
Early Physiological, Cytological and Antioxidative Responses of the Edible Halophyte Chenopodium quinoa Exposed to Salt Stress
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
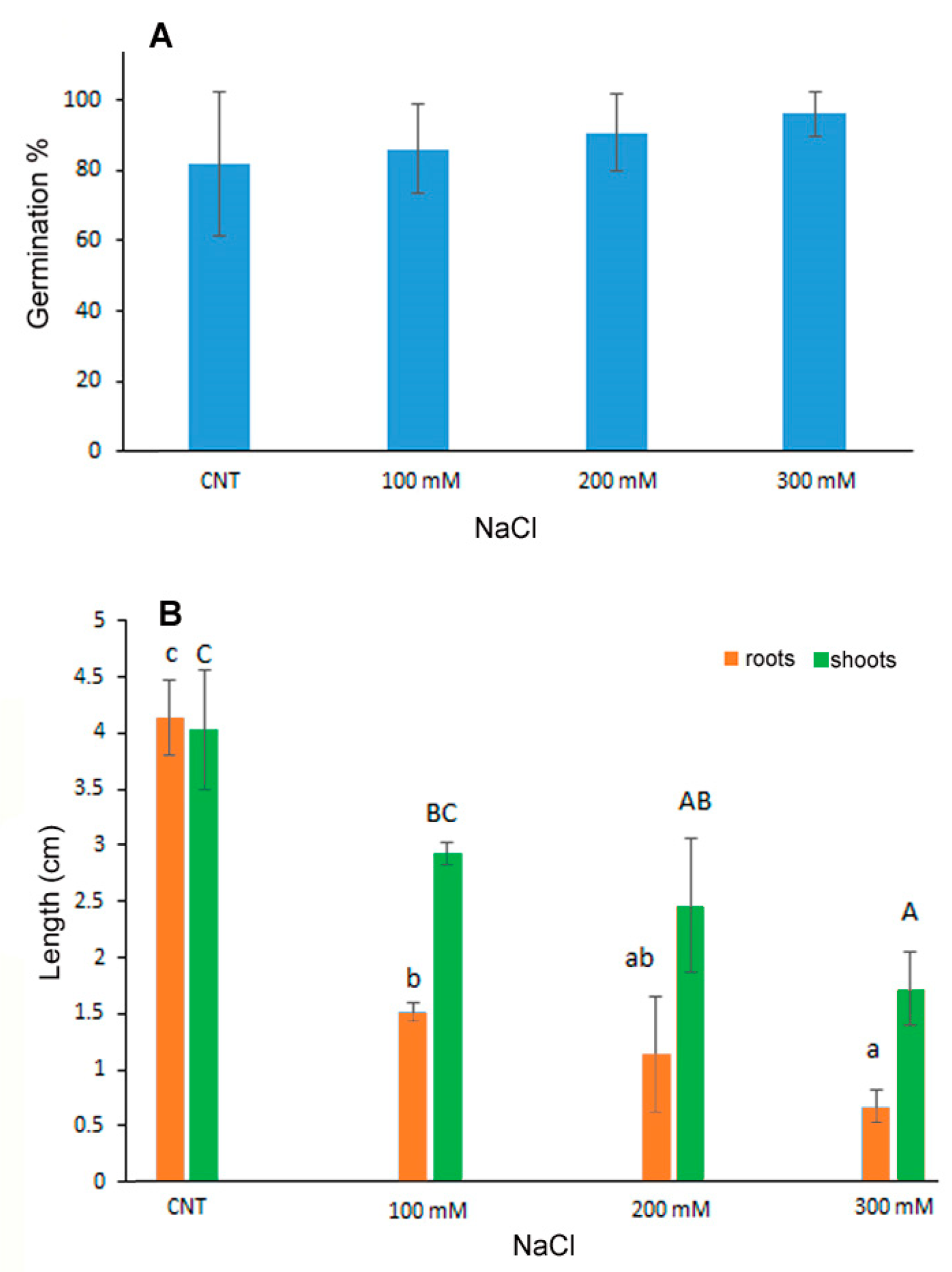
2.1. Seed Germination and Seedling Growth Conditions
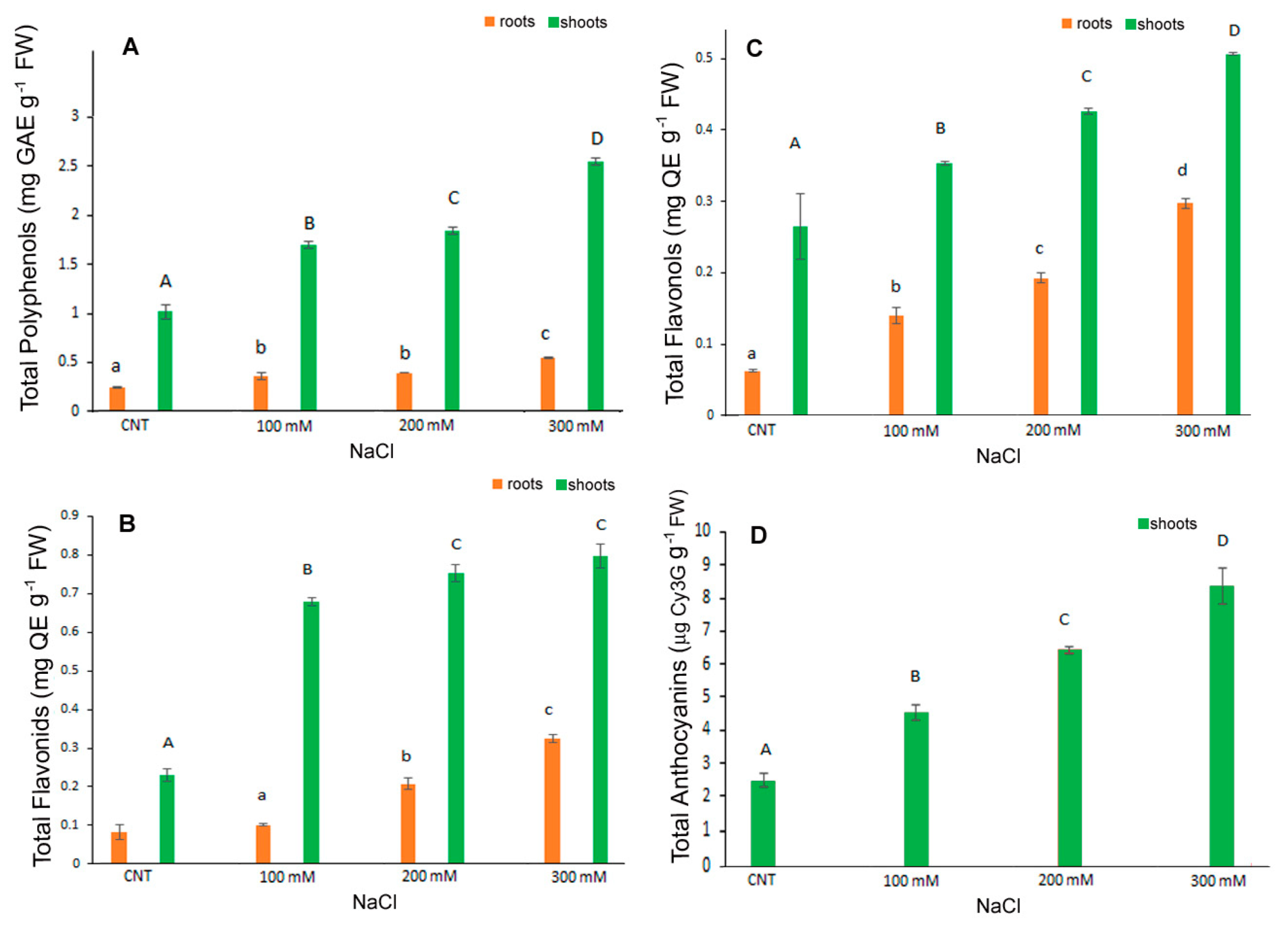
2.2. Total Content of Polyphenols, Flavonoids, Flavonols and Anthocyanins
2.3. Antioxidant Activity by Chemical Assays and Antioxidant Enzymes
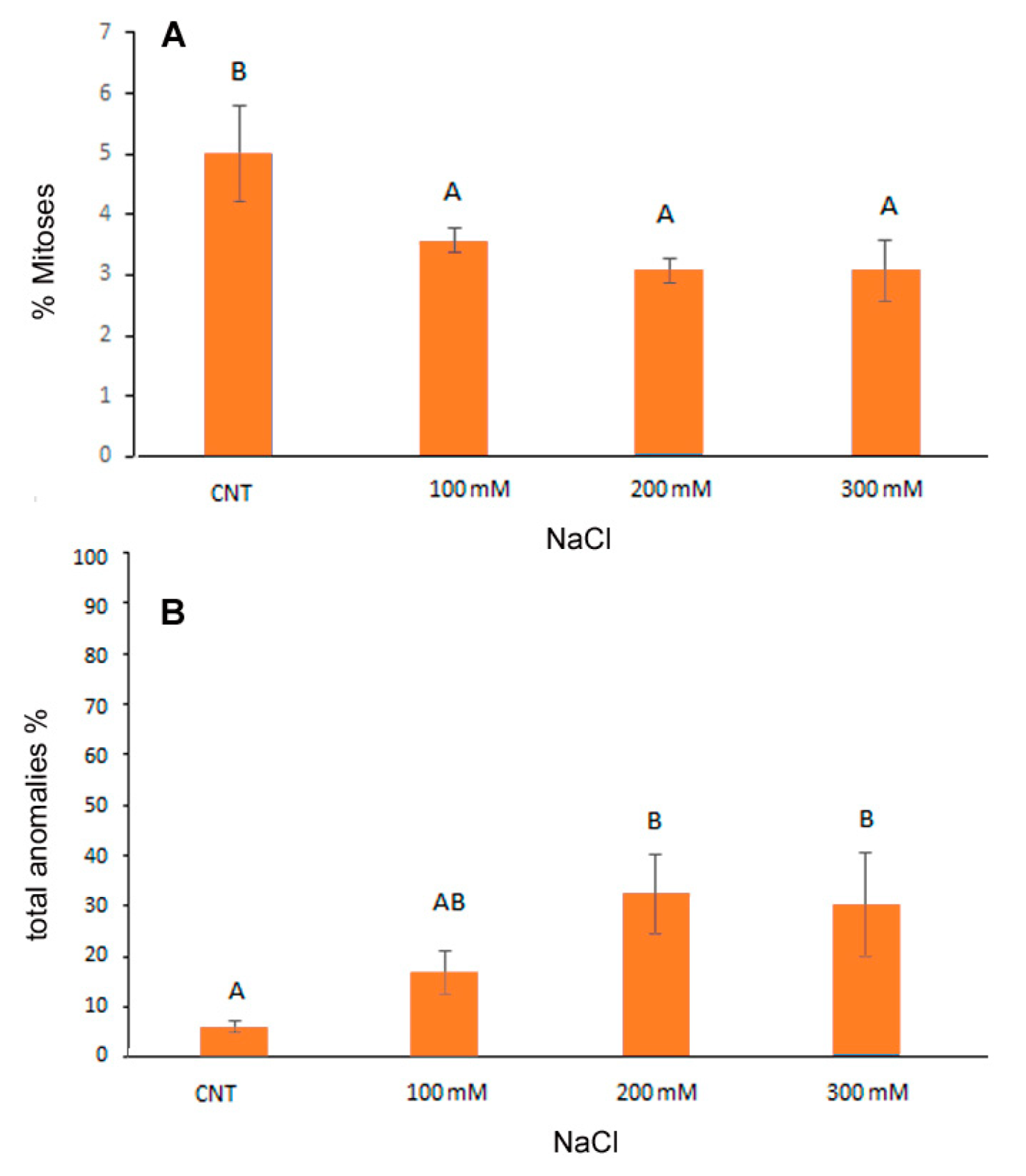
2.4. Cytological Analysis of Root Meristems
2.5. Mineral Nutrient Content
2.6. Statistical Analysis
3. Results and Discussion
3.1. Seed Germination and Seedling Growth
3.2. Cytogenetic Analysis of Root Meristems
3.3. Phenolic Compounds and Antioxidant Activity
3.4. Antioxidant Enzyme Activities
3.5. Mineral Nutrient Content
3.6. Pearson’s Correlation and PCA Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pellegrini, M.; Lucas-Gonzales, R.; Ricci, A.; Fontecha, J.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical, fatty acid, polyphenolic profile, techno-functional and antioxidant properties of flours obtained from quinoa (Chenopodium quinoa Willd.) seeds. Ind. Crops Prod. 2018, 111, 38–46. [Google Scholar] [CrossRef]
- Präger, A.; Munz, S.; Nkebiwe, M.P.; Mast, B.; Graeff-Hönninger, S. Yield and Quality Characteristics of Different Quinoa (Chenopodium quinoa Willd.) Cultivars Grown under Field Conditions in Southwestern Germany. Agronomy 2018, 8, 197. [Google Scholar] [CrossRef]
- Filho, A.M.M.; Pirozi, M.R.; Borges, J.T.D.S.; Sant’Ana, H.M.P.; Chaves, J.B.P.C.; Coimbra, J.S.D.R. Quinoa: Nutritional, functional and antinutritional aspects. Crit. Rev. Food Sci. Nutr. 2017, 57, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Ceyhun-Sezgin, A.; Sanlier, N. A new generation plant for the conventional cuisine: Quinoa (Chenopodium quinoa Willd.) [Review]. Trends Food Sci. Technol. 2019, 86, 51–58. [Google Scholar] [CrossRef]
