

Liver Antioxidant Capacity and Steatosis in Laying Hens Exposed to Various Quantities of Lupin (Lupinus angustifolius) Seeds in the Diet

 , , ,
, , ,  , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Diets
2.3. Blood and Tissue Samples
2.4. Histopathological Examination of Liver Tissue
2.5. Determination of Malondialdehyde (MDA) Concentrations in Tissue
2.6. Isolation of Microsomal and Cytosolic Fractions
2.7. Evaluation of DT-Diaphorase Activity
2.8. Assessment of Ethoxycoumarin O-Deethylase Activity
2.9. Evaluation of Catalase Activity
2.10. Assay of Catalase and Superoxide Dismutase in Chicken Serum
2.11. Statistical Analysis
3. Results
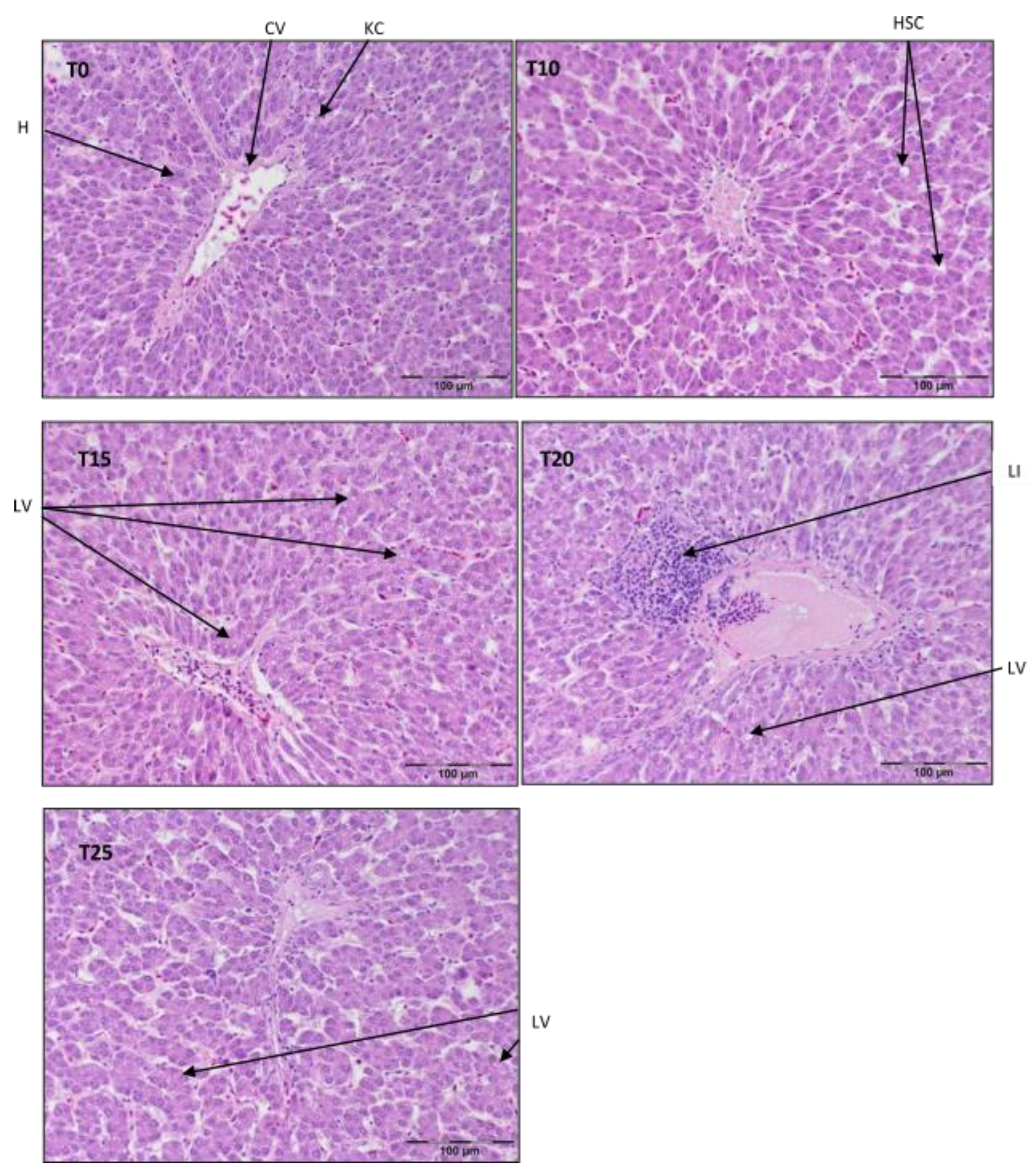
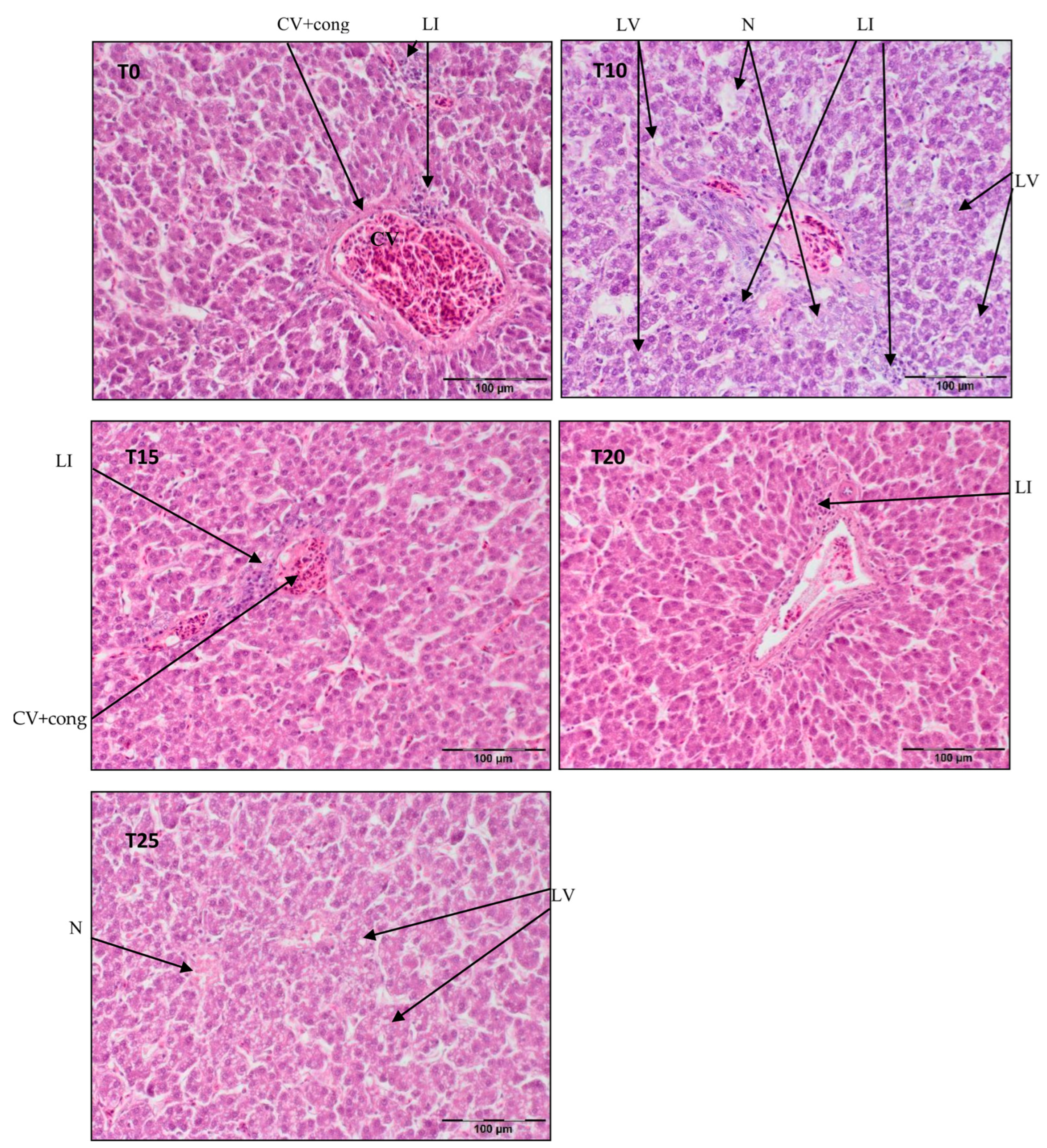
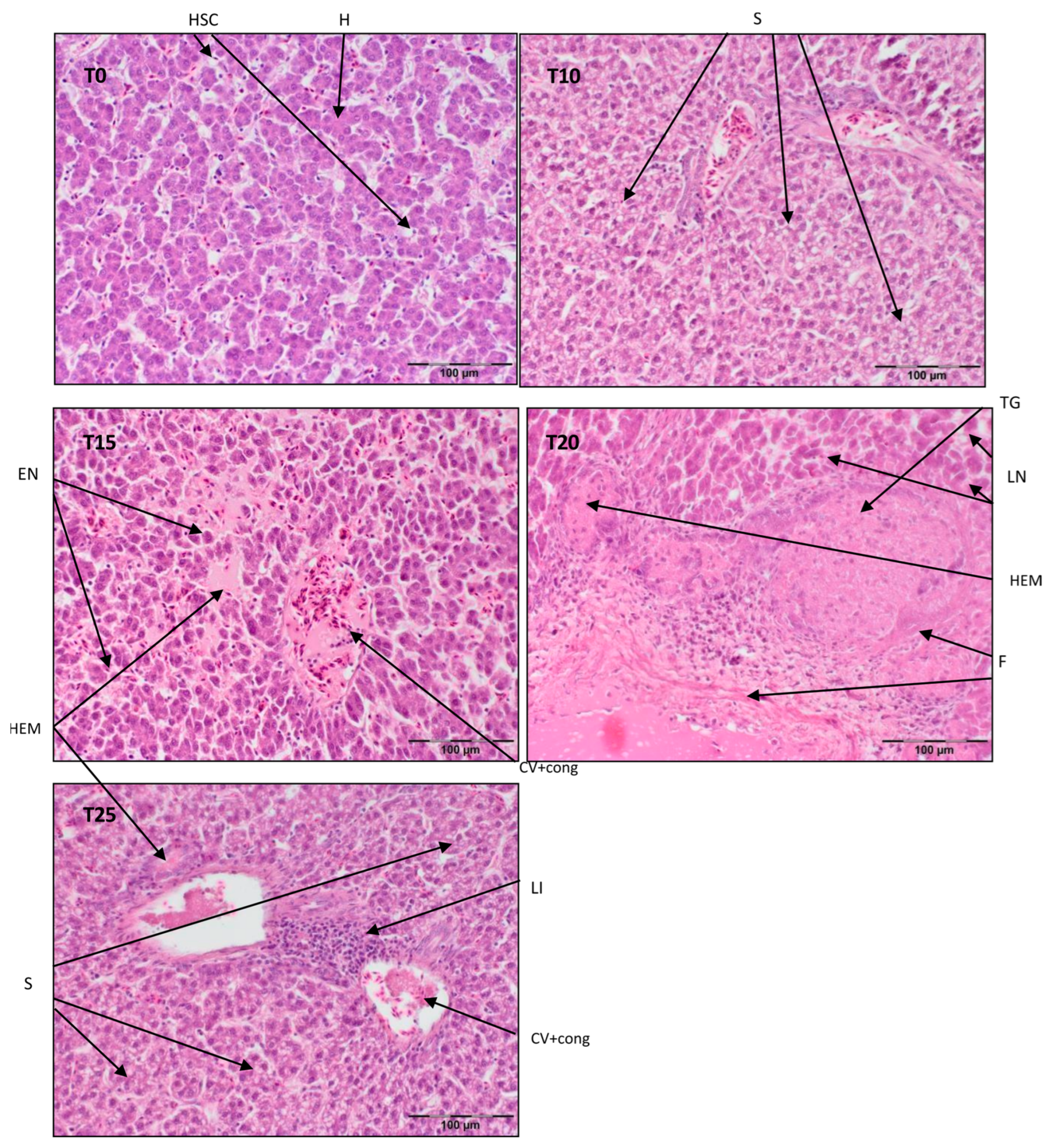
3.1. Liver Histopathology
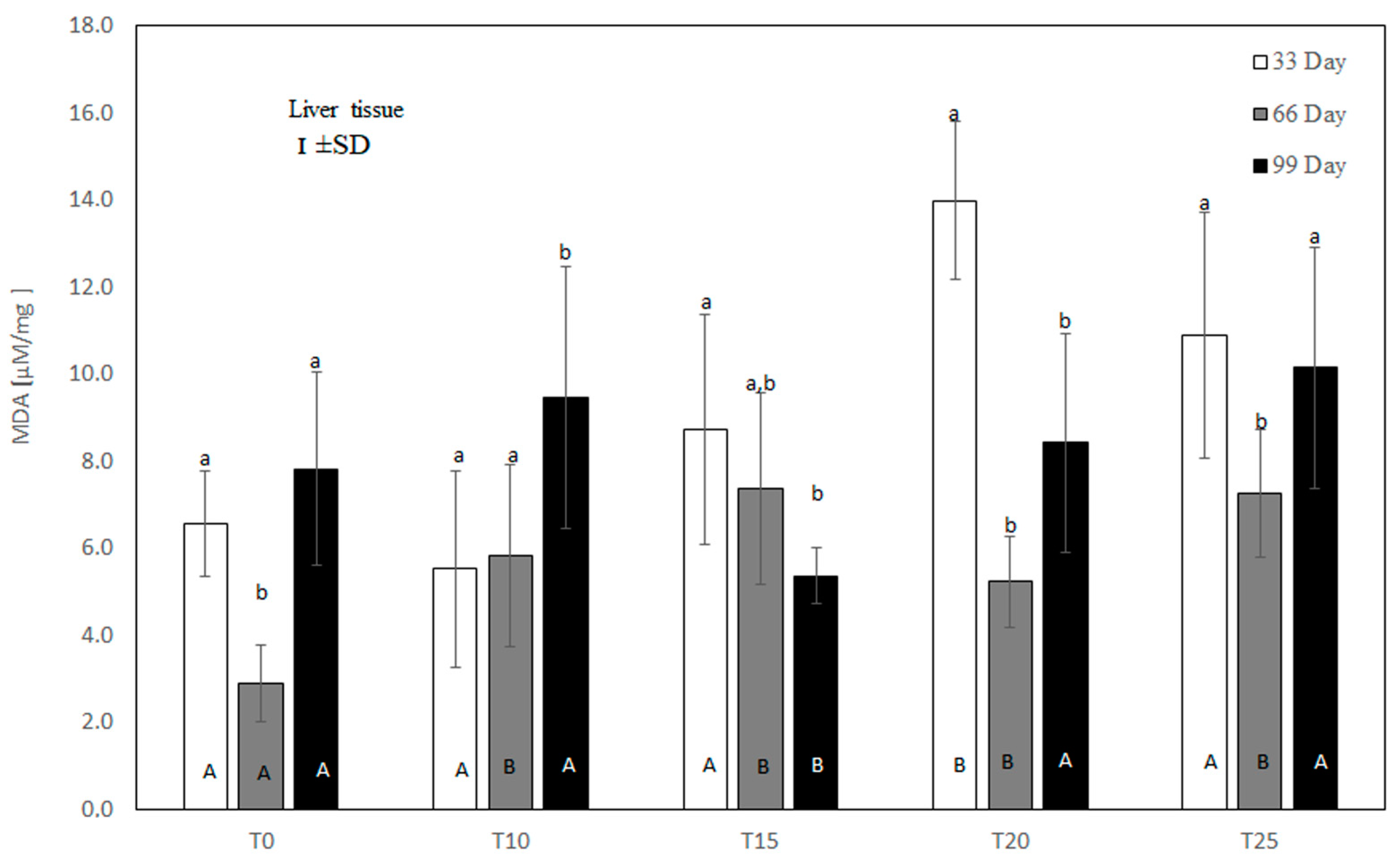
3.2. Malondialdehyde (MDA) Concentration in the Liver of Hens
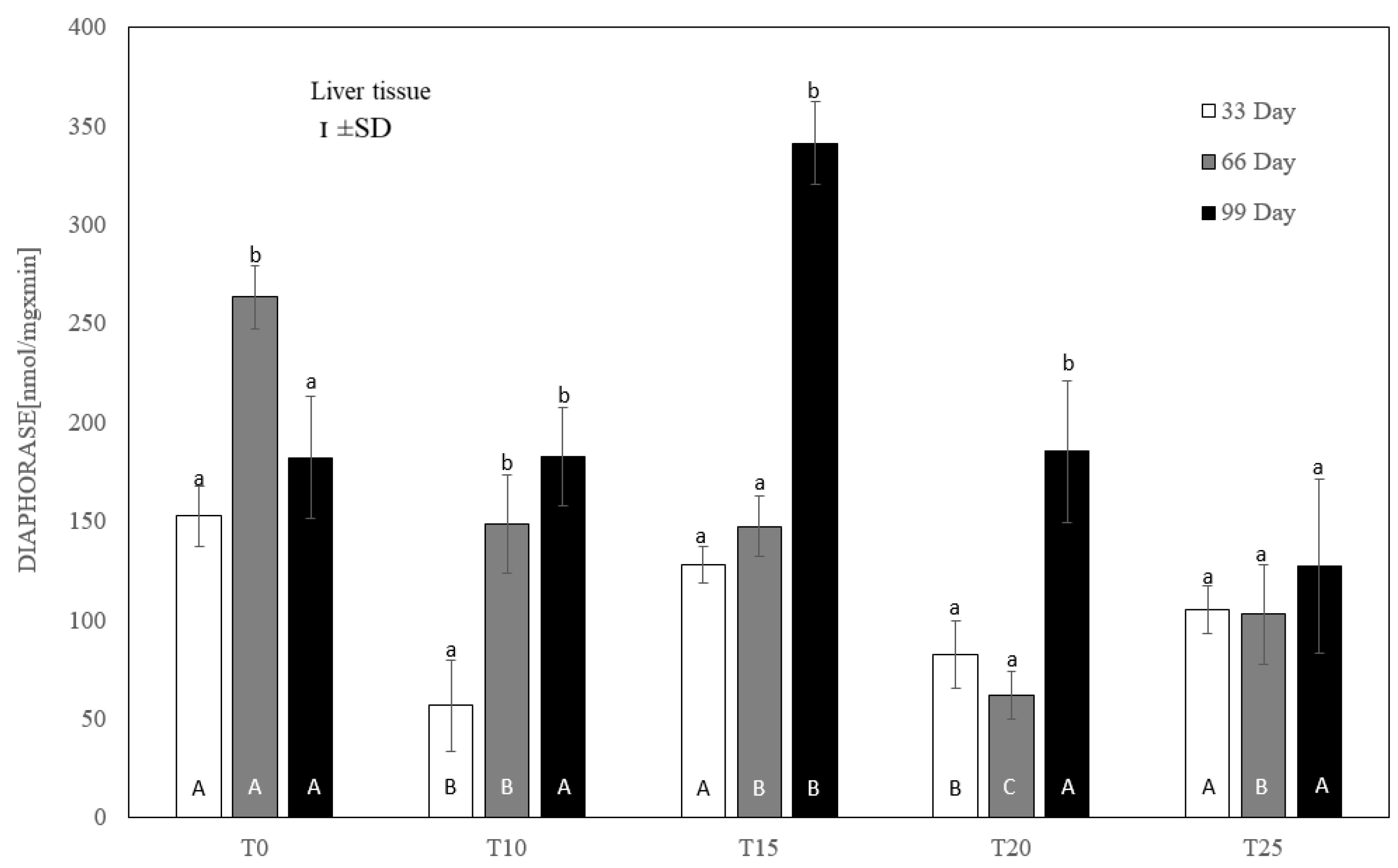
3.3. Cytosolic and Microsomal Antioxidant Capacity—Mean Activity of DT-Diaphorase, Catalase (CAT), and Ethoxycoumarin O-Deethylase in the Liver of Hens
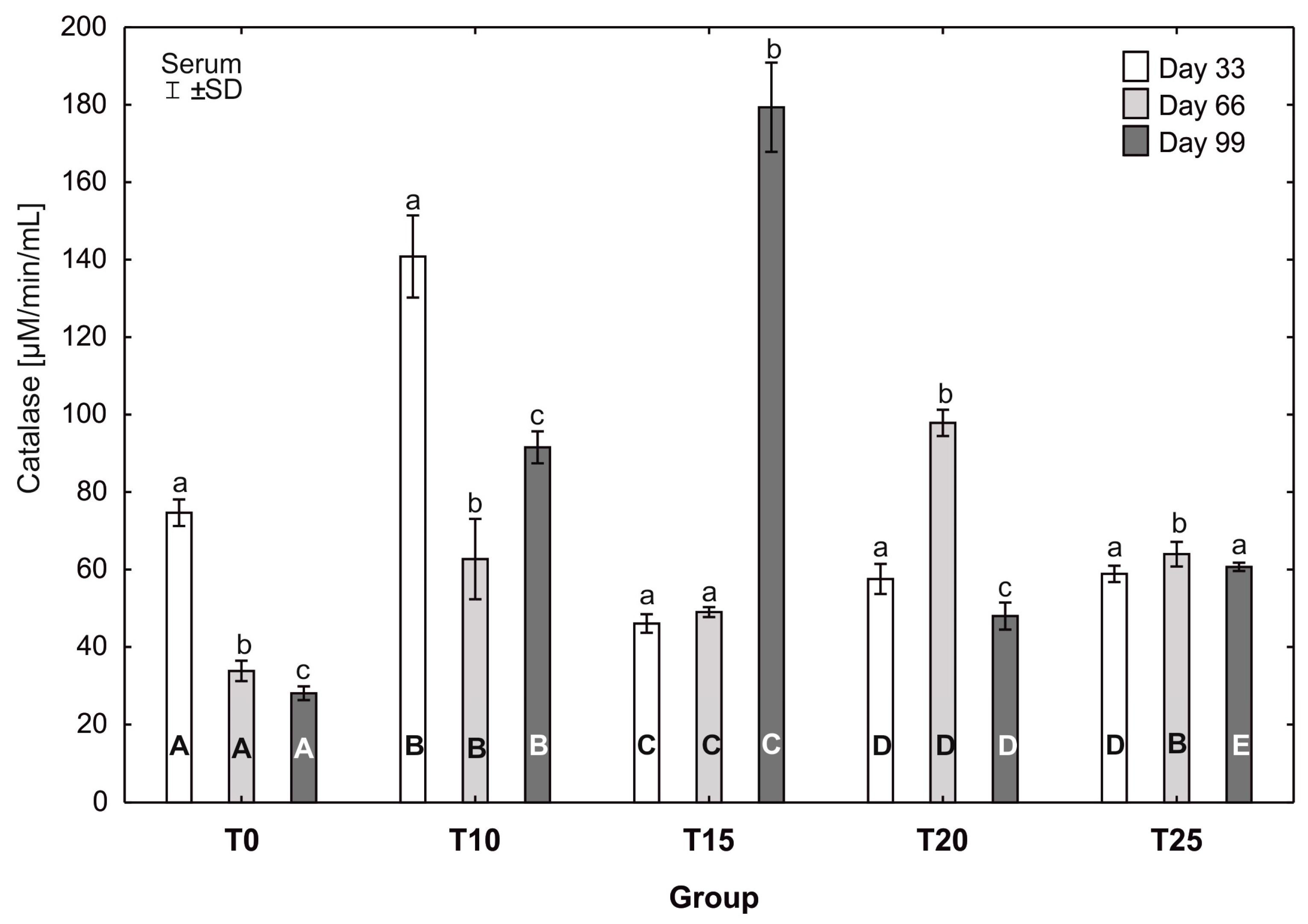
3.4. Mean Activity of Catalase (CAT) and Superoxide Dismutase (SOD) in the Serum of Hens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Yue, H.; Wu, S.; Zhang, H.; Qi, G. Nutritional modulation of health, egg quality and environmental pollution of the layers. Anim. Nutr. 2017, 3, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, E.; Kucharska-Gaca, J.; Kuźniacka, J.; Lewko, L.; Gornowicz, E.; Biesek, J.; Adamski, M. Egg quality depending on the diet with different sources of protein and age of the hens. Sci. Rep. 2021, 11, 2638. [Google Scholar] [CrossRef]
- Bryden, W.L.; Li, X.; Ruhnke, I.; Zhang, D.; Shini, S. Nutrition, feeding and laying hen welfare. Anim. Prod. Sci. 2021, 61, 893–914. [Google Scholar] [CrossRef]
- He, W.; Li, P.; Wu, G. Amino acid nutrition and metabolism in chickens. In Amino Acids in Nutrition and Health; Wu, G., Ed.; Advances in experimental medicine and biology; Springer: Cham, Switzerland, 2021; Volume 1285. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Tiwari, R.; Yatoo, M.I.; Karthik, K.; Michalak, I.; Dhama, K. Nutritional significance of amino acids, vitamins and minerals as nutraceuticals in poultry production and health—A comprehensive review. Vet. Q. 2020, 41, 1–29. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Wang, H.; Wang, J.; Wu, S.; Qi, G. Effect of dietary protein sources on production performance, egg quality, and plasma parameters of laying hens. Asian-Australas. J. Anim. Sci. 2017, 30, 400–409. [Google Scholar] [CrossRef][Green Version]
