
Impact of Chromium Picolinate on Leydig Cell Steroidogenesis and Antioxidant Balance Using an In Vitro Insulin Resistance Model

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture—BLTK1 Murine Leydig Cells
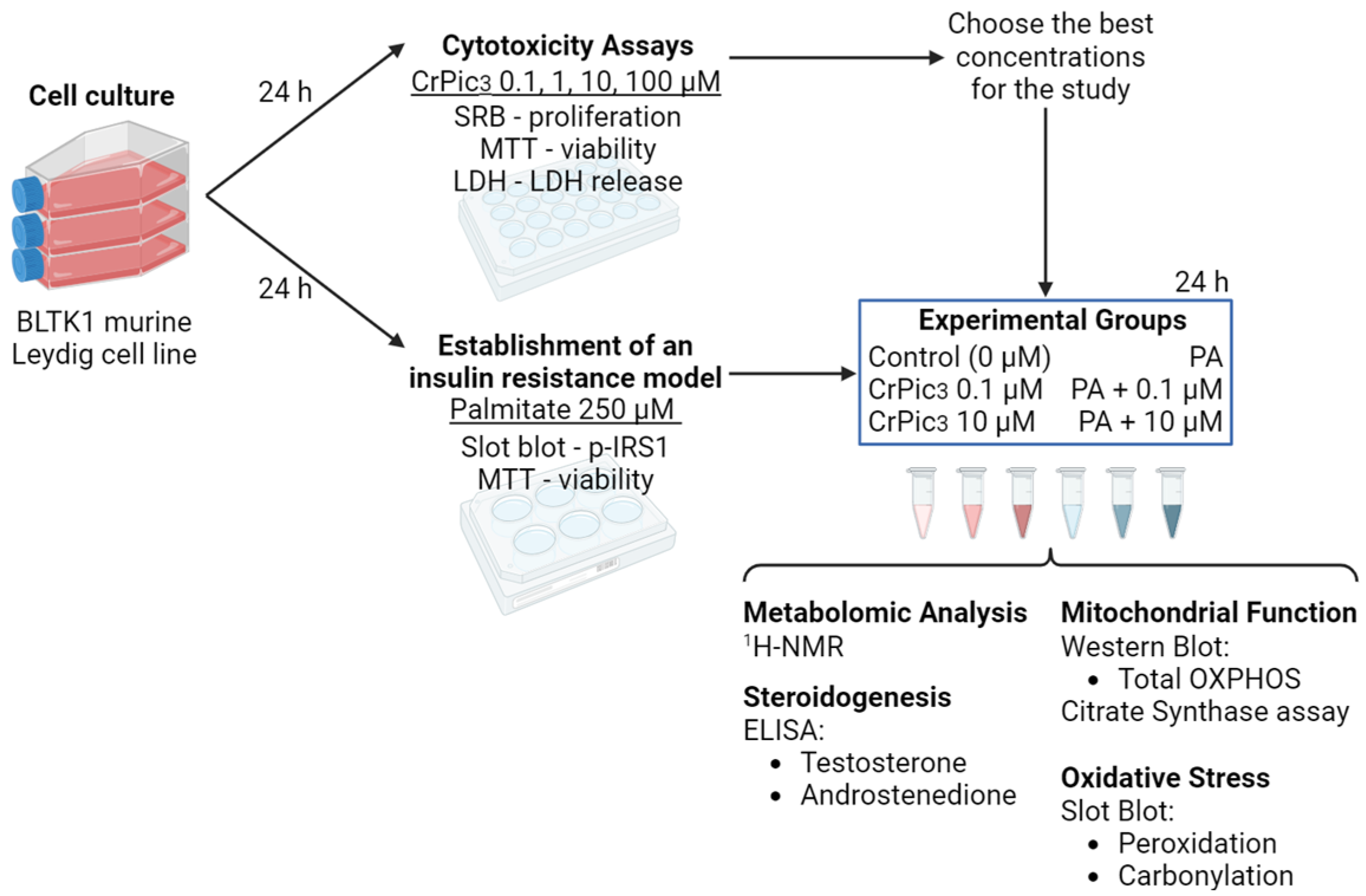
2.3. Experimental Groups and Design
2.4. Evaluation of Cytotoxic Profile of the CrPic3 and PA
2.4.1. Sulforhodamine B Cytotoxicity Assay
2.4.2. Lactate Dehydrogenase Release Assay
2.4.3. 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide Assay
2.5. Exometabolome Analysis via 1H-NMR Spectroscopy
2.6. Protein Extraction and Quantification
2.7. Western Blot
2.8. Slot-Blot
2.9. Citrate Synthase Activity Assay
2.10. Competitive ELISA
2.11. Statistical Analysis
3. Results
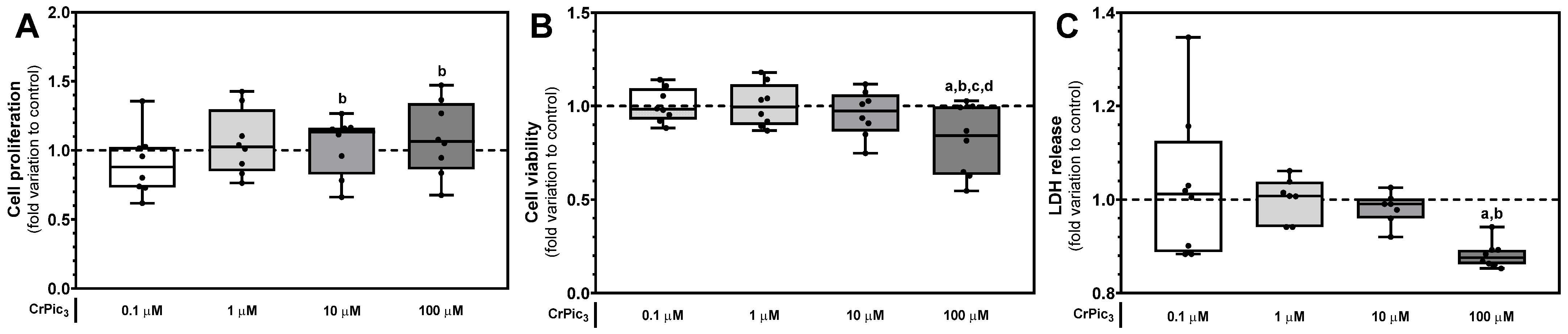
3.1. High Doses of CrPic3 Are Cytotoxic, While Lower Doses Do Not Cause Significant Damage to LCs
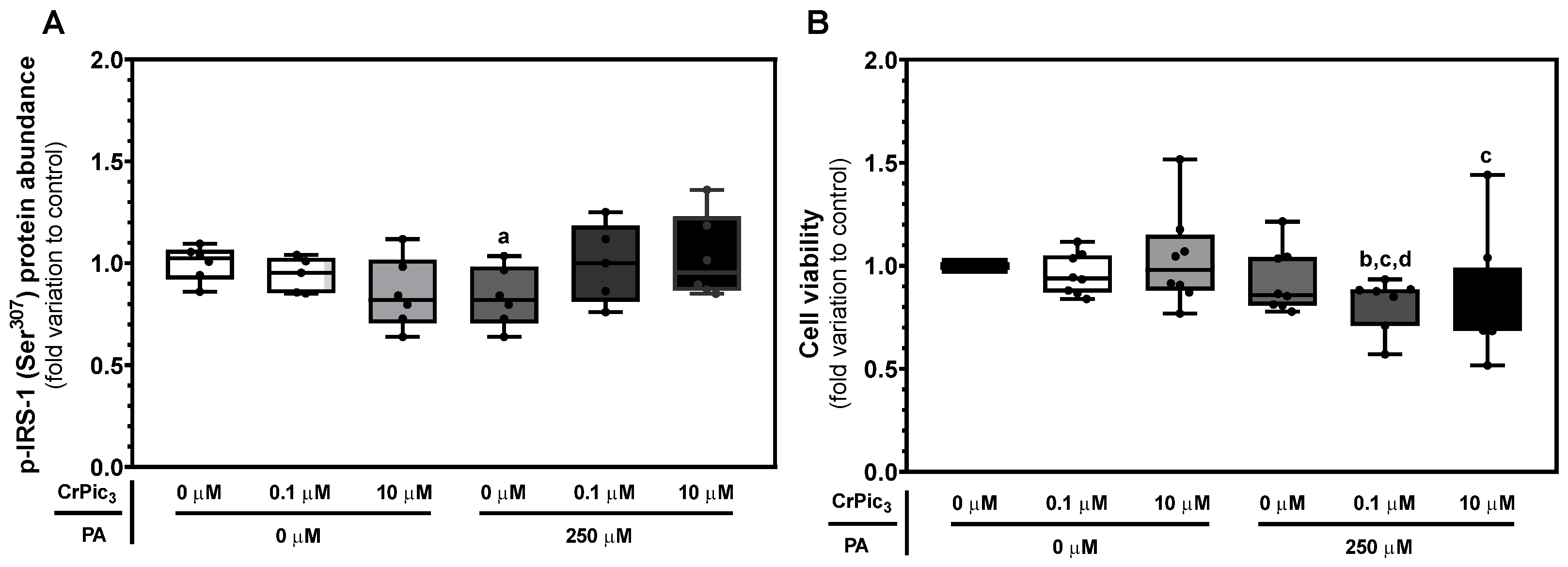
3.2. An In Vitro Model of Insulin Resistance Was Successfully Established in LCs, and CrPic3 Was Not Able to Revert This Condition
3.3. Exometabolome Profiling of LC Exposed to CrPic3 and PA
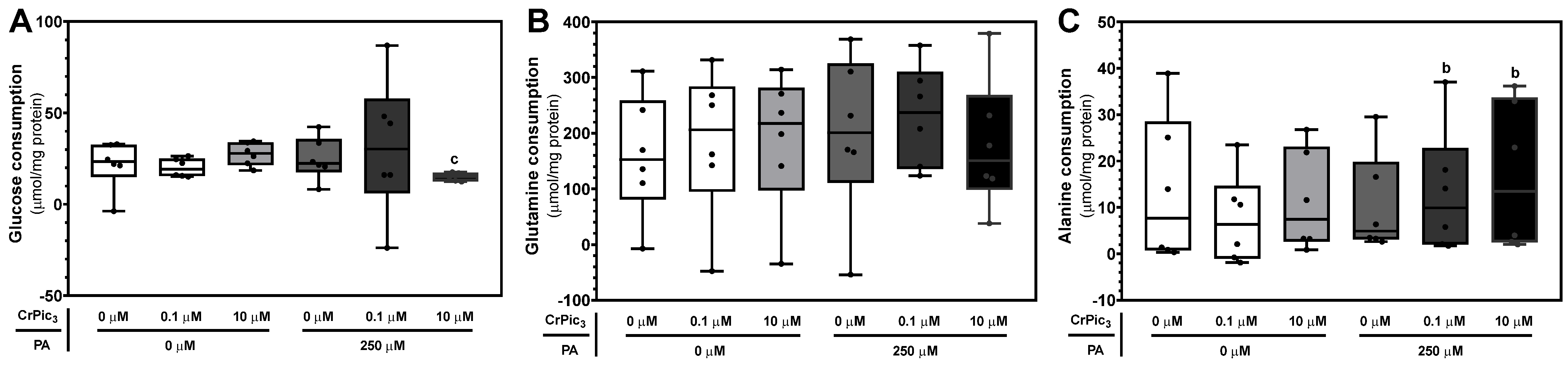
3.3.1. CrPic3 Exposure Did Not Alter Glucose or Glutamine Consumption in LCs under IR Conditions
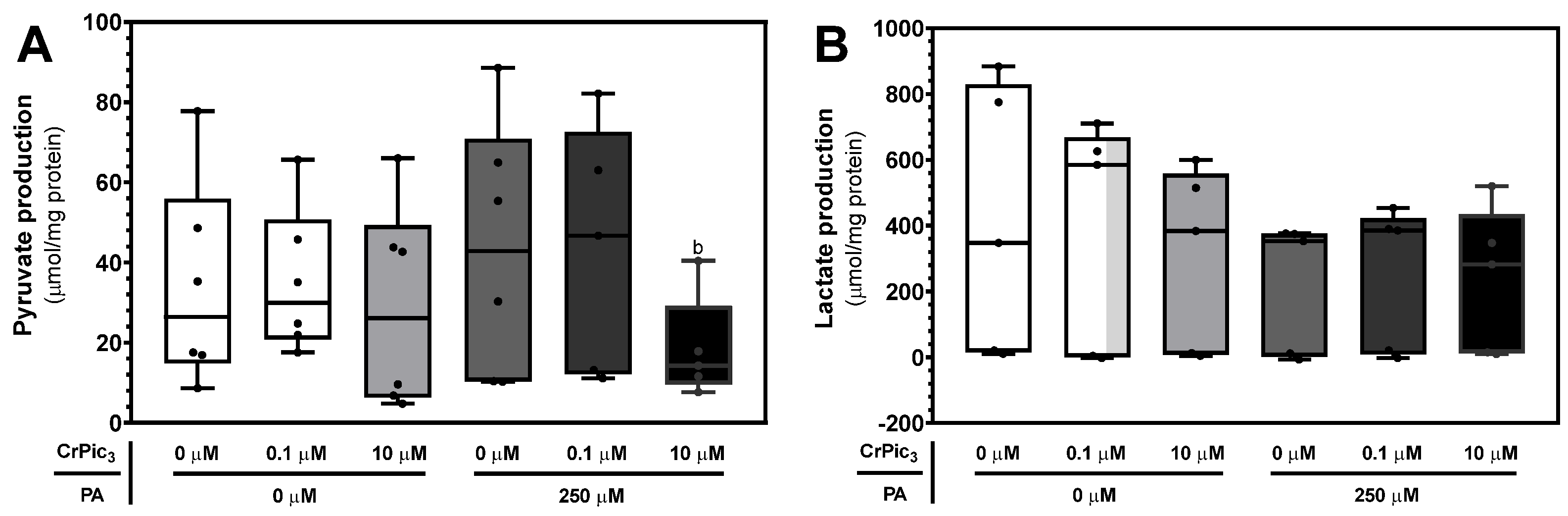
3.3.2. CrPic3 Did Not Alter the Production of Pyruvate or Lactate by LCs Even in IR Conditions
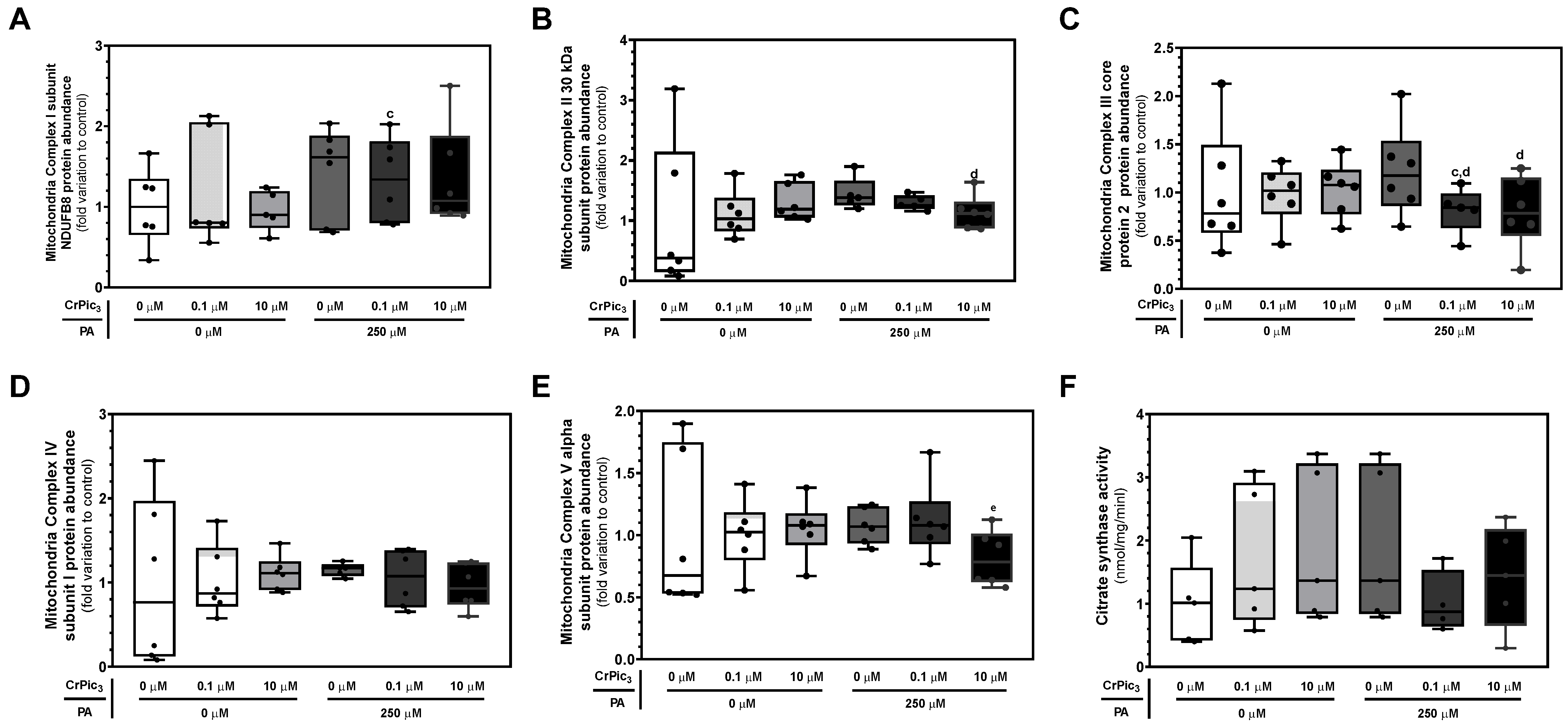
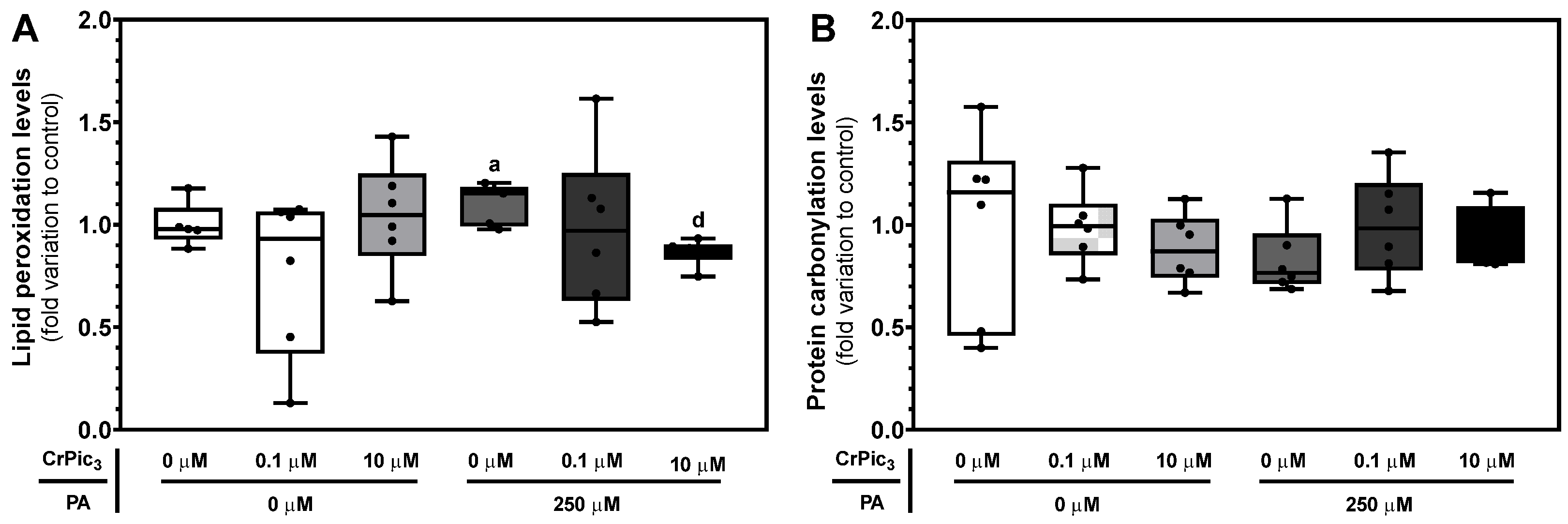
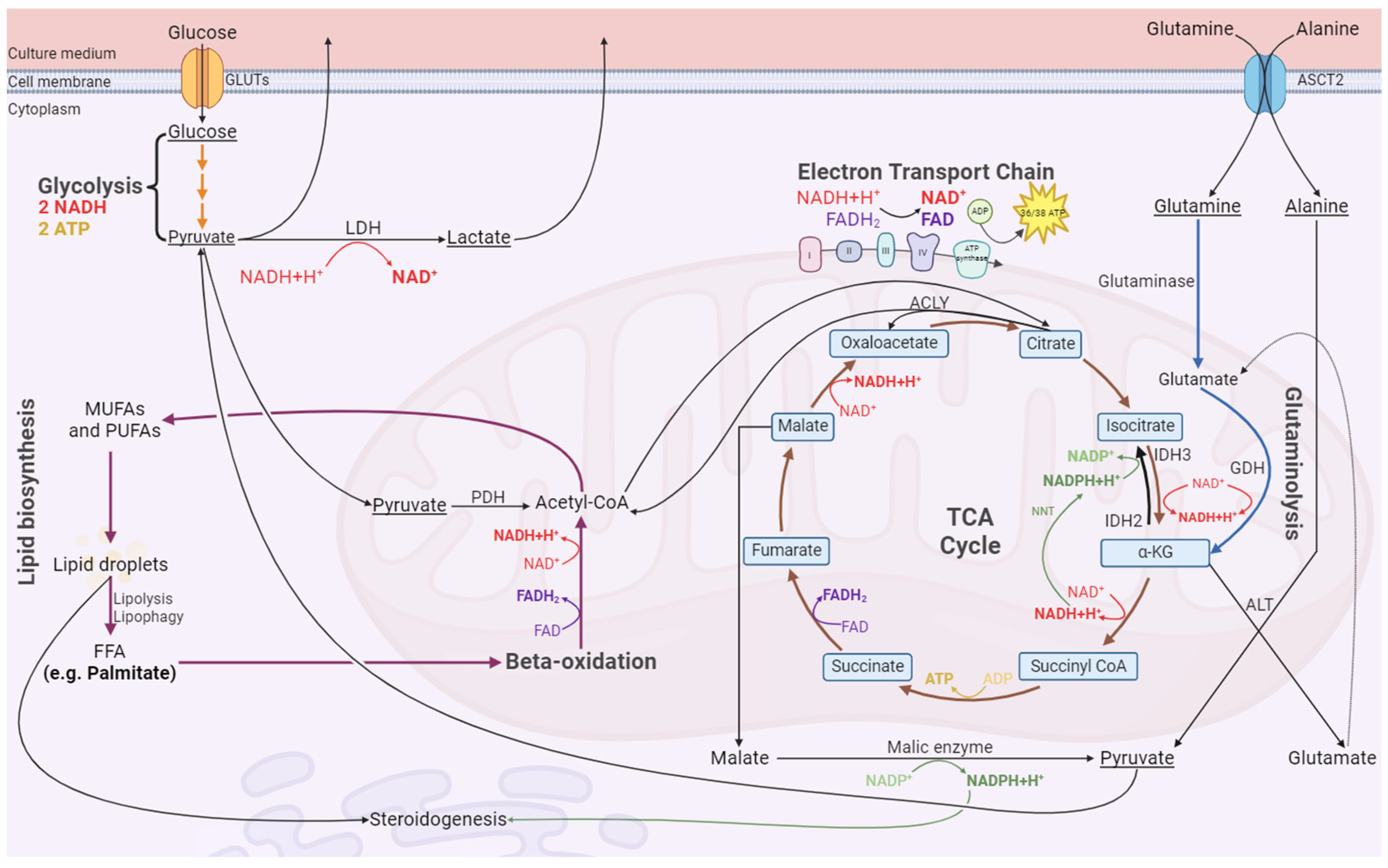
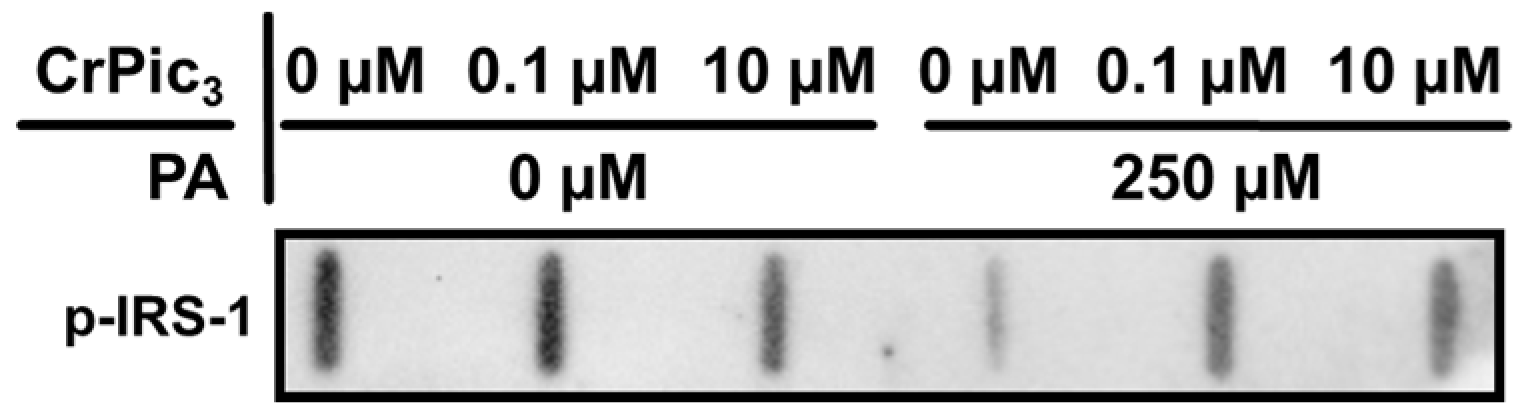
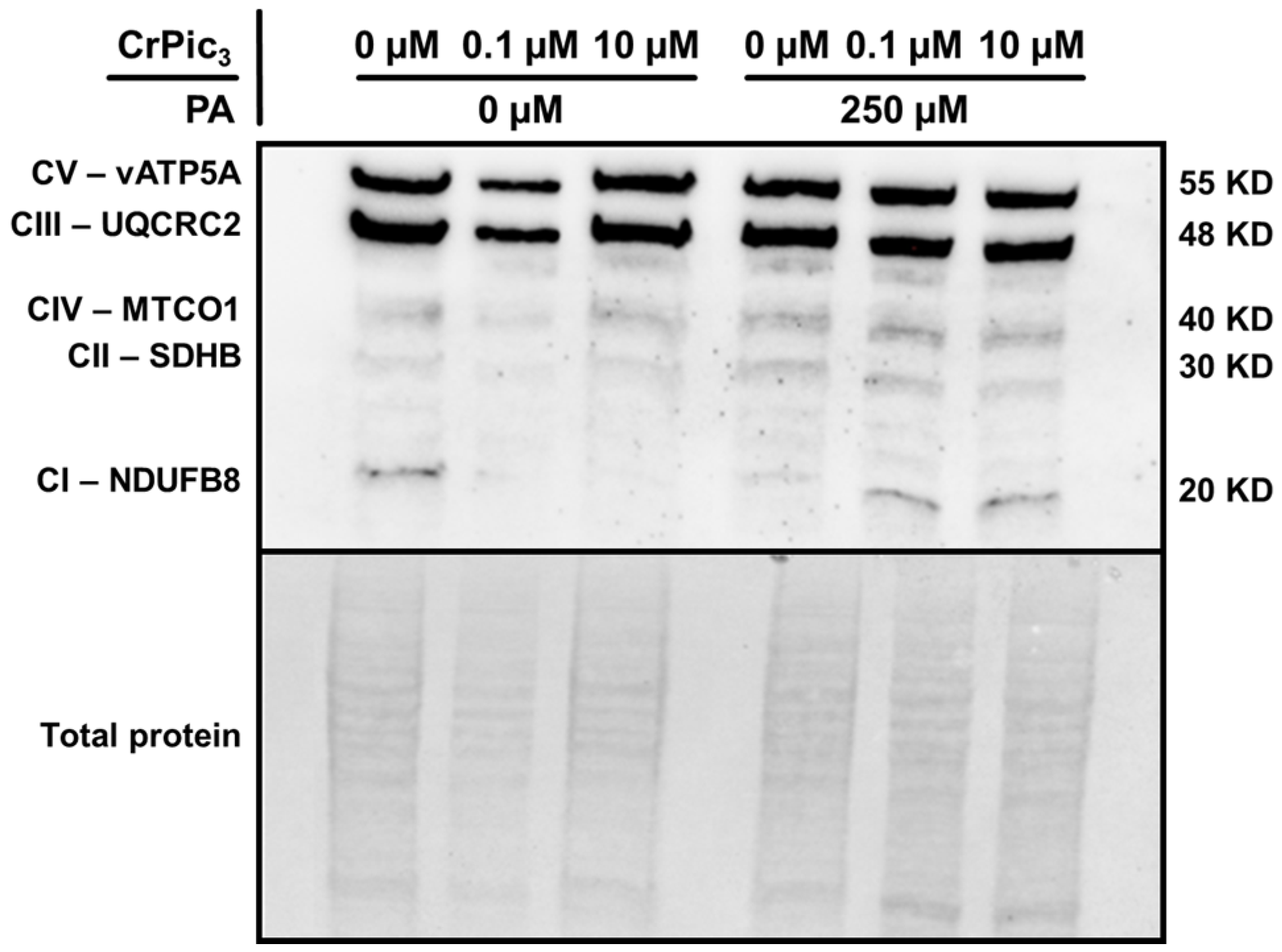
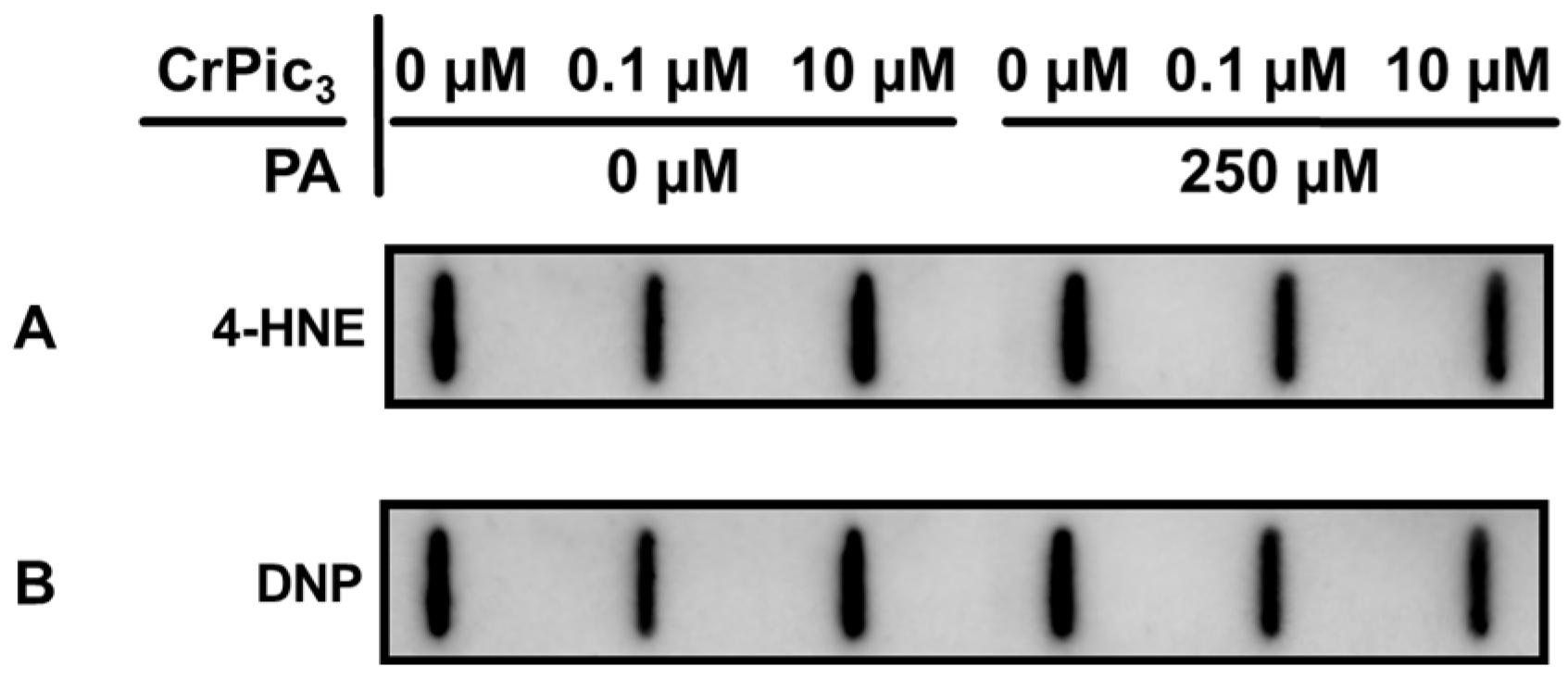
3.4. Exposure of LCs to CrPic3 Reduces the Expression of Mitochondrial Complexes Associated with Oxidative Stress and Reduces Oxidative Damage to Lipids Caused by IR
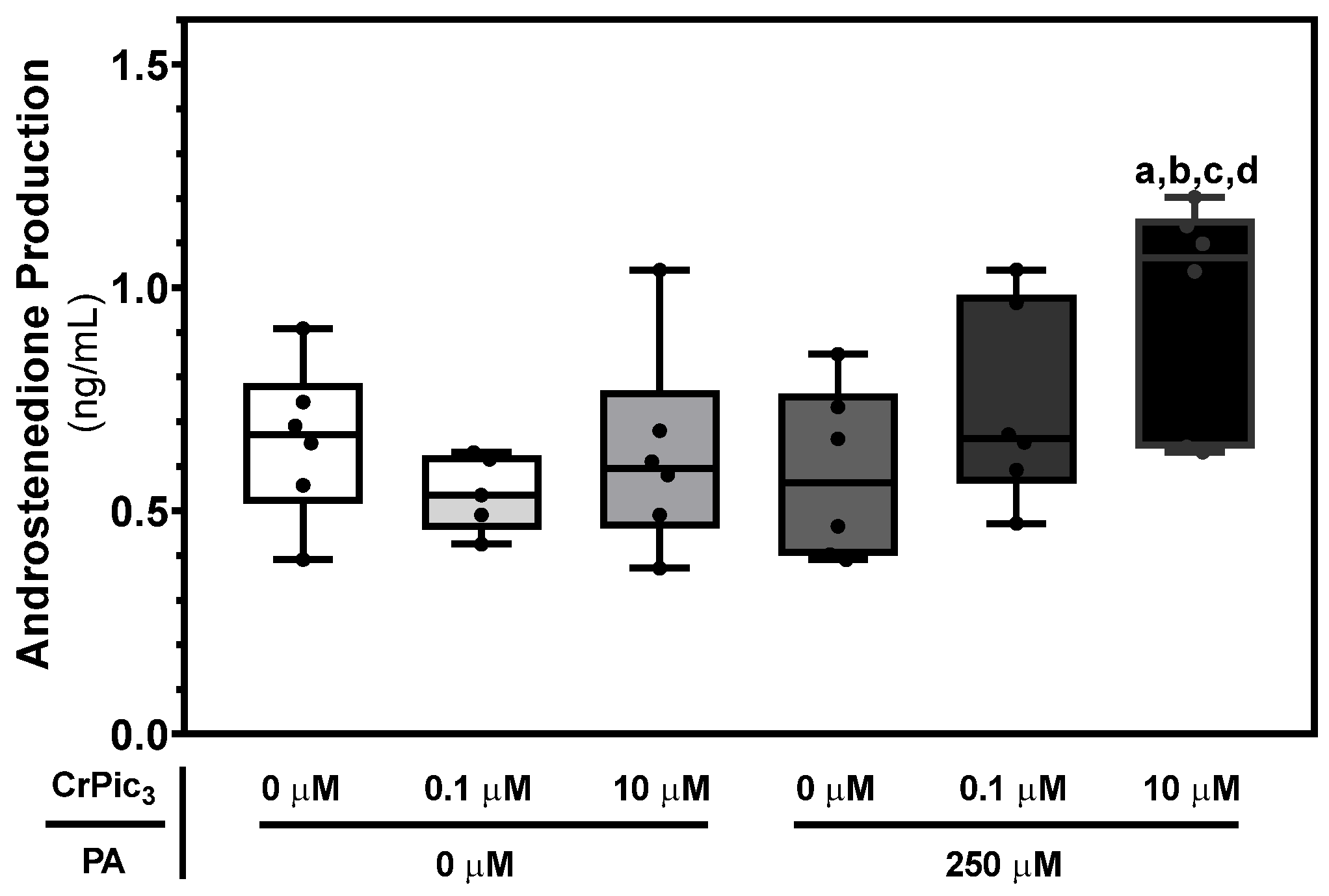
3.5. LC Exposure to CrPic3 Only Increases Androstenedione Production in the Presence of PA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Immunoblots



References
- Zegers-Hochschild, F.; Adamson, G.D.; de Mouzon, J.; Ishihara, O.; Mansour, R.; Nygren, K.; Sullivan, E.; Vanderpoel, S.; International Committee for Monitoring Assisted Reproductive Technology; World Health Organization. International Committee for Monitoring Assisted Reproductive Technology (ICMART) and the World Health Organization (WHO) revised glossary of ART terminology, 2009. Fertil. Steril. 2009, 92, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- Sharlip, I.D.; Jarow, J.P.; Belker, A.M.; Lipshultz, L.I.; Sigman, M.; Thomas, A.J.; Schlegel, P.N.; Howards, S.S.; Nehra, A.; Damewood, M.D.; et al. Best practice policies for male infertility. Fertil. Steril. 2002, 77, 873–882. [Google Scholar] [CrossRef]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, X.; Qi, K.; Wang, T.; Qi, Y.; Cui, L.; Wang, F.; Song, M. Effects of environmental contaminants on fertility and reproductive health. Int. J. Environ. Sci. Technol. 2019, 77, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Terekeci, H.; Sandal, S.; Kelestimur, F. Endocrine disrupting chemicals: Exposure, effects on human health, mechanism of action, models for testing and strategies for prevention. Rev. Endocr. Metab. Disord. 2020, 21, 127–147. [Google Scholar] [CrossRef] [PubMed]