- Bazile, D.; Jacobsen, S.E.; Verniau, A. The Global Expansion of Quinoa: Trends and Limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef]
- Vita, F.; Ghignone, S.; Bazihizina, N.; Rasouli, F.; Sabbatini, L.; Kiani-Pouya, A.; Kiferle, C.; Shabala, S.; Balestrini, R.; Mancuso, S. Early responses to salt stress in quinoa genotypes with opposite behavior. Physiol. Plant. 2021, 173, 1392–1420. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa Abiotic Stress Responses: A Review. Plants 2018, 7, 106. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Oses, R.; Acuña-Rodríguez, I.S.; Antognoni, F.; Martinez-Mosqueira, E.A.; Coulibaly, A.; Canahua-Murillo, A.; Pinto, M.; Zurita-Silva, A.; et al. Quinoa biodiversity and sustainability for food security under climate change. A review. Agron. Sustain. Dev. 2014, 34, 349–359. [Google Scholar] [CrossRef]
- Kiani-Pouya, A.; Roessner, U.; Jayasinghe, N.S.; Lutz, A.; Rupasinghe, T.; Bazihizina, N.; Bohm, J.; Alharbi, S.; Hedrich, R.; Shabala, S. Epidermal bladder cells confer salinity stress tolerance in the halophyte quinoa and Atriplex species. Plant Cell Environ. 2017, 40, 1900–1915. [Google Scholar] [CrossRef]
- Maamri, K.; Zidane, O.D.; Chaabena, A.; Fiene, G.; Bazile, D. Adaptation of Some Quinoa Genotypes (Chenopodium quinoa Willd.), Grown in a Saharan Climate in Algeria. Life 2022, 12, 1854. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, S.E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.E. Quinoa—A model crop for understanding salt tolerance mechanisms in halophytes. Plant Biosyst. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Aloisi, I.; Parrotta, L.; Ruiz, K.B.; Landi, C.; Bini, L.; Cai, G.; Biondi, S.; Del Duca, S. New Insight into Quinoa Seed Quality under Salinity: Changes in Proteomic and Amino Acid Profiles, Phenolic Content, and Antioxidant Activity of Protein Extracts. Front. Plant Sci. 2016, 7, 656. [Google Scholar] [CrossRef]
