
Promotive Role of 5-Aminolevulinic Acid or Salicylic Acid Combined with Citric Acid on Sunflower Growth by Regulating Manganese Absorption

 ,
,  ,
, 
Abstract
1. Introduction
2. Experimental Section
2.1. Plant Materials and Growth Conditions
2.2. Plant Biomass, Chlorophyll Content, Malondialdehyde (MDA) and Reactive Oxygen Species (ROS) Analysis
2.3. Antioxidant and Non-Antioxidant Enzyme Activities
2.4. Heavy Metal Analysis
2.5. Transmission Electron Microscopy
2.6. Gene Expression Analysis
2.7. Statistical Analysis
3. Results
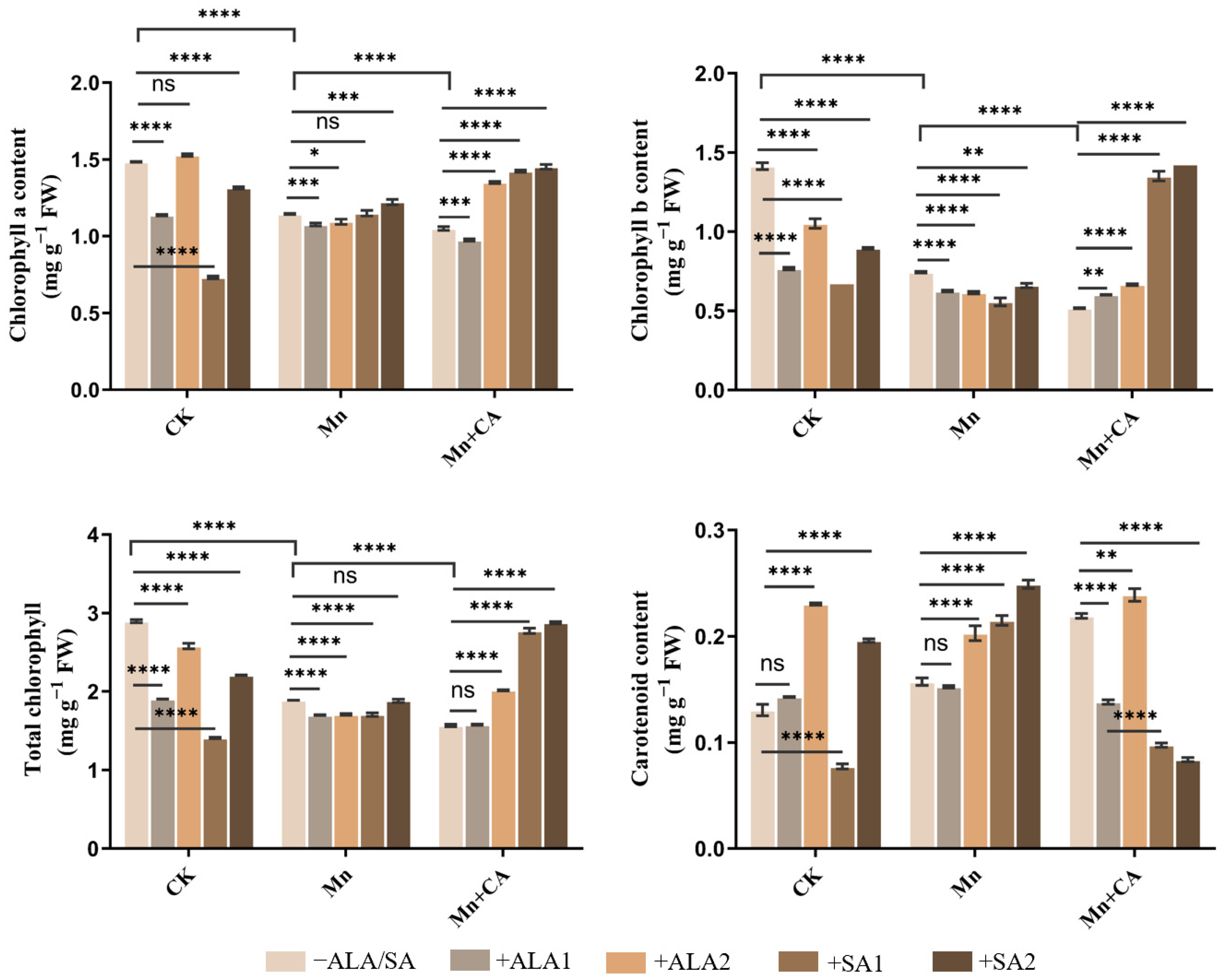
3.1. Plant Growth, Leaf Chlorophyll and Carotenoid Contents
3.2. Reactive Oxygen Species (ROS) and Lipid Peroxidation (MDA)
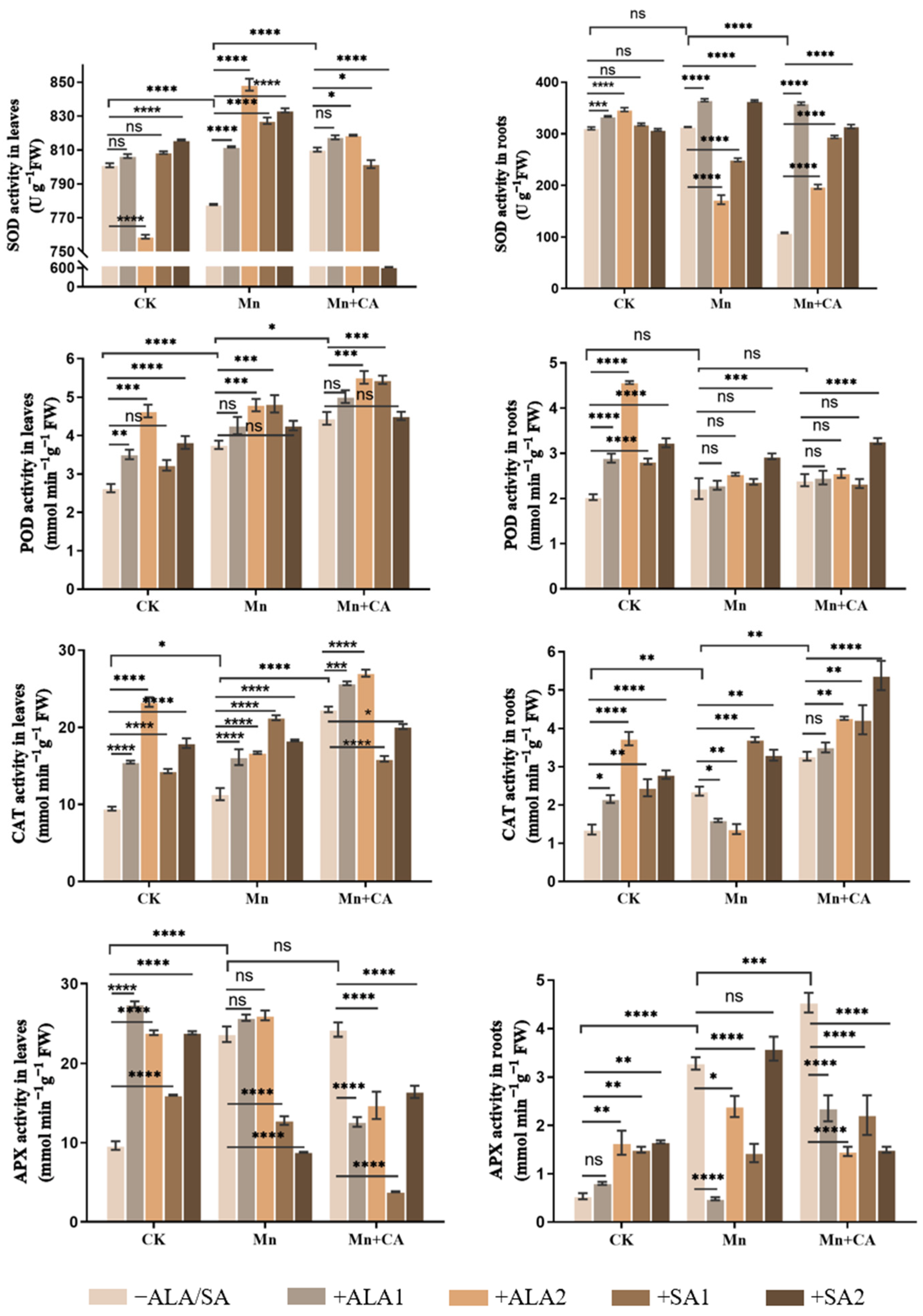
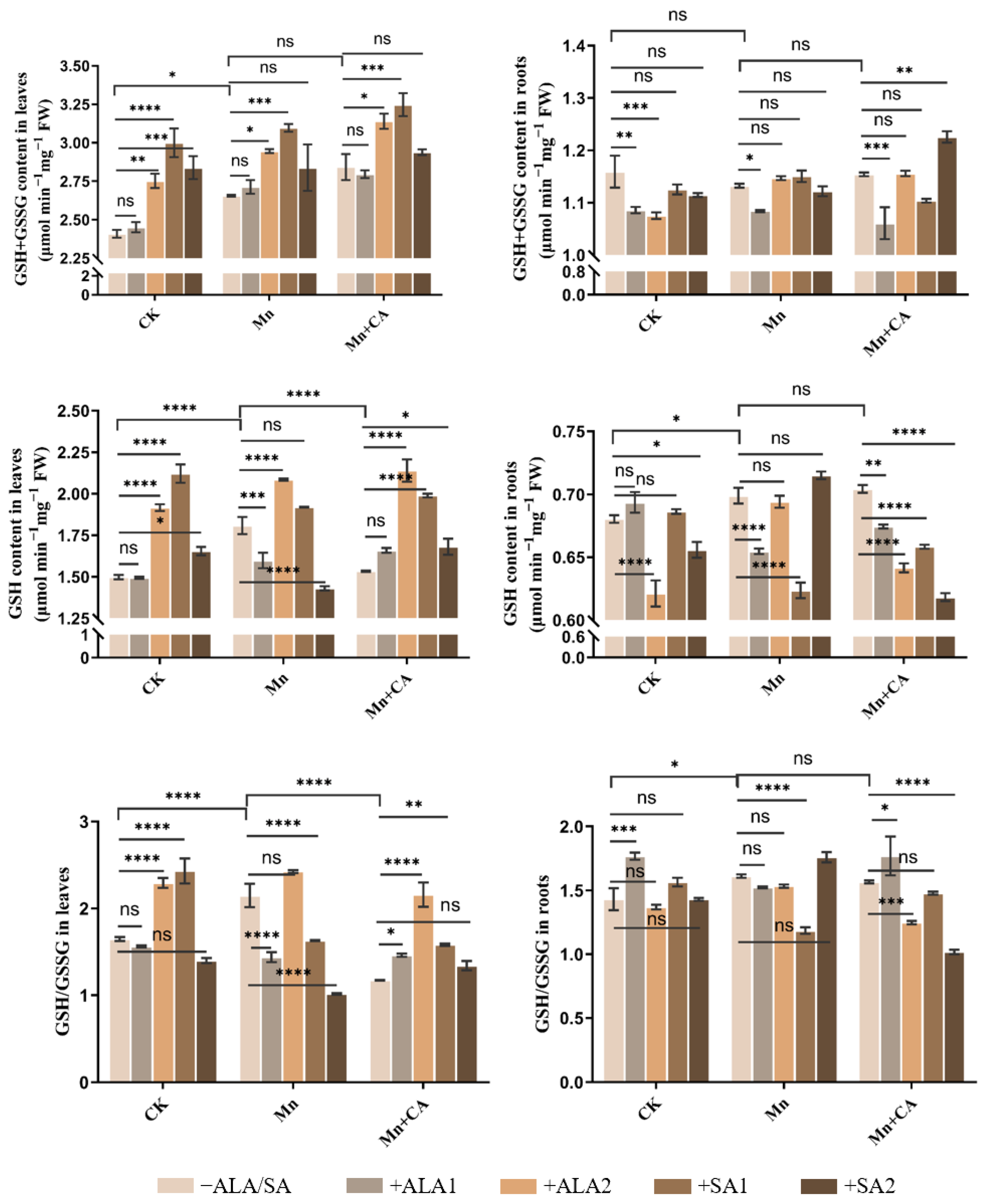
3.3. Antioxidant and Non-Antioxidant Enzyme Activities
3.4. Uptake and Translocation of Mn
3.5. Uptake of Other Elements
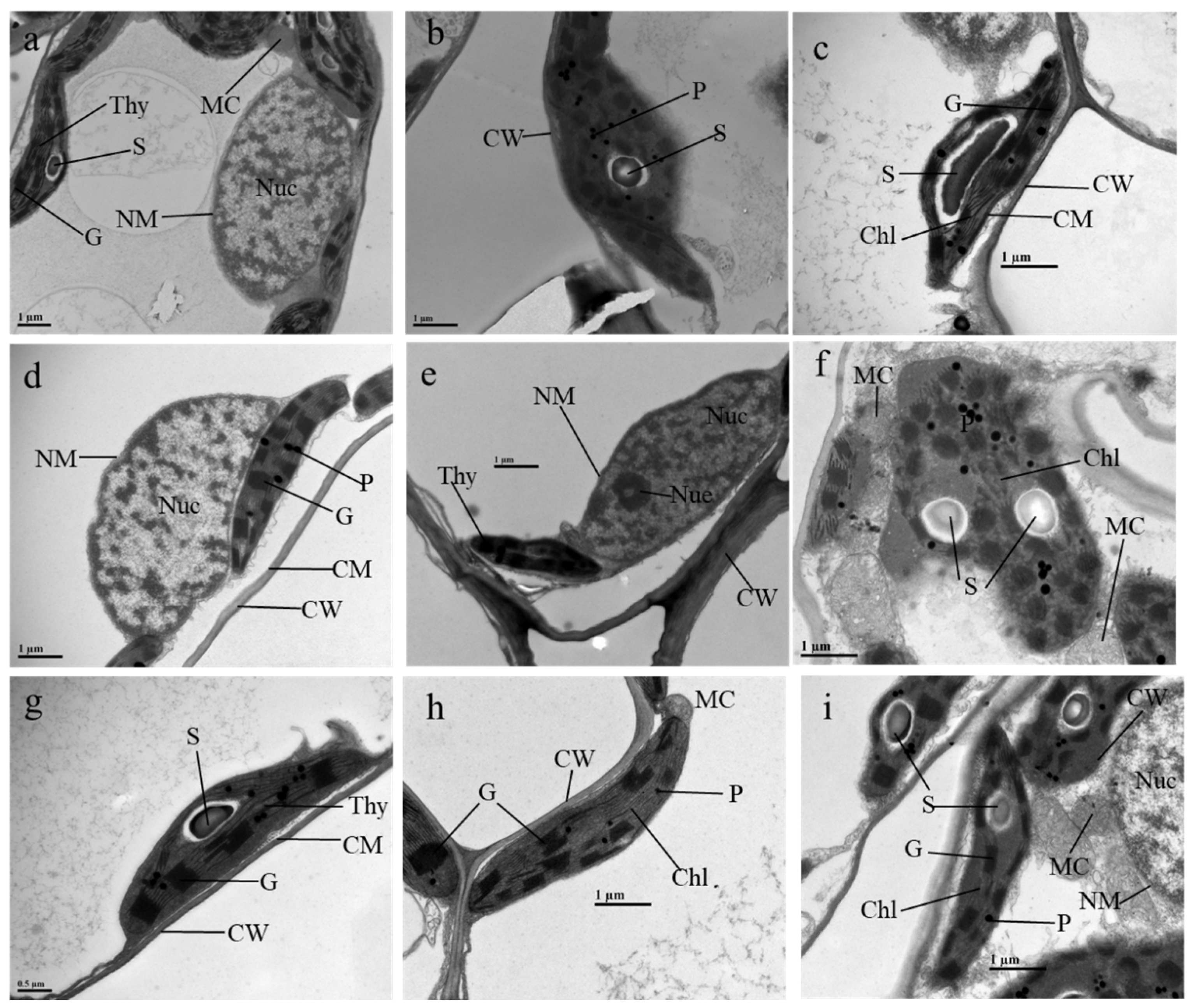
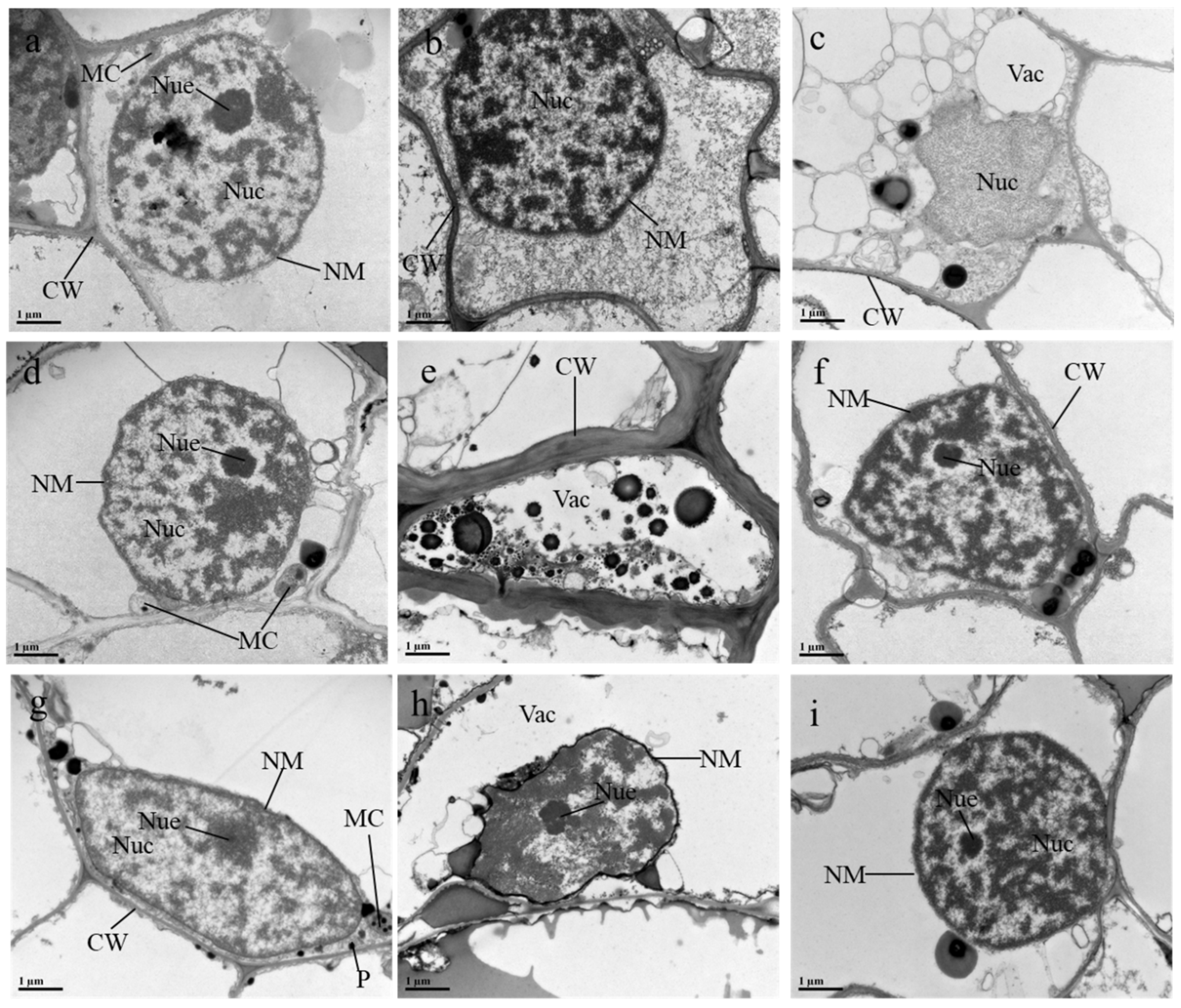
3.6. Effects of ALA/SA on Cellular Organelles of Mn- and CA-Treated Plants
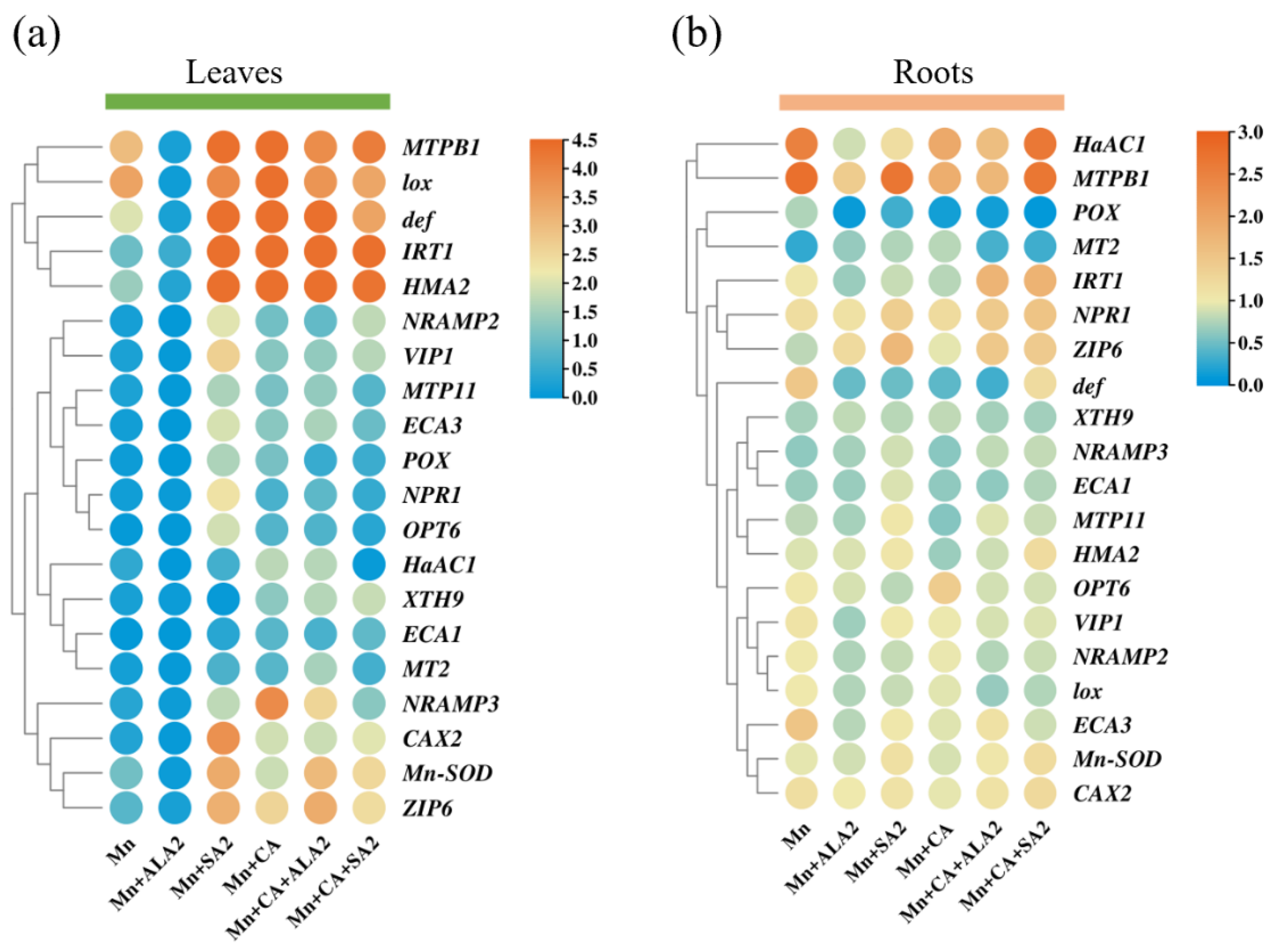
3.7. Effects of ALA/SA on Gene Expression under Mn and CA Treatments
3.8. Correlations among Different Parameters
4. Discussion
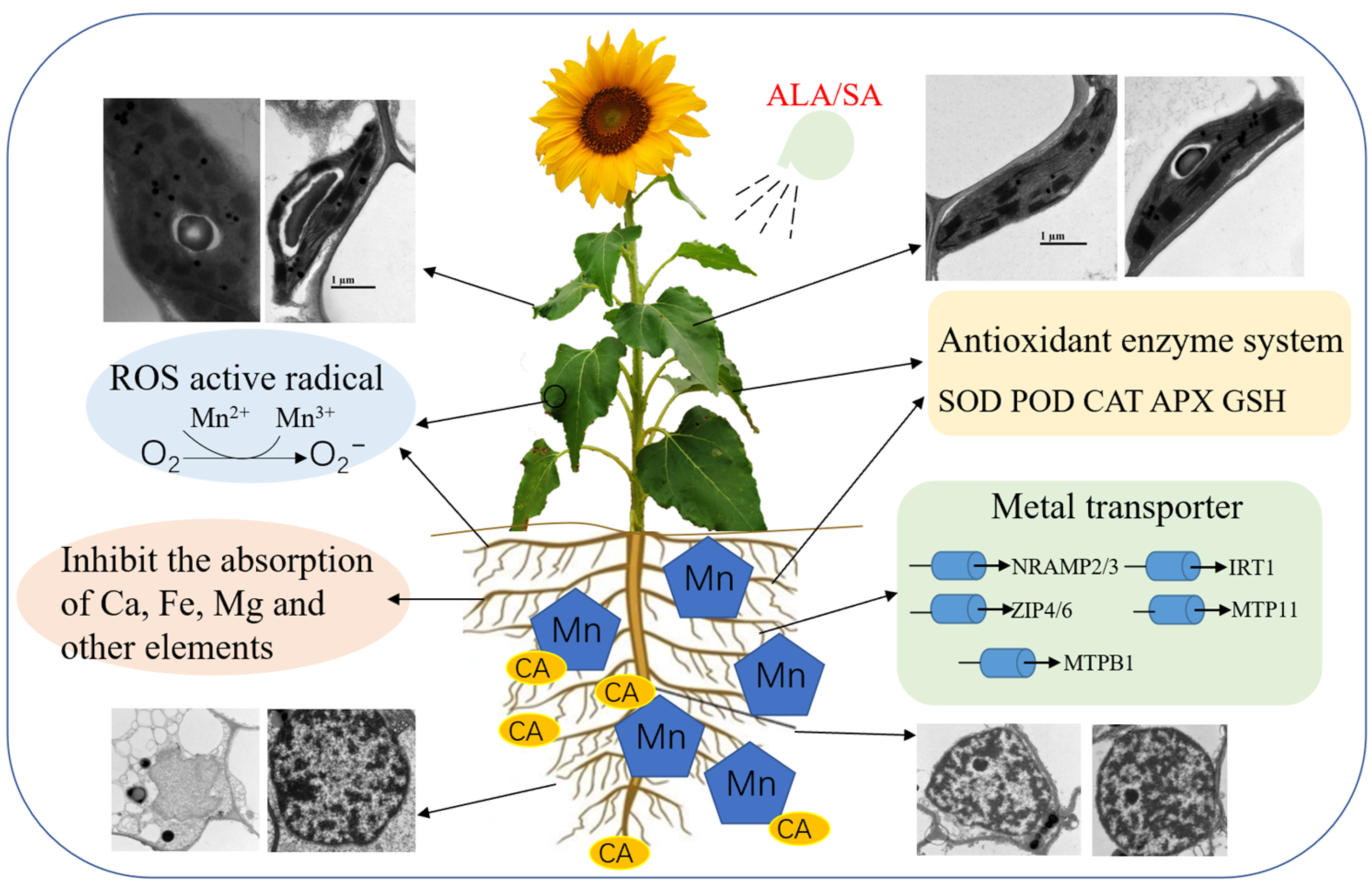
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Millaleo, R.; Reyes-Díaz, M.; Alberdi, M.; Ivanov, A.G.; Krol, M.; Hüner, N.P.A. Excess manganese differentially inhibits photosystem I versus II in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, C.; Liu, C.; Fu, A.; Luan, S. A Golgi-localized manganese transporter functions in pollen tube tip growth to control male fertility in Arabidopsis. Plant Commun. 2021, 2, 100178. [Google Scholar] [CrossRef] [PubMed]
- Millaleo, R.; Reyes-Diaz, M.; Ivanov, A.G.; Mora, M.L.; Alberdi, M. Manganese as essential and toxic element for plants: Transport, accumulation and resistance mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 470–481. [Google Scholar] [CrossRef]