- Różewicz, M.; Grabiński, J.; Sułek, A. Possibilities and limitations in the use of legumes from domestic cultivation in poultry feed in the context of fodder protein deficit. Pol. J. Agron. 2018, 35, 32–44. [Google Scholar] [CrossRef]
- EFSA GMO Panel Working Group on Animal Feeding Trials. Safety and nutritional assessment of GM plants and derived food and feed: The role of animal feeding trials. Food Chem. Toxicol. 2008, 46, 2–70. [Google Scholar] [CrossRef]
- Tufarelli, V.; Selvaggi, M.; Dario, C.; Laudadio, V. Genetically Modified Feeds in Poultry Diet: Safety, Performance, and Product Quality. Crit. Rev. Food Sci. Nutr. 2015, 55, 562–569. [Google Scholar] [CrossRef]
- Kuźniacka, J.; Biesek, J.; Banaszak, M.; Grabowicz, M.; Adamski, M. The Quality of Eggs from Rosa 1 Hens Fed Diets Containing Seeds of Legume Plants (Lupinus luteus L., Lupinus angustifolius, and Pisum sativum) in Two Laying Phases. Animals 2020, 10, 1942. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, S.I.; Kim, I.H. Effects of lupin seed supplementation on egg production performance, and qualitative egg traits in laying hens. Vet. Med. 2016, 61, 701–709. [Google Scholar] [CrossRef]
- Lemus-Conejo, A.; Rivero-Pino, F.; Montserrat-de la Paz, S.; Millan-Linares, M.C. Nutritional composition and biological activity of narrow-leafed lupins (Lupinus angustifolius L.) hydrolysates and seeds. Food Chem. 2023, 420, 136104. [Google Scholar] [CrossRef]
- Lima-Cabello, E.; Morales-Santana, S.; León, J.; Alche, V.; Clemente, A.; Alche, J.D.; Jimenez-Lopez, J.C. Narrow-leafed lupin (Lupinus angustifolius L.) seed β-conglutins reverse the induced insulin resistance in pancreatic cells. Food Funct. 2018, 9, 5176–5188. [Google Scholar] [CrossRef]
- Millán-Linares, M.D.C.; Yust, M.D.M.; Alcaide-Hidalgo, J.M.; Millán, F.; Pedroche, J. Lupine protein hydrolysates inhibit enzymes involved in the inflammatory pathway. Food Chem. 2014, 151, 141–147. [Google Scholar] [CrossRef]
- Bettzieche, A.; Brandsch, C.; Schmidt, M.; Weisse, K.; Eder, K.; Stangl, G.I. Differing effect of protein isolates from different cultivars of blue lupine on plasma lipoproteins of hypercholesterolemic rats. Biosci. Biotechnol. Biochem. 2008, 72, 3114–3121. [Google Scholar] [CrossRef]
- Fontanari, G.G.; Batistuti, J.P.; da Cruz, R.J.; Saldiva, P.H.N.; Arêas, J.A.G. Cholesterol-lowering effect of whole lupine (Lupinus albus) seed and its protein isolate. Food Chem. 2012, 132, 1521–1526. [Google Scholar] [CrossRef]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; del Carmen Millán-Linares, M.; del Mar Yust, M.; Pedroche, J.; Millán, F.; Lardone, P.J.; Carrera-Sánchez, C.; Guerrero, J.M.; Carrillo-Vico, A. Lupine protein hydrolysates decrease the inflammatory response and improve the oxidative status in human peripheral lymphocytes. Food Res. Int. 2019, 126, 108585. [Google Scholar] [CrossRef]