- Bellani, L.; Muccifora, S.; Giorgetti, L. Impact of sprouting and cooking on antioxidant compounds and activity in different Italian varieties of Lens culinaris L. J. Food Meas. Charact. 2020, 14, 333–342. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetric of total phenols with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical Activity and Polyphenol Composition of Local Brassicaceae Edible Varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Romani, A.; Mancini, P.A.; Tatti, S.; Vincieri, F. Polyphenols and polysaccharides in Tuscan grapes and wines. Ital. J. Food Sci. 1996, 1, 13–24. [Google Scholar]
- Landi, M.; Guidi, L.; Pardossi, A.; Tattini, M.; Gould, K.S. Photoprotection by foliar anthocyanins mitigates effects of boron toxicity in sweet basil (Ocimum basilicum). Planta 2014, 240, 941–953. [Google Scholar] [CrossRef]
- Boudjou, S.; Oomah, B.D.; Zaidi, F.; Hosseinian, F. Phenolics content and antioxidant and anti-inflammatory activities of legume fractions. Food Chem. 2013, 138, 1543–1550. [Google Scholar] [CrossRef]
- Ninfali, P.; Mea, G.; Giorgini, S.; Rocchi, M.; Bacchiocca, M. Antioxidant capacity of vegetables, spices and dressings relevant to nutrition. Br. J. Nutr. 2005, 93, 257–266. [Google Scholar] [CrossRef]
- Pereira, G.J.G.; Molina, S.M.G.; Lea, P.J.; Azevedo, R.A. Activity of antioxidant enzymes in response to cadmium in Crotalaria juncea. Plant Soil 2002, 239, 123–132. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Muccifora, S.; Giorgetti, L.; Corsini, M.; Di Florio, G.; Bellani, L. Nano and submicron fluorescent polystyrene particles internalization and translocation in seedlings of Cichorium endivia L. Environ. Sci. Nano 2022, 9, 4585. [Google Scholar] [CrossRef]
- Böhm, N.; Sprenger, E. Fluorescence cytophotometry: A valuable method for the quantitative determination of nuclear Feulgen-DNA. Histochemie 1968, 16, 100–118. [Google Scholar] [CrossRef]