- Gao, H.L.; Xie, W.X.; Yang, C.H.; Xu, J.Y.; Li, J.J.; Wang, H.; Chen, X.; Huang, C.F. NRAMP2, a trans-Golgi network-localized manganese transporter, is required for Arabidopsis root growth under manganese deficiency. New Phytol. 2018, 217, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Blamey, F.P.C.; Mckenna, B.A.; Li, C.; Cheng, M.M.; Tang, C.X.; Jiang, H.B.; Howard, D.L.; Paterson, D.J.; Kappen, P.; Wang, P.; et al. Manganese distribution and speciation help to explain the effects of silicate and phosphate on manganese toxicity in four crop species. New Phytol. 2018, 217, 1146–1160. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, X.Q.; Hu, Z.M.; Shao, J.F.; Che, J.; Chen, R.F.; Dong, X.Y.; Shen, R.F. Aluminium alleviates manganese toxicity to rice by decreasing root symplastic Mn uptake and reducing availability to shoots of Mn stored in roots. Ann. Bot. 2015, 116, 237–246. [Google Scholar] [CrossRef]
- Najeeb, U.; Xu, L.; Ali, S.; Jilani, G.; Gong, H.J.; Shen, W.Q.; Zhou, W.J. Citric acid enhances the phytoextraction of manganese and plant growth by alleviating the ultrastructural damages in Juncus effusus L. J. Hazard. Mater. 2009, 170, 1156–1163. [Google Scholar] [CrossRef]
- McGrath, S.P.; Zhao, F.J.; Lombi, E. Phytoremediation of metals, metalloids, and radionuclides. Adv. Agron. 2002, 75, 1–56. [Google Scholar] [CrossRef]
- Xu, L.; Li, J.J.; Najeeb, U.; Li, X.; Pan, J.M.; Huang, Q.; Zhou, W.J.; Liang, Z.S. Synergistic effects of EDDS and ALA on phytoextraction of cadmium as revealed by biochemical and ultrastructural changes in sunflower (Helianthus annuus L.) tissues. J. Hazard. Mater. 2021, 407, 124764. [Google Scholar] [CrossRef]
- Ali, B.; Tao, Q.J.; Zhou, Y.F.; Gill, R.A.; Ali, S.; Rafiq, M.T.; Xu, L.; Zhou, W.J. 5-aminolevolinic acid mitigates the cadmium-induced changes in Brassica napus as revealed by the biochemical and ultra-structural evaluation of roots. Ecotox. Environ. Saf. 2013, 92, 271–280. [Google Scholar] [CrossRef]
- Ali, B.; Wang, B.; Ali, S.; Ghani, M.A.; Hayat, M.T.; Yang, C.; Xu, L.; Zhou, W.J. 5-Aminolevulinic acid ameliorates the growth, photosynthetic gas exchange capacity, and ultrastructural changes under cadmium stress in Brassica napus L. J. Plant Growth Regul. 2013, 32, 604–614. [Google Scholar] [CrossRef]
