

Changes in Phenolic Compounds and Antioxidant Capacity of Artocarpus heterophyllus Lam. (Jackfruit) Pulp during In Vitro Gastrointestinal Digestion
 ,
, 
Abstract
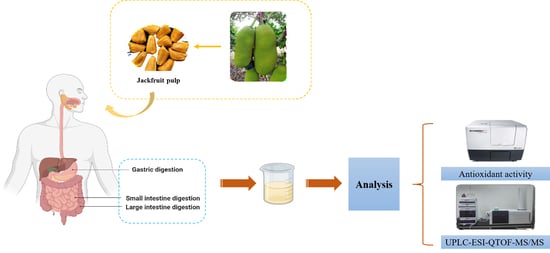
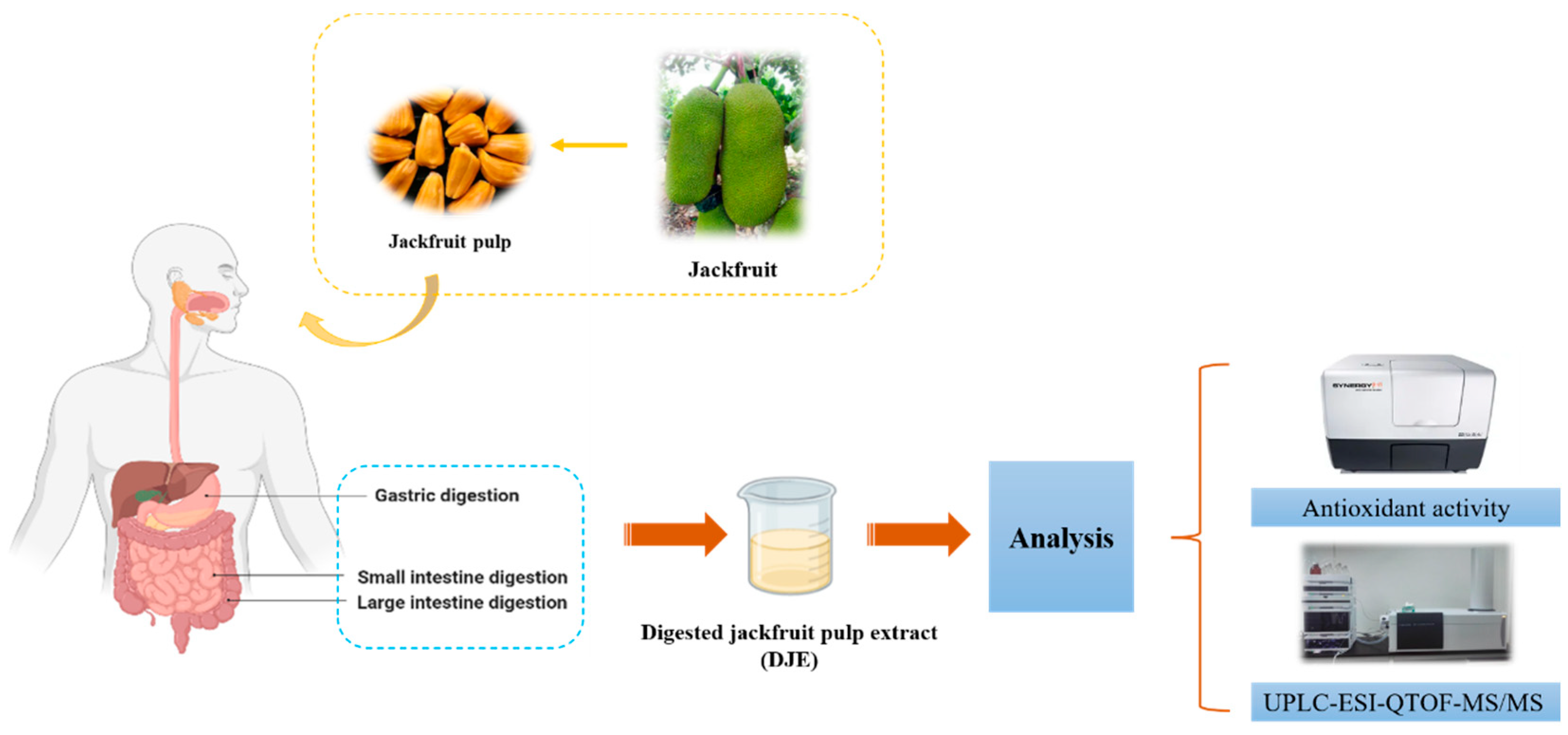
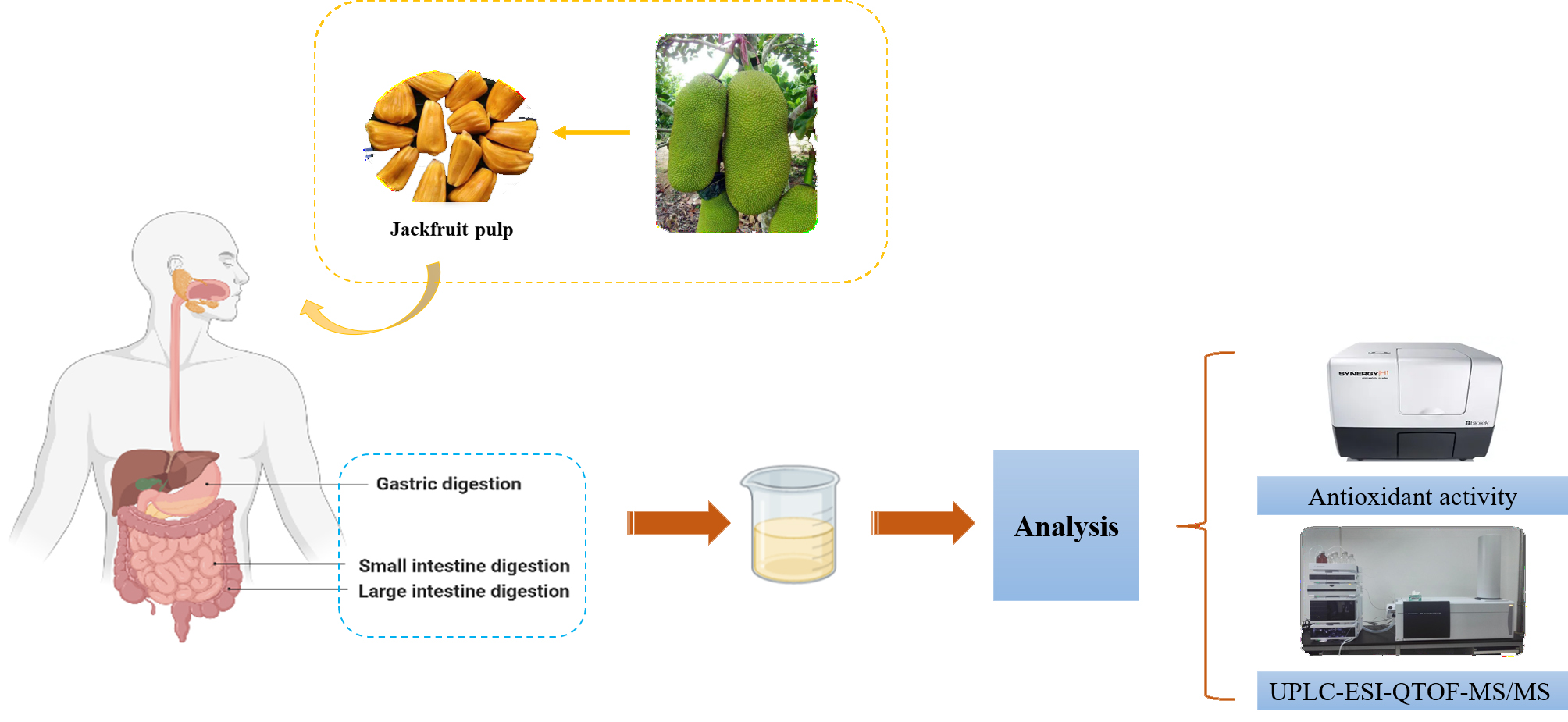
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
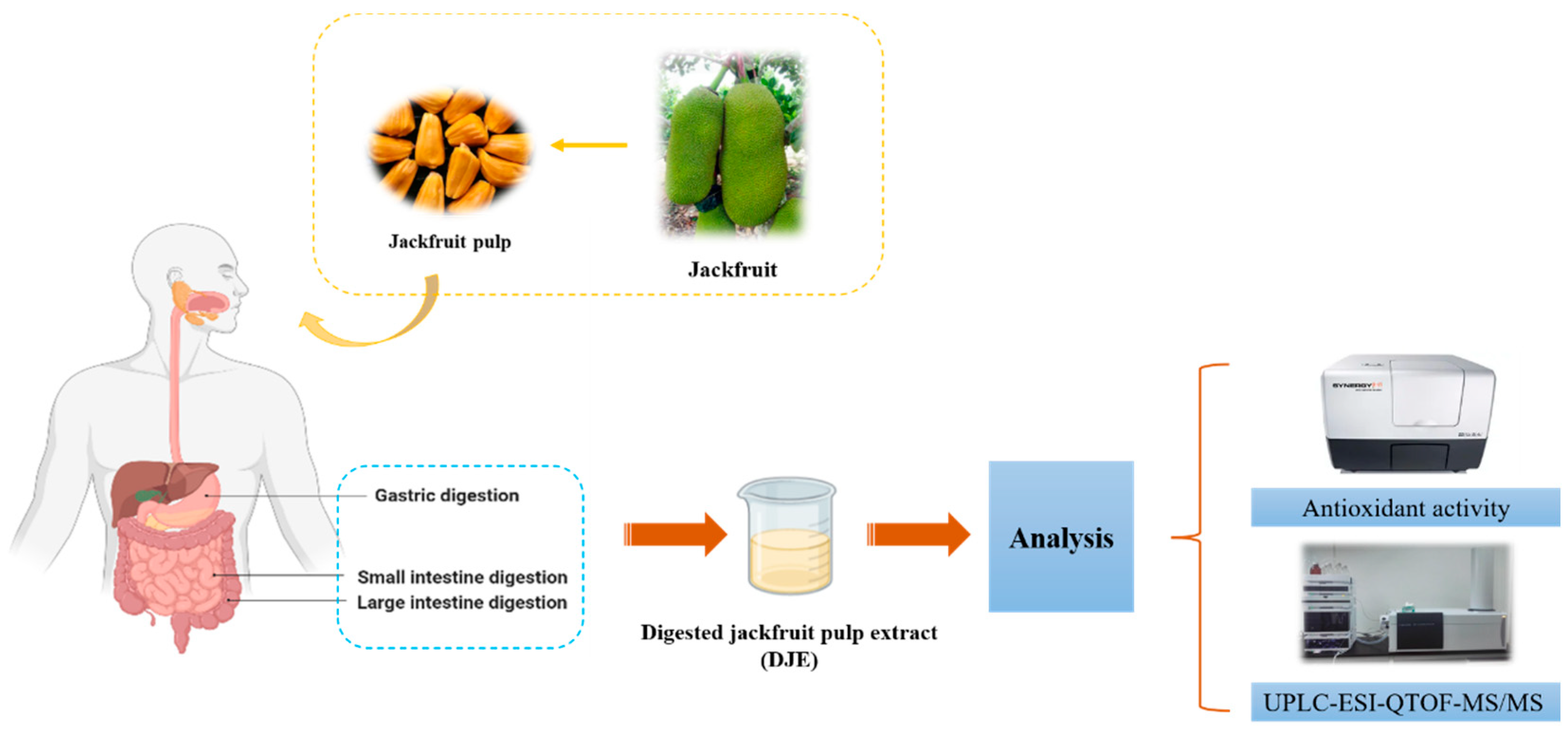
2.2. In Vitro Gastrointestinal Digestion
2.3. Extraction of Digested Jackfruit Pulp Samples
2.4. Determination of Total Phenol Content (TPC)
2.5. Analysis of Phenolic Compounds
2.6. Antioxidant Activity Assay
2.7. Statistical Analysis
3. Results and Discussion
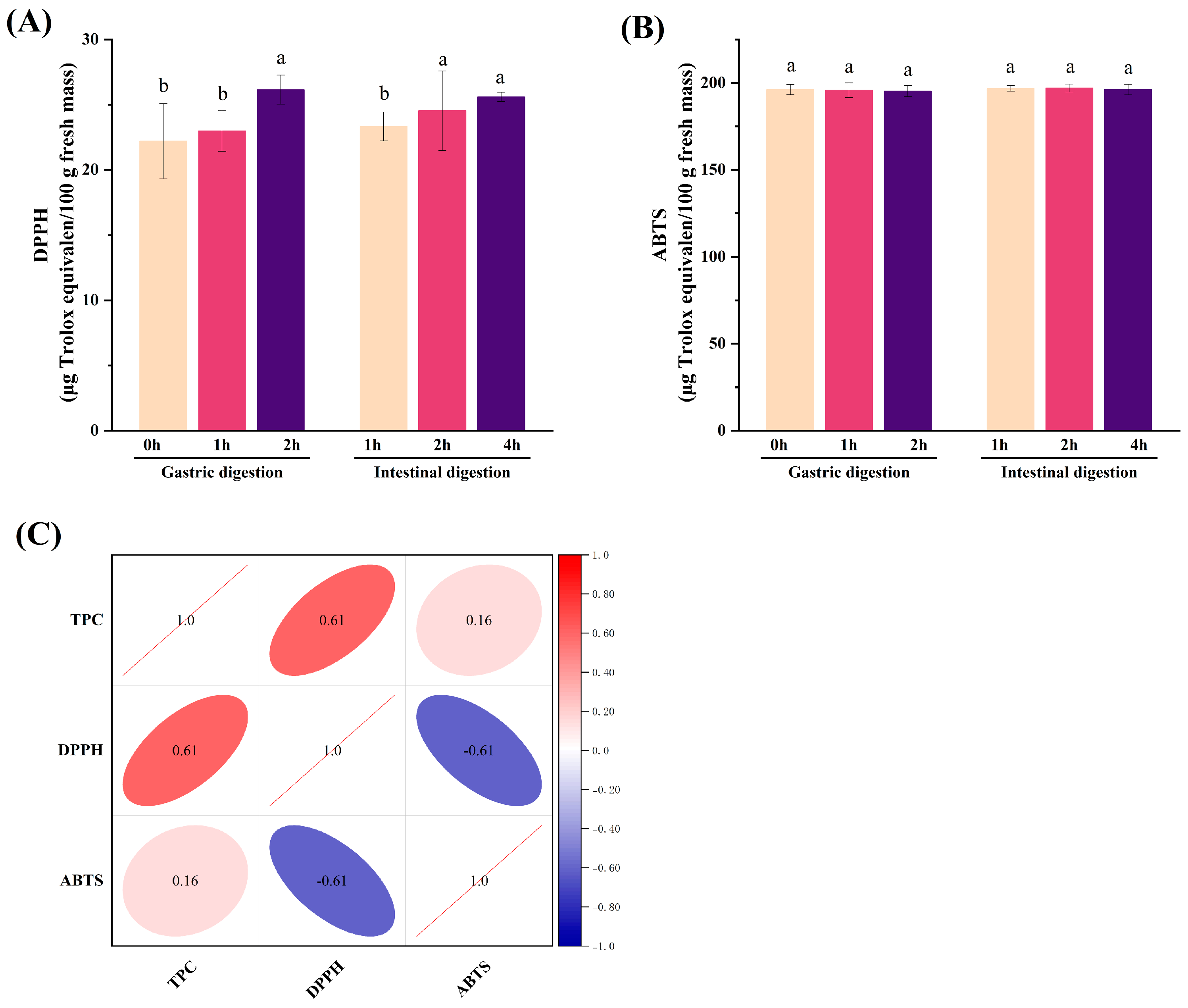
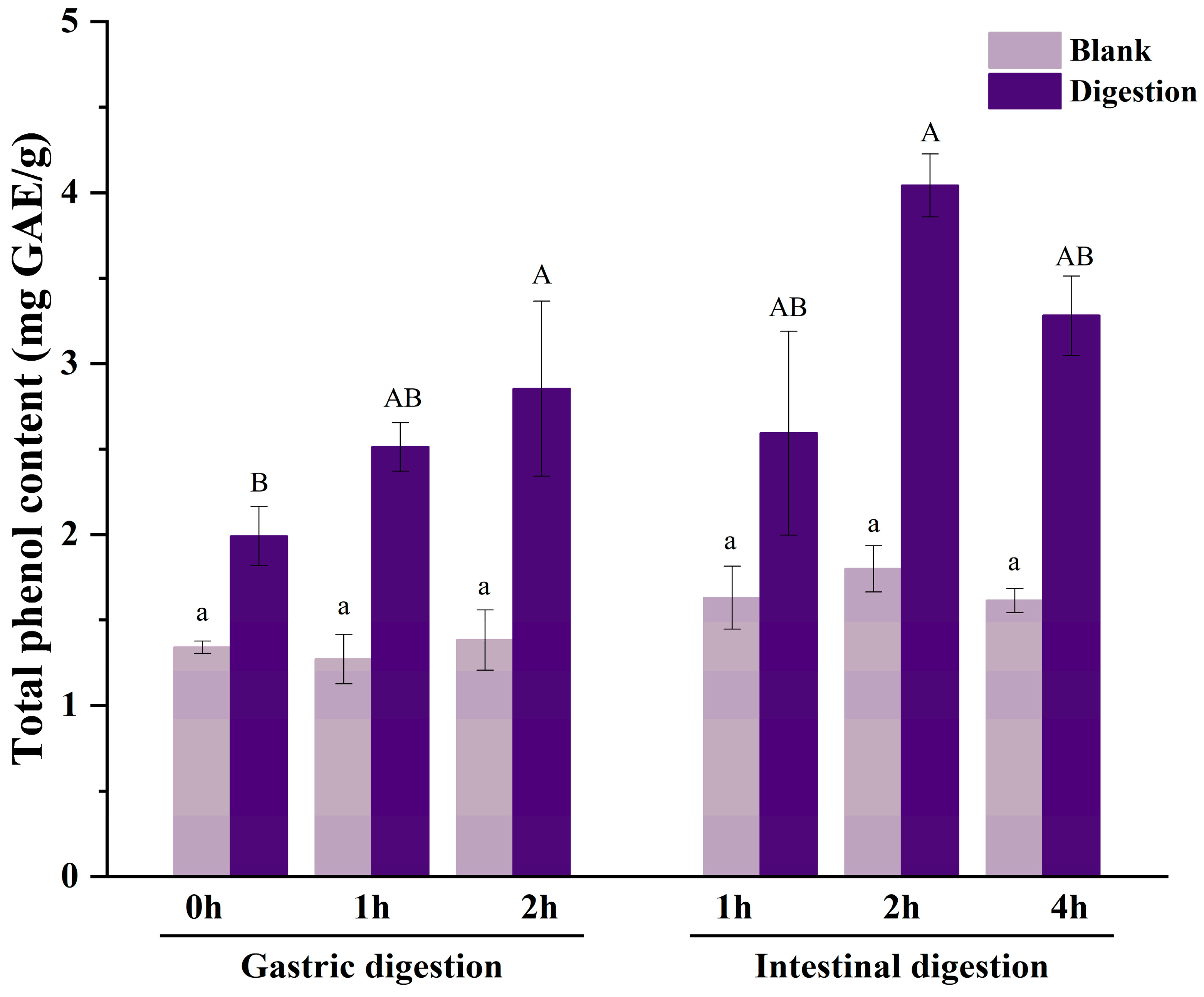
3.1. TPC
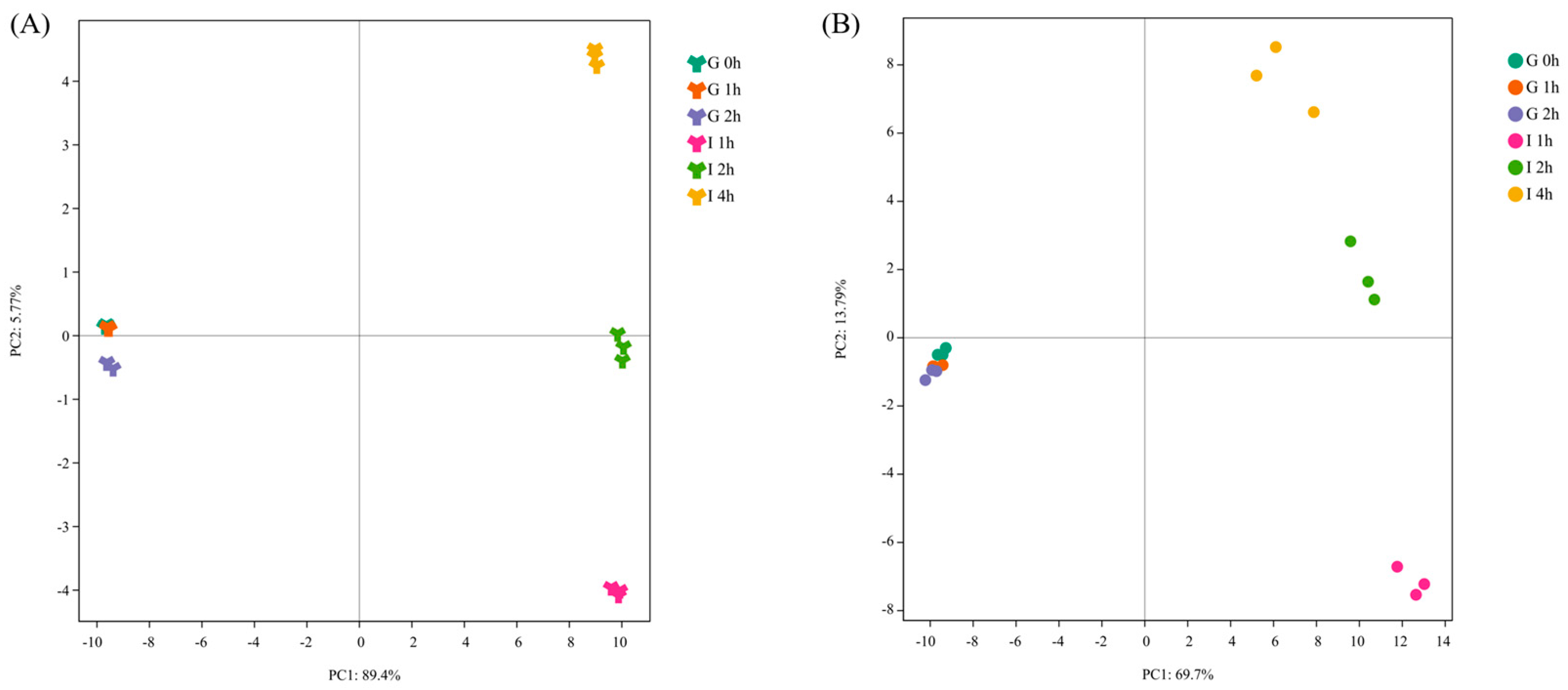
3.2. Identification of Polyphenols and their Decomposition Products after In Vitro Digestion
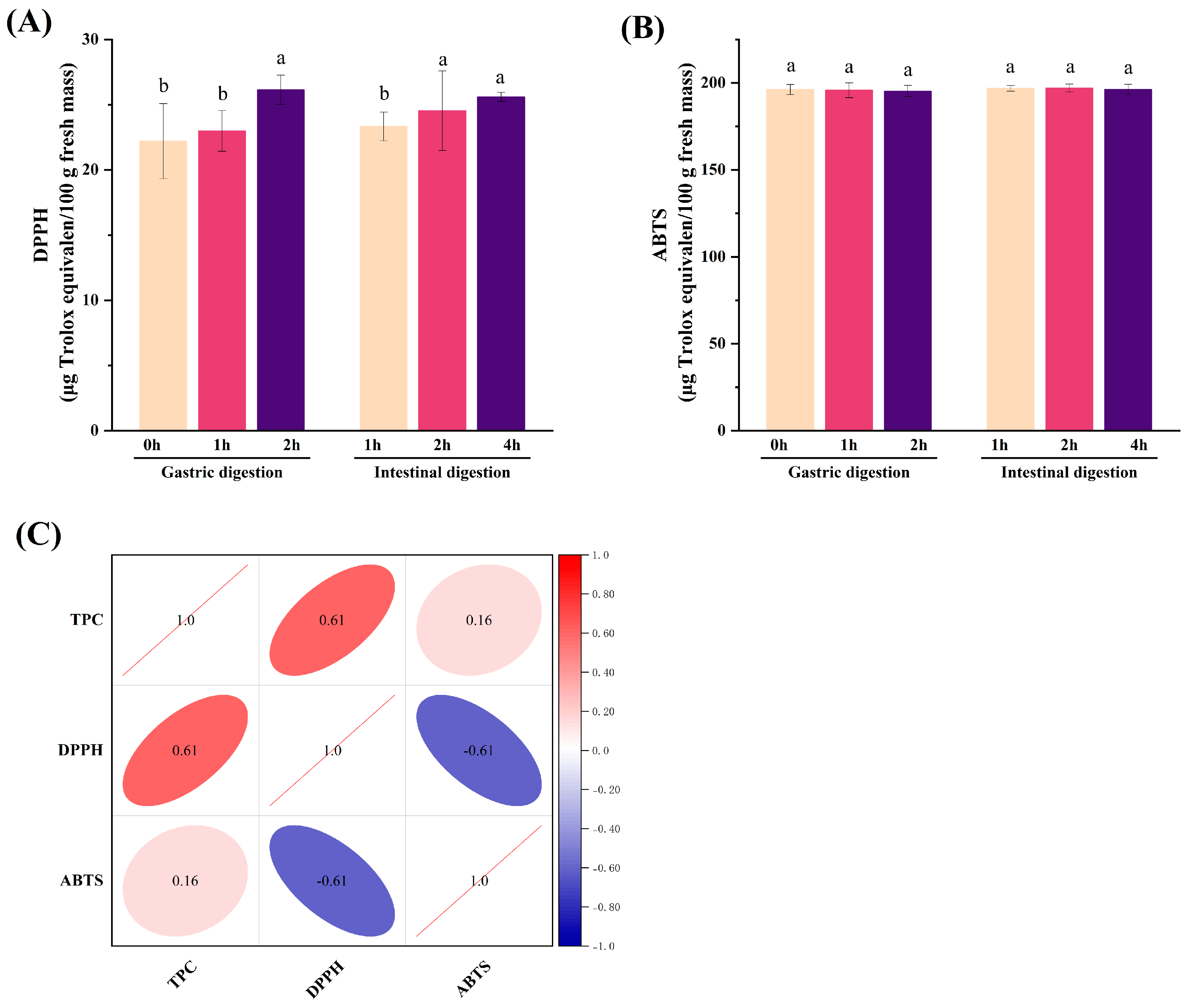
3.3. Antioxidant Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shafiq, M.; Mehmood, S.; Yasmin, A.; Khan, S.J.; Khan, N.H.; Ali, S. Evaluation of phytochemical, nutritional and antioxidant activity of indigenously grown jackfruit (Artocarpus heterophyllus Lam). J. Sci. Res. 2017, 9, 135–143. [Google Scholar] [CrossRef]
- Gupta, A.; Marquess, A.R.; Pandey, A.K.; Bishayee, A. Jackfruit (Artocarpus heterophyllus Lam.) in health and disease: A critical review. Crit. Rev. Food Sci. Nutr. 2022, 63, 6344–6378. [Google Scholar] [CrossRef] [PubMed]
- Vazhacharickal, P.J.; Sajeshkumar, N.K.; Mathew, J.J.; Kuriakose, A.C.; Abraham, B.; Mathew, R.J.; Albin, A.N.; Thomson, D.; Thomas, R.S.; Varghese, N.; et al. Chemistry and medicinal properties of jackfruit (Artocarpus heterophyllus): A review on current status of knowledge. Int. J. Innov. Res. Rev. 2015, 3, 83–95. [Google Scholar]
- Zhang, L.; Tu, Z.; Xie, X.; Wang, H.; Wang, H.; Wang, Z.; Sha, X.; Lu, Y. Jackfruit (Artocarpus heterophyllus Lam.) peel: A better source of antioxidants and α-glucosidase inhibitors than pulp, flake and seed, and phytochemical profile by HPLC-QTOF-MS/MS. Food Chem. 2017, 234, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Agunloye, O.M.; Oboh, G.; Ademiluyi, A.O.; Ademosun, A.O.; Akindahunsi, A.A.; Oyagbemi, A.A.; Omobowale, T.O.; Ajibade, T.O.; Adedapo, A.A. Cardio-protective and antioxidant properties of caffeic acid and chlorogenic acid: Mechanistic role of angiotensin converting enzyme, cholinesterase and arginase activities in cyclosporine induced hypertensive rats. Biomed. Pharmacother. 2019, 109, 450–458. [Google Scholar] [CrossRef]