- Belski, R. Fiber, protein, and lupin-enriched foods: Role for improving cardiovascular health. Adv. Food Nutr. Res. 2012, 66, 147–215. [Google Scholar] [CrossRef] [PubMed]
- Okagu, I.U.; Ndefo, J.C.; Aham, E.C.; Obeme-Nmom, J.I.; Agboinghale, P.E.; Aguchem, R.N.; Nechi, R.N.; Lammi, C. Lupin-derived bioactive peptides: Intestinal transport, bioavailability and health benefits. Nutrients 2021, 13, 3266. [Google Scholar] [CrossRef] [PubMed]
- Olkowski, A.A.; Olkowski, B.I.; Amarowicz, R.; Classen, H.L. Adverse effects of dietary lupine in broiler chickens. Poult. Sci. 2001, 80, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos Pineda, J.; Nolasco Rodríguez, G.; Monteon, J.A.; García López, P.M.; Ruiz Lopez, M.A.; García Estrada, J. Histological evaluation of brain damage caused by crude quinolizidine alkaloid extracts from lupines. Histol. Histopathol. 2005, 20, 1147–1153. [Google Scholar] [CrossRef]
- Stanek, M.; Rotkiewicz, T.; Sobotka, W.; Bogusz, J.; Otrocka-Domagała, I.; Rotkiewicz, A. The effect of alkaloids presents in blue lupine (Lupinus angustifolius) seeds on the growth rate, selected biochemical blood indicators and histopathological changes in the liver of rats. Acta Vet. 2015, 84, 55–62. [Google Scholar] [CrossRef]
- Lemus-Conejo, A.; Grao-Cruces, E.; Toscano, R.; Varela, L.M.; Claro, C.; Pedroche, J.; Millan, F.; Millan-Linares, M.C.; Montserrat-de la Paz, S. A lupine (Lupinus angustifolious L.) peptide prevents non-alcoholic fatty liver disease in high-fat-diet-induced obese mice. Food Funct. 2020, 11, 2943–2952. [Google Scholar] [CrossRef] [PubMed]
- Andor, B.; Danciu, C.; Alexa, E.; Zupko, I.; Hogea, E.; Cioca, A.; Coricovac, D.; Pinzaru, I.; Patrascu, J.M.; Mioc, M.; et al. Germinated and ungerminated seeds extract from two lupinus species: Biological compounds characterization and in vitro and in vivo evaluations. Evid. Based-Complement. Altern. Med. 2016, 2016, 7638542. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.K.; Karnpanit, W.; Nasar-Abbas, S.M.; Huma, Z.E.; Jayasena, V. Phytochemical composition and bioactivities of lupin: A review. Int. J. Food Sci. Technol. 2015, 50, 2004–2012. [Google Scholar] [CrossRef]
- Kouris-Blazos, A.; Belski, R. Health benefits of legumes and pulses with a focus on Australian sweet lupins. Asia Pac. J. Clin. Nutr. 2016, 25, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, M.; Bustos, M.; Mendez, D.; Fuentes, E.; Palomo, I.; Lutz, M. In vitro assay of Quinoa (Chenopodium quinoa Willd.) and lupin (Lupinus spp.) extracts on human platelet aggregation. Plant Foods Hum. Nutr. 2020, 75, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Castillo, R.F.; García Pérez, R.; González Díaz, A.; Liñán González, A. Therapeutic applications and effects of Lupinus angustifolius (blue lupin) and its components: A systematic review and meta-analysis. Foods 2023, 12, 2749. [Google Scholar] [CrossRef] [PubMed]