- Ali, B.; Xu, X.; Gill, R.A.; Yang, S.; Ali, S.; Tahir, M.; Zhou, W.J. Promotive role of 5-aminolevulinic acid on mineral nutrients and antioxidative defense system under lead toxicity in Brassica napus. Ind. Crop. Prod. 2014, 52, 617–626. [Google Scholar] [CrossRef]
- Wang, J.; Lv, M.; Islam, F.; Gill, R.A.; Yang, C.; Ali, B.; Yan, G.J.; Zhou, W.J. Salicylic acid mediates antioxidant defense system and ABA pathway related gene expression in Oryza sativa against quinclorac toxicity. Ecotox. Environ. Saf. 2016, 133, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Shi, Y.; Long, M.J.; Islam, F.; Yang, C.; Yang, S.; He, Y.; Zhou, W.J. Estimation of quinclorac toxicity alleviated by salicylic acid in rice seedlings based on visible/near-infrared hyperspectral imaging. Plant Methods 2020, 16, 30. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Xu, S.; Yang, S.J.; Chen, Y.Y. Salicylic acid alleviates cadmium-induced inhibition of growth and photosynthesis through upregulating antioxidant defense system in two melon cultivars (Cucumis melo L.). Protoplasma 2015, 252, 911–924. [Google Scholar] [CrossRef] [PubMed]
- Nazar, R.; Umar, S.; Khan, N.A. Exogenous salicylic acid improves photosynthesis and growth through increase in ascorbate-glutathione metabolism and S assimilation in mustard under salt stress. Plant Signal. Behav. 2015, 10, e1003751. [Google Scholar] [CrossRef]
- Fayez, K.A.; Bazaid, S.A. Improving drought and salinity tolerance in barley by application of salicylic acid and potassium nitrate. J. Saudi Soc. Agric. Sci. 2014, 13, 45–55. [Google Scholar] [CrossRef]
- Khan, M.I.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Mahmood, Q.; Islam, E.; Jin, X.F.; Li, T.Q.; Yang, X.E.; Liu, D. The effect of EDDS addition on the phytoextraction efficiency from Pb contaminated soil by Sedum alfredii Hance. J. Hazard. Mater. 2009, 168, 530–535. [Google Scholar] [CrossRef]
- Najeeb, U.; Jilani, G.; Gill, S.A.; Sarwar, M.; Xu, L.; Zhou, W.J. Insights into cadmium induced physiological and ultra-structural disorders in Juncus effusus L. and its remediation through exogenous citric acid. J. Hazard. Mater. 2011, 186, 565–574. [Google Scholar] [CrossRef]