- Ali, F.; Rahul; Jyoti, S.; Naz, F.; Ashafaq, M.; Shahid, M.; Siddique, Y.H. Therapeutic potential of luteolin in transgenic Drosophila model of Alzheimer’s disease. Neurosci. Lett. 2019, 692, 90–99. [Google Scholar] [CrossRef] [PubMed]
- De Paulo Farias, D.; Neri-Numa, I.A.; de Araújo, F.F.; Pastore, G.M. A critical review of some fruit trees from the Myrtaceae family as promising sources for food applications with functional claims. Food Chem. 2020, 306, 125630. [Google Scholar] [CrossRef]
- Elsayed, R.H.; Kamel, E.M.; Mahmoud, A.M.; El-Bassuony, A.A.; Bin-Jumah, M.; Lamsabhi, A.M.; Ahmed, S.A. Rumex dentatus L. phenolics ameliorate hyperglycemia by modulating hepatic key enzymes of carbohydrate metabolism, oxidative stress and PPARγ in diabetic rats. Food Chem. Toxicol. 2020, 138, 111202. [Google Scholar] [CrossRef]
- Liu, F.; Li, L.; Lu, W.; Ding, Z.; Huang, W.; Li, Y.T.; Cheng, C.; Shan, W.S.; Xu, J.; He, W.; et al. Scutellarin ameliorates cartilage degeneration in osteoarthritis by inhibiting the Wnt/β-catenin and MAPK signaling pathways. Int. Immunopharmacol. 2020, 78, 105954. [Google Scholar] [CrossRef]
- Alminger, M.; Aura, A.M.; Bohn, T.; Dufour, C.; El, S.N.; Gomes, A.; Karakaya, S.; Martínez-Cuesta, M.C.; McDougall, G.J.; Requena, T.; et al. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef]
- Hao, Y.; Yang, J.; Cui, J.; Fan, Y.; Li, N.; Wang, C.; Liu, Y.; Dong, Y. Stability and mechanism of phenolic compounds from raspberry extract under in vitro gastrointestinal digestion. LWT Food Sci. Technol. 2021, 139, 110552. [Google Scholar] [CrossRef]
- Khochapong, W.; Ketnawa, S.; Ogawa, Y.; Punbusayakul, N. Effect of in vitro digestion on bioactive compounds, antioxidant and antimicrobial activities of coffee (Coffea arabica L.) pulp aqueous extract. Food Chem. 2021, 348, 129094. [Google Scholar] [CrossRef] [PubMed]
- Lucas-González, R.; Viuda-Martos, M.; Pérez Álvarez, J.A.; Fernández-López, J. Changes in bioaccessibility, polyphenol profile and antioxidant potential of flours obtained from persimmon fruit (Diospyros kaki) co-products during in vitro gastrointestinal digestion. Food Chem. 2018, 256, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Aschoff, J.K.; Kaufmann, S.; Kalkan, O.; Neidhart, S.; Carle, R.; Schweiggert, R.M. In vitro bioaccessibility of