

Vitamin D from UV-Irradiated Mushrooms as a Way for Vitamin D Supplementation: A Systematic Review on Classic and Nonclassic Effects in Human and Animal Models

 , , , , and
, , , , and
Abstract
1. Introduction
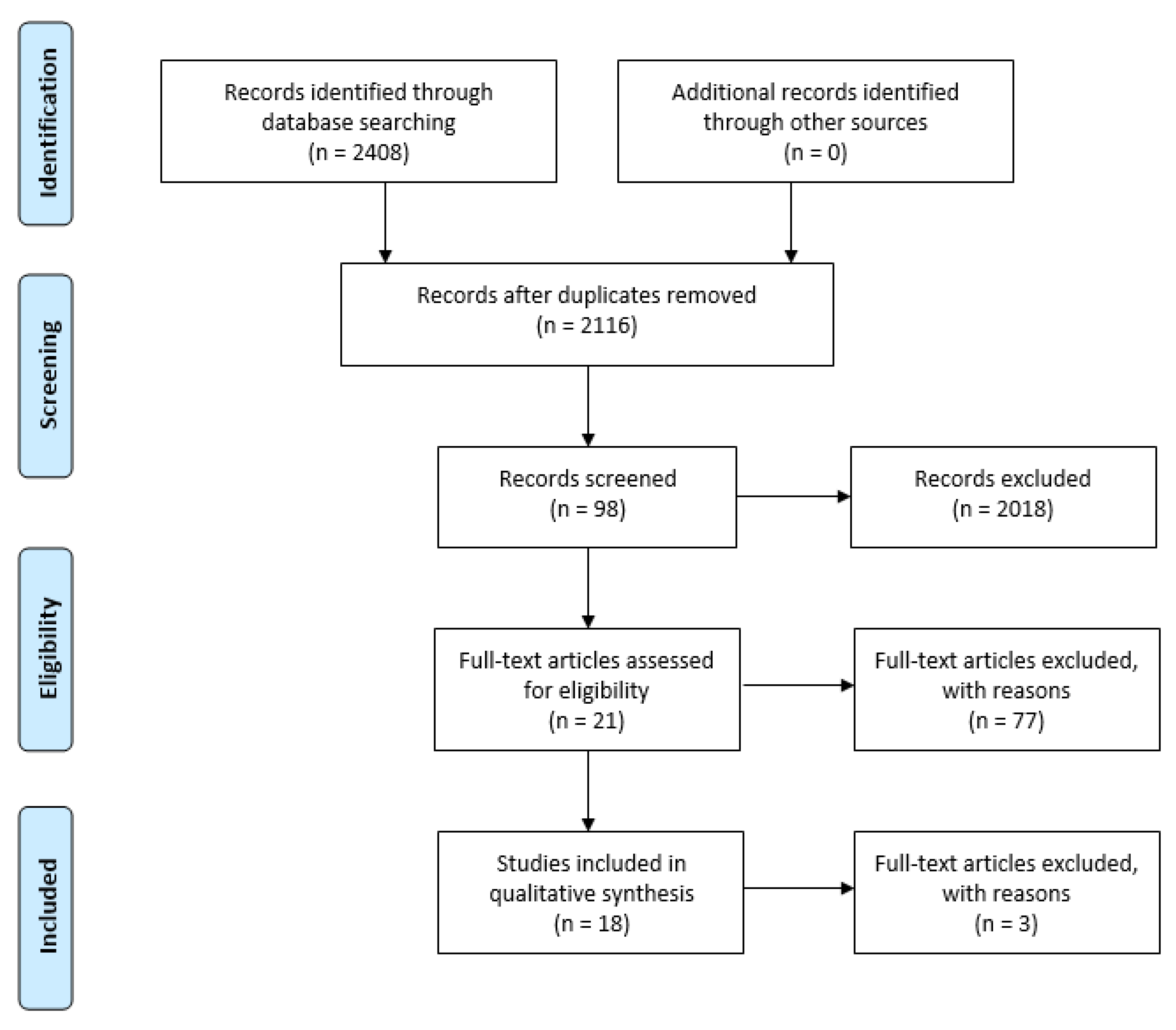
2. Materials and Methods
3. Results
3.1. Studies Conducted on Human Samples
3.2. Studies Conducted on Animal Samples
4. Discussion
4.1. Classical and Nonclassical Function of Vitamin D in Human Studies
4.2. Classical and Nonclassical Function of Vitamin D in Animal Studies
4.3. Comparison of Animal and Human Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cardwell, G.; Bornman, J.F.; James, A.P.; Black, L.J. A Review of Mushrooms as a Potential Source of Dietary Vitamin D. Nutrients 2018, 10, 1498. [Google Scholar] [CrossRef]
- Thu, Z.M.; Ko Myo, K.; Aung, H.T.; Clericuzio, M.; Armijos, C.; Vidari, G. Bioactive Phytochemical Constituents of Wild Edible Mushrooms from Southeast Asia. Molecules 2020, 25, 1972. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Xiao, J.; Xu, B. A Critical Review on Health Promoting Benefits of Edible Mushrooms through Gut Microbiota. Int. J. Mol. Sci. 2017, 18, 1934. [Google Scholar] [CrossRef]
- Strumińska-Parulska, D.; Falandysz, J. A Review of the Occurrence of Alpha-Emitting Radionuclides in Wild Mushrooms. Int. J. Environ. Res. Public Health 2020, 17, 8220. [Google Scholar] [CrossRef] [PubMed]
- Falandysz, J.; Borovička, J. Macro and trace mineral constituents and radionuclides in mushrooms: Health benefits and risks. Appl. Microbiol. Biotechnol. 2013, 97, 477–501. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.G.; Kerlan, V.; Desailloud, R. Non-Classical Effects of Vitamin D: Non-Bone Effects of Vitamin D. Ann. Endocrinol. 2021, 82, 43–51. [Google Scholar] [CrossRef]
- Helde-Frankling, M.; Björkhem-Bergman, L. Vitamin D in Pain Management. Int. J. Mol. Sci. 2017, 18, 2170. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N.; Shirvani, A.; Holick, M.F. Vitamin D for skeletal and non-skeletal health: What we should know. J. Clin. Orthop. Trauma 2019, 10, 1082–1093. [Google Scholar] [CrossRef]
- Borel, P.; Caillaud, D.; Cano, N.J. Vitamin D Bioavailability: State of the Art. Crit. Rev. Food Sci. Nutr. 2015, 55, 1193–1205. [Google Scholar] [CrossRef]
- Keegan, R.J.H.; Lu, Z.; Bogusz, J.M.; Williams, J.E.; Holick, M.F. Photobiology of Vitamin D in Mushrooms and Its Bioavailability in Humans. Dermatoendocrinol 2013, 5, 165–176. [Google Scholar] [CrossRef]
- Slominski, A.; Semak, I.; Wortsman, J.; Zjawiony, J.; Li, W.; Zbytek, B.; Tuckey, R.C. An alternative pathway of vitamin D2 metabolism. FEBS J. 2006, 273, 2891–2901. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Kim, T.-K.; Shehabi, H.Z.; Tang, E.K.; Benson, H.A.E.; Semak, I.; Lin, Z.; Yates, C.R.; Wang, J.; Li, W.; et al. In vivo production of novel vitamin D2 hydroxy-derivatives by human placentas, epidermal keratinocytes, Caco-2 colon cells and the adrenal gland. Mol. Cell Endocrinol. 2014, 383, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Li, W.; Kim, T.-K.; Semak, I.; Wang, J.; Zjawiony, J.K.; Tuckey, R.C. Novel activities of CYP11A1 and their potential physiological significance. J. Steroid Biochem. Mol. Biol. 2015, 151, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Kim, T.-K.; Janjetovic, Z.; Tuckey, R.C.; Bieniek, R.; Yue, J.; Li, W.; Chen, J.; Nguyen, M.N.; Tang, E.K.Y.; et al. 20-Hydroxyvitamin D2is a noncalcemic analog of vitamin D with potent antiproliferative and prodifferentiation activities in normal and malignant cells. Am. J. Physiol.-Cell Physiol. 2011, 300, C526–C541. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Semak, I.; Zjawiony, J.; Wortsman, J.; Gandy, M.N.; Li, J.; Zbytek, B.; Li, W.; Tuckey, R.C. Enzymatic Metabolism of Ergosterol by Cytochrome P450scc to Biologically Active 17α,24-Dihydroxyergosterol. Chem. Biol. 2005, 12, 931–939. [Google Scholar] [CrossRef]
