
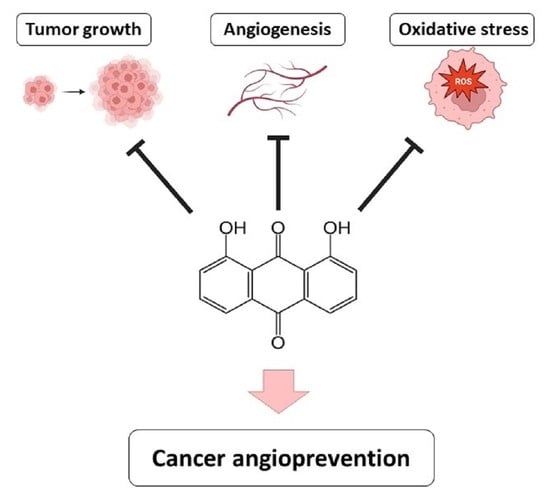
Danthron, an Anthraquinone Isolated from a Marine Fungus, Is a New Inhibitor of Angiogenesis Exhibiting Interesting Antitumor and Antioxidant Properties
 , ,
, ,  ,
,  and
and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of Danthron from the Fermentation Broth of Chromolaenicola sp. (HL-114-33-R04)
2.3. Cell Culture
2.4. Chicken Chorioallantoic Membrane (CAM) Assay
2.5. Tube Formation on Matrigel by Endothelial Cells
2.6. Cell Survival Assay (MTT Assay)
2.7. Proliferation Assay (EdU)
2.8. Cell Cycle Analysis
2.9. Hoechst 33258 Staining
2.10. Zymographic Assay for the Detection of Matrix Metalloproteinase (MMP)
2.11. Invasion Assay
2.12. Wound-Healing Assay to Determine Cell Migration
2.13. Determination of Intracellular Reactive Oxygen Species (ROS) (DCFH-DA Assay)
2.14. Determination of Sulfhydryl Groups
2.15. Statistical Analysis
3. Results
3.1. Isolation of Danthron Form the Fermentation Broth of a Marine Fungus
3.2. Danthron Inhibits Angiogenesis In Vivo in a Dose-Dependent Manner
3.3. Danthron Inhibits the Tube Formation on Matrigel by HUVECs at Non Toxic Doses
3.4. Danthron Inhibits the Growth of Endothelial and Tumor Cells
3.5. Danthron Inhibits the Proteolytic and Invasive Capabilities of Endothelial and Tumor Cells
3.6. Danthron Induces Changes in the Cell Cycle Subpopulations and Apoptosis Only in Tumor Cells
3.7. Danthron Reduces Intracellular ROS Production and Increases the Amount of Intracellular Sulfhydryl Groups in Endothelial and Tumor Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Quesada, A.R.; Medina, M.Á.; Muñoz-Chápuli, R.; Ponce, Á.L. Do not say ever never more: The ins and outs of antiangiogenic therapies. Curr. Pharm. Des. 2010, 16, 3932–3957. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- García-Caballero, M.; Torres-Vargas, J.A.; Marrero, A.D.; Martínez-Poveda, B.; Medina, M.Á.; Quesada, A.R. Angioprevention of Urologic Cancers by Plant-Derived Foods. Pharmaceutics 2022, 14, 256. [Google Scholar] [CrossRef]
- Marrero, A.D.; Quesada, A.R.; Martínez-Poveda, B.; Medina, M.Á. Antiangiogenic Phytochemicals Constituent of Diet as Promising Candidates for Chemoprevention of Cancer. Antioxidants 2022, 11, 302. [Google Scholar] [CrossRef]
- García-Caballero, M.; Marí-Beffa, M.; Medina, M.Á.; Quesada, A.R. Dimethylfumarate inhibits angiogenesis in vitro and in vivo: A possible role for its antipsoriatic effect? J. Investig. Dermatol. 2011, 131, 1347–1355. [Google Scholar] [CrossRef]
- García-Vilas, J.A.; Pino-Ángeles, A.; Martínez-Poveda, B.; Quesada, A.R.; Medina, M.Á. The noni anthraquinone damnacanthal is a multi-kinase inhibitor with potent anti-angiogenic effects. Cancer Lett. 2017, 385, 1–11. [Google Scholar] [CrossRef]
- Carrillo, P.; Martínez-Poveda, B.; Medina, M.Á.; Quesada, A.R. The strigolactone analog GR-24 inhibits angiogenesis in vivo and in vitro by a mechanism involving cytoskeletal reorganization and VEGFR2 signalling. Biochem. Pharmacol. 2019, 168, 366–383. [Google Scholar] [CrossRef]