carotenoids, flavonoids, and vitamin C from differently processed oranges and orange juices [Citrus sinensis (L.) Osbeck]. J. Agric. Food Chem. 2015, 63, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Konsue, N.; Bunyameen, N.; Donlao, N. Utilization of young jackfruit (Artocarpus heterophyllus Lam.) as a plant-based food ingredient: Influence of maturity on chemical attributes and changes during in vitro digestion. LWT Food Sci. Technol. 2023, 180, 114721. [Google Scholar] [CrossRef]
- Zhu, K.; Yao, S.; Zhang, Y.; Liu, Q.; Xu, F.; Wu, G.; Dong, W.; Tan, L. Effects of in vitro saliva, gastric and intestinal digestion on the chemical properties, antioxidant activity of polysaccharide from Artocarpus heterophyllus Lam. (Jackfruit) pulp. Food Hydrocoll. 2019, 87, 952–959. [Google Scholar] [CrossRef]
- Qu, D.; Liu, C.; Jiang, M.; Feng, L.; Chen, Y.; Han, J. After in vitro digestion, jackfruit flake affords protection against Acrylamide-induced oxidative damage. Molecules 2019, 24, 3322. [Google Scholar] [CrossRef]
- Pavan, V.; Sancho, R.A.S.; Pastore, G.M. The effect of in vitro digestion on the antioxidant activity of fruit extracts (Carica papaya, Artocarpus heterophillus and Annona marcgravii). LWT Food Sci. Technol. 2014, 59, 1247–1251. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Cheng, M.; He, J.; Wang, H.; Li, C.; Wu, G.; Zhu, K.; Chen, X.; Zhang, Y.; Tan, L. Comparison of microwave, ultrasound and ultrasound-microwave assisted solvent extraction methods on phenolic profile and antioxidant activity of extracts from jackfruit (Artocarpus heterophyllus Lam.) pulp. LWT Food Sci. Technol. 2023, 173, 114395. [Google Scholar] [CrossRef]
- Dong, R.; Yu, Q.; Liao, W.; Liu, S.; He, Z.; Hu, X.; Chen, Y.; Xie, J.; Nie, S.; Xie, M. Composition of bound polyphenols from carrot dietary fiber and its in vivo and in vitro antioxidant activity. Food Chem. 2021, 339, 127879. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wu, T.; Chu, X.; Tang, S.; Cao, W.; Liang, F.; Fang, Y.; Pan, S.; Xu, X. Fermented blueberry pomace with antioxidant properties improves fecal microbiota community structure and short chain fatty acids production in an in vitro mode. LWT Food Sci. Technol. 2020, 125, 109260. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Grijalva, E.P.; Angulo-Escalante, M.A.; Leon-Felix, J.; Heredia, J.B. Effect of In Vitro Digestion on the Total Antioxidant Capacity and Phenolic Content of 3 Species of Oregano (Hedeoma patens, Lippia graveolens, Lippia palmeri). J. Food Sci. 2017, 82, 2832. [Google Scholar] [CrossRef] [PubMed]