- Bellani, L.; Siracusa, G.; Giorgetti, L.; Di Gregorio, S.; Ruffini Castiglione, M.; Spanò, C.; Muccifora, S.; Bottega, S.; Pini, R.; Tassi, E. TiO2 nanoparticles in a biosolid-amended soil and their implication in soil nutrients, microorganisms and Pisum sativum nutrition. Ecotoxicol. Environ. Saf. 2020, 190, 110095. [Google Scholar] [CrossRef]
- Causin, H.F.; Bordón, D.A.E.; Burrieza, H.P. Salinity tolerance mechanisms during germination and early seedling growth in Chenopodium quinoa Wild. genotypes with different sensitivity to saline stress. Environ. Exp. Bot. 2020, 172, 10399. [Google Scholar] [CrossRef]
- Manaa, A.; Goussi, R.; Derbali, W.; Cantamessa, S.; Abdelly, C.; Barbato, R. Salinity tolerance of quinoa (Chenopodium quinoa Willd) as assessed by chloroplast ultrastructure and photosynthetic performance. Environ. Exp. Bot. 2019, 162, 103–114. [Google Scholar] [CrossRef]
- Derbali, W.; Goussi, R.; Koyro, H.W.; Abdelly, C.; Manaa, A. Physiological and biochemical markers for screening salt tolerant quinoa genotypes at early seedling stage. J. Plant Interact. 2020, 15, 27–38. [Google Scholar] [CrossRef]
- Stoleru, V.; Slabu, C.; Vitanescu, M.; Pères, C.; Cojocaru, A.; Covasa, M.; Mihalache, G. Tolerance of Three Quinoa Cultivars (Chenopodium quinoa Willd.) to Salinity and Alkalinity Stress During Germination Stage. Agronomy 2019, 9, 287. [Google Scholar] [CrossRef]
- Krak, K.; Vít, P.; Belyayev, A.; Douda, J.; Hreusová, L.; Mandák, B. Allopolyploid origin of Chenopodium album str. (Chenopodiaceae): A molecular and cytogenetic insight. PLoS ONE 2016, 11, e0161063. [Google Scholar] [CrossRef]
- Mandák, B.; Krak, K.; Vít, P.; Pavlíková, Z.; Lomonosova, M.N.; Habibi, F.; Wang, L.; Jellen, E.N.; Douda, J. How genome size variation is linked with evolution within Chenopodium sensu lato. Perspect. Plant Ecol. Evol. Syst. 2016, 23, 18–32. [Google Scholar] [CrossRef]
