

Pyrroloquinoline Quinone Improves Ram Sperm Quality through Its Antioxidative Ability during Storage at 4 °C

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Extenders
2.2. Semen Collection and Processing
2.3. Evaluation of Sperm Motility by Computer-Assisted Sperm Analysis (CASA) System
2.4. Detection of Sperm Plasma Membrane Integrity and Acrosome Integrity
2.5. Evaluation of Mitochondrial Membrane Potential
2.6. Detection of Sperm Reactive Oxygen Species
2.7. Measure of the Sperm ATP Level
2.8. Measurement of Sperm MDA Levels
2.9. Determination of SOD Activity
2.10. Western Blotting
2.11. Immunoprecipitation
2.12. Sperm Capacitation
2.13. Chlortetracycline (CTC) Analysis
2.14. Sperm–Zona Pellucida (ZP) Binding Capacity
2.15. Statistical Analysis
3. Results
3.1. PQQ Improved Sperm Motility Parameters during Storage at 4 °C
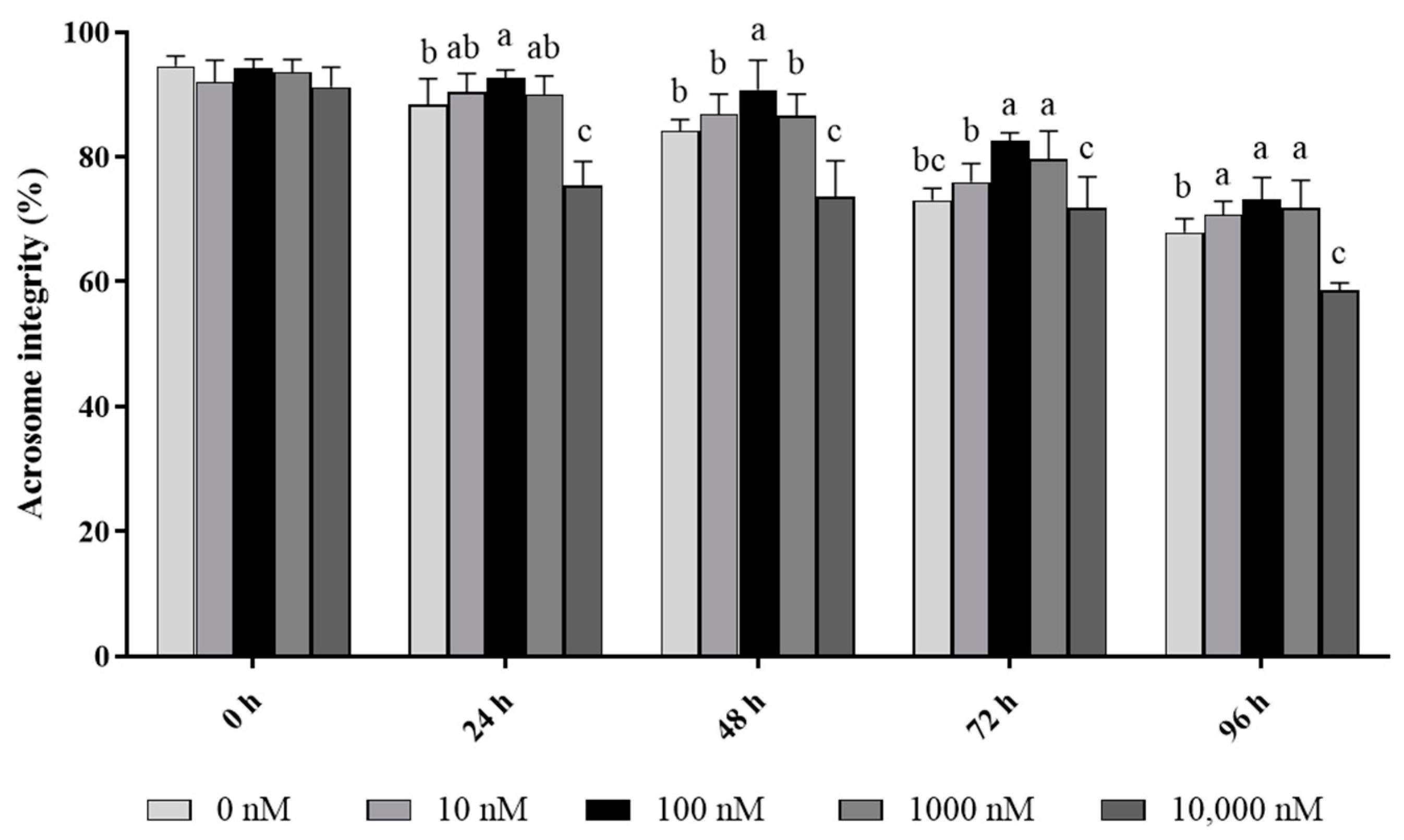
3.2. PQQ Improved Ram Sperm Membrane Integrity and Acrosome Integrity during Storage at 4 °C
3.3. PQQ Improved ram Sperm Mitochondrial Membrane Potentials and ATP Levels during Storage at 4 °C
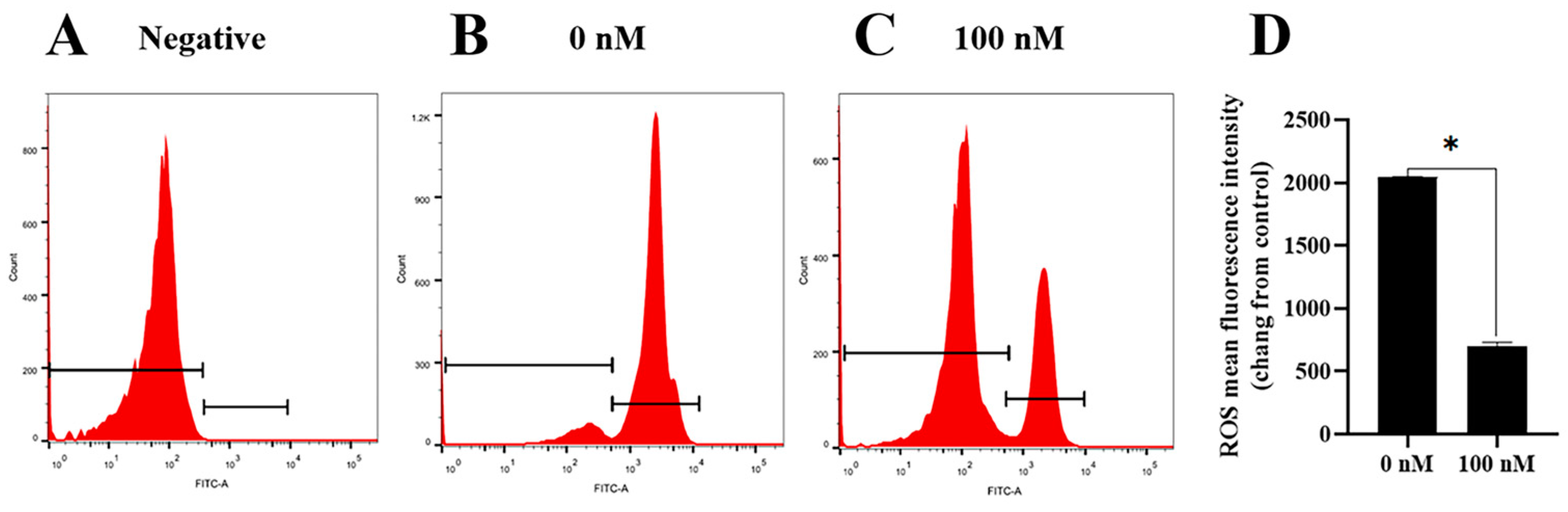
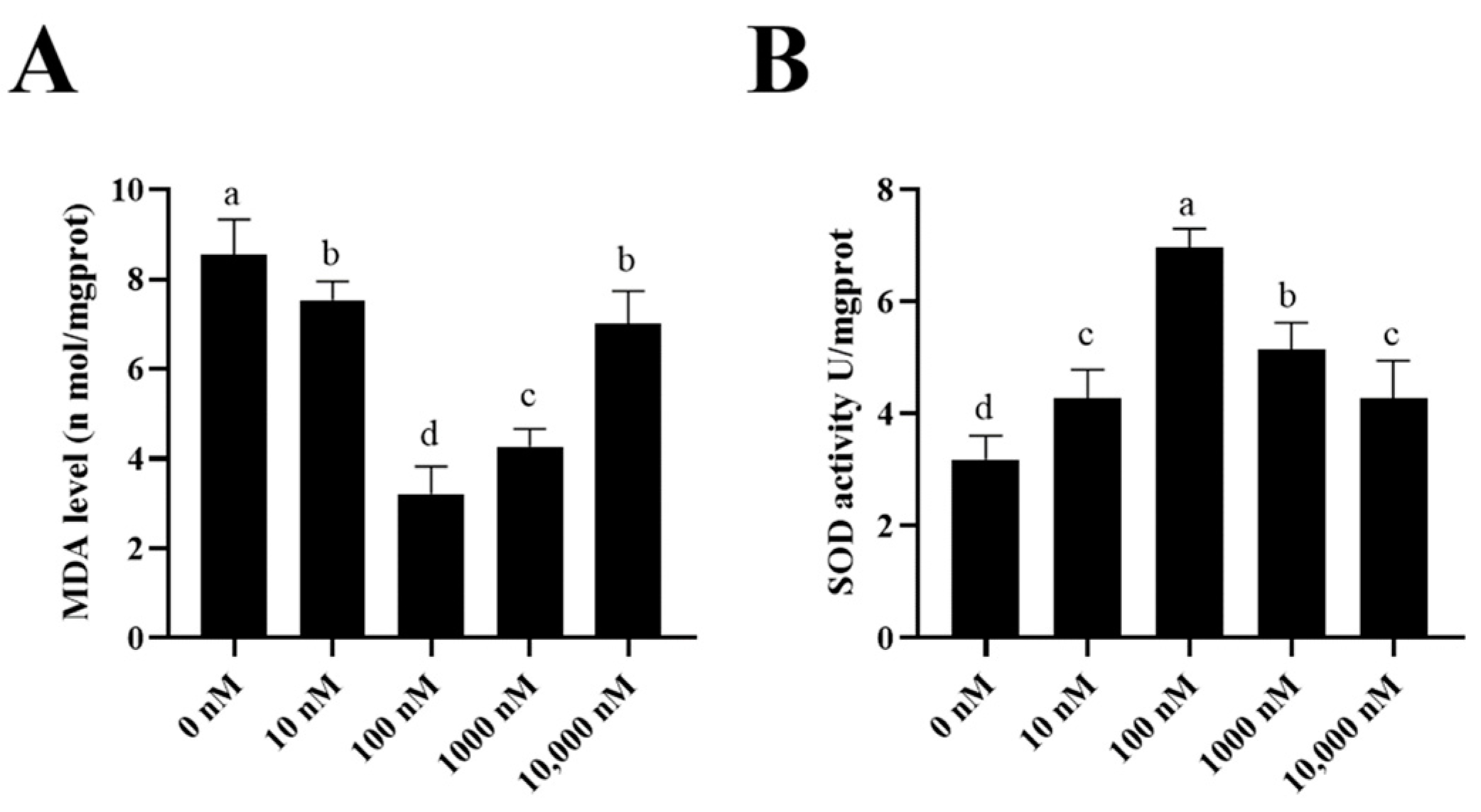
3.4. PQQ Reduced the Ram Sperm Oxidative Stress during Storage at 4 °C
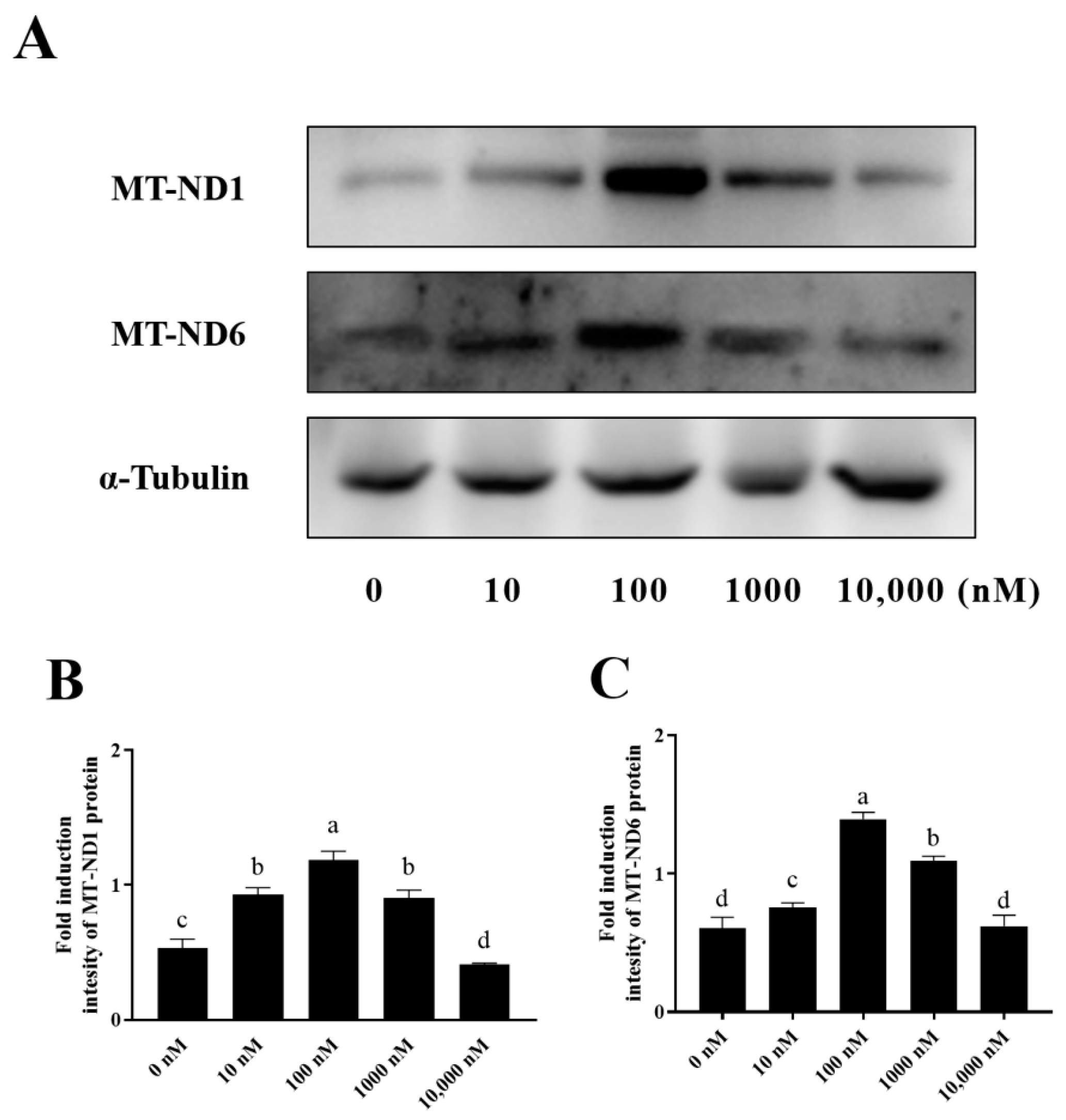
3.5. PQQ Maintained Ram Sperm Mitochondrial Protein Levels by Decreasing ROS-Induced Protein Damage
3.6. The Addition of PQQ Increased Ram Sperm with Capacitation after Inducing Capacitation In Vitro
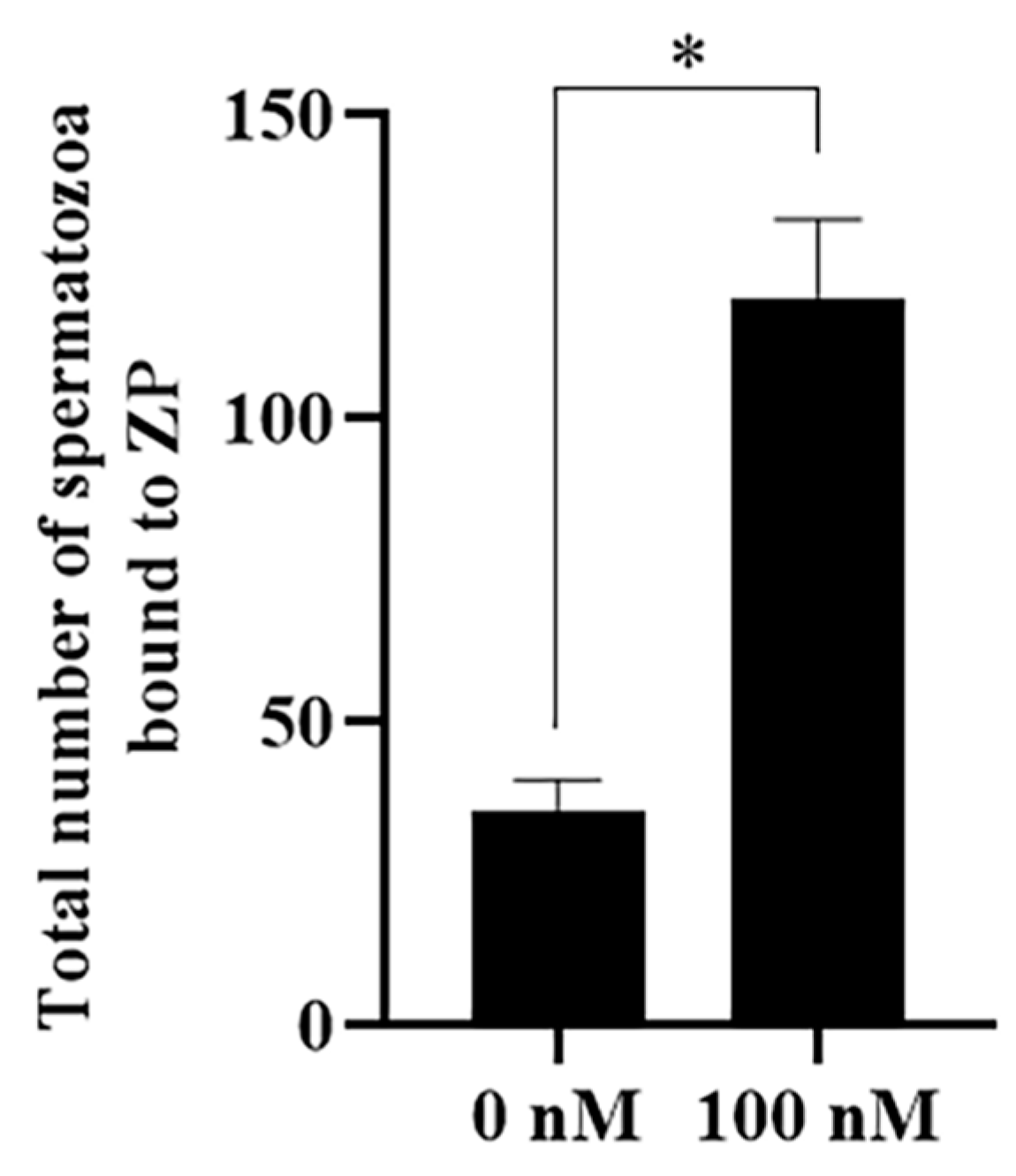
3.7. PQQ Improved Sperm–Zona Pellucida Binding Capacity
4. Discussion
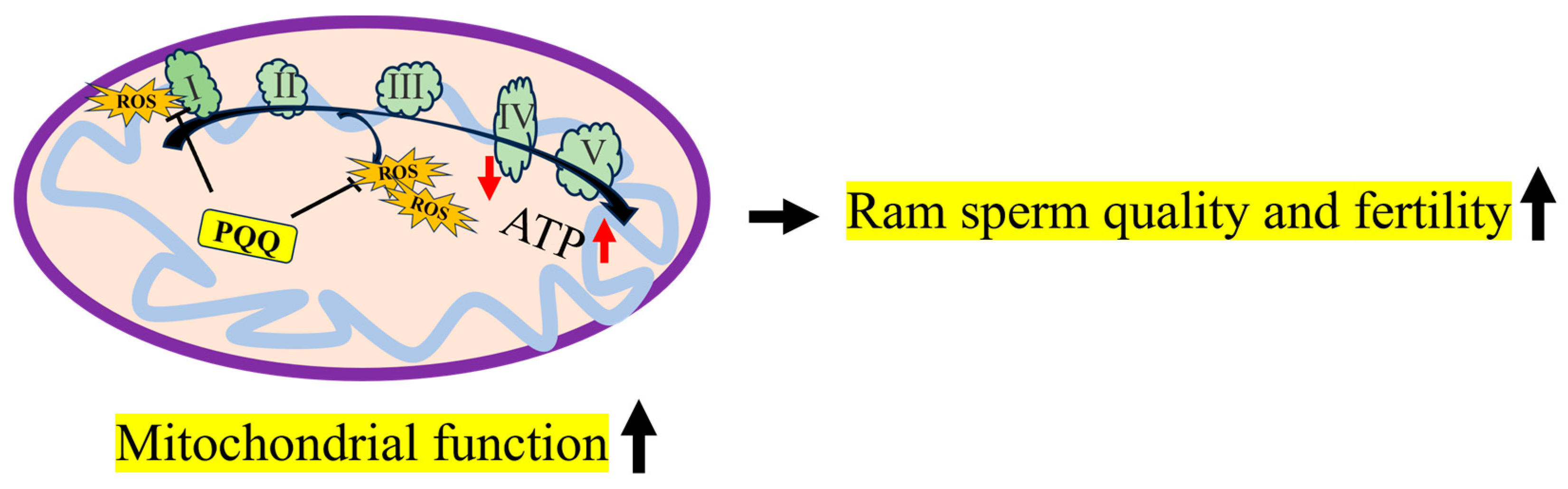
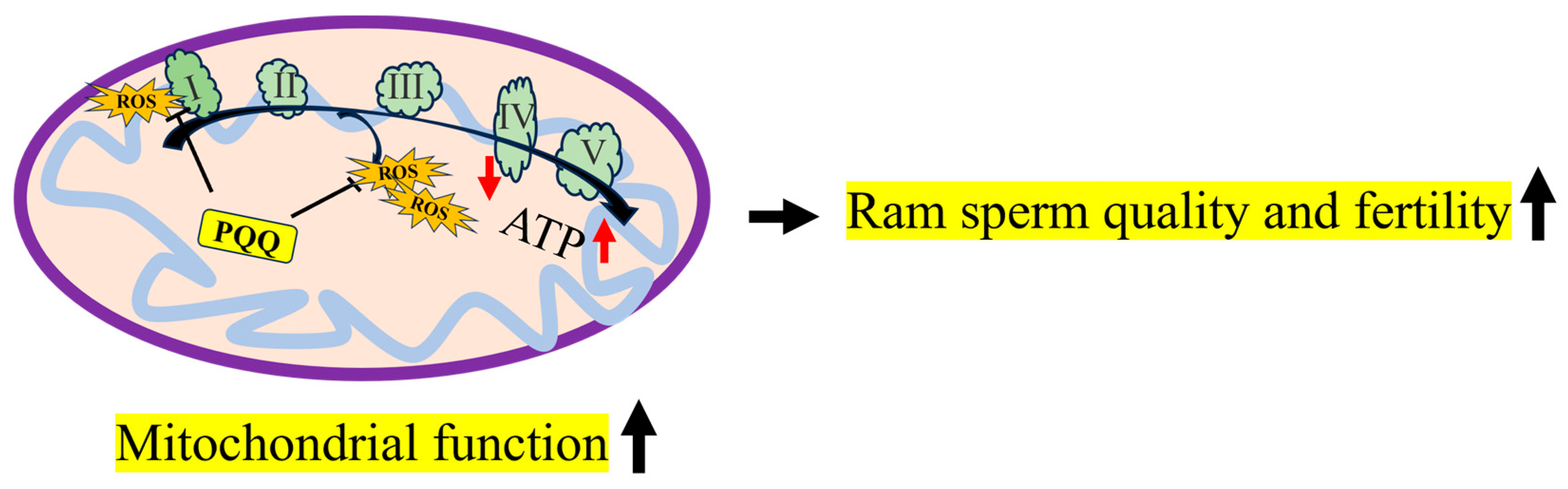
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anel, L.; Alvarez, M.; Martinez-Pastor, F.; Garcia-Macias, V.; Anel, E.; de Paz, P. Improvement strategies in ovine artificial insemination. Reprod. Domest. Anim. 2006, 41 (Suppl. S2), 30–42. [Google Scholar] [CrossRef] [PubMed]
- Salamon, S.; Maxwell, W.M. Storage of ram semen. Anim. Reprod. Sci. 2000, 62, 77–111. [Google Scholar] [CrossRef] [PubMed]
- Mata-Campuzano, M.; Álvarez-Rodríguez, M.; Tamayo-Canul, J.; López-Urueña, E.; de Paz, P.; Anel, L.; Martínez-Pastor, F.; Álvarez, M. Refrigerated storage of ram sperm in presence of Trolox and GSH antioxidants: Effect of temperature, extender and storage time. Anim. Reprod. Sci. 2014, 151, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.S.; Barretto, L.S.; Arruda, R.P.; Perri, S.H.; Mingoti, G.Z. Effect of antioxidants during bovine in vitro fertilization procedures on spermatozoa and embryo development. Reprod. Domest. Anim. 2010, 45, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Makker, K.; Sharma, R. Clinical relevance of oxidative stress in male factor infertility: An update. Am. J. Reprod. Immunol. 2008, 59, 2–11. [Google Scholar] [CrossRef]
