


Indian Almond (Terminalia catappa Linn.) Leaf Extract Extends Lifespan by Improving Lipid Metabolism and Antioxidant Activity Dependent on AMPK Signaling Pathway in Caenorhabditis elegans under High-Glucose-Diet Conditions

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. LC-MS/MS Analysis
2.3. Total Phenol Contents (TPC) and Total Flavonoid Contents (TFC)
2.4. Radical Scavenging Capacity
2.5. Worm Study
2.5.1. Cultivation
2.5.2. Acute Toxicity
2.5.3. Oil Red O Staining
2.5.4. Triglyceride (TG) Assay
2.5.5. Determination of Stress Resistance
2.5.6. DCF–DA Assay
2.5.7. Daf-16 Nuclear Localization
2.5.8. Lifespan Assay
2.5.9. Statistical Analysis
3. Results
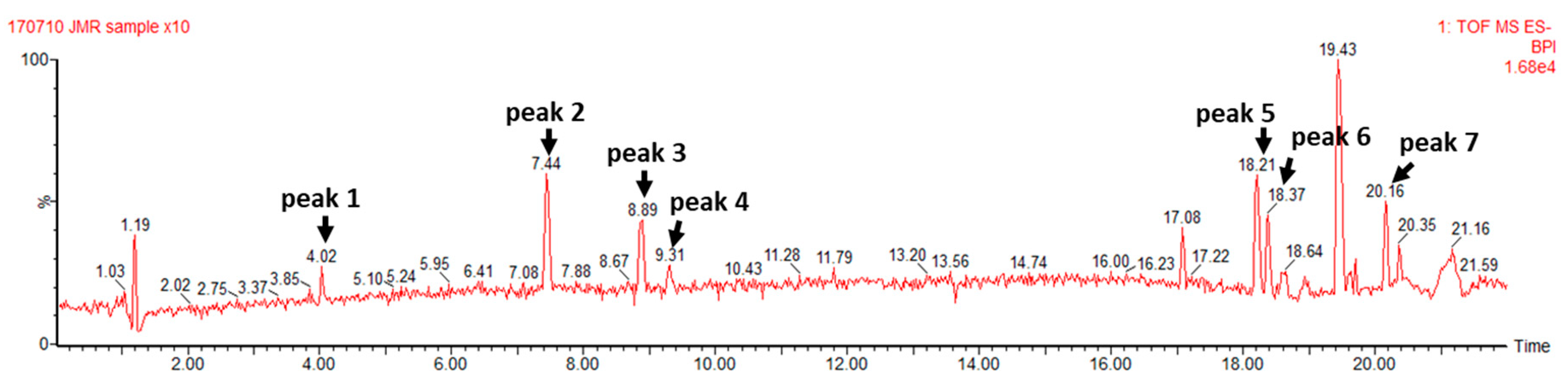
3.1. TCE Components Profile
3.2. Total Phenol and Total Flavonoid Contents and Antioxidant Activity of TCE
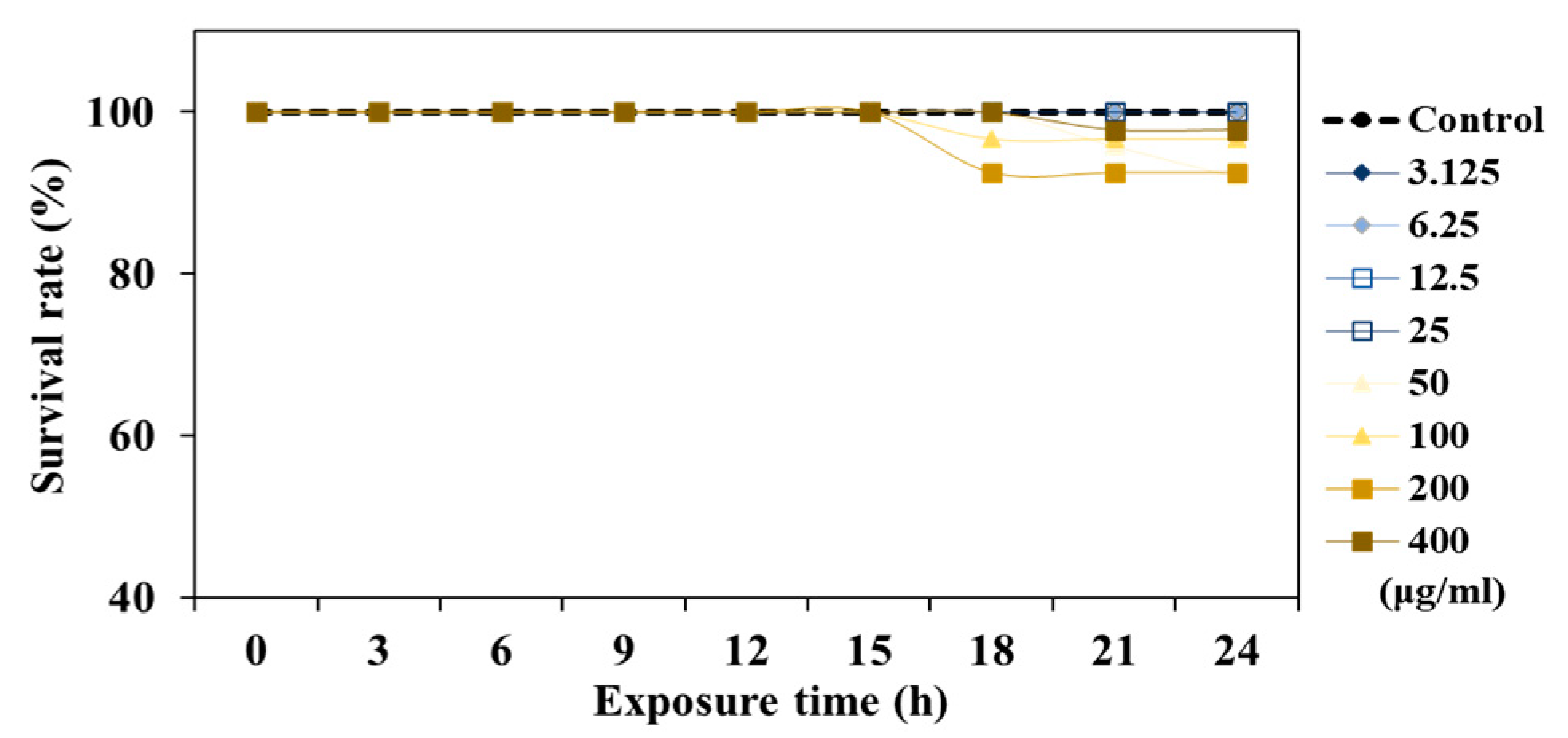
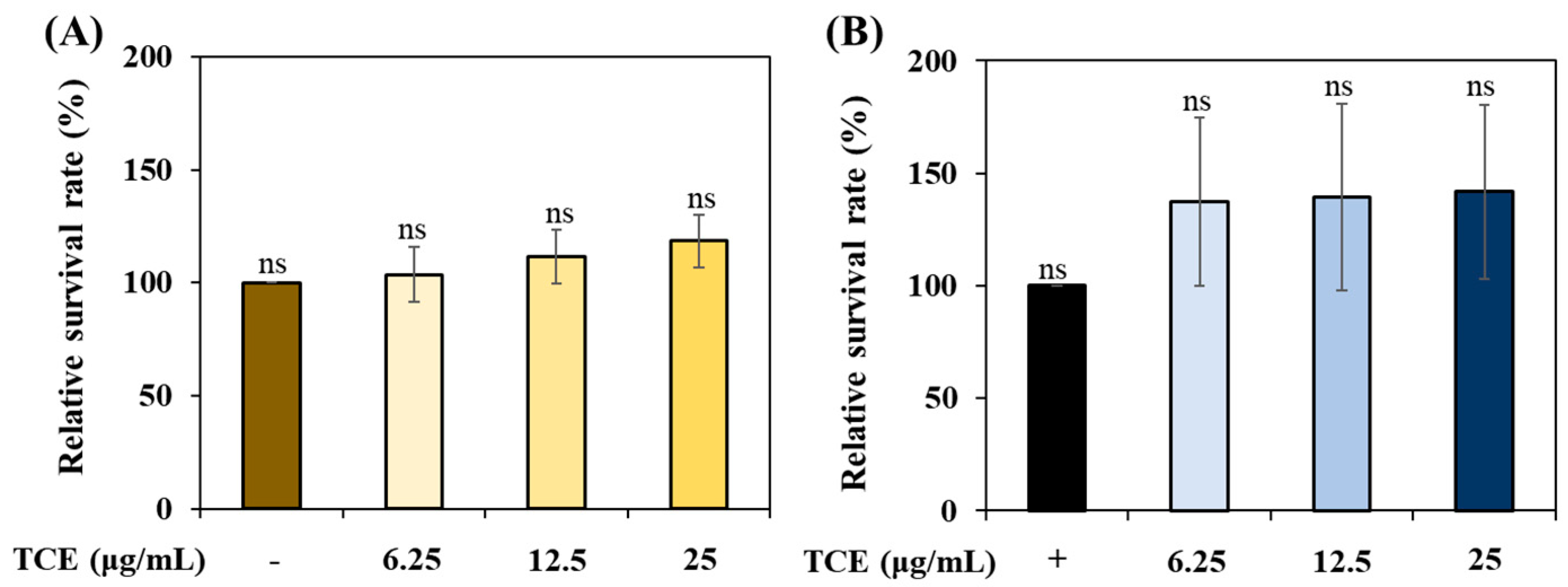
3.3. Safety of TCE
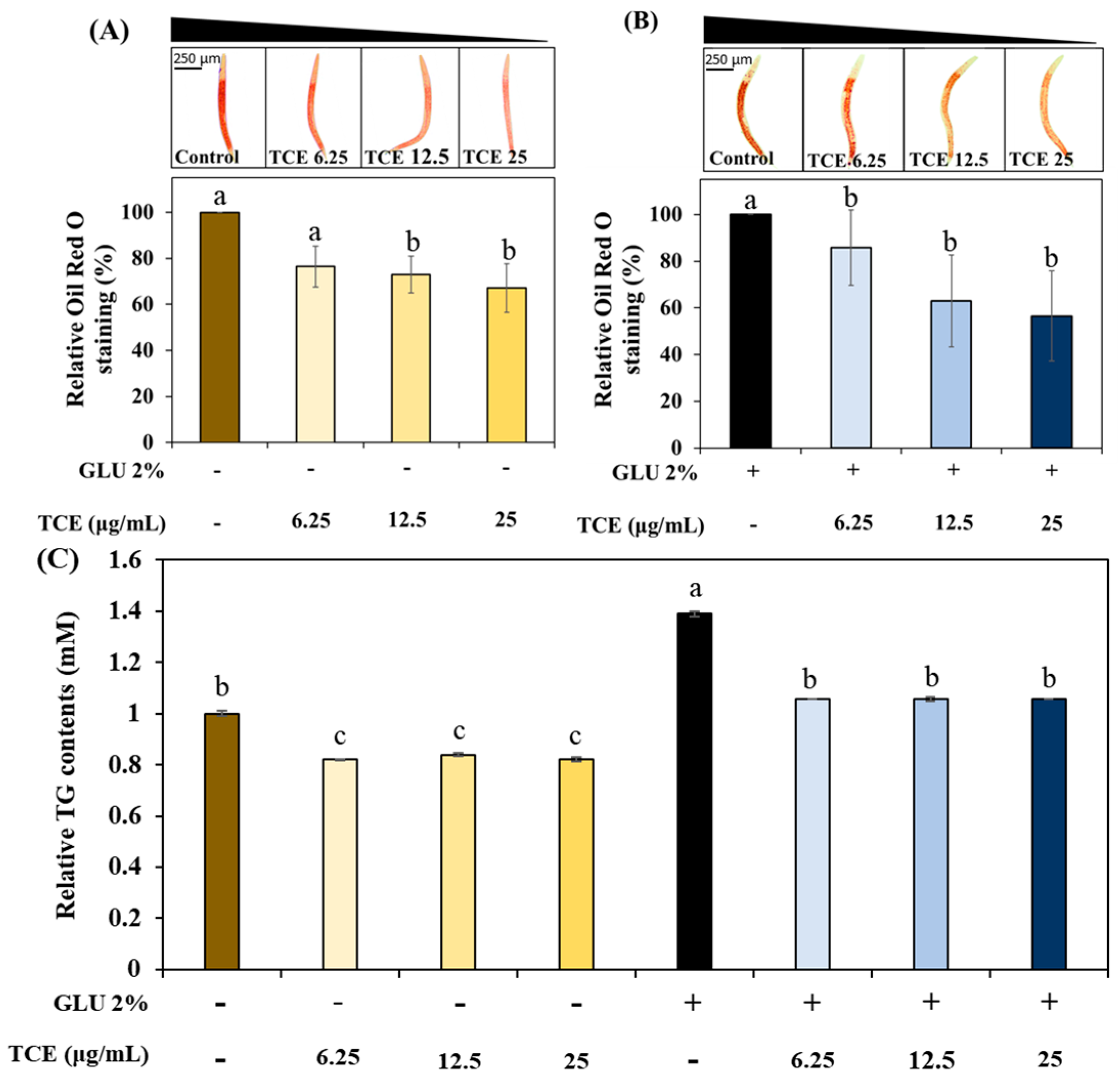
3.4. Inhibitory Effect on Lipid Accumulation of TCE
3.5. Evaluation of TCE Stress Resistance under Various Stress Conditions
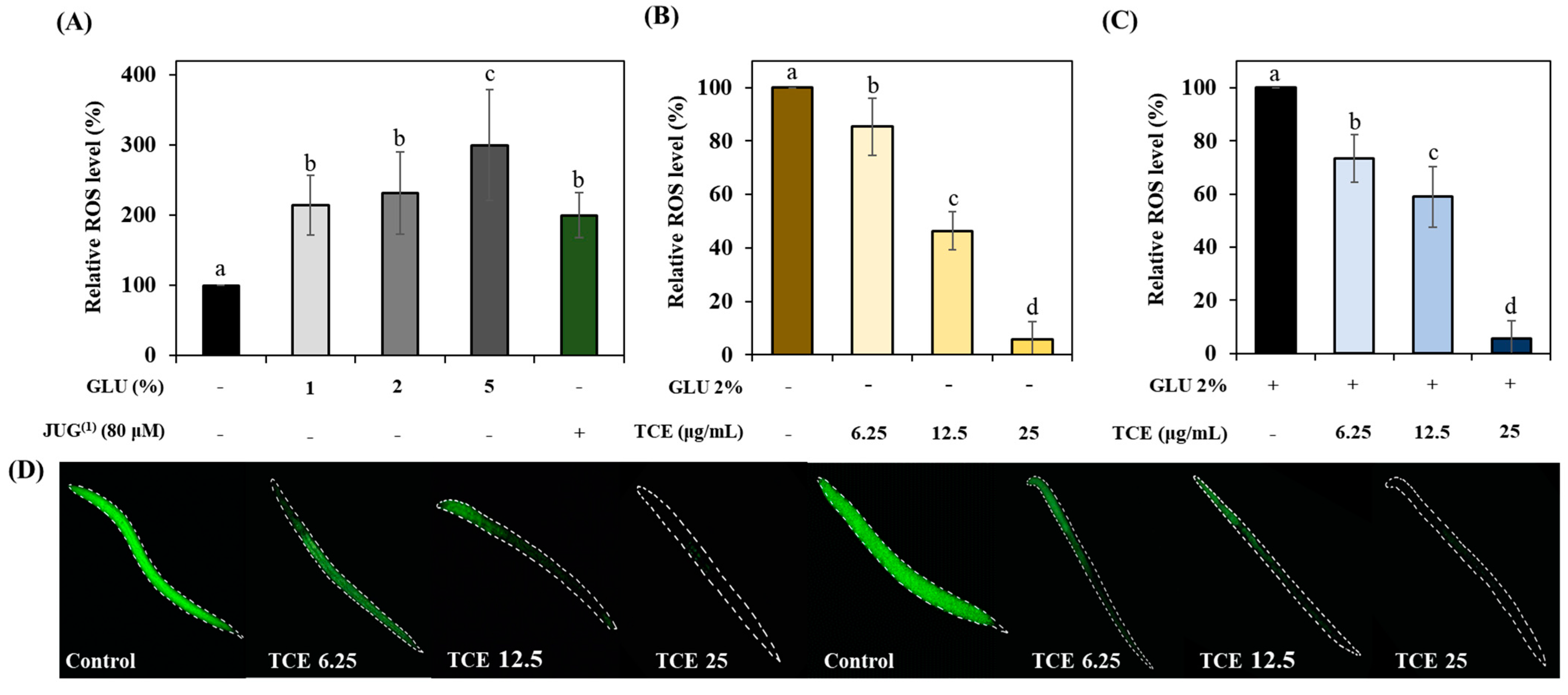
3.6. Inhibitory Effects on Glucose-Induced ROS Accumulation of TCE
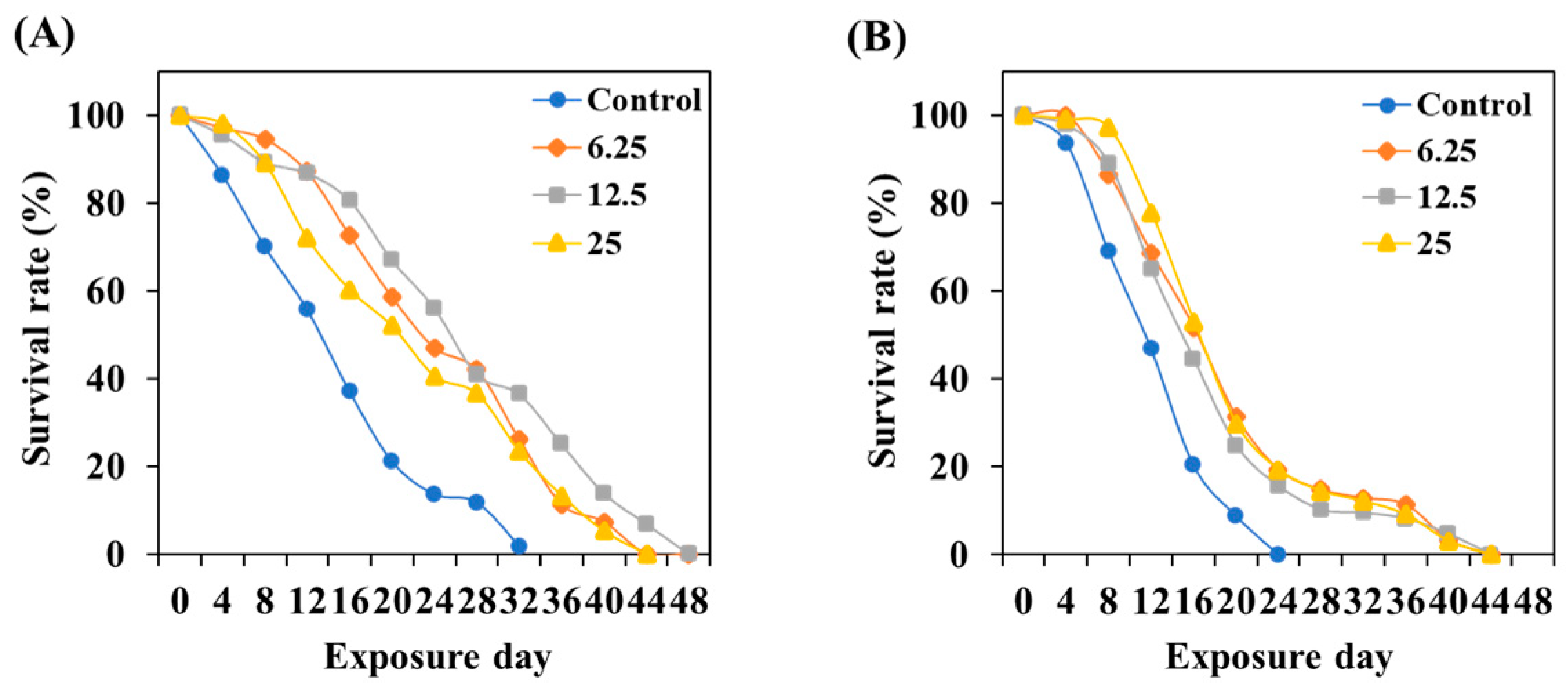
3.7. TCE Effect on Lifespan Prolonging
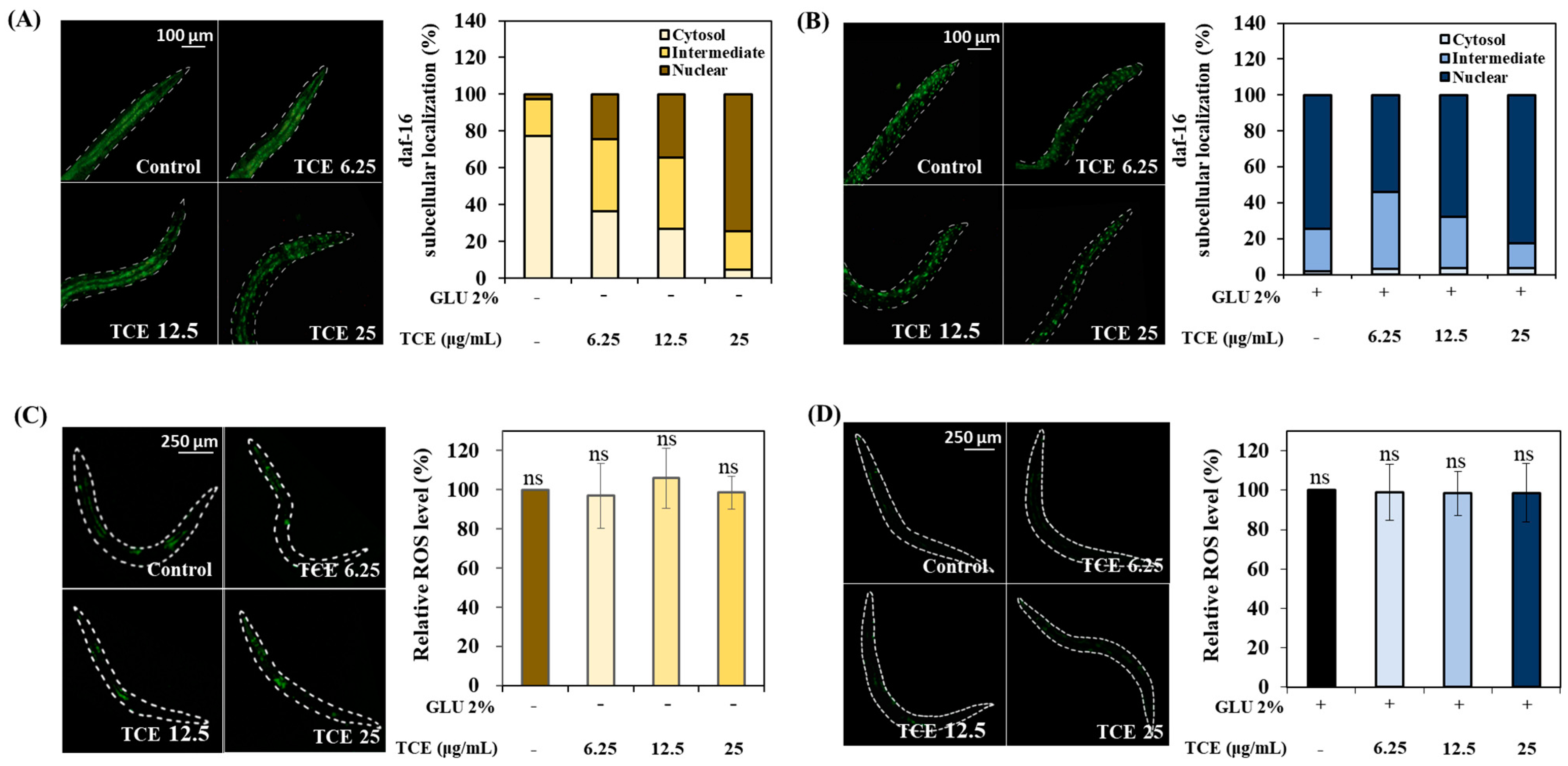
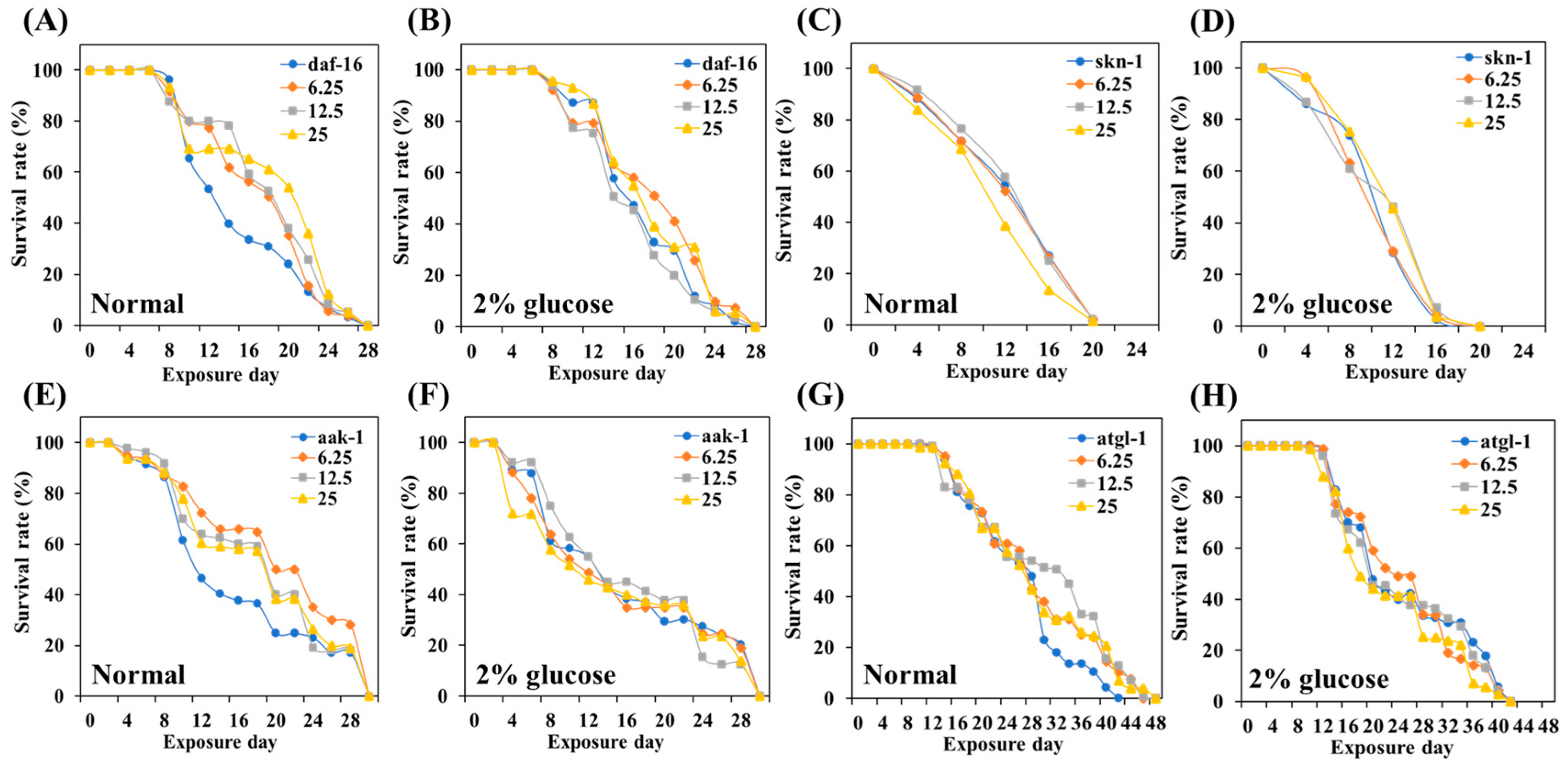
3.8. Role of Daf-16 in TCE Effect
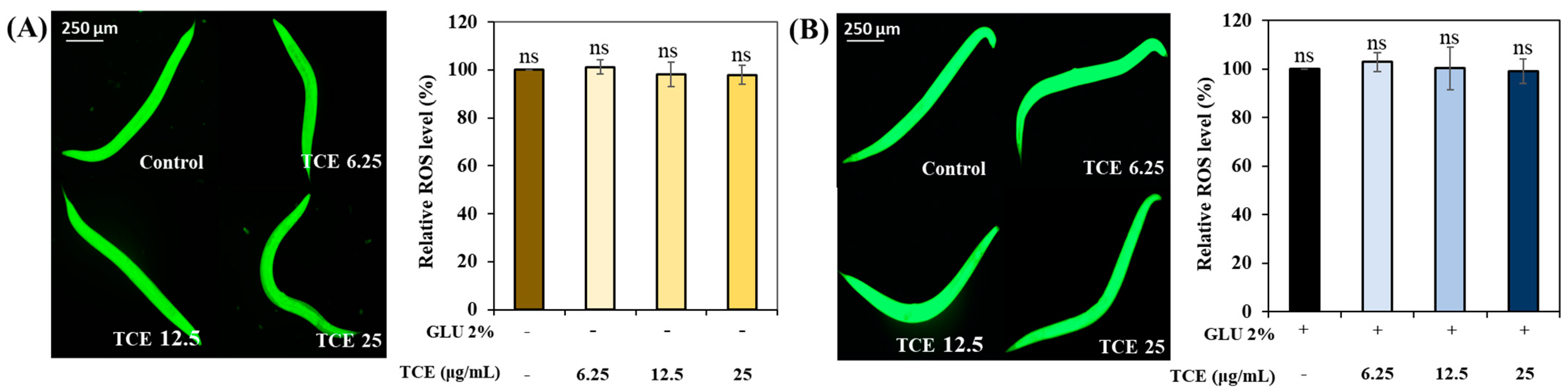
3.9. Role of Skn-1 in TCE Effect
3.10. The Role of Various Metabolic Stress Regulators in the Lifespan-Extending Effect of TCE
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pi-Sunyer, X. The medical risks of obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Francisqueti, F.V.; Nascimento, A.F.; Minatel, I.O.; Dias, M.C.; Luvizotto, R.d.A.M.; Berchieri-Ronchi, C.; Ferreira, A.L.A.; Corrêa, C.R. Metabolic syndrome and inflammation in adipose tissue occur at different times in animals submitted to a high-sugar/fat diet. J. Nutr. Sci. 2017, 6, e41. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.T.d.; Fernandes, I.d.C.; Sousa, G.G.d.; Santos, T.A.P.d.; Paiva, N.C.N.d.; Carneiro, C.M.; Evangelista, E.A.; Barboza, N.R.; Guerra-Sá, R. High-sugar diet leads to obesity and metabolic diseases in ad libitum-fed rats irrespective of caloric intake. Arch. Endocrinol. Metab. 2020, 64, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Segal, M.S.; Sautin, Y.; Nakagawa, T.; Feig, D.I.; Kang, D.-H.; Gersch, M.S.; Benner, S.; Sánchez-Lozada, L.G. Potential role of sugar (fructose) in the epidemic of hypertension, obesity and the metabolic syndrome, diabetes, kidney disease, and cardiovascular disease. Am. J. Clin. Nutr. 2007, 86, 899–906. [Google Scholar] [PubMed]
- Wormbook. Available online: http://www.wormbook.org/chapters/www_feeding/feeding.html (accessed on 21 May 2012).
- McKay, R.M.; McKay, J.P.; Avery, L.; Graff, J.M. C. elegans: A model for exploring the genetics of fat storage. Dev. Cell 2003, 4, 131–142. [Google Scholar] [CrossRef]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef]
- Dwyer, D.S.; Donohoe, D.; Lu, X.H.; Aamodt, E.J. Mechanistic connections between glucose/lipid disturbances and weight gain induced by antipsychotic drugs. Int. Rev. Neurobiol. 2005, 65, 211–247. [Google Scholar]
- Zheng, J.; Enright, F.; Keenan, M.; Finley, J.; Zhou, J.; Ye, J.; Greenway, F.; Senevirathne, R.N.; Gissendanner, C.R.; Manaois, R. Resistant starch, fermented resistant starch, and short-chain fatty acids reduce intestinal fat deposition in Caenorhabditis elegans. J. Agric. Food Chem. 2010, 58, 4744–4748. [Google Scholar] [CrossRef]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–283. [Google Scholar] [CrossRef]