- Li, J.J.; Liu, A.K.; Najeeb, U.; Zhou, W.J.; Liu, H.; Yan, G.J.; Gill, A.R.; Yun, X.P.; Bai, Q.J.; Xu, L. Genome-wide investigation and expression analysis of membrane-bound fatty acid desaturase genes under different biotic and abiotic stresses in sunflower (Helianthus annuus L.). Int. J. Biol. Macromol. 2021, 175, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.Y.; Wang, D. Phytoremediation of uranium and cadmium contaminated soils by sunflower (Helianthus annuus L.) enhanced with biodegradable chelating agents. J. Clean. Prod. 2020, 263, 121491. [Google Scholar] [CrossRef]
- Luo, C.L.; Shen, Z.G.; Li, X.D. Enhanced phytoextraction of Cu, Pb, Zn and Cd with EDTA and EDDS. Chemosphere 2005, 59, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.; Rajguru, A.B.; Dudhe, M.Y.; Mathur, J. Efficacy of lead (Pb) phytoextraction of five varieties of Helianthus annuus L. from contaminated soil. Environ. Technol. Inno. 2020, 18, 100718. [Google Scholar] [CrossRef]
- Chen, L.; Hu, W.F.; Long, C.; Wang, D. Exogenous plant growth regulator alleviate the adverse effects of U and Cd stress in sunflower (Helianthus annuus L.) and improve the efficacy of U and Cd remediation. Chemosphere 2021, 262, 127809. [Google Scholar] [CrossRef] [PubMed]
- Benavides, B.J.; Drohan, P.J.; Spargo, J.T.; Maximova, S.N.; Guiltinan, M.J.; Miller, D.A. Cadmium phytoextraction by Helianthus annuus (sunflower), Brassica napus cv Wichita (rapeseed), and Chyrsopogon zizanioides (vetiver). Chemosphere 2021, 265, 129086. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. BBA-Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Zhang, W.F.; Zhang, F.; Raziuddin, R.; Gong, H.J.; Yang, Z.M.; Lu, L.; Ye, Q.F.; Zhou, W.J. Effects of 5-aminolevulinic acid on oilseed rape seedling growth under herbicide toxicity stress. J. Plant Growth Regul. 2008, 27, 159–169. [Google Scholar] [CrossRef]