- Ashrafi, G.; Schwarz, T.L. The pathways of mitophagy for quality control and clearance of mitochondria. Cell Death Differ. 2013, 20, 31–42. [Google Scholar] [CrossRef]
- Liang, J.; Lv, C.; Yang, H.; Zhang, Y.; Raza, S.H.A.; Wu, G.; Quan, G. Ultrastructural Modification of Ram Sperm Frozen with Cyclohexanediol and Trehalose. Biopreserv. Biobank. 2022, 20, 348–356. [Google Scholar] [CrossRef]
- Jang, H.; Kim, S.J.; Yuk, S.M.; Han, D.S.; Ha, U.S.; Hong, S.H.; Lee, J.Y.; Hwang, T.K.; Hwang, S.Y.; Kim, S.W. Effects of anthocyanin extracted from black soybean seed coat on spermatogenesis in a rat varicocele-induced model. Reprod. Fertil. Dev. 2012, 24, 649–655. [Google Scholar] [CrossRef]
- Chianese, R.; Pierantoni, R. Mitochondrial Reactive Oxygen Species (ROS) Production Alters Sperm Quality. Antioxidants 2021, 10, 92. [Google Scholar] [CrossRef]
- Kocabaş, F.K.; Kocabaş, M.; Aksu, Ö.; Çakir Sahilli, Y. Ascorbic acid ameliorated the sperm quality of rainbow trout (Oncorhynchus mykiss) against arsenic toxicity: Impact on oxidative stress, fertility ability and embryo development. J. Environ. Sci. Health Part C Toxicol. Carcinog. 2022, 40, 119–132. [Google Scholar] [CrossRef]
- Babaei, A.; Asadpour, R.; Mansouri, K.; Sabrivand, A.; Kazemi-Darabadi, S. Lycopene protects sperm from oxidative stress in the experimental varicocele model. Food Sci. Nutr. 2021, 9, 6806–6817. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.K.; Kumar, A.; Gupta, S.; Sharma, P.; Kumar, G.; Sachan, V.; Yadav, B.; Yadav, S.; Saxena, A.; Swain, D.K. Antioxidant additive melatonin in tris-based egg yolk extender improves post-thaw sperm attributes in Hariana bull. Anim. Reprod. Sci. 2023, 251, 107214. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Fan, X.; Zeng, Y.; Wang, L.; Zhu, Z.; Li, R.; Tian, X.; Wang, Y.; Lin, Y.; Wu, D.; et al. Resveratrol protects boar sperm in vitro via its antioxidant capacity. Zygote 2020, 28, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Fan, X.; Lv, Y.; Zhang, N.; Fan, C.; Zhang, P.; Zeng, W. Vitamin E Analogue Improves Rabbit Sperm Quality during the Process of Cryopreservation through Its Antioxidative Action. PLoS ONE 2015, 10, e0145383. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, T.; Yang, J.; Huang, Q.; Wu, H.; Pan, Y.; Wang, H.; Qian, Y. Epimedium protects against dyszoospermia in mice with Pex3 knockout by exerting antioxidant effects and regulating the expression level of P16. Cell Death Dis. 2022, 13, 69. [Google Scholar] [CrossRef]
- Bucak, M.N.; Coyan, K.; Oztürk, C.; Güngör, S.; Omür, A.D. Methionine supplementation improves ram sperm parameters during liquid storage at 5 °C. Cryobiology 2012, 65, 335–337. [Google Scholar] [CrossRef]