- Chatterjee, J.; Majumder, A.L. Salt-induced abnormalities on root tip mitotic cells of Allium cepa: Prevention by inositol pretreatment. Protoplasma 2010, 245, 165–172. [Google Scholar] [CrossRef]
- Younis, M.E.; Rizwan, M.; Tourky, S.M.N. Assessment of early physiological and biochemical responses in chia (Salvia hispanica L.) sprouts under salt stress. Acta Physiol. Plant. 2021, 43, 121. [Google Scholar] [CrossRef]
- Antognoni, F.; Potente, G.; Biondi, S.; Mandrioli, R.; Marincich, L.; Ruiz, K.B. Free and Conjugated Phenolic Profiles and Antioxidant Activity in Quinoa Seeds and Their Relationship with Genotype and Environment. Plants 2021, 10, 1046. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995. [Google Scholar]
- Rouphael, Y.; Petropoulos, A.S.; Cardarelli, M.T.; Colla, G. Salinity as eustressor for enhancing quality of vegetables. Sci. Hortic. 2018, 234, 361–369. [Google Scholar] [CrossRef]
- Maathuis, F.J.M. Sodium in plants: Perception, signalling, and regulation of sodium fluxes. J. Exp. Bot. 2014, 65, 849–858. [Google Scholar] [CrossRef]
- Waters, S.; Gilliham, M.; Hrmova, M. Plant High-Affinity Potassium (HKT) Transporters Involved in Salinity Tolerance: Structural Insights to Probe Differences in Ion Selectivity. Int. J. Mol. Sci. 2013, 14, 7660–7680. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Z. The role of calcium in salt toxicity. Plant Cell Environ. 1992, 15, 625–632. [Google Scholar] [CrossRef]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | NaCl 100 mM | NaCl 200 mM | NaCl 300 mM | |
|---|---|---|---|---|
| %Prophases | 24.1 ± 8.0 b | 24.2 ± 5.8 b | 23.8 ± 3.7 b | 9.0 ± 3.1 a |
| %Metaphases | 34.1 ± 5.5 b | 15.1 ± 4.9 a | 30.6 ± 8.3 b | 34.3 ± 4.2 b |
| %Abn Metaphases | 4.1 ± 0.7 a | 11.0 ± 5 ab | 27.1 ± 6.2 b | 23.6 ± 9.7 b |
| %Ana/Telophases | 35.9 ± 3.6 bc | 44.0 ± 5.3 c | 13.0 ± 3.3 a | 26.2 ± 9.6 ab |
| %Abn Ana/Telophases | 1.8 ± 1.7 a | 5.7 ± 3 a | 5.4 ± 1.9 a | 6.8 ± 3.69 a |
| NaCl (mM) | Na+/K+ | |
|---|---|---|
| Roots | Shoots | |
| 0 | 0.04 | 0.05 |