- Bai, J.; Zhu, Y.; He, L.; Zhang, J.; Li, J.; Pan, R.; Zhang, J.; Zhao, Y.; Cui, L.; Lu, H. Saponins from bitter melon reduce lipid accumulation via induction of autophagy in C. elegans and HepG2 cell line. Curr. Res. Food Sci. 2022, 5, 1167–1175. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res. Rev. 2012, 11, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; Roy, R. 5′-AMP-Activated Protein Kinase Signaling in Caenorhabditis elegans. AMP-Act. Protein Kinase 2016, 107, 375–388. [Google Scholar]
- Greer, E.L.; Dowlatshahi, D.; Banko, M.R.; Villen, J.; Hoang, K.; Blanchard, D.; Gygi, S.P.; Brunet, A. An AMPK-FOXO pathway mediates longevity induced by a novel method of dietary restriction in C. elegans. Curr. Biol. 2007, 17, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Ankeny, R.A. The natural history of Caenorhabditis elegans research. Nat. Rev. Genet. 2001, 2, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Fatt, H.V.; Dougherty, E.C. Genetic control of differential heat tolerance in two strains of the nematode Caenorhabditis elegans. Science 1963, 141, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Nigon, V.; Dougherty, E.C. Reproductive patterns and attempts at reciprocal crossing of Rhabditis elegans Maupas, 1900, and Rhabditis briggsae Dougherty and Nigon, 1949 (Nematoda: Rhabditidae). J. Exp. Zool. 1949, 112, 485–503. [Google Scholar] [CrossRef]
- Kosztelnik, M.; Kurucz, A.; Papp, D.; Jones, E.; Sigmond, T.; Barna, J.; Traka, M.H.; Lorincz, T.; Szarka, A.; Banhegyi, G. Suppression of AMPK/aak-2 by NRF2/SKN-1 down-regulates autophagy during prolonged oxidative stress. FASEB J. 2019, 33, 2372. [Google Scholar] [CrossRef]
- Zhang, H.; Davies, K.J.; Forman, H.J. Oxidative stress response and Nrf2 signaling in aging. Free Radic. Biol. Med. 2015, 88, 314–336. [Google Scholar] [CrossRef]
- Chen, W.-L.; Chen, Y.-L.; Chiang, Y.-M.; Wang, S.-G.; Lee, H.-M. Fenofibrate lowers lipid accumulation in myotubes by modulating the PPARα/AMPK/FoxO1/ATGL pathway. Biochem. Pharmacol. 2012, 84, 522–531. [Google Scholar] [CrossRef]