- Jonscher, K.R.; Chowanadisai, W.; Rucker, R.B. Pyrroloquinoline-Quinone Is More Than an Antioxidant: A Vitamin-like Accessory Factor Important in Health and Disease Prevention. Biomolecules 2021, 11, 1441. [Google Scholar] [CrossRef]
- Pierpoint, W.S. PQQ in plants. Trends Biochem. Sci. 1990, 15, 299. [Google Scholar] [CrossRef]
- Paz, M.A.; Flückiger, R.; Torrelio, B.M.; Gallop, P.M. Methoxatin (PQQ), coenzyme for copper-dependent amine and mixed-function oxidation in mammalian tissues. Connect. Tissue Res. 1989, 20, 251–257. [Google Scholar] [CrossRef]
- Salmon, V.M.; Castonguay, F.; Demers-Caron, V.; Leclerc, P.; Bailey, J.L. Cholesterol-loaded cyclodextrin improves ram sperm cryoresistance in skim milk-extender. Anim. Reprod. Sci. 2017, 177, 1–11. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, M.; Lin, Z. Pyrroloquinoline quinone (PQQ) alleviated sepsis-induced acute liver injury, inflammation, oxidative stress and cell apoptosis by downregulating CUL3 expression. Bioengineered 2021, 12, 2459–2468. [Google Scholar] [CrossRef]
- Harris, C.B.; Chowanadisai, W.; Mishchuk, D.O.; Satre, M.A.; Slupsky, C.M.; Rucker, R.B. Dietary pyrroloquinoline quinone (PQQ) alters indicators of inflammation and mitochondrial-related metabolism in human subjects. J. Nutr. Biochem. 2013, 24, 2076–2084. [Google Scholar] [CrossRef]
- Boretti, A. PQQ Supplementation and SARS-CoV-2 Spike Protein-Induced Heart Inflammation. Nat. Prod. Commun. 2022, 17, 1934578x221080929. [Google Scholar] [CrossRef]
- Kumazawa, T.; Hiwasa, T.; Takiguchi, M.; Suzuki, O.; Sato, K. Activation of Ras signaling pathways by pyrroloquinoline quinone in NIH3T3 mouse fibroblasts. Int. J. Mol. Med. 2007, 19, 765–770. [Google Scholar] [CrossRef]
- Tchaparian, E.; Marshal, L.; Cutler, G.; Bauerly, K.; Chowanadisai, W.; Satre, M.; Harris, C.; Rucker, R.B. Identification of transcriptional networks responding to pyrroloquinoline quinone dietary supplementation and their influence on thioredoxin expression, and the JAK/STAT and MAPK pathways. Biochem. J. 2010, 429, 515–526. [Google Scholar] [CrossRef]
- Hara, H.; Hiramatsu, H.; Adachi, T. Pyrroloquinoline quinone is a potent neuroprotective nutrient against 6-hydroxydopamine-induced neurotoxicity. Neurochem. Res. 2007, 32, 489–495. [Google Scholar] [CrossRef]
- Murase, K.; Hattori, A.; Kohno, M.; Hayashi, K. Stimulation of nerve growth factor synthesis/secretion in mouse astroglial cells by coenzymes. Biochem. Mol. Biol. Int. 1993, 30, 615–621. [Google Scholar]
- Li, H.H.; Liu, S.Q.; Peng, H.; Zhang, N. Pyrroloquinoline quinone enhances regeneration of transected sciatic nerve in rats. Chin. J. Traumatol. 2005, 8, 225–229. [Google Scholar]
- Dai, X.; Yi, X.; Wang, Y.; Xia, W.; Tao, J.; Wu, J.; Miao, D.; Chen, L. PQQ Dietary Supplementation Prevents Alkylating Agent-Induced Ovarian Dysfunction in Mice. Front. Endocrinol. 2022, 13, 781404. [Google Scholar] [CrossRef]