- Slominski, R.; Raman, C.; Elmets, C.; Jetten, A.; Slominski, A.; Tuckey, R. The significance of CYP11A1 expression in skin physiology and pathology. Mol. Cell Endocrinol. 2021, 530, 111238. [Google Scholar] [CrossRef]
- Sánchez, C. Modern aspects of mushroom culture technology. Appl. Microbiol. Biotechnol. 2004, 64, 756–762. [Google Scholar] [CrossRef]
- Rai, R.D.; Yadav, M.C. Mushroom Biology and Biotechnology National Agricultural Technology Project (NATP) A-7 Enttiled “Development of Improved Strains of Mushroom and Domestication of Morchella Spp. Using Molecular Techniques” View Project OFF-FLAVOUR GENERATION SOYBEAN View Project. In Mushroom Biology and Biotechnology; Chapter: 3; MSI: Solan, India, 2007; pp. 41–61. [Google Scholar]
- Barney, D.L. Growing Mushrooms Commercially Risks and Opportunities; University of Idaho: Moscow, ID, USA, 2009. [Google Scholar]
- Sánchez, C. Cultivation of Pleurotus ostreatus and other edible mushrooms. Appl. Microbiol. Biotechnol. 2010, 85, 1321–1337. [Google Scholar] [CrossRef]
- Rühl, M. Ligninolytic Enzyme Activities Alternate with Mushroom Production during Industrial Cultivation of Pleurotus Ostreatus on Wheat Straw-Based Substrate Monocular Depth Estimation in Forest Environments View Project Explore the Terpenoid Diversity View Project. Curr. Trends Biotechnol. Pharm. 2008, 2, 478–492. [Google Scholar]
- Marrone, G.; Urciuoli, S.; Di Lauro, M.; Ruzzolini, J.; Ieri, F.; Vignolini, P.; Di Daniele, F.; Guerriero, C.; Nediani, C.; Di Daniele, N.; et al. Extra Virgin Olive Oil and Cardiovascular Protection in Chronic Kidney Disease. Nutrients 2022, 14, 4265. [Google Scholar] [CrossRef]
- Leung, M.F.; Cheung, P.C.K. Vitamins D and D2 in Cultivated Mushrooms under Ultraviolet Irradiation and Their Bioavailability in Humans: A Mini-Review. Int. J. Med. Mushrooms 2021, 23, 40390. [Google Scholar] [CrossRef]
- Xu, Z.; Meenu, M.; Xu, B. Effects of UV-C treatment and ultrafine-grinding on the biotransformation of ergosterol to vitamin D2, physiochemical properties, and antioxidant properties of shiitake and Jew’s ear. Food Chem. 2020, 309, 125738. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhang, M.; Mujumdar, A.S. UV induced conversion during drying of ergosterol to vitamin D in various mushrooms: Effect of different drying conditions. Trends Food Sci. Technol. 2020, 105, 200–210. [Google Scholar] [CrossRef]
- Salemi, S.; Saedisomeolia, A.; Azimi, F.; Zolfigol, S.; Mohajerani, E.; Mohammadi, M.; Yaseri, M. Optimizing the production of vitamin D in white button mushrooms (Agaricus bisporus) using ultraviolet radiation and measurement of its stability. LWT 2021, 137, 110401. [Google Scholar] [CrossRef]
- Heo, T.-Y.; Kim, Y.-N.; Park, I.B.; Lee, D.-U. Amplification of Vitamin D2 in the White Button Mushroom (Agaricus bisporus) by UV-B Irradiation and Jet-Milling for Its Potential Use as a Functional Ingredient. Foods 2020, 9, 1713. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Chen, W.; Li, X.; Yue, T.; Zhang, Z.; Feng, Z.; Li, C.; Bu, X.; Li, Q.X.; Hu, C.Y.; et al. Ultraviolet Irradiation Increased the Concentration of Vitamin D2 and Decreased the Concentration of Ergosterol in Shiitake Mushroom (Lentinus Edodes) and Oyster Mushroom (Pleurotus Ostreatus) Powder in Ethanol Suspension. ACS Omega 2020, 5, 7361–7368. [Google Scholar] [CrossRef]