| 100 | 1.41 | 0.90 |
| 200 | 2.56 | 2.44 |
| 300 | 5.15 | 3.74 |
| Variables | Length | TPC | TFC | Flavonols | Anthocyanin | DPPH | ORAC | SOD | GPX | APX | CAT | Ca | Na | Mg | K | Root 0 mM | Root 100 mM | Root 200 mM | Root 300 mM | Shoot 0 mM | Shoot 100 mM | Shoot 200 mM | Shoot 300 mM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Length | 1 | −0.185 | −0.114 | −0.225 | 0.090 | −0.102 | 0.157 | −0.183 | −0.642 | −0.663 | −0.730 | 0.085 | −0.685 | 0.583 | 0.824 | 0.550 | −0.243 | −0.359 | −0.500 | 0.515 | 0.179 | 0.041 | −0.182 |
| TPC | 1 | 0.906 | 0.880 | 0.929 | 0.986 | 0.834 | 0.956 | 0.609 | 0.622 | 0.196 | −0.031 | 0.802 | 0.008 | −0.495 | −0.309 | −0.254 | −0.238 | −0.165 | −0.309 | 0.054 | 0.445 | 0.776 | |
| TFC | 1 | 0.938 | 0.927 | 0.948 | 0.925 | 0.846 | 0.513 | 0.431 | 0.094 | −0.291 | 0.731 | 0.066 | −0.474 | −0.425 | −0.400 | −0.257 | −0.097 | −0.227 | 0.382 | 0.482 | 0.542 | ||
| Flavonols | 1 | 0.892 | 0.899 | 0.891 | 0.876 | 0.604 | 0.554 | 0.256 | −0.241 | 0.782 | 0.203 | −0.579 | −0.593 | −0.381 | −0.238 | 0.046 | −0.041 | 0.2 | 0.394 | 0.615 | |||
| Anthocyanin | 1 | 0.966 | 0.966 | 0.889 | 0.362 | 0.357 | −0.079 | −0.195 | 0.590 | 0.283 | −0.280 | −0.328 | −0.328 | −0.328 | −0.328 | −0.030 | 0.217 | 0.447 | 0.678 | ||||
| DPPH | 1 | 0.900 | 0.922 | 0.530 | 0.524 | 0.096 | −0.120 | 0.747 | 0.055 | −0.442 | −0.318 | −0.295 | −0.266 | −0.219 | −0.261 | 0.191 | 0.431 | 0.738 | |||||
| ORAC | 1 | 0.789 | 0.257 | 0.212 | −0.176 | −0.361 | 0.496 | 0.355 | −0.222 | −0.408 | −0.35 | −0.340 | −0.323 | 0.075 | 0.374 | 0.438 | 0.533 | ||||||
| SOD | 1 | 0.634 | 0.654 | 0.258 | 0.011 | 0.787 | 0.145 | −0.486 | −0.352 | −0.285 | −0.237 | −0.077 | −0.176 | −0.137 | 0.566 | 0.698 | |||||||
| GPX | 1 | 0.932 | 0.839 | 0.291 | 0.893 | −0.327 | −0.787 | −0.312 | −0.201 | −0.097 | 0.588 | −0.482 | −0.201 | 0.182 | 0.523 | ||||||||
| APX | 1 | 0.829 | 0.367 | 0.879 | −0.336 | −0.811 | −0.287 | −0.175 | 0.062 | 0.454 | −0.444 | −0.357 | 0.091 | 0.657 | |||||||||