- Hoque, S.A.M.; Umehara, T.; Kawai, T.; Shimada, M. Adverse effect of superoxide-induced mitochondrial damage in granulosa cells on follicular development in mouse ovaries. Free Radic. Biol. Med. 2021, 163, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Wang, Z.; Guo, Y.; Sheng, X.; Xing, K.; Ni, H.; Wang, X.; Xiao, L.; Qi, X. Research Note: Dietary supplementation with pyrroloquinoline quinone disodium (PQQ.Na(2)) improves oxidative status and semen quality in aging layer breeder roosters. Poult. Sci. 2022, 101, 101812. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Kawai, T.; Umehara, T.; Hoque, S.A.M.; Zeng, W.; Shimada, M. Negative effects of ROS generated during linear sperm motility on gene expression and ATP generation in boar sperm mitochondria. Free Radic. Biol. Med. 2019, 141, 159–171. [Google Scholar] [CrossRef] [PubMed]
- van der Horst, G. Computer Aided Sperm Analysis (CASA) in domestic animals: Current status, three D tracking and flagellar analysis. Anim. Reprod. Sci. 2020, 220, 106350. [Google Scholar] [CrossRef]
- Galarza, D.A.; López-Sebastián, A.; Santiago-Moreno, J. Supplementing a skimmed milk-egg yolk-based extender with L-carnitine helps maintain the motility, membrane integrity and fertilizing capacity of chilled ram sperm. Reprod. Domest. Anim. 2020, 55, 805–813. [Google Scholar] [CrossRef]
- Mehmood, A.; Anwar, M.; Naqvi, S.M. Motility, acrosome integrity, membrane integrity and oocyte cleavage rate of sperm separated by swim-up or Percoll gradient method from frozen-thawed buffalo semen. Anim. Reprod. Sci. 2009, 111, 141–148. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, W.; Li, R.; Zeng, W. Reducing the Glucose Level in Pre-treatment Solution Improves Post-thaw Boar Sperm Quality. Front. Vet. Sci. 2022, 9, 856536. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, Q.; Wang, S.; Ma, R.; Jing, J.; Yang, Y.; Feng, Y.; Zou, Z.; Zhang, Y.; Ge, X.; et al. Mitochondria-related miR-574 reduces sperm ATP by targeting ND5 in aging males. Aging 2020, 12, 8321–8338. [Google Scholar] [CrossRef]
- Najafi, A.; Daghigh Kia, H.; Mehdipour, M.; Shamsollahi, M.; Miller, D.J. Does fennel extract ameliorate oxidative stress frozen-thawed ram sperm? Cryobiology 2019, 87, 47–51. [Google Scholar] [CrossRef]
- Zhang, W.; Min, L.; Li, Y.; Lang, Y.; Hoque, S.A.M.; Adetunji, A.O.; Zhu, Z. Beneficial Effect of Proline Supplementation on Goat Spermatozoa Quality during Cryopreservation. Animals 2022, 12, 2626. [Google Scholar] [CrossRef]
- Miguel-Jiménez, S.; Pina-Beltrán, B.; Gimeno-Martos, S.; Carvajal-Serna, M.; Casao, A.; Pérez-Pe, R. NADPH Oxidase 5 and Melatonin: Involvement in Ram Sperm Capacitation. Front. Cell Dev. Biol. 2021, 9, 655794. [Google Scholar] [CrossRef]