- Anand, A.; Divya, N.; Kotti, P. An updated review of Terminalia catappa. Pharmacogn. Rev. 2015, 9, 93. [Google Scholar] [CrossRef]
- Abiodun, O.O.; Rodríguez-Nogales, A.; Algieri, F.; Gomez-Caravaca, A.M.; Segura-Carretero, A.; Utrilla, M.P.; Rodriguez-Cabezas, M.E.; Galvez, J. Antiinflammatory and immunomodulatory activity of an ethanolic extract from the stem bark of Terminalia catappa L. (Combretaceae): In vitro and in vivo evidences. J. Ethnopharmacol. 2016, 192, 309–319. [Google Scholar] [CrossRef]
- Fan, Y.; Xu, L.; Gao, J.; Wang, Y.; Tang, X.; Zhao, X.; Zhang, Z. Phytochemical and antiinflammatory studies on Terminalia catappa. Fitoterapia 2004, 75, 253–260. [Google Scholar] [CrossRef]
- Ratnasooriya, W.; Dharmasiri, M.; Rajapakse, R.; De Silva, M.; Jayawardena, S.; Fernando, P.; De Silva, W.; Nawela, A.; Warusawithana, R.; Jayakody, J. Tender leaf extract of Terminalia catappa antinociceptive activity in rats. Pharm. Biol. 2002, 40, 60–66. [Google Scholar] [CrossRef]
- Liu, T.-Y.; Ho, L.-K.; Tsai, Y.-C.; Chiang, S.-H.; Chao, T.-W.; Li, J.-H.; Chi, C.-W. Modification of mitomycin C-induced clastogenicity by Terminalia catappa L. in vitro and in vivo. Cancer Lett. 1996, 105, 113–118. [Google Scholar] [CrossRef]
- Nair, R.; Chanda, S. Antimicrobial activity of Terminalia catappa, Manilkara zapota and Piper betel leaf extract. Indian J. Pharm. Sci. 2008, 70, 390. [Google Scholar]
- Yang, S.-F.; Chen, M.-K.; Hsieh, Y.-S.; Yang, J.-S.; Zavras, A.-I.; Hsieh, Y.-H.; Su, S.-C.; Kao, T.-Y.; Chen, P.-N.; Chu, S.-C. Antimetastatic effects of Terminalia catappa L. on oral cancer via a down-regulation of metastasis-associated proteases. Food Chem. Toxicol. 2010, 48, 1052–1058. [Google Scholar] [CrossRef]