- Zhou, W.J.; Leul, M. Uniconazole-induced alleviation of freezing injury in relation to changes in hormonal balance, enzyme activities and lipid peroxidation in winter rape. Plant Growth Regul. 1998, 21, 41–47. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen-peroxide is scavenged by ascorbate-specific peroxidase in spinach-chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic-acid in spinach (Spinacia oleracea) chloroplasts, the effect of hydrogen-peroxide and of paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Yang, C.; Liu, H.; Cao, M.T.; Yan, G.J.; Si, P.; Zhou, W.J.; Xu, L. 5-Aminolevolinic acid enhances sunflower resistance to Orobanche cumana (Broomrape). Ind. Crop. Prod. 2019, 140, 111467. [Google Scholar] [CrossRef]
- Xing, B.C.; Yang, D.F.; Guo, W.L.; Liang, Z.S.; Yan, X.J.; Zhu, Y.H.; Yan, L. Ag+ as a more effective elicitor for production of tanshinones than phenolic acids in Salvia miltiorrhiza hairy roots. Molecules. 2015, 20, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Z.; Li, D.Y.; Yang, D.F.; Xue, Z.Y.; Li, J.; Xing, B.C.; Yan, K.J.; Han, R.L.; Liang, Z.S. SmKFB5 protein regulates phenolic acid biosynthesis by controlling the degradation of phenylalanine ammonia-lyase in Salvia miltiorrhiza. J. Exp. Bot. 2021, 72, 4915–4929. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Li, X.; Han, P.; Liu, H.; Gong, J.C.; Zhou, W.J.; Shi, B.X.; Liu, A.K.; Xu, L. Genome-wide investigation of bHLH genes and expression analysis under different biotic and abiotic stresses in Helianthus annuus L. Int. J. Biol. Macromol. 2021, 189, 72–83. [Google Scholar] [CrossRef]
- Touria, E.J.; Douglas, A.S. Manganese toxicity in plants. J. Plant Nutr. 1998, 21, 353–386. [Google Scholar] [CrossRef]
- Xiong, J.; Yang, Y.J.; Fu, G.F.; Tao, L.X. Novel roles of hydrogen peroxide (H2O2) in regulating pectin synthesis and demethylesterification in the cell wall of rice (Oryza sativa) root tips. New Phytol. 2015, 206, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Youssef, T.; Awad, M.A. Mechanisms of enhancing photosynthetic gas exchange in date palm seedlings (Phoenix dactylifera L.) under salinity stress by a 5-aminolevulinic acid-based fertilizer. J. Plant Growth Regul. 2008, 27, 1–9. [Google Scholar] [CrossRef]
- Lin, W.D.; Li, Y.L.; Lu, Q.W.; Lu, H.F.; Li, J.M. Combined analysis of the metabolome and transcriptome identified candidate genes involved in phenolic acid biosynthesis in the leaves of Cyclocarya paliurus. Int. J. Mol. Sci. 2020, 21, 1337. [Google Scholar] [CrossRef]