- Tischer Seraglio, S.K.; Valese, A.C.; Daguer, H.; Bergamo, G.; Azevedo, M.S.; Nehring, P.; Gonzaga, L.V.; Fett, R.; Oliveira Costa, A.C. Effect of in vitro gastrointestinal digestion on the bioaccessibility of phenolic compounds, minerals, and antioxidant capacity of Mimosa scabrella Bentham honeydew honeys. Food Res. Int. 2017, 99, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shi, J.; Yi, J.; Zhang, X.; Ma, Q.; Cai, S. The effect of in vitro simulated gastrointestinal digestion on phenolic bioaccessibility and bioactivities of Prinsepia utilis Royle fruits. LWT Food Sci. Technol. 2021, 138, 110782. [Google Scholar] [CrossRef]
- Tu, F.; Xie, C.; Li, H.; Lei, S.; Li, J.; Huang, X.; Yang, F. Effect of in vitro digestion on chestnut outer-skin and inner-skin bioaccessibility: The relationship between biotransformation and antioxidant activity of polyphenols by metabolomics. Food Chem. 2021, 363, 130277. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, J.; Wei, Z.; Shahidi, F. Effect of in vitro digestion on phenolics and antioxidant activity of red and yellow colored pea hulls. Food Chem. 2021, 337, 127606. [Google Scholar] [CrossRef]
- Peña-Vázquez, G.I.; Dominguez-Fernández, M.T.; Camacho-Zamora, B.D.; Hernandez-Salazar, M.; Urías-Orona, V.; De Peña, M.; de la Garza, A.L. In vitro simulated gastrointestinal digestion impacts bioaccessibility and bioactivity of Sweet orange (Citrus sinensis) phenolic compounds. J. Funct. Foods 2022, 88, 104891. [Google Scholar] [CrossRef]
- Velderrain-Rodríguez, G.; Quirós-Sauceda, A.; Mercado-Mercado, G.; Ayala-Zavala, J.F.; Astiazarán-García, H.; Robles-Sánchez, R.M.; Wall-Medrano, A.; Sayago-Ayerdi, S.; González-Aguilar, G.A. Effect of dietary fiber on the bioaccessibility of phenolic compounds of mango, papaya and pineapple fruits by an in vitro digestion model. Food Sci. Technol. 2016, 36, 188–194. [Google Scholar] [CrossRef]
- Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F.; Tomás-Barberán, F. In Vitro Availability of Flavonoids and Other Phenolics in Orange Juice. Food Chem. 2001, 49, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, A.; Kumar, B. Identification and characterization of phenolics and terpenoids from ethanolic extracts of Phyllanthus species by HPLC-ESI-QTOF-MS/MS. J. Pharm. Anal. 2017, 7, 214–222. [Google Scholar] [CrossRef] [PubMed]