- Ohlander, S.J.; Lindgren, M.C.; Lipshultz, L.I. Testosterone and Male Infertility. Urol. Clin. N. Am. 2016, 43, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Flück, C.E.; Pandey, A.V. Testicular Steroidogenesis. In Endocrinology of the Testis and Male Reproduction; Springer: Berlin, Germany, 2017; pp. 343–371. [Google Scholar] [CrossRef]
- Huhtaniemi, I.; Teerds, K. Leydig Cells. In Encyclopedia of Reproduction, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 1, pp. 30–38. [Google Scholar]
- Badr, F.M.; El-Habit, O. Heavy Metal Toxicity Affecting Fertility and Reproduction of Males. In Bioenvironmental Issues Affecting Men’s Reproductive and Sexual Health; Academic Press: Cambridge, MA, USA, 2018; Volume 35, pp. 293–304. [Google Scholar]
- Genchi, G.; Lauria, G.; Catalano, A.; Carocci, A.; Sinicropi, M.S. The Double Face of Metals: The Intriguing Case of Chromium. Appl. Sci. 2021, 11, 638. [Google Scholar] [CrossRef]
- Wu, Y.H.; Lin, J.C.; Wang, T.Y.; Lin, T.J.; Yen, M.C.; Liu, Y.H.; Wu, P.L.; Chen, F.W.; Shih, Y.L.; Yeh, I.J. Hexavalent chromium intoxication induces intrinsic and extrinsic apoptosis in human renal cells. Mol. Med. Rep. 2020, 21, 851–857. [Google Scholar] [CrossRef]
- Handa, K.; Jindal, R. Estimating the hepatotoxic impact of hexavalent chromium on Ctenopharyngodon idellus through a multi-biomarker study. Environ. Adv. 2021, 5, 100108. [Google Scholar] [CrossRef]
- Yatera, K.; Morimoto, Y.; Ueno, S.; Noguchi, S.; Kawaguchi, T.; Tanaka, F.; Suzuki, H.; Higashi, T. Cancer Risks of Hexavalent Chromium in the Respiratory Tract. J. UOEH 2018, 40, 157–172. [Google Scholar] [CrossRef]
- Chandra, A.K.; Chatterjee, A.; Ghosh, R.; Sarkar, M.; Chaube, S.K. Chromium induced testicular impairment in relation to adrenocortical activities in adult albino rats. Reprod. Toxicol. 2007, 24, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Abbas, T.; Khawaja, A.; Qaisar, F.; Sajjad, A. Effects of chromium on testes and protective role of mulberry. J. Sheikh Zayed Med. Coll. 2017, 8, 1200–1204. [Google Scholar]
- Chandra, A.K.; Chatterjee, A.; Ghosh, R.; Sarkar, M. Vitamin E-supplementation protect chromium (VI)-induced spermatogenic and steroidogenic disorders in testicular tissues of rats. Food Chem. Toxicol. 2010, 48, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Pure Earth. World’s Worst Pollution Problems: The New Top Six Toxic Threats: A Priority List for Remediation; Pure Earth: New York, NY, USA, 2015. [Google Scholar]
- Vincent, J.B. Chromium: Celebrating 50 years as an essential element? Dalton Trans. 2010, 39, 3787–3794. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Chromium Supplementation in Human Health, Metabolic Syndrome, and Diabetes; De Gruyter: Berlin, Germany, 2019; Volume 19, pp. 231–251. [Google Scholar]
- Martin, J.; Wang, Z.Q.; Zhang, X.H.; Wachtel, D.; Volaufova, J.; Matthews, D.E.; Cefalu, W.T. Chromium Picolinate Supplementation Attenuates Body Weight Gain and Increases Insulin Sensitivity in Subjects With Type 2 Diabetes. Diabetes Care 2006, 29, 1826–1832. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Komorowski, J.R.; Tuzcu, M.; Sahin, N.; Juturu, V.; Orhan, C.; Ulas, M.; Sahin, K. Chromium picolinate modulates serotonergic properties and carbohydrate metabolism in a rat model of diabetes. Biol. Trace Elem. Res. 2012, 149, 50–56. [Google Scholar] [CrossRef] [PubMed]