- Siger, A.; Czubinski, J.; Kachlicki, P.; Dwiecki, K.; Lampart-Szczapa, E.; Nogala-Kalucka, M. Antioxidant activity and phenolic content in three lupin species. J. Food Compos. Anal. 2012, 25, 190–197. [Google Scholar] [CrossRef]
- Kamran, F.; Phillips, M.; Harman, D.G.; Reddy, N. Antioxidant activities of lupin (Lupinus angustifolius) protein hydrolysates and their potential for nutraceutical and functional foods. Food Chem. Adv. 2023, 2, 100297. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef]
- Gutteridge, J.M. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin. Chem. 1995, 41, 1819–1828. [Google Scholar] [CrossRef]
- Montuschi, P.; Barnes, P.J.; Roberts, L.J., 2nd. Isoprostanes: Markers and mediators of oxidative stress. FASEB J. 2004, 18, 1791–1800. [Google Scholar] [CrossRef]
- Barteczko, J.; Rutkowski, A. Nutritional Recommendations and Nutritional Value of Feed: Poultry Feeding Standards; Kielanowski Institute of Animal Physiology and Nutrition, National Academy of Sciences: Jabłonna, Poland, 2005. [Google Scholar]
- Van Barneveld, R. Understanding the nutritional chemistry of lupin (Lupinus spp.) seed to improve livestock production efficiency. Nutr. Res. Rev. 1999, 12, 203–230. [Google Scholar] [CrossRef] [PubMed]
- Perez-Maldonado, R.A.; Mannion, P.F.; Farrell, D.J. Optimum inclusion of field peas, faba beans, chick peas and sweet lupins in poultry diets. I. Chemical composition and layer experiments. Br. Poult. Sci. 1999, 40, 667–673. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Hy-Line International. Hy-Line Variety Brown Commercial Management Guide 2002–2004; Hy-Line International: West Des Moines, IA, USA, 2002. [Google Scholar]
- Wójcik, M.; Bobowiec, R.; Lisiecka, U.; Śmiech, A. Expression of receptor interacting protein 1 and receptor interacting protein 3 oval cells in a rat model of hepatocarcinogenesis. Exp. Ther. Med. 2018, 15, 4448–4456. [Google Scholar] [CrossRef] [PubMed]
- Wójcik, M.; Bobowiec, R.; Silmanowicz, P. Changes of bile secretion and liver peroxydation under influence of glicocortycoids in sheep. Ann. Univ. Mariae Curie-Skłodowska DD Med. Vet. 2005, 60, 132–140. [Google Scholar]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.J. Protein determination—Method matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, P.G.; Longo, V.; Naldi, F.; Panattoni, G.; Ursino, F. Xenobiotic-metabolizing enzymes in human respiratory nasal mucosa. Biochem. Pharmacol. 1991, 41, 177–184. [Google Scholar] [CrossRef]