- Hu, D.; Yang, X.; Hu, C.; Feng, Z.; Chen, W.; Shi, H. Comparison of Ergosterol and Vitamin D2 in Mushrooms Agaricus bisporus and Cordyceps militaris Using Ultraviolet Irradiation Directly on Dry Powder or in Ethanol Suspension. ACS Omega 2021, 6, 29506–29515. [Google Scholar] [CrossRef] [PubMed]
- Vieth, R. Vitamin D supplementation: Cholecalciferol, calcifediol, and calcitriol. Eur. J. Clin. Nutr. 2020, 74, 1493–1497. [Google Scholar] [CrossRef]
- Tripkovic, L.; Lambert, H.; Hart, K.; Smith, C.P.; Bucca, G.; Penson, S.; Chope, G.; Hyppönen, E.; Berry, J.; Vieth, R.; et al. Comparison of vitamin D2 and vitamin D3 supplementation in raising serum 25-hydroxyvitamin D status: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2012, 95, 1357–1364. [Google Scholar] [CrossRef]
- Bjelakovic, G.; Gluud, L.L.; Nikolova, D.; Whitfield, K.; Wetterslev, J.; Simonetti, R.G.; Bjelakovic, M.; Gluud, C. Vitamin D supplementation for prevention of mortality in adults. Cochrane Database Syst. Rev. 2014, 2014, CD007470. [Google Scholar] [CrossRef]
- Houghton, L.A.; Vieth, R. The case against ergocalciferol (vitamin D2) as a vitamin supplement. Am. J. Clin. Nutr. 2006, 84, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Kido, S.; Chosa, E.; Tanaka, R. The Effect of Six Dried and UV-C-Irradiated Mushrooms Powder on Lipid Oxidation and Vitamin D Contents of Fish Meat. Food Chem. 2023, 398, 133917. [Google Scholar] [CrossRef] [PubMed]
- Ao, T.; Kikuta, J.; Ishii, M. The Effects of Vitamin D on Immune System and Inflammatory Diseases. Biomolecules 2021, 11, 1624. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, R.M.; Ali, M.I.; Abdel-Salam, F.F. Nutritional and Quality Characteristics of Some Foods Fortified with Dried Mushroom Powder as a Source of Vitamin D. Int. J. Food Sci. 2022, 2022, 2792084. [Google Scholar] [CrossRef] [PubMed]
- Liberati, M.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Zajac, I.T.; Barnes, M.; Cavuoto, P.; Wittert, G.; Noakes, M. The Effects of Vitamin D-Enriched Mushrooms and Vitamin D3 on Cognitive Performance and Mood in Healthy Elderly Adults: A Randomised, Double-Blinded, Placebo-Controlled Trial. Nutrients 2020, 12, 3874. [Google Scholar] [CrossRef]
- Mehrotra, A.; Calvo, M.S.; Beelman, R.B.; Levy, E.; Siuty, J.; Kalaras, M.D.; Uribarri, J. Bioavailability of vitamin D2 from enriched mushrooms in prediabetic adults: A randomized controlled trial. Eur. J. Clin. Nutr. 2014, 68, 1154–1160. [Google Scholar] [CrossRef]
- Stephensen, C.B.; Zerofsky, M.; Burnett, D.J.; Lin, Y.-P.; Hammock, B.D.; Hall, L.M.; McHugh, T. Ergocalciferol from Mushrooms or Supplements Consumed with a Standard Meal Increases 25-Hydroxyergocalciferol but Decreases 25-Hydroxycholecalciferol in the Serum of Healthy Adults. J. Nutr. 2012, 142, 1246–1252. [Google Scholar] [CrossRef]
- Shanely, R.A.; Nieman, D.C.; Knab, A.M.; Gillitt, N.D.; Meaney, M.P.; Jin, F.; Sha, W.; Cialdella-Kam, L. Influence of vitamin D mushroom powder supplementation on exercise-induced muscle damage in vitamin D insufficient high school athletes. J. Sports Sci. 2014, 32, 670–679. [Google Scholar] [CrossRef]