- McLeod, M.; Golden, R. Chromium treatment of depression. Int. J. Neuropsychopharmacol. 2000, 3, 311–314. [Google Scholar] [CrossRef]
- Sahin, K.; Tuzcu, M.; Orhan, C.; Sahin, N.; Kucuk, O.; Ozercan, I.H.; Juturu, V.; Komorowski, J.R. Anti-diabetic activity of chromium picolinate and biotin in rats with type 2 diabetes induced by high-fat diet and streptozotocin. Br. J. Nutr. 2013, 110, 197–205. [Google Scholar] [CrossRef]
- Sundaram, B.; Aggarwal, A.; Sandhir, R. Chromium picolinate attenuates hyperglycemia-induced oxidative stress in streptozotocin-induced diabetic rats. J. Trace Elem. Med. Biol. 2013, 27, 117–121. [Google Scholar] [CrossRef]
- Doddigarla, Z.; Parwez, I.; Abidi, S.; Ahmad, J. Effect of Chromium Picolinate and Melatonin either in Single or in a Combination in Alloxan Induced Male Wistar Rats. J. Biomed. Sci. 2017, 06, 100051. [Google Scholar] [CrossRef]
- Al-Bishri, W.M. Attenuating impacts of chromium and nano resveratrol against hyperglycemia induced oxidative stress in diabetic rats. Int. J. Pharm. Res. Allied Sci. 2017, 6, 61–69. [Google Scholar]
- Kolahian, S.; Sadri, H.; Shahbazfar, A.A.; Amani, M.; Mazadeh, A.; Mirani, M. The Effects of Leucine, Zinc, and Chromium Supplements on Inflammatory Events of the Respiratory System in Type 2 Diabetic Rats. PLoS ONE 2015, 10, e0133374. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, Z.M.; Lugo, J.P. Impact of chromium dinicocysteinate supplementation on inflammation, oxidative stress, and insulin resistance in type 2 diabetic subjects: An exploratory analysis of a randomized, double-blind, placebo-controlled study. Food Nutr. Res. 2016, 60, 31762. [Google Scholar] [CrossRef] [PubMed]
- Rehab, M.E.; Ashraf, M.E. The protective effect of Panax ginseng against chromium picolonate induced testicular changes. Afr. J. Pharm. Pharmacol. 2014, 8, 346–355. [Google Scholar] [CrossRef]
- Zakaria, A.D.; Fayed, A.H.; Hedaya, S.A.; Gad, S.B.; Hafez, M.H. Effect of chromium Picolinate on some reproductive aspects in male rats. Alex. J. Vet. Sci. 2011, 34, 113–124. [Google Scholar]
- Ezzat, W. Effect of supplementing diet with sodium bentonite and/or organic chromium on productive, physiological performance and immune response of matrouh chickens strain. 1- during growth period. Egypt. Poult. Sci. J. 2016, 36, 841–857. [Google Scholar] [CrossRef]
- Mehrim, A.I. Physiological, biochemical and histometric responses of Nile tilapia (Oreochromis niloticus L.) by dietary organic chromium (chromium picolinate) supplementation. J. Adv. Res. 2014, 5, 303–310. [Google Scholar] [CrossRef]
- Dallago, B.S.L.; Braz, S.; Marçola, T.G.; McManus, C.; Caldeira, D.F.; Campeche, A.; Gomes, E.F.; Paim, T.P.; Borges, B.O.; Louvandini, H. Blood Parameters and Toxicity of Chromium Picolinate Oral Supplementation in Lambs. Biol. Trace Elem. Res. 2015, 168, 91–102. [Google Scholar] [CrossRef]
- Moreira, R.; Martins, A.D.; Alves, M.G.; de Lourdes Pereira, M.; Oliveira, P.F. A Comprehensive Review of the Impact of Chromium Picolinate on Testicular Steroidogenesis and Antioxidant Balance. Antioxidants 2023, 12, 1572. [Google Scholar] [CrossRef]