- Dhruve, P.; Nauman, M.; Kale, R.K.; Singh, R.P. A Novel hepatoprotective activity of alangium salviifolium in mouse model. Drug Chem. Toxicol. 2022, 45, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Vivarelli, F.; Canistro, D.; Franchi, P.; Sapone, A.; Vornoli, A.; Della Croce, C.; Longo, V.; Lucarini, M.; Paolini, M. Disruption of redox homeostasis and carcinogen metabolizing enzymes changes by administration of vitamin E to rats. Life Sci. 2016, 145, 166–173. [Google Scholar] [CrossRef]
- Wheeler, C.R.; Salzman, J.A.; Elsayed, N.M.; Omaye, S.T.; Korte, D.W., Jr. Automated assays for superoxide dismutase, catalase, glutathione peroxidase, and glutathione reductase activity. Anal. Biochem. 1990, 184, 193–199. [Google Scholar] [CrossRef]
- Endale, H.T.; Tesfaye, W.; Mengstie, T.A. ROS induced lipid peroxidation and their role in ferroptosis. Front. Cell Dev. Biol. 2023, 11, 1226044. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Santos-Sánchez, G.; Cruz-Chamorro, I.; Álvarez-Ríos, A.I.; Fernández-Santos, J.M.; Vázquez-Román, M.V.; Rodríguez-Ortiz, B.; Álvarez-Sánchez, N.; Álvarez-López, A.I.; Millán-Linares, M.D.C.; Millán, F.; et al. Lupinus angustifolius protein hydrolysates reduce abdominal adiposity and ameliorate metabolic associated fatty liver disease (MAFLD) in western diet fed-ApoE-/- mice. Antioxidants 2021, 10, 1222. [Google Scholar] [CrossRef]
- Aydın, S.; Kaya, T.; Erman, O.; Yılmaz, Ö. The Protective effects of lupine (Lupinus albus) fruit extract against to destructive effects of hyperglycemia in type I diabetic rats. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 1466–1475. [Google Scholar] [CrossRef] [PubMed]
- Całyniuk, B.; Grochowska-Niedworok, E.; Walkiewicz, K.W.; Kawecka, S.; Popiołek, E.; Fatyga, E. Malondialdehyde (MDA)—Product of lipid peroxidation as a marker of homeostasis disorders and aging. Ann. Acad. Med. Siles. 2016, 70, 224–228. [Google Scholar] [CrossRef]
- Peverill, W.; Powell, L.W.; Skoien, R. Evolving concepts in the pathogenesis of NASH: Beyond steatosis and inflammation. Int. J. Mol. Sci. 2014, 15, 8591–8638. [Google Scholar] [CrossRef]
- Gresta, F.; Avola, G.; Abbate, V.; Magazzù, G.; Chiofalo, B. Lupin seed for the crop-livestock food chains. Ital. J. Agron. 2010, 4, 333–340. [Google Scholar] [CrossRef]
- Musco, N.; Cutrignelli, M.I.; Calabrò, S.; Tudisco, R.; Infascelli, F.; Grazioli, R.; Lo Presti, V.; Gresta, F.; Chiofalo, B. Comparison of nutritional and antinutritional traits among different species (Lupinus albus L., Lupinus luteus L., Lupinus angustifolius L.) and varieties of lupin seeds. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Paternostro, R.; Sieghart, W.; Trauner, M.; Pinter, M. Cancer and hepatic steatosis. ESMO Open 2021, 6, 100185. [Google Scholar] [CrossRef] [PubMed]