- De Bellis, R.; Chiarantini, L.; Potenza, L.; Gorassini, A.; Verardo, G.; De Marco, R.; Benayada, L.; Stocchi, V.; Cristina Albertini, M.; Fraternale, D. High production of secondary metabolites and biological activities of Cydonia oblonga Mill. pulp fruit callus. J. Funct. Foods 2022, 94, 105133. [Google Scholar] [CrossRef]
- Kiselova-Kaneva, Y.; Galunska, B.; Nikolova, M.; Dincheva, I.; Badjakov, I. High resolution LC-MS/MS characterization of polyphenolic composition and evaluation of antioxidant activity of Sambucus ebulus fruit tea traditionally used in Bulgaria as a functional food. Food Chem. 2022, 367, 130759. [Google Scholar] [CrossRef] [PubMed]
- Alves Filho, E.G.; Sousa, V.M.; Rodrigues, S.; de Brito, E.S.; Fernandes, F.A.N. Green ultrasound-assisted extraction of chlorogenic acids from sweet potato peels and sonochemical hydrolysis of caffeoylquinic acids derivatives. Ultrason. Sonochem. 2020, 63, 104911. [Google Scholar] [CrossRef]
- Yang, L.; Marney, L.; Magana, A.A.; Choi, J.; Wright, K.; Mcferrin, J.; Gray, N.E.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Quantification of caffeoylquinic acids and triterpenes as targeted bioactive compounds of Centella Asiatica in extracts and formulations by liquid chromatography mass spectrometry. J. Chromatogr. Open 2023, 4, 100091. [Google Scholar] [CrossRef]
- Acero, N.; Gradillas, A.; Beltran, M.; Garcia, A.; Mingarro, D.M. Comparison of phenolic compounds profile and antioxidant properties of different sweet cherry (Prunus avium L.) varieties. Food Chem. 2019, 279, 260–271. [Google Scholar] [CrossRef]
- Daud, M.; Fatanah, D.N.; Abdullah, N.; Ahmad, R. Evaluation of antioxidant potential of Artocarpus heterophyllus L. J33 variety fruit waste from different extraction methods and identification of phenolic constituents by LCMS. Food Chem. 2017, 232, 621–632. [Google Scholar] [CrossRef]
- Torres, A.; Aguilar-Osorio, G.; Camacho, M.; Basurto, F.; Navarro-Ocana, A. Characterization of polyphenol oxidase from purple sweet potato (Ipomoea batatas L. Lam) and its affinity towards acylated anthocyanins and caffeoylquinic acid derivatives. Food Chem. 2021, 356, 129709. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Arráez-Román, D.; Lozano-Sánchez, J.