- Kananen, K.; Markkula, M.; El-Hefnawy, T.; Zhang, F.-P.; Paukku, T.; Su, J.-G.J.; Hsueh, A.J.; Huhtaniemi, I. The mouse inhibin α-subunit promoter directs SV40 T-antigen to Leydig cells in transgenic mice. Mol. Cell. Endocrinol. 1996, 119, 135–146. [Google Scholar] [CrossRef]
- Rahman, N.A.; Huhtaniemi, I.T. Testicular cell lines. Mol. Cell. Endocrinol. 2004, 228, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Engeli, R.T.; Fürstenberger, C.; Kratschmar, D.V.; Odermatt, A. Currently available murine Leydig cell lines can be applied to study early steps of steroidogenesis but not testosterone synthesis. Heliyon 2018, 4, e00527. [Google Scholar] [CrossRef] [PubMed]
- Forgacs, A.L.; Ding, Q.; Jaremba, R.G.; Huhtaniemi, I.T.; Rahman, N.A.; Zacharewski, T.R. BLTK1 Murine Leydig Cells: A Novel Steroidogenic Model for Evaluating the Effects of Reproductive and Developmental Toxicants. Toxicol. Sci. 2012, 127, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Liu, Y.; Lu, N.; Li, Y.; Sun, C.-H. cis-9, trans-11-Conjugated linoleic acid activates AMP-activated protein kinase in attenuation of insulin resistance in C2C12 myotubes. J. Agric. Food Chem. 2009, 57, 4452–4458. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.G.; Oliveira, P.F.; Martins, F.O.; Oliveira, P.J.; Carvalho, R.A. Gender-dependent metabolic remodeling during heart preservation in cardioplegic celsior and histidine buffer solution. J. Cardiovasc. Pharmacol. 2012, 59, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Maloy, A.; Alexander, S.; Andreas, A.; Nyunoya, T.; Chandra, D. Stain-Free total-protein normalization enhances the reproducibility of Western blot data. Anal. Biochem. 2022, 654, 114840. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.R.; Alves, M.G.; Bernardino, R.L.; Martins, A.D.; Moreira, A.C.; Silva, J.; Barros, A.; Sousa, M.; Silva, B.M.; Oliveira, P.F. Dose-dependent effects of caffeine in human Sertoli cells metabolism and oxidative profile: Relevance for male fertility. Toxicology 2015, 328, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Coore, H.; Denton, R.; Martin, B.; Randle, P. Regulation of adipose tissue pyruvate dehydrogenase by insulin and other hormones. Biochem. J. 1971, 125, 115–127. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 6th ed.; W. H. Freeman: New York, NY, USA, 2013. [Google Scholar]
- Idell-Wenger, J.A.; Grotyohann, L.W.; Neely, J.R. Coenzyme A and carnitine distribution in normal and ischemic hearts. J. Biol. Chem. 1978, 253, 4310–4318. [Google Scholar] [CrossRef]
- Haider, S.G. Cell Biology of Leydig Cells in the Testis; Elsevier: Amsterdam, The Netherlands, 2004; Volume 233. [Google Scholar]
- Schwarz, K.; Mertz, W. Chromium(III) and the glucose tolerance factor. Arch. Biochem. Biophys. 1959, 85, 292–295. [Google Scholar] [CrossRef]
- Stearns, D.M. Is chromium a trace essential metal? Biofactors 2000, 11, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Casalegno, C.; Schifanella, O.; Zennaro, E.; Marroncelli, S.; Briant, R. Collate Literature Data on Toxicity of Chromium (Cr) and Nickel (Ni) in Experimental Animals and Humans; Wiley Online Library: Hoboken, NJ, USA, 2015; Volume 12, pp. 1–287. [Google Scholar]