- Danson, S.; Ward, T.H.; Butler, J.; Ranson, M. DT-diaphorase: A target for new anticancer drugs. Cancer Treat. Rev. 2004, 30, 437–449. [Google Scholar] [CrossRef]
- Cadenas, E. Antioxidant and prooxidant functions of DT-diaphorase in quinone metabolism. Biochem. Pharmacol. 1995, 49, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.; de Ovalle, S.; Cabrera, M.; Cerecetto, H.; González, M. Searching phase II enzymes inducers, from Michael acceptor-[1,2]dithiolethione hybrids, as cancer chemopreventive agents. Future Med. Chem. 2015, 7, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wu, K.; Knox, R. Structure-function studies of DT-diaphorase (NQO1) and NRH: Quinone oxidoreductase (NQO2). Free Radic. Biol. Med. 2000, 29, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Solt, D.B.; Farber, E. Phenotypic diversity as an early property of putative preneoplastic hepatocyte populations in liver carcinogenesis. Cancer Res. 1980, 40, 725–733. [Google Scholar] [PubMed]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidant systems in poultry biology: Superoxide dismutase. J. Anim. Res. Nutr. 2016, 1, 8. [Google Scholar] [CrossRef]
- Santos-Sánchez, G.; Cruz-Chamorro, I.; Álvarez-Ríos, A.I.; Álvarez-Sánchez, N.; Rodríguez-Ortiz, B.; Álvarez-López, A.I.; Fernández-Pachón, M.S.; Pedroche, J.; Millán, F.; Millán-Linares, M.D.C.; et al. Bioactive Peptides from Lupin (Lupinus angustifolius) prevent the early stages of atherosclerosis in western diet-fed ApoE-/- mice. J. Agric. Food Chem. 2022, 70, 8243–8253. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Shang, W.; Strappe, P.; Zhou, Z.; Blanchard, C. Peptides derived from lupin proteins confer potent protection against oxidative stress. J. Sci. Food Agric. 2018, 98, 5225–5234. [Google Scholar] [CrossRef] [PubMed]
- Ścibior, D.; Czeczot, H. Catalase: Structure, properties, functions. Postep. Hig. Med. Dosw. 2006, 60, 170–180. [Google Scholar]
- Niikawa, N.; Fukushima, Y.; Taniguchi, N.; Iizuka, S.; Kajii, T. Chromosome abnormalities involving 11p13 and low erythrocyte catalase activity. Hum. Genet. 1982, 60, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Yasmineh, W.G.; Kaur, T.P.; Blazar, B.R.; Theologides, A. Serum catalase as marker of graft-vs-host disease in allogeneic bone marrow transplant recipients: Pilot study. Clin. Chem. 1995, 41, 1574–1580. [Google Scholar] [CrossRef]
- Chandoga, J. Properties and biogenesis of peroxisomes. Bratisl. Lek. Listy 1994, 95, 543–557. [Google Scholar]