- Wen, K.-C.; Shih, I.; Hu, J.-C.; Liao, S.-T.; Su, T.-W.; Chiang, H.-M. Inhibitory effects of Terminalia catappa on UVB-induced photodamage in fibroblast cell line. Evid.-Based Complement. Altern. Med. 2010, 2011, 904532. [Google Scholar]
- Jang, M.; Choi, H.Y.; Kim, G.H. Phenolic components rich ethyl acetate fraction of Orostachys japonicus inhibits lipid accumulation by regulating reactive oxygen species generation in adipogenesis. J. Food Biochem. 2019, 43, e12939. [Google Scholar] [CrossRef]
- Gullon, B.; Pintado, M.E.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. In vitro gastrointestinal digestion of pomegranate peel (Punica granatum) flour obtained from co-products: Changes in the antioxidant potential and bioactive compounds stability. J. Funct. Foods 2015, 19, 617–628. [Google Scholar] [CrossRef]
- Li, Z.; Lan, Y.; Miao, J.; Chen, X.; Chen, B.; Liu, G.; Wu, X.; Zhu, X.; Cao, Y. Phytochemicals, antioxidant capacity and cytoprotective effects of jackfruit (Artocarpus heterophyllus Lam.) axis extracts on HepG2 cells. Food Biosci. 2021, 41, 100933. [Google Scholar] [CrossRef]
- Pandey, S.; Tiwari, S.; Kumar, A.; Niranjan, A.; Chand, J.; Lehri, A.; Chauhan, P.S. Antioxidant and anti-aging potential of Juniper berry (Juniperus communis L.) essential oil in Caenorhabditis elegans model system. Ind. Crops Prod. 2018, 120, 113–122. [Google Scholar] [CrossRef]
- Bai, J.; Farias-Pereira, R.; Jang, M.; Zhang, Y.; Lee, S.M.; Kim, Y.-S.; Park, Y.; Ahn, J.B.; Kim, G.-H.; Kim, K.-H. Azelaic acid promotes caenorhabditis elegans longevity at low temperature via an increase in fatty acid desaturation. Pharm. Res. 2021, 38, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Rathor, L.; Pant, A.; Nagar, A.; Tandon, S.; Trivedi, S.; Pandey, R. Trachyspermum ammi L. (Carom) oil induces alterations in SOD-3, GST-4 expression and prolongs lifespan in Caenorhabditis elegans. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 1355–1362. [Google Scholar] [CrossRef]