- Bromfield, E.G.; Aitken, R.J.; Gibb, Z.; Lambourne, S.R.; Nixon, B. Capacitation in the presence of methyl-β-cyclodextrin results in enhanced zona pellucida-binding ability of stallion spermatozoa. Reproduction 2014, 147, 153–166. [Google Scholar] [CrossRef]
- Clulow, J.R.; Evans, G.; Maxwell, W.M.; Morris, L.H. Evaluation of the function of fresh and frozen-thawed sex-sorted and non-sorted stallion spermatozoa using a heterologous oocyte binding assay. Reprod. Fertil. Dev. 2010, 22, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Mugnier, S.; Dell’Aquila, M.E.; Pelaez, J.; Douet, C.; Ambruosi, B.; De Santis, T.; Lacalandra, G.M.; Lebos, C.; Sizaret, P.Y.; Delaleu, B.; et al. New insights into the mechanisms of fertilization: Comparison of the fertilization steps, composition, and structure of the zona pellucida between horses and pigs. Biol. Reprod. 2009, 81, 856–870. [Google Scholar] [CrossRef]
- Tanihara, F.; Nakai, M.; Men, N.T.; Kato, N.; Kaneko, H.; Noguchi, J.; Otoi, T.; Kikuchi, K. Roles of the zona pellucida and functional exposure of the sperm-egg fusion factor ‘IZUMO’ during in vitro fertilization in pigs. Anim. Sci. J. 2014, 85, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Taberner, E.; Morató, R.; Mogas, T.; Miró, J. Ability of Catalonian donkey sperm to penetrate zona pellucida-free bovine oocytes matured in vitro. Anim. Reprod. Sci. 2010, 118, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.J.; Fisher, P.R. Mitochondria in Health and Disease. Cells 2019, 8, 680. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Jiang, C.; Qin, J.; Ke, K.; Ding, F. Involvement of ERK1/2 pathway in neuroprotective effects of pyrroloquinoline quinine against rotenone-induced SH-SY5Y cell injury. Neuroscience 2014, 270, 183–191. [Google Scholar] [CrossRef]
- Losano, J.D.A.; Angrimani, D.S.R.; Dalmazzo, A.; Rocha, C.C.; Brito, M.M.; Perez, E.G.A.; Tsunoda, R.H.; Góes, P.A.A.; Mendes, C.M.; Assumpção, M.; et al. Effect of Vitamin E and Polyunsaturated Fatty Acids on Cryopreserved Sperm Quality in Bos taurus Bulls Under Testicular Heat Stress. Anim. Biotechnol. 2018, 29, 100–109. [Google Scholar] [CrossRef]
- Ramalho-Santos, J.; Amaral, S. Mitochondria and mammalian reproduction. Mol. Cell. Endocrinol. 2013, 379, 74–84. [Google Scholar] [CrossRef]
- Miller, W.L. Steroid hormone synthesis in mitochondria. Mol. Cell. Endocrinol. 2013, 379, 62–73. [Google Scholar] [CrossRef]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Guha, M.; Kashina, A.; Avadhani, N.G. Mitochondrial dysfunction and mitochondrial dynamics-The cancer connection. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 602–614. [Google Scholar] [CrossRef]