| CAT | 1 | 0.453 | 0.665 | −0.37 | −0.743 | −0.222 | −0.164 | 0.088 | 0.829 | −0.357 | −0.353 | −0.122 | 0.301 | ||||||||||
| Ca | 1 | −0.009 | −0.058 | 0.078 | 0.672 | −0.218 | −0.181 | 0.342 | −0.103 | −0.568 | −0.253 | 0.310 | |||||||||||
| Na | 1 | −0.340 | −0.878 | −0.516 | −0.178 | 0.023 | 0.319 | −0.522 | −0.041 | 0.299 | 0.616 | ||||||||||||
| Mg | 1 | 0.466 | −0.115 | −0.158 | −0.475 | −0.175 | 0.895 | −0.135 | 0.152 | 0.011 | |||||||||||||
| K | 1 | 0.6 | 0.232 | −0.412 | −0.417 | 0.433 | 0.046 | −0.046 | −0.436 | ||||||||||||||
| −1 | −0.9 | −0.8 | −0.7 | −0.6 | −0.5 | −0.4 | −0.3 | −0.2 | −0.1 | 0 | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 | 0.6 | 0.7 | 0.8 | 0.9 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souid, A.; Bellani, L.; Tassi, E.L.; Ben Hamed, K.; Longo, V.; Giorgetti, L. Early Physiological, Cytological and Antioxidative Responses of the Edible Halophyte Chenopodium quinoa Exposed to Salt Stress. Antioxidants 2023, 12, 1060. https://doi.org/10.3390/antiox12051060
Souid A, Bellani L, Tassi EL, Ben Hamed K, Longo V, Giorgetti L. Early Physiological, Cytological and Antioxidative Responses of the Edible Halophyte Chenopodium quinoa Exposed to Salt Stress. Antioxidants. 2023; 12(5):1060. https://doi.org/10.3390/antiox12051060
Chicago/Turabian StyleSouid, Aymen, Lorenza Bellani, Eliana Lanfranca Tassi, Karim Ben Hamed, Vincenzo Longo, and Lucia Giorgetti. 2023. "Early Physiological, Cytological and Antioxidative Responses of the Edible Halophyte Chenopodium quinoa Exposed to Salt Stress" Antioxidants 12, no. 5: 1060. https://doi.org/10.3390/antiox12051060
APA StyleSouid, A., Bellani, L., Tassi, E. L., Ben Hamed, K., Longo, V., & Giorgetti, L. (2023). Early Physiological, Cytological and Antioxidative Responses of the Edible Halophyte Chenopodium quinoa Exposed to Salt Stress. Antioxidants, 12(5), 1060. https://doi.org/10.3390/antiox12051060