- Marrero, A.D.; Castilla, L.; Espartero, J.L.; Madrona, A.; Quesada, A.R.; Medina, M.Á.; Martínez-Poveda, B. A comparative study of the antiangiogenic activity of hydroxytyrosyl alkyl ethers. Food Chem. 2020, 333, 127476. [Google Scholar] [CrossRef]
- Rodríguez-Nieto, S.; González-Iriarte, M.; Carmona, R.; Muñoz-Chápuli, R.; Medina, M.A.; Quesada, A.R. Antiangiogenic activity of aeroplysinin-1, a brominated compound isolated from a marine sponge. FASEB J. 2002, 16, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.E.; González-Iriarte, M.; Barrero, A.F.; Salvador-Tormo, N.; Muñoz-Chápuli, R.; Medina, M.A.; Quesada, A.R. Study of puupehenone and related compounds as inhibitors of angiogenesis. Int. J. Cancer 2004, 110, 31–38. [Google Scholar] [CrossRef] [PubMed]
- García-Caballero, M.; Marí-Beffa, M.; Cañedo, L.; Medina, M.Á.; Quesada, A.R. Toluquinol, a marine fungus metabolite, is a new angiosuppresor that interferes with the Akt pathway. Biochem. Pharmacol. 2013, 85, 1727–1740. [Google Scholar] [CrossRef]
- García-Caballero, M.; Cañedo, L.; Fernández-Medarde, A.; Medina, M.Á.; Quesada, A.R. The marine fungal metabolite, AD0157, inhibits angiogenesis by targeting the Akt signaling pathway. Mar. Drugs 2014, 12, 279–299. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine Compounds and Cancer: Updates 2022. Mar. Drugs 2022, 20, 759. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.Y.; Li, H.J.; Li, Q.Y.; Wu, Y.C. Application of marine natural products in drug research. Bioorg. Med. Chem. 2021, 35, 116058. [Google Scholar] [CrossRef] [PubMed]
- Sebak, M.; Molham, F.; Greco, C.; Tammam, M.A.; Sobeh, M.; El-Demerdash, A. Chemical diversity, medicinal potentialities, biosynthesis, and pharmacokinetics of anthraquinones and their congeners derived from marine fungi: A comprehensive update. RSC Adv. 2022, 12, 24887–24921. [Google Scholar] [CrossRef]
- Hafez Ghoran, S.; Taktaz, F.; Ayatollahi, S.A.; Kijjoa, A. Anthraquinones and Their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities. Mar. Drugs 2022, 20, 474. [Google Scholar] [CrossRef]
- Arnaoutova, I.; Kleinman, H.K. In vitro angiogenesis: Endothelial cell tube formation on gelled basement membrane extract. Nat. Protoc. 2010, 5, 628–635. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Tao, L.Y.; Liang, Y.J.; Chen, L.M.; Mi, Y.J.; Zheng, L.S.; Wang, F.; She, Z.G.; Lin, Y.C.; To, K.K.; et al. Anthracenedione derivatives as anticancer agents isolated from secondary metabolites of the mangrove endophytic fungi. Mar. Drugs 2010, 8, 1469–1481. [Google Scholar] [CrossRef]
- Yamazaki, H.; Rotinsulu, H.; Kaneko, T.; Murakami, K.; Fujiwara, H.; Ukai, K.; Namikoshi, M. A new dibenz[b,e]oxepine derivative, 1-hydroxy-10-methoxy-dibenz[b,e]oxepin-6,11-dione, from a marine-derived fungus, Beauveria bassiana TPU942. Mar. Drugs 2012, 10, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Nash, B.D.; Duffy, S.; Avery, V.M. Albopunctatone, an antiplasmodial anthrone-anthraquinone from the Australian ascidian Didemnum albopunctatum. J. Nat. Prod. 2012, 75, 1206–1209. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.J.; Higuchi, R.; Miyamoto, T.; Ono, M.; Kuwano, M.; Mawatari, S.F. Bryoanthrathiophene, a new antiangiogenic constituent from the bryozoan Watersipora subtorquata (d’Orbigny, 1852). J. Nat. Prod. 2002, 65, 1344–1345. [Google Scholar] [CrossRef] [PubMed]