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Phytochemical Characterisation of Green Beans (Phaseolus vulgaris L.) by Using High-performance Liquid Chromatography Coupled with Time-of-flight Mass Spectrometry. Phytochem. Anal. 2013, 24, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, X.; Zhan, L.; Xu, C.; Sun, L.; Jiang, H.; Sun, C.; Li, X. LC-Q-TOF-MS characterization of polyphenols from white bayberry fruit and its antidiabetic effect in KK-Ay mice. ACS Omega 2020, 5, 17839–17849. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Lin, G.; Lin, H.; Chang, S. Characteristics of proanthocyanidins in leaves of Chamaecyparis obtusa var. formosana as strong α-glucosidase inhibitors. J. Sci. Food Agric. 2018, 98, 3806–3814. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiao, J.; Chen, L.; Hu, C.; Chen, P.; Xie, B.; Sun, Z. Identification of A-series oligomeric procyanidins from pericarp of Litchi chinensis by FT-ICR-MS and LC-MS. Food Chem. 2012, 135, 31–38. [Google Scholar] [CrossRef]
- Xue, C.; Guo, J.; Qian, D.; Duan, J.; Shang, E.; Shu, Y.; Lu, Y. Identification of the potential active components of Abelmoschus manihot in rat blood and kidney tissue by microdialysis combined with ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometry. J. Chromatogr. B 2011, 879, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Tejada, S.; Pinya, S.; Martorell, M.; Capó, X.; Tur, J.A.; Pons, A.; Sureda, A. Potential anti-inflammatory effects of hesperidin from the genus Citrus. Curr. Med. Chem. 2017, 24, 4929. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y. Chemical constituents of Fortunella margarita fruits. J. Chin. Med. Mater. 2014, 37, 435–438. (In Chinese) [Google Scholar]
- Quail, W.; Tao, Y.; Lu, M.; Yuan, B.; Chen, J.; Zeng, M.; Qin, F.; Guo, F.; He, Z. Stability of the phenolic compounds and antioxidant capacity of five fruit (apple, orange, grape, pomelo and kiwi) juices during in vitro: Imulated gastrointestinal digestion. Int. J. Food Sci. Technol. 2018, 53, 1131–1139. [Google Scholar]
- Barreira, J.C.M.; Ferreira, I.C.F.R.; Oliveira, M.B.P.P.; Pereira, J.A. Antioxidant activities of the extracts from chestnut flower, leaf, skins and fruit. Food Chem. 2008, 107, 1106–1113. [Google Scholar] [CrossRef]
- Pods Dek, A.; Majewska, I.; Redzynia, M.; Sosnowska, D.; Kozio Kiewicz, M. In vitro inhibitory effect on digestive enzymes and antioxidant potential of commonly consumed fruits. J. Agric. Food Chem. 2014, 62, 4610–4617. [Google Scholar] [CrossRef]