- Stepien, M.; O’Mahony, L.; O’Sullivan, A.; Collier, J.; Fraser, W.D.; Gibney, M.J.; Nugent, A.P.; Brennan, L. Human and Clinical Nutrition: Effect of supplementation with vitamin D2-enhanced mushrooms on vitamin D status in healthy adults. J. Nutr. Sci. 2013, 2, e29. [Google Scholar] [CrossRef]
- Nieman, D.C.; Gillitt, N.D.; Andrew Shanely, R.; Dew, D.; Meaney, M.P.; Luo, B. Vitamin D2 Supplementation Amplifies Eccentric Exercise-Induced Muscle Damage in NASCAR Pit Crew Athletes. Nutrients 2013, 6, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.S.; Babu, U.S.; Garthoff, L.H.; Woods, T.O.; Dreher, M.; Hill, G.; Nagaraja, S. Vitamin D2 from light-exposed edible mushrooms is safe, bioavailable and effectively supports bone growth in rats. Osteoporos. Int. 2013, 24, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Jasinghe, V.J.; Perera, C.O.; Barlow, P.J. Vitamin D2 from Irradiated Mushrooms Significantly Increases Femur Bone Mineral Density in Rats. J. Toxicol. Environ. Health 2006, 69, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Babu, U.S.; Balan, K.V.; Garthoff, L.H.; Calvo, M.S. Vitamin D2 from UVB light exposed mushrooms modulates immune response to LPS in rats. Mol. Nutr. Food Res. 2014, 58, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Dowley, A.; Sweeney, T.; Conway, E.; Vigors, S.; Yadav, S.; Wilson, J.; Gabrielli, W.; O’Doherty, J.V. Effects of Dietary Supplementation with Mushroom or Vitamin D2-Enriched Mushroom Powders on Gastrointestinal Health Parameters in the Weaned Pig. Animals 2021, 11, 3603. [Google Scholar] [CrossRef]
- Malik, M.A.; Jan, Y.; Al-Keridis, L.A.; Haq, A.; Ahmad, J.; Adnan, M.; Alshammari, N.; Ashraf, S.A.; Panda, B.P. Effect of Vitamin-D-Enriched Edible Mushrooms on Vitamin D Status, Bone Health and Expression of CYP2R1, CYP27B1 and VDR Gene in Wistar Rats. J. Fungi 2022, 8, 864. [Google Scholar] [CrossRef]
- Bennett, L.; Kersaitis, C.; Macaulay, S.L.; Münch, G.; Niedermayer, G.; Nigro, J.; Payne, M.; Sheean, P.; Vallotton, P.; Zabaras, D.; et al. Vitamin D2-Enriched Button Mushroom (Agaricus bisporus) Improves Memory in Both Wild Type and APPswe/PS1dE9 Transgenic Mice. PLoS ONE 2013, 8, e76362. [Google Scholar] [CrossRef]
- Conway, E.; Sweeney, T.; Dowley, A.; Maher, S.; Rajauria, G.; Yadav, S.; Wilson, J.; Gabrielli, W.; O’Doherty, J.V. The effects of mushroom powder and vitamin D2 -enriched mushroom powder supplementation on the growth performance and health of newly weaned pigs. J. Anim. Physiol. Anim. Nutr. 2022, 106, 517–527. [Google Scholar] [CrossRef]
- Lee, G.-S.; Byun, H.-S.; Yoon, K.-H.; Lee, J.-S.; Choi, K.-C.; Jeung, E.-B. Dietary calcium and vitamin D2 supplementation with enhanced Lentinula edodes improves osteoporosis-like symptoms and induces duodenal and renal active calcium transport gene expression in mice. Eur. J. Nutr. 2009, 48, 75–83. [Google Scholar] [CrossRef]
- Won, D.J.; Seong, K.S.; Jang, C.H.; Lee, J.S.; Ko, J.A.; Bae, H.; Park, H.J. Effects of vitamin D2-fortified shiitake mushroom on bioavailability and bone structure. Biosci. Biotechnol. Biochem. 2019, 83, 942–951. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Yu, H.-T.; Kao, J.-P.; Yang, C.-C.; Chiang, S.-S.; Mishchuk, D.O.; Mau, J.-L.; Slupsky, C.M. Consumption of vitamin D2 enhanced mushrooms is associated with improved bone health. J. Nutr. Biochem. 