- Marczuk-Krynicka, D.; Hryniewiecki, T.; Paluszak, J.; Krauss, H.; Nowak, D. High Fat Content in diets and oxidative stress in livers of non-diabetic and diabetic rats. Pol. J. Environ. Stud. 2009, 18, 249–253. [Google Scholar]
- Rindler, P.M.; Plafker, S.M.; Szweda, L.I.; Kinter, M. High dietary fat selectively increases catalase expression within cardiac mitochondria. J. Biol. Chem. 2013, 288, 1979–1990. [Google Scholar] [CrossRef]
- Machado, M.V.; Michelotti, G.A.; Xie, G.; Almeida Pereira, T.; Boursier, J.; Bohnic, B.; Guy, C.D.; Diehl, A.M. Mouse models of diet-induced nonalcoholic steatohepatitis reproduce the heterogeneity of the human disease. PLoS ONE 2015, 10, e0127991. [Google Scholar] [CrossRef]
- Noeman, S.A.; Hamooda, H.E.; Baalash, A.A. Biochemical study of oxidative stress markers in the liver, kidney and heart of high fat diet induced obesity in rats. Diabetol. Metab. Syndr. 2011, 3, 17. [Google Scholar] [CrossRef]
- Leclercq, I.A.; Farrell, G.C.; Field, J.; Bell, D.R.; Gonzalez, F.J.; Robertson, G.R. CYP2E1 and CYP4A as microsomal catalysts of lipid peroxides in murine nonalcoholic steatohepatitis. J. Clin. Investig. 2000, 105, 1067–1075. [Google Scholar] [CrossRef]
- Lieber, C.S. Cytochrome P-4502E1: Its physiological and pathological role. Physiol. Rev. 1997, 77, 517–544. [Google Scholar] [CrossRef]
- Puccinelli, E.; Gervasi, P.G.; Longo, V. Xenobiotic metabolizing cytochrome P450 in pig, a promising animal model. Curr. Drug Metab. 2011, 12, 507–525. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.Y.; Lee, D.W.; Park, K.H. Inhibition of 7-ethoxycoumarin O-deethylase activity in rat liver microsomes by naturally occurring flavonoids: Structure-activity relationships. Xenobiotica 1998, 28, 117–126. [Google Scholar] [CrossRef]
- Tanimizu, N.; Ichinohe, N.; Suzuki, H.; Mitaka, T. Prolonged oxidative stress and delayed tissue repair exacerbate acetaminophen-induced liver injury in aged mice. Aging 2020, 12, 18907–18927. [Google Scholar] [CrossRef]
- Gu, Y.F.; Chen, Y.P.; Jin, R.; Wang, C.; Wen, C.; Zhou, Y.M. Age-related changes in liver metabolism and antioxidant capacity of laying hens. Poult. Sci. 2021, 100, 101478. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components, % | Group | ||||
|---|---|---|---|---|---|
| T0 | T10 | T15 | T20 | T30 | |
| Wheat | 57.78 | 53.324 | 50.737 | 48.153 | 45.568 |
| Post-extraction soybean meal | 23.71 | 17.678 | 14.727 | 11.776 | 8.825 |
| Lupinus angustifolius | - | 10 | 15 | 20 | 25 |
| Calcium carbonate | 10.219 | 9.812 | 9.793 | 9.773 | 9.754 |
| Rapeseed oil | 5.976 | 6.77 | 7.268 | 7.766 | 8.264 |
| Monocalcium phosphate | 0.99 | 1.002 | 1.009 | 1.015 | 1.022 |