- Tsunemitsu, Y.; Genga, M.; Okada, T.; Yamaji, N.; Ma, J.F.; Miyazaki, A.; Kato, S.; Iwasaki, K.; Ueno, D. A member of cation diffusion facilitator family, MTP11, is required for manganese tolerance and high fertility in rice. Planta 2018, 248, 231–241. [Google Scholar] [CrossRef]
- Alejandro, S.; Holler, S.; Meier, B.; Peiter, E. Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.F.; Zhang, Z.Q.; Deng, J.P.; Miao, C.C.; Wang, Z.Q.; Wallrad, L.; Javed, L.; Fu, D.L.; Zhang, T.; Kudla, J.; et al. Ca2+-dependent successive phosphorylation of vacuolar transporter MTP8 by CBL2/3-CIPK3/9/26 and CPK5 is critical for manganese homeostasis in Arabidopsis. Mol. Plant 2021, 15, 419–437. [Google Scholar] [CrossRef]
- Cailliatte, R.; Schikora, A.; Briat, J.F.; Mari, S.; Curie, C. High-affinity manganese uptake by the metal transporter NRAMP1 is essential for Arabidopsis growth in low manganese conditions. Plant Cell 2010, 22, 904–917. [Google Scholar] [CrossRef]
- Gao, Y.Q.; Chen, J.G.; Chen, Z.R.; An, D.; Lv, Q.Y.; Han, M.L.; Wang, Y.L.; Salt, D.E.; Chao, D.Y. A new vesicle trafficking regulator CTL1 plays a crucial role in ion homeostasis. PLoS Biol. 2017, 15, e2002978. [Google Scholar] [CrossRef] [PubMed]
- Oomen, R.; Wu, J.; Lelièvre, F.; Blanchet, S.; Richaud, P.; Barbier-Brygoo, H.; Aarts, M.G.M.; Thomine, S. Functional characterization of NRAMP3 and NRAMP4 from the metal hyperaccumulator Thlaspi caerulescens. New Phytol. 2008, 181, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Castaings, L.; Caquot, A.; Loubet, S.; Curie, C. The high-affinity metal transporters NRAMP1 and IRT1 team up to take up iron under sufficient metal provision. Sci. Rep. 2016, 6, 37222. [Google Scholar] [CrossRef]
- Zhang, J.L.; Wang, L.L.; Zheng, Y.P.; Feng, J.; Ruan, Y.M.; Diao, S.; Chen, S.N.; Jiang, B.; Shen, Z.G.; Lu, H.F. Effects of Bemisia tabaci (Gennadius) infestation and squash silverleaf disorder on Cucurbita pepo L. leaf. Sci. Hortic. 2017, 217, 8–16. [Google Scholar] [CrossRef]