2015, 26, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Drori, A.; Rotnemer-Golinkin, D.; Avni, S.; Danay, O.; Levanon, D.; Tam, J.; Zolotarev, L.; Ilan, Y. Attenuating the rate of total body fat accumulation and alleviating liver damage by oral administration of vitamin D-enriched edible mushrooms in a diet-induced obesity murine model is mediated by an anti-inflammatory paradigm shift. BMC Gastroenterol. 2017, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.K.; Kelly, A.K.; Rajauria, G.; Jakobsen, J.; Clarke, L.C.; Monahan, F.J.; Dowling, K.G.; Hull, G.; Galvin, K.; Cashman, K.D.; et al. The use of synthetic and natural vitamin D sources in pig diets to improve meat quality and vitamin D content. Meat Sci. 2018, 143, 60–68. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Technological and Environmental Factors | Comment | Reference |
|---|---|---|
| Type of effective irradiation | Increased effectiveness using UVB irradiation and sun exposure, rather than UVA and UVC irradiation. | [1,23,24,25,26,27,28,29,36] |
| Irradiation surface | The smaller the sample, the larger the exposure area and thus the greater the conversion of vitamin D2. Again, irradiating all sides of the cut allows a greater synthesis of vitamin D2 than simply exposing only one side. | [1,23,24,25] |
| Ideal reaction temperature | Between 25–29 °C. Data still insufficient and preliminary. | [1,23,24,25,36] |
| Exposure intensity | Higher efficacy for values between 1.14 and 1.36 W/m2 (values referring exclusively to irradiation with pulsed rays). There are no data correlating the conversion of ergosterols to ergocalciferol with the environmental UV index and/or factors such as latitude and time of day. Data still insufficient and preliminary. | [1,23,24,25] |
| Product type | Higher conversion efficiency in freeze-dried and heat-dried mushrooms than in fresh mushrooms. Data still insufficient and preliminary. | [1,23,25,36] |
| Sample storage | In fresh mushrooms, loss of about 23% vitamin D2 in mushrooms stored at 3–2 °C due to sample degradation, but no significant loss for storage at 4 °C. At room temperature, dried samples appear to have losses of about 50% after 18 months. Still insufficient and preliminary data. | [1,23,24,25] |
| Culinary transformations | Cooking induces a loss of vitamin D2, which differs depending on the type of cooking (greater in baking than in pan-frying). Data still insufficient and preliminary. | [1,23] |
| Subjects | Reaction Conditions and Sample Type | Vitamin D2 EM Supplementation and Species | Vitamin D Nonclassic Effects | Vitamin D Classic Effects | Reference |
|---|---|---|---|---|---|
| N = 436 (healthy, ≥60 years old, M-F) | 6 months freeze-dried, powder and capsuled, UVB Lamp | 600 IU/die, Agaricus bisporus | No benefit on cognitive functions and mood | Increase in 25(OH)D2. Decline in total 25(OHD). Such decline was observed to be less negative and slower in the D2 arm (EM) than controls. | [38] |
| N = 36 (pre-diabetic, BMI < 25, vitamin D-deficient adults, 49 ± 12 years old, M-F) | 4 months fresh sliced cooked mushrooms, UVB Lamp | arm 1: 600 IU/die arm 2: 4000 IU/die Agaricus bisporus | No positive or significant results on metabolic syndrome markers | Modest, but significant increase in serum 25(OH)D2 over time. The amount of 25(OH)D3 and total 25(OH)D remained unchanged. | [39] |
| N = 40 (healthy adults, 20–50 years old, M-F) | 6 weeks fresh sliced cooked mushrooms cooked mushrooms, UVB Lamp | 25 µg ergocalciferol/die arm 1: 8.8 µg/die arm 2: 17.1 µg/die Agaricus bisporus | Not studied by the authors | Increase in serum 25(OH)D2 in both arms, which was correlated to the treatment. No influence of any arm treatment on total 25(OH)D while 25(OH)D3 levels declined. | [40] |