- Steinberg, H.O.; Chaker, H.; Leaming, R.; Johnson, A.; Brechtel, G.; Baron, A.D. Obesity/insulin resistance is associated with endothelial dysfunction. Implications for the syndrome of insulin resistance. J. Clin. Investig. 1996, 97, 2601–2610. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ballantyne, C.M. Metabolic inflammation and insulin resistance in obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef] [PubMed]
- AsghariHanjani, N.; Vafa, M. The role of IGF-1 in obesity, cardiovascular disease, and cancer. Med. J. Islam. Repub. Iran 2019, 33, 56. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645. [Google Scholar] [CrossRef]
- Koliaki, C.; Liatis, S.; Kokkinos, A. Obesity and cardiovascular disease: Revisiting an old relationship. Metabolism 2019, 92, 98–107. [Google Scholar] [CrossRef]
- Zhang, H.; Qin, J.; Lan, X.; Zeng, W.; Zhou, J.; Huang, T.-E.; Xiao, W.-L.; Wang, Q.-Q.; Sun, S.; Su, W. Handelin extends lifespan and healthspan of Caenorhabditis elegans by reducing ROS generation and improving motor function. Biogerontology 2022, 23, 115–128. [Google Scholar] [CrossRef]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; de Bittencourt, P.I.H., Jr. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Zhang, Z.; Ko, S.H.; Fisher, A.L.; Liu, Z.; Chen, L. Microtubule regulators act in the nervous system to modulate fat metabolism and longevity through DAF-16 in C. elegans. Aging Cell 2019, 18, e12884. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, T.K.; Steinbaugh, M.J.; Hourihan, J.M.; Ewald, C.Y.; Isik, M. SKN-1/Nrf, stress responses, and aging in Caenorhabditis elegans. Free Radic. Biol. Med. 2015, 88, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.-L.; Zheng, S.-Q.; Wu, G.-S.; Luo, H.-R. Aspirin extends the lifespan of Caenorhabditis elegans via AMPK and DAF-16/FOXO in dietary restriction pathway. Exp. Gerontol. 2013, 48, 499–506. [Google Scholar] [CrossRef]