- Sarabhai, T.; Koliaki, C.; Mastrototaro, L.; Kahl, S.; Pesta, D.; Apostolopoulou, M.; Wolkersdorfer, M.; Bönner, A.C.; Bobrov, P.; Markgraf, D.F.; et al. Dietary palmitate and oleate differently modulate insulin sensitivity in human skeletal muscle. Diabetologia 2022, 65, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Braga, P.C.; Bernardino, R.L.; Guerra-Carvalho, B.; Carrageta, D.F.; Oliveira, P.F.; Rodrigues, A.S.; Alves, M.G. The progression from mild to severe hyperglycemia coupled with insulin resistance causes mitochondrial dysfunction and alters the metabolic secretome of epithelial kidney cells. Exp. Cell Res. 2023, 431, 113744. [Google Scholar] [CrossRef] [PubMed]
- Rui, L.; Aguirre, V.; Kim, J.K.; Shulman, G.I.; Lee, A.; Corbould, A.; Dunaif, A.; White, M.F. Insulin/IGF-1 and TNF-alpha stimulate phosphorylation of IRS-1 at inhibitory Ser307 via distinct pathways. J. Clin. Investig. 2001, 107, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Hwang, D.; Bataille, F.; Lefevre, M.; York, D.; Quon, M.J.; Ye, J. Serine phosphorylation of insulin receptor substrate 1 by inhibitor κB kinase complex. J. Biol. Chem. 2002, 277, 48115–48121. [Google Scholar] [CrossRef] [PubMed]
- Chavez, J.A.; Knotts, T.A.; Wang, L.-P.; Li, G.; Dobrowsky, R.T.; Florant, G.L.; Summers, S.A. A Role for Ceramide, but Not Diacylglycerol, in the Antagonism of Insulin Signal Transduction by Saturated Fatty Acids. J. Biol. Chem. 2003, 278, 10297–10303. [Google Scholar] [CrossRef]
- Schmitz-Peiffer, C.; Craig, D.L.; Biden, T.J. Ceramide generation is sufficient to account for the inhibition of the insulin-stimulated PKB pathway in C2C12 skeletal muscle cells pretreated with palmitate. J. Biol. Chem. 1999, 274, 24202–24210. [Google Scholar] [CrossRef]
- Chen, W.Y.; Chen, C.J.; Liu, C.H.; Mao, F.C. Chromium supplementation enhances insulin signalling in skeletal muscle of obese KK/HlJ diabetic mice. Diabetes Obes. Metab. 2009, 11, 293–303. [Google Scholar] [CrossRef]
- Kim, S.T.; Moley, K.H. The expression of GLUT8, GLUT9a, and GLUT9b in the mouse testis and sperm. Reprod. Sci. 2007, 14, 445–455. [Google Scholar] [CrossRef]
- Maria, Z.; Campolo, A.R.; Scherlag, B.J.; Ritchey, J.W.; Lacombe, V.A. Dysregulation of insulin-sensitive glucose transporters during insulin resistance-induced atrial fibrillation. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 987–996. [Google Scholar] [CrossRef]
- Bibert, S.; Hess, S.K.; Firsov, D.; Thorens, B.; Geering, K.; Horisberger, J.-D.; Bonny, O. Mouse GLUT9: Evidences for a urate uniporter. Am. J. Physiol. Ren. Physiol. 2009, 297, F612–F619. [Google Scholar] [CrossRef] [PubMed]
- Augustin, R.; Carayannopoulos, M.O.; Dowd, L.O.; Phay, J.E.; Moley, J.F.; Moley, K.H. Identification and characterization of human glucose transporter-like protein-9 (GLUT9): Alternative splicing alters trafficking. J. Biol. Chem. 2004, 279, 16229–16236. [Google Scholar] [CrossRef] [PubMed]