- Zhang, J.L.; Lu, J.W.; Mantri, N.; Jiang, L.; Ying, S.J.; Chen, S.N.; Feng, X.Y.; Cao, Y.Z.; Chen, Z.C.; Ren, L.C.; et al. An effective combination storage technology to prolong storability, preserve high nutrients and antioxidant ability of astringent persimmon. Sci. Hortic. 2018, 241, 304–312. [Google Scholar] [CrossRef]
- Lian, J.; Zhao, L.; Wu, J.; Xiong, H.; Bao, Y.; Zeb, A.; Tang, J.; Liu, W. Foliar spray of TiO2 nanoparticles prevails over root application in reducing Cd accumulation and mitigating Cd-induced phytotoxicity in maize (Zea mays L.). Chemosphere 2020, 239, 124794. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Zhang, H.Q.; Yang, Y.J.; Fu, G.F.; Tao, L.X.; Xiong, J. Effects and oxygen-regulated mechanisms of water management on cadmium (Cd) accumulation in rice (Oryza sativa). Sci. Total. Environ. 2022, 846, 157484. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Chang, J.D.; Chen, R.J.; Li, H.B.; Lu, H.F.; Tao, L.X.; Xiong, J. Comparison on cellular mechanisms of iron and cadmium accumulation in rice: Prospects for cultivating Fe-rich but Cd-free rice. Rice 2016, 9, 39. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Shoot (g) | Root (g) | Plant Height (cm) | ||
|---|---|---|---|---|---|
| FW | DW | FW | DW | ||
| CK | 3.58 ± 0.06 gh | 0.5198 ± 0.04 fg | 1.602 ± 0.02 e | 0.1169 ± 0.004 cd | 36.67 ± 0.16 ef |
| ALA1 | 4.22 ± 0.03 cd | 0.6223 ± 0.01 bcd | 1.737 ± 0.01 de | 0.1358 ± 0.017 cd | 38.66 ± 0.88 d |
| ALA2 | 3.30 ± 0.07 h | 0.4636 ± 0.01 h | 1.650 ± 0.05 e | 0.1236 ± 0.002 cd | 33.38 ± 0.47 h |
| SA1 | 3.98 ± 0.14 def | 0.5705 ± 0.02 def | 1.741 ± 0.07 de | 0.1365 ± 0.005 cd | 36.17 ± 0.44 f |
| SA2 | 4.19 ± 0.03 cd | 0.6063 ± 0.02 bcd | 1.763 ± 0.03 cde | 0.1380 ± 0.007 cd | 39.17 ± 0.17 cd |
| Mn | 4.07 ± 0.05 de | 0.5740 ± 0.01 de | 1.627 ± 0.01 e | 0.1195 ± 0.004 cd | 38.15 ± 0.67 d |
| Mn + ALA1 | 5.14 ± 0.08 ab | 0.6544 ± 0.01 b | 2.043 ± 0.17 bcd | 0.1405 ± 0.007 bcd | 40.83 ± 0.17 ab |
| Mn + ALA2 | 5.26 ± 0.01 ab | 0.6584 ± 0.01 b | 1.544 ± 0.04 e | 0.1094 ± 0.003 de | 42.00 ± 0.29 a |
| Mn + SA1 | 4.98 ± 0.03 b | 0.6314 ± 0.04 bc | 2.280 ± 0.13 ab | 0.1689 ± 0.024 ab | 40.21 ± 0.27 bc |
| Mn + SA2 | 3.71 ± 0.05 fg | 0.5258 ± 0.01 efg | 1.641 ± 0.02 e | 0.1223 ± 0.008 cd | 35.83 ± 0.33 fg |