- Wang, S.; Yi, X.; Wu, Z.; Guo, S.; Dai, W.; Wang, H.; Shi, Q.; Zeng, K.; Guo, W.; Li, C. CAMKK2 defines ferroptosis sensitivity of melanoma cells by regulating AMPK—NRF2 pathway. J. Investig. Dermatol. 2022, 142, 189–200.e188. [Google Scholar] [CrossRef]
- Koyama, T.; Nakajima, C.; Nighimoto, S.; Takami, M.; Woo, J.-T.; Yazawa, K. Suppressive effects of the leaf of Terminalia catappa L. on osteoclast differentiation in vitro and bone weight loss in vivo. J. Nutr. Sci. Vitaminol. 2012, 58, 129–135. [Google Scholar] [CrossRef][Green Version]
- Jang, M.; Choi, S.I. Schisandrin C isolated from Schisandra chinensis fruits inhibits lipid accumulation by regulating adipogenesis and lipolysis through AMPK signaling in 3T3-L1 adipocytes. J. Food Biochem. 2022, 46, e14454. [Google Scholar] [CrossRef]
- Wang, K.; Chen, S.; Zhang, C.; Huang, J.; Wu, J.; Zhou, H.; Jin, L.; Qian, X.; Jin, J.; Lyu, J. Enhanced ROS production leads to excessive fat accumulation through DAF-16 in Caenorhabditis elegans. Exp. Gerontol. 2018, 112, 20–29. [Google Scholar] [CrossRef]
- Bai, J.; Farias-Pereira, R.; Zhang, Y.; Jang, M.; Park, Y.; Kim, K.H. C. elegans ACAT regulates lipolysis and its related lifespan in fasting through modulation of the genes in lipolysis and insulin/IGF-1 signaling. BioFactors 2020, 46, 754–765. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LC Condition (Waters® ACQUITYTM UPLC) | |
|---|---|
| Column | CORTECSTM UPLC® C18 1.6 μm (2.1 × 100 mm) |
| Temperature: 35 °C | |
| Mobile phase | (A) 0.1% formic acid in water and (B) 0.1% formic acid in MethanolGradient: 5% solvent B for 1 min, 5–30% solvent B for the next 4 min, 30–32% solvent B for 2 min, 32–50% solvent B for 7 min, 50–90% solvent B for 4 min, 90% solvent B for 2 min, and a linear step of 90-2% solvent B for 3 min. |
| Flow rate | 0.25 mL/min |
| MS Condition (SYNAPTTM G2) | |