- Baars, A.J.; Vermeulen, R.J.; Breimer, D.D. Gas chromatographic determination of the laxative 1, 8-dihydroxyanthraquinone in urine and faeces. J. Chromatogr. 1976, 120, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.E.E. Martindale. The Extra Pharmacopeia, 29th ed.; The Pharmaceutical Press: London, UK, 1989; pp. 1073–1112. [Google Scholar]
- Huang, Q.; Lu, G.; Shen, H.M.; Chung, M.C.; Ong, C.N. Anti-cancer properties of anthraquinones from rhubarb. Med. Res. Rev. 2007, 27, 609–630. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, Q.; Kang, L.; Yuan, L.; Chen, G.; Cui, X.; Wang, L.; Zhang, T.; Wang, L. Rheum officinale and Salvia miltiorrhiza inhibit renal fibrosis via miR-21/PTEN/Akt signaling pathway in vitro and in vivo. J. Ethnopharmacol. 2023, 304, 115928. [Google Scholar] [CrossRef]
- National Toxicology Program. Danthron (1,8-dihydroxyanthraquinone). Rep. Carcinog. 2002, 10, 76–77, Erratum in Rep. Carcinog. 2004, 11, III77–III78. [Google Scholar]
- Bennett, M.; Cresswell, H. Factors influencing constipation in advanced cancer patients: A prospective study of opioid dose, dantron dose and physical functioning. Palliat. Med. 2003, 17, 418–422. [Google Scholar] [CrossRef]
- Liudvytska, O.; Kolodziejczyk-Czepas, J. A Review on Rhubarb-Derived Substances as Modulators of Cardiovascular Risk Factors-A Special Emphasis on Anti-Obesity Action. Nutrients 2022, 14, 2053. [Google Scholar] [CrossRef]
- Nguyen, L.T.H.; Ahn, S.H.; Shin, H.M.; Yang, I.J. Anti-Psoriatic Effect of Rheum palmatum L. and Its Underlying Molecular Mechanisms. Int. J. Mol. Sci. 2022, 23, 16000. [Google Scholar] [CrossRef]
- Muhlemann, H. Uber Anthrachinone und Anthrachinonglykoside [Anthrachinons and anthrachinon glycosides]. Pharm. Acta Helv. 1952, 27, 9–17. [Google Scholar] [PubMed]
- Ocaña, M.C.; Martínez-Poveda, B.; Marí-Beffa, M.; Quesada, A.R.; Medina, M.Á. Fasentin diminishes endothelial cell proliferation, differentiation and invasion in a glucose metabolism-independent manner. Sci. Rep. 2020, 10, 6132. [Google Scholar] [CrossRef] [PubMed]
- Torres-Vargas, J.A.; Cheng-Sánchez, I.; Martínez-Poveda, B.; Medina, M.Á.; Sarabia, F.; García-Caballero, M.; Quesada, A.R. Characterization of the activity and the mechanism of action of a new toluquinol derivative with improved potential as an antiangiogenic drug. Biomed. Pharmacother. 2022, 155, 113759. [Google Scholar] [CrossRef] [PubMed]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.; Jones, E.G.; Bhat, D.J.; Jeewon, R.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 171–175. [Google Scholar] [CrossRef]
- Richardson, M.; Singh, G. Observations on the use of the avian chorioallantoic membrane (CAM) model in investigations into angiogenesis. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2003, 3, 155–185. [Google Scholar] [CrossRef]