- Nowicka-Bauer, K.; Lepczynski, A.; Ozgo, M.; Kamieniczna, M.; Fraczek, M.; Stanski, L.; Olszewska, M.; Malcher, A.; Skrzypczak, W.; Kurpisz, M.K. Sperm mitochondrial dysfunction and oxidative stress as possible reasons for isolated asthenozoospermia. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2018, 69, 403–417. [Google Scholar] [CrossRef]
- Mundy, A.J.; Ryder, T.A.; Edmonds, D.K. Asthenozoospermia and the human sperm mid-piece. Hum. Reprod. 1995, 10, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Pelliccione, F.; Micillo, A.; Cordeschi, G.; D’Angeli, A.; Necozione, S.; Gandini, L.; Lenzi, A.; Francavilla, F.; Francavilla, S. Altered ultrastructure of mitochondrial membranes is strongly associated with unexplained asthenozoospermia. Fertil. Steril. 2011, 95, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Vercellino, I.; Sazanov, L.A. The assembly, regulation and function of the mitochondrial respiratory chain. Nat. Rev. Mol. Cell Biol. 2022, 23, 141–161. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, M.; Pyakurel, A.; Glytsou, C.; Scorrano, L. The cell biology of mitochondrial membrane dynamics. Nat. Rev. Mol. Cell Biol. 2020, 21, 204–224. [Google Scholar] [CrossRef]
- St John, J.C.; Bowles, E.J.; Amaral, A. Sperm mitochondria and fertilisation. Soc. Reprod. Fertil. Suppl. 2007, 65, 399–416. [Google Scholar]
- Zhao, C.; Guo, X.J.; Shi, Z.H.; Wang, F.Q.; Huang, X.Y.; Huo, R.; Zhu, H.; Wang, X.R.; Liu, J.Y.; Zhou, Z.M.; et al. Role of translation by mitochondrial-type ribosomes during sperm capacitation: An analysis based on a proteomic approach. Proteomics 2009, 9, 1385–1399. [Google Scholar] [CrossRef]
- de Mateo, S.; Martínez-Heredia, J.; Estanyol, J.M.; Domínguez-Fandos, D.; Vidal-Taboada, J.M.; Ballescà, J.L.; Oliva, R. Marked correlations in protein expression identified by proteomic analysis of human spermatozoa. Proteomics 2007, 7, 4264–4277. [Google Scholar] [CrossRef]
- Varuzhanyan, G.; Chan, D.C. Mitochondrial dynamics during spermatogenesis. J. Cell Sci. 2020, 133, jcs235937. [Google Scholar] [CrossRef] [PubMed]
- Shivaji, S.; Kota, V.; Siva, A.B. The role of mitochondrial proteins in sperm capacitation. J. Reprod. Immunol. 2009, 83, 14–18. [Google Scholar] [CrossRef]
- Parte, P.P.; Rao, P.; Redij, S.; Lobo, V.; D’Souza, S.J.; Gajbhiye, R.; Kulkarni, V. Sperm phosphoproteome profiling by ultra performance liquid chromatography followed by data independent analysis (LC-MS(E)) reveals altered proteomic signatures in asthenozoospermia. J. Proteom. 2012, 75, 5861–5871. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Pesini, E.; Diez, C.; Lapeña, A.C.; Pérez-Martos, A.; Montoya, J.; Alvarez, E.; Arenas, J.; López-Pérez, M.J. Correlation of sperm motility with mitochondrial enzymatic activities. Clin. Chem. 1998, 44, 1616–1620. [Google Scholar] [CrossRef]