| Ionization Mode | ESI- |
| Temperature | Source: 120 °C, and desolvation: 350 °C |
| Voltage | Capillary: 2.5 kV, sampling cone: 35 V, extraction cone: 4.0 V |
| Gas flow | Cone gas: 100 L/h, and desolvation gas: 800 L/h |
| TPC (1) | TFC (2) | DPPH | ABTS | ||
|---|---|---|---|---|---|
| IC50 (3) (μg/mL) | μM TE (4)/g | IC50 (μg/mL) | μM TE/g | ||
| 0.694 ± 0.06 | 0.268 ± 0.011 | 18.816 ± 0.038 | 84.326 ± 0.228 | 125.773 ± 0.465 | 2.227 ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Lee, S.-b.; Cho, M.; Choe, S.; Jang, M. Indian Almond (Terminalia catappa Linn.) Leaf Extract Extends Lifespan by Improving Lipid Metabolism and Antioxidant Activity Dependent on AMPK Signaling Pathway in Caenorhabditis elegans under High-Glucose-Diet Conditions. Antioxidants 2024, 13, 14. https://doi.org/10.3390/antiox13010014
Kim Y, Lee S-b, Cho M, Choe S, Jang M. Indian Almond (Terminalia catappa Linn.) Leaf Extract Extends Lifespan by Improving Lipid Metabolism and Antioxidant Activity Dependent on AMPK Signaling Pathway in Caenorhabditis elegans under High-Glucose-Diet Conditions. Antioxidants. 2024; 13(1):14. https://doi.org/10.3390/antiox13010014
Chicago/Turabian StyleKim, Yebin, Seul-bi Lee, Myogyeong Cho, Soojin Choe, and Miran Jang. 2024. "Indian Almond (Terminalia catappa Linn.) Leaf Extract Extends Lifespan by Improving Lipid Metabolism and Antioxidant Activity Dependent on AMPK Signaling Pathway in Caenorhabditis elegans under High-Glucose-Diet Conditions" Antioxidants 13, no. 1: 14. https://doi.org/10.3390/antiox13010014
APA StyleKim, Y., Lee, S.-b., Cho, M., Choe, S., & Jang, M. (2024). Indian Almond (Terminalia catappa Linn.) Leaf Extract Extends Lifespan by Improving Lipid Metabolism and Antioxidant Activity Dependent on AMPK Signaling Pathway in Caenorhabditis elegans under High-Glucose-Diet Conditions. Antioxidants, 13(1), 14. https://doi.org/10.3390/antiox13010014