| N = 33 (athletes students with serum 25(OH)D less than 30 ng/mL, 16.2 ± 0.19 years old) | 6 weeks powder and capsuled, UVB Lamp | 600 IU/die Agaricus bisporus | No positive results muscle system/function, exercise-induced muscle damage or DOMS | Increase in either 25(OH)D2 or total 25(OH)D. Levels of 25(OH)D3 decreased. | [41] |
| N = 90 (healthy adults 40–65 years old, M-F) | 4 weeks Freeze-dried, powder, UVB Lamp | 600 IU/die Agaricus bisporus | Significant decrease in Plasminogen activator inhibitor-1 (PAI-1) | No significant changes in total 25(OH)D and 25(OH)D3 values. Increase in 25(OH)D2 levels. | [42] |
| N = 28 (healthy athlete professional pilots, age not declared, M) | 6 weeks powdered, hot dried, UVB Lamp | 3800 IU/die Agaricus bisporus | No influence on muscle system/function, exercise-induced muscle damage or DOMS and negative effects on their markers such as CK, LDH, serum myoglobin and DOMS as well as amplified exercise-induced muscle damage (EIMD) | No significant change in total 25(OH)D values, but significant increase in serum 25(OH)D2 levels and a significant decrease in serum 25(OH)D3 levels. | [43] |
| Subjects | Reaction Conditions and Sample Type | Vitamin D2 EM Supplementation and Species | Vitamin D Nonclassic Effects | Vitamin D Classic Effects | Reference |
|---|---|---|---|---|---|
| 300 rats 3 weeks ol—F | 10 weeks powdered, freeze-dried, UVB Lamp | arm 1: 300 IU/day arm 2: 600 IU/day Agaricus bisporus | Vitamin D2 from mushrooms was bioavailable and effective in suppressing PTH levels. The mean PTH level in the group of rats fed with the 5.0% of unexposed mushroom diet was also significantly lower (p < 0.002) than in the vitamin D-deficient fed control group. Rats fed the 5.0% UVB mushroom diet had a mean 25(OH)D plasma level of 159 ± 29 ng/mL, while the control group had a mean 25(OH)Dtot final level of 32 ± 11 ng/mL. The tested treatment, appeared to stimulate bone formation in all of the experimental group, inducing significantly longer femurs, p < 0.001), and a positive effect on cortical bone (The treatment also had a positive effect on cortical bone (higher mean midshaft cortical thickness and pMOI, p < 0.01). | [44] | |
| 30 rats (age not declared—M) | 4 weeks powdered, hot dried, UVB Lamp | 1 mg/die Lentinula edodes | The serum 25OHD concentration of the active group was 129 ± 42 (SD 22 ± 00) nmol/L in contrast to the control group (6 ± 06) (SD 1 ± 09) nmol/L. The serum calcium level of the active group was significantly lower compared to the controls. Significantly higher BMD and femur lengths in the active group. Decreased serum PTH levels, increased serum ionized Ca levels and an age-related decrease in duodenal Ca absorption have been previously reported. | [45] | |
| 100 rats (3 wk old) | 10 weeks Powdered, freeze-dried, UVB Lamp | arm 1: 300 IU/die arm 2: 600 IU/die Agaricus bisporus | Plasma TNF-a and MIP2 were significantly lower in rats fed 2.5 and 5% UVB-irradiated mushrooms compared with controls; IL-1b was significantly higher in rats fed 5% non-irradiated and 2.5% UVB-irradiated mushrooms. NK-cell activity was improved in the 5% UVB-exposed mushroom group compared to controls. UVB-exposed mushrooms in the diet caused a significant reduction in chemokines and cytokines in response to LPS stimulation. | Rats fed 5.0% UVB-exposed mushroom diet (600 IU) had a mean plasma 25(OH)Dtot (155.4 ± 12.8 ng/mL) in respect of the control. | [46] |