- Park, Y.J.; Pang, M.G. Mitochondrial Functionality in Male Fertility: From Spermatogenesis to Fertilization. Antioxidants 2021, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Barati, E.; Nikzad, H.; Karimian, M. Oxidative stress and male infertility: Current knowledge of pathophysiology and role of antioxidant therapy in disease management. Cell. Mol. Life Sci. CMLS 2020, 77, 93–113. [Google Scholar] [CrossRef]
- Khosravizadeh, Z.; Khodamoradi, K.; Rashidi, Z.; Jahromi, M.; Shiri, E.; Salehi, E.; Talebi, A. Sperm cryopreservation and DNA methylation: Possible implications for ART success and the health of offspring. J. Assist. Reprod. Genet. 2022, 39, 1815–1824. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Scarlata, E. Oxidative stress and reproductive function: The protection of mammalian spermatozoa against oxidative stress. Reproduction 2022, 164, F67–F78. [Google Scholar] [CrossRef]
- Li, R.; Wu, X.; Zhu, Z.; Lv, Y.; Zheng, Y.; Lu, H.; Zhou, K.; Wu, D.; Zeng, W.; Dong, W.; et al. Polyamines protect boar sperm from oxidative stress in vitro. J. Anim. Sci. 2022, 100, skac069. [Google Scholar] [CrossRef]
- Purdy, P.H.; Graham, J.K.; Azevedo, H.C. Evaluation of boar and bull sperm capacitation and the acrosome reaction using flow cytometry. Anim. Reprod. Sci. 2022, 246, 106846. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters | Time (h) | 0 nM | 10 nM | 100 nM | 1000 nM | 10,000 nM |
|---|---|---|---|---|---|---|
| Total motility (%) | 24 | 87.9 ±11.4 b | 90.0 ± 3.9 ab | 93.3 ± 1.2 a | 88.5 ± 3.0 b | 77.8 ± 4.3 c |
| 48 | 74.9 ± 5.0 bc | 80.2 ± 6.1 b | 86.0 ± 7.4 a | 82.0 ± 6.5 b | 70.8 ± 3.4 c | |
| 72 | 59.2 ± 3.1 c | 67.3 ± 5.4 b | 72.1 ± 2.1 a | 71.4 ± 0.5 a | 58.3 ± 0.5 c | |
| 96 | 40.3 ± 0.5 b | 41.8 ± 1.6 ab | 43.6 ± 0.5 a | 38.8 ± 2.2 b | 39.2 ± 2.0 b | |
| Progressive motility (%) | 24 | 60.5 ± 7.8 bc | 71.4± 4.1 b | 73.2 ± 4.1 a | 68.1 ± 1.6 b | 51.9 ± 1.8 c |
| 48 | 53.5 ± 6.1 bc | 53.4 ± 7.2 bc | 71.3 ± 8.4 a | 57.2 ± 7.6 b | 41.4 ± 2.9 c | |
| 72 | 41.2 ± 5.2 b | 43.6 ± 1.3 ab | 48.5 ± 0.6 a | 45.1 ± 0.2 a | 39.6 ± 1.6 b | |
| 96 | 22.3 ± 0.7 b | 22.6 ± 1.2 b | 24.5 ± 1.4 a | 24.9 ± 0.7 a | 20.6 ± 0.6 c | |
| VCL (μm/s) | 24 | 67.8 ± 3.5 b | 60.8 ± 8.7 b | 76.6± 6.0 a | 66.2± 3.0 b | 54.2 ± 1.2 c |
| 48 | 67.6 ± 4.3 b | 70.2 ± 4.3 b | 78.9 ± 10 a | 74.9± 11.2 a | 63.6 ± 1.2 c | |
| 72 | 59.3 ± 3.6 b | 62.6 ± 0.3 b | 67.5 ± 8.2 a | 66.2 ± 4.9 a | 52.3 ± 0.5 c | |
| 96 | 36.5 ± 1.2 b | 36.3 ± 0.5 b | 42.7 ± 0.4 a | 38.5 ± 1.5 b | 32.5 ± 8.0 c | |
| VSL (μm/s) | 24 | 38.5 ± 7.8 b | 47.2 ± 5.0 ab | 58.0 ± 3.1 a | 41.4 ± 4.2 b | 30.3 ± 2.2 c |
| 48 | 36.7 ± 5.0 b | 35.8 ± 4.9 b | 45.5 ± 6.5 a | 36.4 ± 7.5 b | 29.6 ± 0.6 c | |
| 72 | 36.5 ± 4.2 b | 34.8 ± 4.1 b | 44.9 ± 5.6 a | 38.3 ± 7.8 b | 26.3 ± 2.9 c | |
| 96 | 18.5 ± 0.6 b | 20.3 ± 2.3 b | 22.6 ± 4.2 a | 29.2 ± 2.3 a | 15.6 ± 0.9 c | |