- Jayawardena, N.; Watawana, M.I.; Waisundara, V.Y. The total antioxidant capacity, total phenolics content and starch hydrolase inhibitory activity of fruit juices following pepsin (gastric) and pancreatin (duodenal) digestion. J. Consum. Prot. Food Saf. 2015, 10, 349–357. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Agilent 1290 Infinity II UPLC, 6530B Hybrid Q-TOF-MS |
|---|---|
| Column | Agilent Zorbax Eclipse Plus C18 column (3.0 mm × 150 mm, 1.8 μm) |
| Mobile phase | 0.1%formic acid in water (A) and acetonitrile (B) |
| Phase gradient | 0–1.5 min, 5% B; 1.5–15 min, 5–60% B; 15–25 min, 60–100% B; 30–30.10 min, 100–5% B; 30.10–35 min, 5% B |
| Injection volume | 3 μL |
| Column temperature | 35 ℃ |
| Flow rate | 0.4 mL/min |
| Sheath gas | Helium |
| Sheath gas temperature | 325 ℃ |
| Sheath gas flow rate | 11 L/min |
| Crash voltage | 140 V |
| Compd | Rt (min) | MS | MS/MS | Formula | Identification | Distribution | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| G0 | G1 | G2 | I1 | I2 | I4 | ||||||
| Hydroxybenzoic acids and derivatives | |||||||||||
| 1 | 3.172 | 171.0291 | 125 | C7H6O5 | Gallic acid | ✓ | |||||
| 2 | 1.431 | 257.0657 | 153 | C9H10O5 | Syringic acid | ✓ | |||||
| Hydroxycinnamic acids and derivatives | |||||||||||
| 3 | 7.522 | 337.0906 | 147, 119 | C16H18O8 | 5-p-Coumaroylquinic acid | ✓ | ✓ | ||||
| 4 | 7.323 | 399.0939 | 173, 179 | C16H18O9 | 1-Caffeoylquinic acid | ✓ | |||||
| 5 | 1.44 | 539.1212 | 163, 319 | C25H24O12 | (E,E)-3,5-di-O-Caffeoylquinic acid | ✓ | |||||
| 6 | 1.674 | 367.1056 | 135 | C17H20O9 | 3-O-Caffeoylquinic acid methyl ester | ✓ | |||||
| 7 | 8.757 | 531.1475 | 204,163 | C26H26O12 | 4,5-di-O-Caffeoylquinic acid ester | ✓ | ✓ | ||||
| 8 | 4.503 | 543.1557 | 326, 163 | C27H28O12 | Ethyl-3,4-dicaffeoylquinate | ✓ | |||||
| Flavanols | |||||||||||
| 9 | 1.563 | 353.0869 | 245 | C15H16O7 | (+)-Catechin hydrate | ✓ | ✓ | ||||
| 10 | 8.304 | 441.0798 | 167, 137, 125 | C22H18O10 | (−)-Epicatechin gallate | ✓ | |||||
| 11 | 10.161 | 459.0919 | 125, 137,139 | C22H18O11 | Gallocatechin | ✓ | |||||
| 12 | 20.856 | 599.1153 | 539, 407, 285 | C30H24O12 | Procyanidin A1 | ✓ | ✓ | ||||
| Flavonols | |||||||||||
| 13 | 1.58 | 397.0781 | 303, 301, 273 | C15H14O9 | Quercetin dihydrate | ✓ | |||||
| 14 | 2.284 | 789.2055 | 591, 489 | C33H40O22 | Quercetin-3-O-beta-D-glucose-7-O-beta-D-gentiobiosiden | ✓ | |||||
| 15 | 1.567 | 447.4016 | 285 | C21H18O12 | Kaempferol-3-glucuronide | ✓ | ✓ | ||||
