. The PI3K/AKT Pathway in Obesity and Type 2 Diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Labra, R.; Subiabre, M.; Toledo, F.; Pardo, F.; Sobrevia, L. Endoplasmic Reticulum Stress and Development of Insulin Resistance in Adipose, Skeletal, Liver, and Foetoplacental Tissue in Diabesity. Mol. Asp. Med. 2019, 66, 49–61. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Yu, Z.L.; Chung, S.K.; Xu, B. The Roles of Dietary Polyphenols at Crosstalk between Type 2 Diabetes and Alzheimer’s Disease in Ameliorating Oxidative Stress and Mitochondrial Dysfunction via PI3K/Akt Signaling Pathways. Ageing Res. Rev. 2024, 99, 102416. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, F.G.; Zhang, W.S.; Pan, A.; Yang, Y.L.; Liu, J.F.; Li, P.; Liu, B.L.; Qi, L.W. Ginsenoside Rg1 Inhibits Glucagon-Induced Hepatic Gluconeogenesis through Akt-FoxO1 Interaction. Theranostics 2017, 7, 4001–4012. [Google Scholar] [CrossRef]
- Tricò, D.; Mengozzi, A.; Baldi, S.; Bizzotto, R.; Olaniru, O.; Toczyska, K.; Huang, G.C.; Seghieri, M.; Frascerra, S.; Amiel, S.A.; et al. Lipid-Induced Glucose Intolerance Is Driven by Impaired Glucose Kinetics and Insulin Metabolism in Healthy Individuals. Metabolism 2022, 134, 155247. [Google Scholar] [CrossRef] [PubMed]
- Xepapadaki, E.; Nikdima, I.; Sagiadinou, E.C.; Zvintzou, E.; Kypreos, K.E. HDL and Type 2 Diabetes: The Chicken or the Egg? Diabetologia 2021, 64, 1917–1926. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xu, L.; Ye, J.; Li, D.; Wang, W.; Li, X.; Wu, L.; Wang, H.; Guan, F.; Li, P. Cidea Promotes Hepatic Steatosis by Sensing Dietary Fatty Acids. Hepatology 2012, 56, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Castro, V.; Calvo, G.; Ávila-Pérez, G.; Dreux, M.; Gastaminza, P. Differential Roles of Lipin1 and Lipin2 in the Hepatitis C Virus Replication Cycle. Cells 2019, 8, 1456. [Google Scholar] [CrossRef]
- Maiese, K. New Insights for Oxidative Stress and Diabetes Mellitus. Oxid. Med. Cell Longev. 2015, 2015, 875961. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative Stress and Diabetic Complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Herrera-Balandrano, D.D.; Chai, Z.; Hutabarat, R.P.; Beta, T.; Feng, J.; Ma, K.; Li, D.; Huang, W. Hypoglycemic and Hypolipidemic Effects of Blueberry Anthocyanins by AMPK Activation: In Vitro and in Vivo Studies. Redox Biol. 2021, 46, 102100. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content (mg/g) |
|---|---|
| Total polyphenols (as gallic acid) | 635.58 ± 11.57 |
| Total flavonoids (as rutin) | 118.97 ± 1.98 |
| Total anthocyanins (as cyanin-3-glucoside) | 246.13 ± 3.80 |
| Compound | Classification | Formula | CAS |
|---|---|---|---|
| Dihydromyricetin | flavanonol | C15 H12 O8 | 27200-12-0 |
| Quercetin | flavonol | C15 H10 O7 | 117-39-5 |
| Rutin | flavonol glycoside | C27 H30 O16 | 153-18-4 |
| Chlorogenic acid | phenolic acid | C16 H18 O9 | 327-97-9 |
| Gallic acid | phenolic acid | C7 H6 O5 | 149-91-7 |
| Gentisic acid | phenolic acid | C7 H6 O4 | 490-79-9 |
| Syringic acid | phenolic acid | C9 H10 O5 | 530-57-4 |
| Methyl gallate | phenolic acid | C8 H8 O5 | 99-24-1 |
| Pyrogallol | phenolic compound | C6 H6 O3 | 87-66-1 |
| Catechol | phenolic compound | C6 H6 O2 | 120-80-9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Y.; Guo, J.; Lin, Y.; You, Y.; Huang, W.; Zhan, J. Evaluation of Hypoglycemic Polyphenolic Compounds in Blueberry Extract: Functional Effects and Mechanisms. Antioxidants 2024, 13, 1490. https://doi.org/10.3390/antiox13121490
Qin Y, Guo J, Lin Y, You Y, Huang W, Zhan J. Evaluation of Hypoglycemic Polyphenolic Compounds in Blueberry Extract: Functional Effects and Mechanisms. Antioxidants. 2024; 13(12):1490. https://doi.org/10.3390/antiox13121490
Chicago/Turabian StyleQin, Yue, Jielong Guo, Yuchen Lin, Yilin You, Weidong Huang, and Jicheng Zhan. 2024. "Evaluation of Hypoglycemic Polyphenolic Compounds in Blueberry Extract: Functional Effects and Mechanisms" Antioxidants 13, no. 12: 1490. https://doi.org/10.3390/antiox13121490
APA StyleQin, Y., Guo, J., Lin, Y., You, Y., Huang, W., & Zhan, J. (2024). Evaluation of Hypoglycemic Polyphenolic Compounds in Blueberry Extract: Functional Effects and Mechanisms. Antioxidants, 13(12), 1490. https://doi.org/10.3390/antiox13121490