| 192 pigs (28 days old, 96 M and 96 F) | 45 days dried powder, UVB lamp | 100 ug/kg/feed Agaricus bisporus | Concerning gastrointestinal morphology, treatment led to a significant improvement of VH (villus height). Pigs supplemented with MPD2 had increased total colonic VFA’s compared with all other groups (p < 0.05). Treatment also increased expression of SLC15A1, (p < 0.05) and FABP2, (p < 0.05) in the duodenum compared with the control group. In the duodenum, treatment caused a reduction in the expression of chemokine CXCL8, (p < 0.05) compared with the ZnO group and a reduction in the expression of cytokine gene IL6, (p < 0.05) compared to the control group. In the ileum, treatment led to an increase in the expression of cytokine gene IL10, (p < 0.05) compared with the control and ZnO group. | Increased expression of vitamin D receptor VDR, (p < 0.05) in the duodenum compared to the control and ZnO group. | [47] |
| 36 rats (4–6 weeks old, M-F) | 4 weeks powdered, hot dried, UVB Lamp | 30 IU/die Lentinula edodes Pleurotus ostreatus Agaricus bisporus | Treatment caused a significant increase in 25(OH)D. A significant increase (p < 0.05) was seen in calcium and phosphorus levels, while a significant decrease in alkaline phosphatase and PTH levels was seen in all treated groups. The results showed a significant increase in trabecular separation and a significant decrease in osteoid area in the selected region of interest in relation to the control group (p < 0.05). | [48] | |
| 21 wild-type (B6C3) and 25 transgenic (APPSwe/ PS1dE9) mice (2 months old, gender not declared) | 7 months powdered, freeze-dried, UV-C Lamp | 54 IU/Kg/die Agaricus bisporus | Treatment resulted in significantly (p < 0.05) higher number of IL-10-positive neurons in the cortex and a significantly (p < 0.01) larger area of neurons was IL-10 positive. Immunolocalization of IL-1β in the irradiation cortex or hippocampus of 9-month-old mice showed no difference between chow type or genotype, but a significant main effect of chow on the total area of neurons in the cortex. The absolute sensitivity of IL-1β staining and the size of the total neuronal area appeared to be lower than that of IL-10, based on the comparison of the stained areas. In conclusion, VDM-fed wild-type and AD transgenic mice showed improved learning and memory performance, significantly reduced amyloid plaque load and glial fibrillary acidic protein, and increased brain interleukin-10 concentration. The results suggest that VDM may be a dietary source of vitamin D2 and other bioactives to prevent memory impairment in dementia. | Treatment led to an increase in vitamin D2. | [49] |
| 192 pigs (age not declared, M-F) | 35 days dried powder, UVB Lamp | 100 ug/kg/feed Agaricus bisporus | No effect of treatment on the number of total bacteria. Pigs in the control group had higher concentrations of total fecal VFA than the treated group. With regard to the coefficient of apparent total tract digestibility (CATTD), the authors found no differences between the groups. | [50] | |
| 55 mice (3 weeks old, M) | 4 weeks Freeze-dried, powder, UVB Lamp | 1 ug/die Active treatment: Ca++ and Vitamin D2 EM Lentinula edodes | Beneficial effects of Ca++-EM treatment on serum calcium levels, mRNA levels of active calcium transport genes (duodenal CABP9K, TRPV6 and renal CABP9K, TRPV5,6) and femur density and length bone histology. | [51] | |