- Zielke, H.R.; Sumbilla, C.M.; Sevdalian, D.A.; Hawkins, R.L.; Ozand, P.T. Lactate: A major product of glutamine metabolism by human diploid fibroblasts. J. Cell. Physiol. 1980, 104, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Reitzer, L.J.; BM, W.; Kennel, D. Evidence that glutamine, not sugar, is the major energy source for cultured HeLa cells. J. Biol. Chem. 1979, 254, 2669–2676. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Mancuso, A.; Daikhin, E.; Nissim, I.; Yudkoff, M.; Wehrli, S.; Thompson, C.B. Beyond aerobic glycolysis: Transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 19345–19350. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Escuredo, J.; Dadhich, R.K.; Dhup, S.; Cacace, A.; Van Hée, V.F.; De Saedeleer, C.J.; Sboarina, M.; Rodriguez, F.; Fontenille, M.-J.; Brisson, L. Lactate promotes glutamine uptake and metabolism in oxidative cancer cells. Cell Cycle 2016, 15, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Venneti, S.; Nagrath, D. Glutaminolysis: A hallmark of cancer metabolism. Annu. Rev. Biomed. Eng. 2017, 19, 163–194. [Google Scholar] [CrossRef]
- Bhagavan, N.; Ha, C.-E. Protein and amino acid metabolism. Essent. Med. Biochem. 2015, 227–268. [Google Scholar]
- Mullen, A.R.; Hu, Z.; Shi, X.; Jiang, L.; Boroughs, L.K.; Kovacs, Z.; Boriack, R.; Rakheja, D.; Sullivan, L.B.; Linehan, W.M. Oxidation of alpha-ketoglutarate is required for reductive carboxylation in cancer cells with mitochondrial defects. Cell Rep. 2014, 7, 1679–1690. [Google Scholar] [CrossRef]
- Jensen, M.V.; Joseph, J.W.; Ronnebaum, S.M.; Burgess, S.C.; Sherry, A.D.; Newgard, C.B. Metabolic cycling in control of glucose-stimulated insulin secretion. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1287–E1297. [Google Scholar] [CrossRef]
- Kavitha, T.S.; Parthasarathy, C.; Sivakumar, R.; Badrinarayanan, R.; Balasubramanian, K. Effects of excess corticosterone on NADPH generating enzymes and glucose oxidation in Leydig cells of adult rats. Hum. Exp. Toxicol. 2006, 25, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.-G.; Tong, L. Structure and function of malic enzymes, a new class of oxidative decarboxylases. Biochemistry 2003, 42, 12721–12733. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Lee, S. Fatty acid metabolism in ovarian cancer: Therapeutic implications. Int. J. Mol. Sci. 2022, 23, 2170. [Google Scholar] [CrossRef] [PubMed]
- Oshima, M.; Pechberty, S.; Bellini, L.; Göpel, S.O.; Campana, M.; Rouch, C.; Dairou, J.; Cosentino, C.; Fantuzzi, F.; Toivonen, S. Stearoyl CoA desaturase is a gatekeeper that protects human beta cells against lipotoxicity and maintains their identity. Diabetologia 2020, 63, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Deik, A.; Gonzalez, C.; Gonzalez, M.E.; Fu, F.; Ferrari, M.; Churchhouse, C.L.; Florez, J.C.; Jacobs, S.B.; Clish, C.B. Polyunsaturated fatty acid desaturation is a mechanism for glycolytic NAD+ recycling. Cell Metab. 2019, 29, 856–870.e7. [Google Scholar] [CrossRef] [PubMed]
- Amrolia, P.; Sullivan, M.; Garside, D.; Baldwin, S.; Cooke, B. An investigation of glucose uptake in relation to steroidogenesis in rat testis and tumour Leydig cells. Biochem. J. 1988, 249, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Van Straaten, H.W.; He, Y.; Van Duist, M.M.; Labruyère, W.T.; Vermeulen, J.L.; Van Dijk, P.J.; Ruijter, J.M.; Lamers, W.H.; Hakvoort, T.B. Cellular concentrations of glutamine synthetase in murine organs. Biochem. Cell Biol. 2006, 84, 215–231. [Google Scholar] [CrossRef]