| VAP (μm/s) | 24 | 47.6 ± 4.8 b | 47.2 ± 5.0 b | 61.6 ± 2.1 a | 38.6 ± 3.9 c | 33.4 ± 4.1 d |
| 48 | 48.8 ± 11.0 b | 46.5 ± 6.5 b | 59.0 ± 9.2 a | 47.7 ± 8.9 b | 36.3 ± 0.7 c | |
| 72 | 45.2 ± 12 b | 47.8 ± 5.6 b | 55.4 ± 11.4 a | 41.6 ± 8.6 c | 31.2 ± 0.9 d | |
| 96 | 39.6 ± 0.5 b | 41.3 ± 5.2 b | 46.2 ± 4.2 a | 45.3 ± 0.7 a | 32.9 ± 5.8 c | |
| BCF (Hz) | 24 | 24.7 ± 1.2 | 24.5 ± 1.4 | 26.4 ± 0.9 | 24.9 ± 0.6 | 24.2 ± 1.2 |
| 48 | 21.3 ± 3.2 | 22.1 ± 4.1 | 22.5 ± 3.2 | 21.9 ± 0.9 | 20.6 ± 1.3 | |
| 72 | 22.3 ± 3.1 | 31.5 ± 1.6 | 23.6 ± 3.6 | 19.6 ± 0.8 | 22.3 ± 0.5 | |
| 96 | 22.6 ± 2.4 | 22.9 ± 1.2 | 24.5 ± 3.2 | 21.3 ± 1.4 | 26.1 ± 0.8 | |
| ALH (μm) | 24 | 8.7 ± 0.6 | 8.3 ± 0.3 | 8.7 ± 0.4 | 8.2 ± 0.3 | 8.4 ± 0.3 |
| 48 | 6.9 ± 0.5 | 7.2 ± 1.2 | 7.3 ± 1.3 | 6.9 ± 2.3 | 7.3 ± 1.2 | |
| 72 | 7.5 ± 0.9 | 7.4 ± 1.2 | 7.3 ± 1.6 | 7.8 ± 1.9 | 7.4 ± 1.6 | |
| 96 | 7.4. ± 1.4 | 8.3 ± 0.9 | 8.5 ± 2.1 | 7.6 ± 0.6 | 7.2 ± 1.5 | |
| STR (%) | 24 | 79.7 ± 1.7 b | 76.4 ± 1.3 b | 81.4 ± 1.4 a | 79.7 ± 3.1 b | 75.6 ± 0.9 c |
| 48 | 66.8 ± 4.3 b | 67.5 ± 2.3 b | 72.1 ± 1.7 a | 55.6 ± 1.7 c | 51.5 ± 0.8 d | |
| 72 | 71.3 ± 2.6 b | 74.9 ± 2.5 ab | 75.3 ± 0.9 a | 55.6 ± 0.9 c | 71.3 ± 1.4 b | |
| 96 | 54.1 ± 1.2 c | 58.6 ± 2.1 bc | 71.3 ± 0.8 a | 64.2 ± 1.5 b | 63.8 ± 1.8 b | |
| LIN (%) | 24 | 49.2 ± 1.5 b | 42.2 ± 6.3 b | 60.1 ± 1.5 a | 46.5 ± 4.4 b | 41.2 ± 0.7 b |
| 48 | 47.4 ± 0.3 ab | 47.4 ± 6.3 ab | 58.3 ± 5.2 a | 47.8 ± 5.3 a | 46.5 ± 0.8 b | |
| 72 | 46.9 ± 7.2 b | 47.3 ± 6.2 b | 55.3 ± 4.6 a | 47.8 ± 12.0 b | 46.2 ± 11.4 b | |
| 96 | 32.1 ± 0.5 c | 34.2 ± 0.8 c | 44.5 ± 1.6 a | 41.2 ± 0.6 b | 43.2 ± 1.2 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Li, W.; Yang, Q.; Zhao, H.; Zhang, W.; Adetunji, A.O.; Hoque, S.A.M.; Kou, X.; Min, L. Pyrroloquinoline Quinone Improves Ram Sperm Quality through Its Antioxidative Ability during Storage at 4 °C. Antioxidants 2024, 13, 104. https://doi.org/10.3390/antiox13010104
Zhu Z, Li W, Yang Q, Zhao H, Zhang W, Adetunji AO, Hoque SAM, Kou X, Min L. Pyrroloquinoline Quinone Improves Ram Sperm Quality through Its Antioxidative Ability during Storage at 4 °C. Antioxidants. 2024; 13(1):104. https://doi.org/10.3390/antiox13010104
Chicago/Turabian StyleZhu, Zhendong, Wenjia Li, Qitai Yang, Haolong Zhao, Weijing Zhang, Adedeji O. Adetunji, S. A. Masudul Hoque, Xin Kou, and Lingjiang Min. 2024. "Pyrroloquinoline Quinone Improves Ram Sperm Quality through Its Antioxidative Ability during Storage at 4 °C" Antioxidants 13, no. 1: 104. https://doi.org/10.3390/antiox13010104
APA StyleZhu, Z., Li, W., Yang, Q., Zhao, H., Zhang, W., Adetunji, A. O., Hoque, S. A. M., Kou, X., & Min, L. (2024). Pyrroloquinoline Quinone Improves Ram Sperm Quality through Its Antioxidative Ability during Storage at 4 °C. Antioxidants, 13(1), 104. https://doi.org/10.3390/antiox13010104