| Premix without coccidiostat 1 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| NaHCO3 | 0.294 | 0.312 | 0.32 | 0.329 | 0.337 |
| DL-Methionine | 0.234 | 0.251 | 0.261 | 0.27 | 0.279 |
| Sodium chloride | 0.171 | 0.156 | 0.15 | 0.143 | 0.136 |
| Lysine HCL | 0.07 | 0.11 | 0.129 | 0.148 | 0.167 |
| Threonine | 0.029 | 0.038 | 0.042 | 0.047 | 0.051 |
| L-Valine | 0.017 | 0.033 | 0.042 | 0.05 | 0.059 |
| Optiphose (0.01%) 2 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| l-Tryptophan | - | 0.004 | 0.012 | 0.02 | 0.028 |
| Nutritional Value of the Mixture a, % | Group | ||||
|---|---|---|---|---|---|
| T0 | T10 | T15 | T20 | T25 | |
| AMEn 1 | 2850 | 2850 | 2850 | 2850 | 2850 |
| Total protein | 17.99 | 17.89 | 17.88 | 17.92 | 17.95 |
| Raw fat | 8.50 | 8.48 | 9.16 | 9.84 | 10.52 |
| Raw fiber | 3 | 3.99 | 4.48 | 4.97 | 5.47 |
| Digestible lysine | 0.84 | 0.84 | 0.84 | 0.84 | 0.84 |
| Digestible threonine | 0.59 | 0.59 | 0.59 | 0.59 | 0.59 |
| Valine | 0.7 | 0.7 | 0.7 | 0.7 | 0.7 |
| Methionine + cystine | 0.76 | 0.76 | 0.76 | 0.76 | 0.76 |
| Available phosphorus | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Calcium | 4.29 | 4.29 | 4.29 | 4.29 | 4.29 |
| Chlorine | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 |
| Sodium | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wójcik, M.; Grabowski, S.; Jarosz, Ł.S.; Szymczak, B.; Longo, V.; della Croce, C.M.; Hejdysz, M.; Cieślak, A.; Gruszczyński, K.; Marek, A. Liver Antioxidant Capacity and Steatosis in Laying Hens Exposed to Various Quantities of Lupin (Lupinus angustifolius) Seeds in the Diet. Antioxidants 2024, 13, 251. https://doi.org/10.3390/antiox13020251
Wójcik M, Grabowski S, Jarosz ŁS, Szymczak B, Longo V, della Croce CM, Hejdysz M, Cieślak A, Gruszczyński K, Marek A. Liver Antioxidant Capacity and Steatosis in Laying Hens Exposed to Various Quantities of Lupin (Lupinus angustifolius) Seeds in the Diet. Antioxidants. 2024; 13(2):251. https://doi.org/10.3390/antiox13020251
Chicago/Turabian StyleWójcik, Marta, Sebastian Grabowski, Łukasz S. Jarosz, Bartłomiej Szymczak, Vincenzo Longo, Clara Maria della Croce, Marcin Hejdysz, Adam Cieślak, Kamil Gruszczyński, and Agnieszka Marek. 2024. "Liver Antioxidant Capacity and Steatosis in Laying Hens Exposed to Various Quantities of Lupin (Lupinus angustifolius) Seeds in the Diet" Antioxidants 13, no. 2: 251. https://doi.org/10.3390/antiox13020251
APA StyleWójcik, M., Grabowski, S., Jarosz, Ł. S., Szymczak, B., Longo, V., della Croce, C. M., Hejdysz, M., Cieślak, A., Gruszczyński, K., & Marek, A. (2024). Liver Antioxidant Capacity and Steatosis in Laying Hens Exposed to Various Quantities of Lupin (Lupinus angustifolius) Seeds in the Diet. Antioxidants, 13(2), 251. https://doi.org/10.3390/antiox13020251