- Fischer, D.; Fluegen, G.; Garcia, P.; Ghaffari-Tabrizi-Wizsy, N.; Gribaldo, L.; Huang, R.Y.; Rasche, V.; Ribatti, D.; Rousset, X.; Pinto, M.T.; et al. The CAM Model-Q&A with Experts. Cancers 2022, 15, 191. [Google Scholar] [CrossRef]
- Cárdenas, C.; Quesada, A.R.; Medina, M.A. Effects of ursolic acid on different steps of the angiogenic process. Biochem. Biophys. Res. Commun. 2004, 320, 402–408. [Google Scholar] [CrossRef]
- Martínez-Poveda, B.; Quesada, A.R.; Medina, M.A. Hyperforin, a bio-active compound of St. John’s Wort, is a new inhibitor of angiogenesis targeting several key steps of the process. Int. J. Cancer 2005, 117, 775–780. [Google Scholar] [CrossRef]
- Crawford, A.D.; Liekens, S.; Kamuhabwa, A.R.; Maes, J.; Munck, S.; Busson, R.; Rozenski, J.; Esguerra, C.V.; de Witte, P.A. Zebrafish bioassay-guided natural product discovery: Isolation of angiogenesis inhibitors from East African medicinal plants. PLoS ONE 2011, 6, e14694. [Google Scholar] [CrossRef]
- Cárdenas, C.; Quesada, A.R.; Medina, M.A. Evaluation of the anti-angiogenic effect of aloe-emodin. Cell. Mol. Life Sci. 2006, 63, 3083–3089. [Google Scholar] [CrossRef]
- Ribatti, D.; Nico, B.; Crivellato, E.; Roccaro, A.M.; Vacca, A. The history of the angiogenic switch concept. Leukemia 2007, 21, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.F.; Wang, H.L.; Chuang, Y.Y.; Tang, Y.J.; Yang, J.S.; Ma, Y.S.; Chiang, J.H.; Lu, C.C.; Yang, J.L.; Lai, T.Y.; et al. Danthron induced apoptosis through mitochondria- and caspase-3-dependent pathways in human brain glioblastoma multiforms GBM 8401 cells. Neurochem. Res. 2010, 35, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Rundhaug, J.E. Matrix metalloproteinases and angiogenesis. J. Cell. Mol. Med. 2005, 9, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Ben-Yosef, Y.; Miller, A.; Shapiro, S.; Lahat, N. Hypoxia of endothelial cells leads to MMP-2-dependent survival and death. Am. J. Physiol. Cell Physiol. 2005, 289, C1321–C1331. [Google Scholar] [CrossRef]
- Lin, C.C.; Chen, J.T.; Yang, J.S.; Lu, H.F.; Hsu, S.C.; Tan, T.W.; Lin, Y.T.; Ma, Y.S.; Ip, S.W.; Wu, J.J.; et al. Danthron inhibits the migration and invasion of human brain glioblastoma multiforme cells through the inhibition of mRNA expression of focal adhesion kinase, Rho kinases-1 and metalloproteinase-9. Oncol. Rep. 2009, 22, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.H.; Yang, J.S.; Ma, C.Y.; Yang, M.D.; Huang, H.Y.; Hsia, T.C.; Kuo, H.M.; Wu, P.P.; Lee, T.H.; Chung, J.G. Danthron, an anthraquinone derivative, induces DNA damage and caspase cascades-mediated apoptosis in SNU-1 human gastric cancer cells through mitochondrial permeability transition pores and Bax-triggered pathways. Chem. Res. Toxicol. 2011, 24, 20–29. [Google Scholar] [CrossRef]