- Chaban, Y.; Boekema, E.J.; Dudkina, N.V. Structures of mitochondrial oxidative phosphorylation supercomplexes and mechanisms for their stabilisation. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 418–426. [Google Scholar] [CrossRef]
- Bassi, G.; Sidhu, S.K.; Mishra, S. The expanding role of mitochondria, autophagy and lipophagy in steroidogenesis. Cells 2021, 10, 1851. [Google Scholar] [CrossRef]
- Ly, L.D.; Xu, S.; Choi, S.-K.; Ha, C.-M.; Thoudam, T.; Cha, S.-K.; Wiederkehr, A.; Wollheim, C.B.; Lee, I.-K.; Park, K.-S. Oxidative stress and calcium dysregulation by palmitate in type 2 diabetes. Exp. Mol. Med. 2017, 49, e291. [Google Scholar] [CrossRef]
- Quinlan, C.L.; Orr, A.L.; Perevoshchikova, I.V.; Treberg, J.R.; Ackrell, B.A.; Brand, M.D. Mitochondrial complex II can generate reactive oxygen species at high rates in both the forward and reverse reactions. J. Biol. Chem. 2012, 287, 27255–27264. [Google Scholar] [CrossRef] [PubMed]
- Kraus, M.; Neuzil, J.; Rohlena, J. Mitochondrial complex II and reactive oxygen species in disease and therapy. Redox Rep. 2020, 25, 26–32. [Google Scholar]
- Chen, Q.; Vazquez, E.J.; Moghaddas, S.; Hoppel, C.L.; Lesnefsky, E.J. Production of reactive oxygen species by mitochondria: Central role of complex III. J. Biol. Chem. 2003, 278, 36027–36031. [Google Scholar] [CrossRef] [PubMed]
- Sergi, D.; Luscombe-Marsh, N.; Naumovski, N.; Abeywardena, M.; O’Callaghan, N. Palmitic acid, but not lauric acid, induces metabolic inflammation, mitochondrial fragmentation, and a drop in mitochondrial membrane potential in human primary myotubes. Front. Nutr. 2021, 8, 663838. [Google Scholar] [CrossRef] [PubMed]
- Palomino, O.M.; Giordani, V.; Chowen, J.; Fernández-Alfonso, M.S.; Goya, L. Physiological doses of oleic and palmitic acids protect human endothelial cells from oxidative stress. Molecules 2022, 27, 5217. [Google Scholar] [CrossRef] [PubMed]
- Haffar, T.; Akoumi, A.; Bousette, N. Lipotoxic palmitate impairs the rate of β-oxidation and citric acid cycle flux in rat neonatal cardiomyocytes. Cell. Physiol. Biochem. 2016, 40, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Egnatchik, R.A.; Leamy, A.K.; Noguchi, Y.; Shiota, M.; Young, J.D. Palmitate-induced activation of mitochondrial metabolism promotes oxidative stress and apoptosis in H4IIEC3 rat hepatocytes. Metabolism 2014, 63, 283–295. [Google Scholar] [CrossRef]
- Koves, T.R.; Ussher, J.R.; Noland, R.C.; Slentz, D.; Mosedale, M.; Ilkayeva, O.; Bain, J.; Stevens, R.; Dyck, J.R.; Newgard, C.B. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008, 7, 45–56. [Google Scholar] [CrossRef]
- Wang, H.; Hu, L.; Li, H.; Lai, Y.-T.; Wei, X.; Xu, X.; Cao, Z.; Cao, H.; Wan, Q.; Chang, Y.-Y. Mitochondrial ATP synthase as a direct molecular target of chromium (III) to ameliorate hyperglycaemia stress. Nat. Commun. 2023, 14, 1738. [Google Scholar] [CrossRef]
- Navin, A.K.; Aruldhas, M.M.; Navaneethabalakrishnan, S.; Mani, K.; Michael, F.M.; Srinivasan, N.; Banu, S.K. Prenatal exposure to hexavalent chromium disrupts testicular steroidogenic pathway in peripubertal F(1) rats. Reprod. Toxicol. 2021, 101, 63–73. [Google Scholar] [CrossRef]