| 16 | 20.865 | 595.1632 | 287, 285, 449 | C27H30O15 | Kaempferol 3-O-robinobioside | ✓ | |||||
| 17 | 15.968 | 625.1758 | 314 | C28H32O16 | Isorhamnetin-3-O-neohesperidoside | ✓ | ✓ | ||||
| 18 | 16.206 | 553.0848 | 493 | C21H18O14 | Hibifolin | ✓ | |||||
| 19 | 7.151 | 497.1851 | 438 | C25H26O7 | Papyriflavonol A | ✓ | |||||
| 20 | 1.679 | 677.2371 | 351 | C33H40O15 | Sagittatoside A | ✓ | |||||
| Flavanones | |||||||||||
| 21 | 3.37 | 633.1822 | 494, 465, 431 | C28H34O15 | Hesperidin | ✓ | ✓ | ||||
| 22 | 6.772 | 271.064 | 177,151, 107 | C15H12O5 | Naringenin | ✓ | ✓ | ||||
| 23 | 7.3 | 373.1289 | 181, 161 | C18H18O5 | Naringenin trimethyl ether | ✓ | |||||
| 24 | 5.147 | 331.0828 | 193, 93 | C16H14O5 | 5-O-Methylnaringenin | ✓ | |||||
| 25 | 29.446 | 467.1715 | 367, 45, 123 | C25H26O6 | Kuwanol C | ✓ | |||||
| Flavones | |||||||||||
| 26 | 15.971 | 615.1728 | 593 | C28H32O14 | Fortunellin | ✓ | |||||
| 27 | 7.301 | 395.1096 | 342, 357 | C20H20O7 | Tangeretin | ✓ | |||||
| 28 | 6.347 | 553.1154 | 521, 477 | C31H20O10 | Bilobetin | ✓ | ✓ | ✓ | |||
| 29 | 2.374 | 289.0703 | 271, 243, 153 | C15H12O6 | Aromadendrin | ✓ | |||||
| 30 | 2.469 | 463.0875 | 162 | C21H20O12 | 6-Hydroxyluteolin-7-glucoside | ✓ | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, M.; He, J.; Gu, Y.; Wu, G.; Tan, L.; Li, C.; Xu, F.; Zhu, K. Changes in Phenolic Compounds and Antioxidant Capacity of Artocarpus heterophyllus Lam. (Jackfruit) Pulp during In Vitro Gastrointestinal Digestion. Antioxidants 2024, 13, 37. https://doi.org/10.3390/antiox13010037
Cheng M, He J, Gu Y, Wu G, Tan L, Li C, Xu F, Zhu K. Changes in Phenolic Compounds and Antioxidant Capacity of Artocarpus heterophyllus Lam. (Jackfruit) Pulp during In Vitro Gastrointestinal Digestion. Antioxidants. 2024; 13(1):37. https://doi.org/10.3390/antiox13010037
Chicago/Turabian StyleCheng, Ming, Jiali He, Yu Gu, Gang Wu, Lehe Tan, Chuan Li, Fei Xu, and Kexue Zhu. 2024. "Changes in Phenolic Compounds and Antioxidant Capacity of Artocarpus heterophyllus Lam. (Jackfruit) Pulp during In Vitro Gastrointestinal Digestion" Antioxidants 13, no. 1: 37. https://doi.org/10.3390/antiox13010037
APA StyleCheng, M., He, J., Gu, Y., Wu, G., Tan, L., Li, C., Xu, F., & Zhu, K. (2024). Changes in Phenolic Compounds and Antioxidant Capacity of Artocarpus heterophyllus Lam. (Jackfruit) Pulp during In Vitro Gastrointestinal Digestion. Antioxidants, 13(1), 37. https://doi.org/10.3390/antiox13010037