- Chiou, S.M.; Chiu, C.H.; Yang, S.T.; Yang, J.S.; Huang, H.Y.; Kuo, C.L.; Chen, P.Y.; Chung, J.G. Danthron triggers ROS and mitochondria-mediated apoptotic death in C6 rat glioma cells through caspase cascades, apoptosis-inducing factor and endonuclease G multiple signaling. Neurochem. Res. 2012, 37, 1790–1800. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Tang, J.Y.; Ou-Yang, F.; Hou, M.F.; Huang, H.W.; Wang, H.R.; Li, K.T.; Fayyaz, S.; Shu, C.W.; Chang, H.W. Oxidative stress-modulating drugs have preferential anticancer effects—Involving the regulation of apoptosis, DNA damage, endoplasmic reticulum stress, autophagy, metabolism, and migration. Semin. Cancer Biol. 2019, 58, 109–117. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Koh, J.Y.; Song, D.K.; Kim, H.C.; Kwon, M.S.; Choi, Y.S.; Wie, M.B. Danthron inhibits the neurotoxicity induced by various compounds causing oxidative damages including beta-amyloid (25–35) in primary cortical cultures. Biol. Pharm. Bull. 2004, 27, 723–726. [Google Scholar] [CrossRef]
- Kaneshiro, T.; Morioka, T.; Inamine, M.; Kinjo, T.; Arakaki, J.; Chiba, I.; Sunagawa, N.; Suzui, M.; Yoshimi, N. Anthraquinone derivative emodin inhibits tumor-associated angiogenesis through inhibition of extracellular signal-regulated kinase 1/2 phosphorylation. Eur. J. Pharmacol. 2006, 553, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Wu, S.Y.; Zhen, Y.S. Inhibitory effects of emodin on angiogenesis. Yao Xue Xue Bao 2004, 39, 254–258. [Google Scholar] [PubMed]
- Rossi, S.; Tabolacci, C.; Lentini, A.; Provenzano, B.; Carlomosti, F.; Frezzotti, S.; Beninati, S. Anthraquinones danthron and quinizarin exert antiproliferative and antimetastatic activity on murine B16-F10 melanoma cells. Anticancer Res. 2010, 30, 445–449. [Google Scholar] [PubMed]
- Chou, Y.T.; Hsu, F.F.; Hu, D.Y.; Chen, Y.C.; Hsu, Y.H.; Hsu, J.T.; Chau, L.Y. Identification of danthron as an isoform-specific inhibitor of HEME OXYGENASE-1/cytochrome P450 reductase interaction with anti-tumor activity. J. Biomed. Sci. 2018, 25, 6. [Google Scholar] [CrossRef]
- Fang, J.; Shing, Y.; Wiederschain, D.; Yan, L.; Butterfield, C.; Jackson, G.; Harper, J.; Tamvakopoulos, G.; Moses, M.A. Matrix metalloproteinase-2 is required for the switch to the angiogenic phenotype in a tumor model. Proc. Natl. Acad. Sci. USA 2000, 97, 3884–3889. [Google Scholar] [CrossRef]
- Itoh, T.; Tanioka, M.; Yoshida, H.; Yoshioka, T.; Nishimoto, H.; Itohara, S. Reduced angiogenesis and tumor progression in gelatinase A-deficient mice. Cancer Res. 1998, 58, 1048–1051. [Google Scholar]
- García-Vilas, J.A.; Quesada, A.R.; Medina, M.Á. Hydroxytyrosol targets extracellular matrix remodeling by endothelial cells and inhibits both ex vivo and in vivo angiogenesis. Food Chem. 2017, 221, 1741–1746. [Google Scholar] [CrossRef]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef]
- Yang, G.; Chang, C.C.; Yang, Y.; Yuan, L.; Xu, L.; Ho, C.T.; Li, S. Resveratrol Alleviates Rheumatoid Arthritis via Reducing ROS and Inflammation, Inhibiting MAPK Signaling Pathways, and Suppressing Angiogenesis. J. Agric. Food Chem. 2018, 66, 12953–12960. [Google Scholar] [CrossRef]
- Khan, F.B.; Singh, P.; Jamous, Y.F.; Ali, S.A.; Abdullah; Uddin, S.; Zia, Q.; Jena, M.K.; Khan, M.; Owais, M.; et al. Multifaceted Pharmacological Potentials of Curcumin, Genistein, and Tanshinone IIA through Proteomic Approaches: An In-Depth Review. Cancers 2022, 15, 249. [Google Scholar] [CrossRef]