- Pihlajamäki, J.; Gylling, H.; Miettinen, T.A.; Laakso, M. Insulin resistance is associated with increased cholesterol synthesis and decreased cholesterol absorption in normoglycemic men. J. Lipid Res. 2004, 45, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Pignataro, O.P.; Radicella, J.P.; Calvo, J.C.; Charreau, E.H. Mitochondrial biosynthesis of cholesterol in Leydig cells from rat testis. Mol. Cell. Endocrinol. 1983, 33, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Tarrahi, M.J.; Tarrahi, M.A.; Rafiee, M.; Mansourian, M. The effects of chromium supplementation on lipidprofile in humans: A systematic review and meta-analysis ofrandomized controlled trials. Pharmacol. Res. 2021, 164, 105308. [Google Scholar] [CrossRef] [PubMed]
- Sawicka, E.; Jurkowska, K.; Piwowar, A. Estrogenowe działanie chromu–ważny metaloestrogen w modulowaniu szlaków endokrynnych. Farm. Pol. 2019, 75, 357–364. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Catalog Number | Host Species | Predicted Molecular Weight (kDa) | Dilution | Vendor |
|---|---|---|---|---|---|
| p-IRS-1 (Ser307) | #2381 | Rabbit | 132 | 1:500 | Cell Signaling, Massachusetts, USA |
| OXPHOS | ab110413 | Mouse | 20, 30, 40, 48, and 55 | 1:2000 | Abcam, United Kingdom |
| 4-HNE | AB5605 | Goat | n.a. | 1:1000 | EMD Millipore, California, USA |
| DNP | D9656 | Rabbit | n.a. | 1:5000 | Sigma-Aldrich, St. Louis, Missouri, USA |
| (CrPic3) (µM) | (PA) (µM) | Metabolites Consumed | Sum of Cs Consumed * | Metabolites Produced | Sum of Cs Produced | Ratio of Cs Produced vs. Consumed | |||
|---|---|---|---|---|---|---|---|---|---|
| Glucose | Glutamine | Alanine | Lactate | Pyruvate | |||||
| 0 | 0 | 21.59 | 160.2 | 13.41 | 650.37 | 407.4 | 34.12 | 1324.56 | 2.04 |
| 0.1 | 0 | 20.04 | 184.5 | 7.53 | 696.33 | 384.4 | 35.11 | 1258.53 | 1.81 |
| 10 | 0 | 27.46 | 187.8 | 11.25 | 761.91 | 303.1 | 28.93 | 996.09 | 1.31 |
| 0 | 250 | 24.93 | 198.9 | 10.29 | 777.15 | 221.7 | 43.3 | 795.00 | 1.03 |
| 0.1 | 250 | 31.29 | 231.7 | 13.12 | 922.20 | 249.5 | 43.21 | 878.13 | 0.95 |
| 10 | 250 | 15.05 | 178.1 | 16.77 | 674.91 | 234.8 | 18.36 | 759.48 | 1.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, R.; Martins, A.D.; Ferreira, R.; Alves, M.G.; Pereira, M.d.L.; Oliveira, P.F. Impact of Chromium Picolinate on Leydig Cell Steroidogenesis and Antioxidant Balance Using an In Vitro Insulin Resistance Model. Antioxidants 2024, 13, 40. https://doi.org/10.3390/antiox13010040
Moreira R, Martins AD, Ferreira R, Alves MG, Pereira MdL, Oliveira PF. Impact of Chromium Picolinate on Leydig Cell Steroidogenesis and Antioxidant Balance Using an In Vitro Insulin Resistance Model. Antioxidants. 2024; 13(1):40. https://doi.org/10.3390/antiox13010040
Chicago/Turabian StyleMoreira, Rúben, Ana D. Martins, Rita Ferreira, Marco G. Alves, Maria de Lourdes Pereira, and Pedro F. Oliveira. 2024. "Impact of Chromium Picolinate on Leydig Cell Steroidogenesis and Antioxidant Balance Using an In Vitro Insulin Resistance Model" Antioxidants 13, no. 1: 40. https://doi.org/10.3390/antiox13010040
APA StyleMoreira, R., Martins, A. D., Ferreira, R., Alves, M. G., Pereira, M. d. L., & Oliveira, P. F. (2024). Impact of Chromium Picolinate on Leydig Cell Steroidogenesis and Antioxidant Balance Using an In Vitro Insulin Resistance Model. Antioxidants, 13(1), 40. https://doi.org/10.3390/antiox13010040