| 48 rats (subjected to sham operation or bilateral ovariectomy, when they were 5 weeks old, F) | 6 weeks Powder, Hot dried, UVB lamp | 750 ug/KgDiet 30 g powder/die Lentinula edodes | Comparison of 25(OH)D2 levels between the sham UV(O) and OVX-UV (O) groups showed that its level decreased by 70% in the absence of estrogen, suggesting that the presence of ovaries is highly associated with vitamin D2 bioavailability. The absence of estrogen had negative effects on trabecular bone structure and the bioavailability of vitamin D and calcium. Although there was a decrease in calcium levels, serum 25(OH)Dtot and other parameters linked to trabecular bone structures including BMD, vitamin D2- EM. These results may help delay bone loss which can be accelerated by the absence of estrogens after menopause. | [52] | |
| 32 mice (some of them subjected to sham operation or bilateral ovariectomy,7 weeks old, F) | 23 weeks Powdered, Freeze-dried, UVC Lamp | 5 ug/die Pleurotus eryngii | Pulsed enhanced vitamin D2 EM can maintain bone health, decrease the activity of bone resorption markers (osteocalcin, PYD, NTX1) and increase bone health related metabolites (osteocalcin, taurine, cretainin, emaic lactate, arginine) in OVX mice through its action as a mushroom (polyphenols and fiber) and its vitamin D2 content. | [53] | |
| Four groups of mice (n = 5/6) mice per group. Concanavalin A induced immune liver damage) | 25 weeks | 1 IU of vitamin D twice daily for 3 days Lentinula edodes | Beneficial effects of EM on liver damage and insulin resistance in a mouse model of NAFLD. A synergistic effect on body fat accumulation was observed. Given the high safety profile of these extracts, the data support their potential use in early-stage patients NASH. | [54] | |
| 120 pigs (60 M ad 60 F, age not declared) | DC | 55 days Freeze-dried, powder, UVB Lamp | 50 μg of vitamin D₂/kg/feed Study focused on the organoleptic quality of the meat (not reported but one from Monaghan Mushrooms that sell only commercial mushrooms) | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rondanelli, M.; Moroni, A.; Zese, M.; Gasparri, C.; Riva, A.; Petrangolini, G.; Perna, S.; Mazzola, G. Vitamin D from UV-Irradiated Mushrooms as a Way for Vitamin D Supplementation: A Systematic Review on Classic and Nonclassic Effects in Human and Animal Models. Antioxidants 2023, 12, 736. https://doi.org/10.3390/antiox12030736
Rondanelli M, Moroni A, Zese M, Gasparri C, Riva A, Petrangolini G, Perna S, Mazzola G. Vitamin D from UV-Irradiated Mushrooms as a Way for Vitamin D Supplementation: A Systematic Review on Classic and Nonclassic Effects in Human and Animal Models. Antioxidants. 2023; 12(3):736. https://doi.org/10.3390/antiox12030736
Chicago/Turabian StyleRondanelli, Mariangela, Alessia Moroni, Marco Zese, Clara Gasparri, Antonella Riva, Giovanna Petrangolini, Simone Perna, and Giuseppe Mazzola. 2023. "Vitamin D from UV-Irradiated Mushrooms as a Way for Vitamin D Supplementation: A Systematic Review on Classic and Nonclassic Effects in Human and Animal Models" Antioxidants 12, no. 3: 736. https://doi.org/10.3390/antiox12030736
APA StyleRondanelli, M., Moroni, A., Zese, M., Gasparri, C., Riva, A., Petrangolini, G., Perna, S., & Mazzola, G. (2023). Vitamin D from UV-Irradiated Mushrooms as a Way for Vitamin D Supplementation: A Systematic Review on Classic and Nonclassic Effects in Human and Animal Models. Antioxidants, 12(3), 736. https://doi.org/10.3390/antiox12030736