- He, X.; Wang, N.; Zhang, Y.; Huang, X.; Wang, Y. The therapeutic potential of natural products for treating pancreatic cancer. Front. Pharmacol. 2022, 13, 1051952. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Garg, V.K.; Mehta, J.K.; Kaur, G.; Mohapatra, R.K.; Dhama, K.; Sak, K.; Kumar, A.; Varol, M.; Aggarwal, D.; et al. Licorice (Glycyrrhiza glabra L.)-Derived Phytochemicals Target Multiple Signaling Pathways to Confer Oncopreventive and Oncotherapeutic Effects. OncoTargets Ther. 2022, 15, 1419–1448. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, S.; Iqbal, J.; Abbasi, B.A.; Ullah, Z.; Yaseen, T.; Kanwal, S.; Mahmood, T.; Sydykbayeva, S.; Ydyrys, A.; Almarhoon, Z.M.; et al. Rosmarinic acid and its derivatives: Current insights on anticancer potential and other biomedical applications. Biomed. Pharmacother. 2023, 162, 114687. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Tertil, M.; Jozkowicz, A.; Dulak, J. Oxidative stress in tumor angiogenesis- therapeutic targets. Curr. Pharm. Des. 2010, 16, 3877–3894. [Google Scholar] [CrossRef]
- Prasad, S.; Gupta, S.C.; Tyagi, A.K. Reactive oxygen species (ROS) and cancer: Role of antioxidative nutraceuticals. Cancer Lett. 2017, 387, 95–105. [Google Scholar] [CrossRef]
- Jing, W.; Liu, C.; Su, C.; Liu, L.; Chen, P.; Li, X.; Zhang, X.; Yuan, B.; Wang, H.; Du, X. Role of reactive oxygen species and mitochondrial damage in rheumatoid arthritis and targeted drugs. Front. Immunol. 2023, 14, 1107670. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidal, I.; Torres-Vargas, J.A.; Sánchez, J.M.; Trigal, M.; García-Caballero, M.; Medina, M.Á.; Quesada, A.R. Danthron, an Anthraquinone Isolated from a Marine Fungus, Is a New Inhibitor of Angiogenesis Exhibiting Interesting Antitumor and Antioxidant Properties. Antioxidants 2023, 12, 1101. https://doi.org/10.3390/antiox12051101
Vidal I, Torres-Vargas JA, Sánchez JM, Trigal M, García-Caballero M, Medina MÁ, Quesada AR. Danthron, an Anthraquinone Isolated from a Marine Fungus, Is a New Inhibitor of Angiogenesis Exhibiting Interesting Antitumor and Antioxidant Properties. Antioxidants. 2023; 12(5):1101. https://doi.org/10.3390/antiox12051101
Chicago/Turabian StyleVidal, Isabel, José Antonio Torres-Vargas, José María Sánchez, Mónica Trigal, Melissa García-Caballero, Miguel Ángel Medina, and Ana R. Quesada. 2023. "Danthron, an Anthraquinone Isolated from a Marine Fungus, Is a New Inhibitor of Angiogenesis Exhibiting Interesting Antitumor and Antioxidant Properties" Antioxidants 12, no. 5: 1101. https://doi.org/10.3390/antiox12051101
APA StyleVidal, I., Torres-Vargas, J. A., Sánchez, J. M., Trigal, M., García-Caballero, M., Medina, M. Á., & Quesada, A. R. (2023). Danthron, an Anthraquinone Isolated from a Marine Fungus, Is a New Inhibitor of Angiogenesis Exhibiting Interesting Antitumor and Antioxidant Properties. Antioxidants, 12(5), 1101. https://doi.org/10.3390/antiox12051101