| Mn + CA | 5.40 ± 0.25 a | 0.7779 ± 0.01 a | 2.480 ± 0.18 a | 0.1746 ± 0.001 a | 40.33 ± 0.17 bc |
| Mn + CA + ALA1 | 3.50 ± 0.08 gh | 0.5146 ± 0.01 g | 1.640 ± 0.12 e | 0.1220 ± 0.002 cd | 35.33 ± 0.33 fg |
| Mn + CA + ALA2 | 4.14 ± 0.06 cd | 0.5868 ± 0.01 cd | 2.117 ± 0.10 bc | 0.1437 ± 0.008 bc | 36.67 ± 0.44 ef |
| Mn + CA + SA1 | 4.47 ± 0.24 c | 0.6292 ± 0.01 bc | 2.103 ± 0.19 bcd | 0.1425 ± 0.003 bc | 37.83 ± 0.73 de |
| Mn + CA + SA2 | 3.78 ± 0.07 efg | 0.5313 ± 0.02 efg | 1.150 ± 0.24 f | 0.0857 ± 0.013 e | 34.50 ± 0.50 gh |
| Treatments | H2O2 (μmol g−1 FW) | O2− (nmol min−1 g−1 FW) | −OH (μmol g−1 FW) | MDA (nmol g−1 FW) | ||||
|---|---|---|---|---|---|---|---|---|
| Leaf | Root | Leaf | Root | Leaf | Root | Leaf | Root | |
| CK | 17.54 ± 0.21 hi | 14.75 ± 0.68 f | 6.33 ± 0.16 h | 3.49 ± 0.02 d | 98.25 ± 0.28 b | 32.35 ± 0.21 d | 39.43 ± 0.65 e | 14.09 ± 0.28 fg |
| ALA1 | 20.66 ± 1.78 fg | 19.61 ± 0.34 bc | 9.22 ± 1.26 cde | 4.11 ± 0.02 a | 62.20 ± 0.61 k | 34.23 ± 0.28 c | 37.90 ± 0.47 f | 12.90 ± 0.32 fgh |
| ALA2 | 19.35 ± 0.71 gh | 12.65 ± 0.09 g | 6.82 ± 0.36 gh | 3.25 ± 0.03 e | 82.39 ± 0.55 ef | 26.52 ± 0.09 h | 47.04 ± 0.16 a | 19.46 ± 1.55 bc |
| SA1 | 28.12 ± 1.63 a | 13.36 ± 0.08 g | 7.72 ± 0.34 fg | 3.62 ± 0.03 cd | 83.61 ± 0.75 de | 34.29 ± 0.11 c | 42.88 ± 0.54 c | 17.10 ± 0.19 de |
| SA2 | 16.56 ± 0.17 i | 18.53 ± 0.59 d | 10.38 ± 0.57 bc | 3.14 ± 0.01 ef | 77.79 ± 0.56 g | 30.43 ± 0.47 ef | 37.50 ± 0.23 f | 16.45 ± 0.49 e |
| Mn | 24.84 ± 0.23 cd | 17.27 ± 0.25 e | 14.44 ± 0.05 a | 4.07 ± 0.04 a | 84.86 ± 0.56 d | 40.18 ± 0.56 a | 44.62 ± 0.31 b | 21.51 ± 0.17 a |
| Mn + ALA1 | 20.99 ± 0.86 fg | 15.29 ± 0.18 f | 9.72 ± 0.10 cd | 3.03 ± 0.01 fg | 87.91 ± 0.33 c | 34.18 ± 0.54 c | 37.37 ± 0.78 f | 17.42 ± 1.22 de |
| Mn + ALA2 | 21.86 ± 0.19 ef | 15.28 ± 0.03 f | 7.59 ± 0.25 fgh | 2.99 ± 0.07 fg | 72.05 ± 0.37 i | 28.71 ± 0.60 g | 45.70 ± 0.64 b | 19.35 ± 0.19 bc |
| Mn + SA1 | 21.66 ± 0.45 ef | 18.36 ± 0.64 d | 11.39 ± 0.14 b | 3.89 ± 0.02 b | 106.21 ± 0.06 a | 36.43 ± 0.28 b | 41.40 ± 0.31 d | 12.47 ± 0.57 fgh |
| Mn + SA2 | 22.13 ± 0.26 ef | 21.05 ± 0.41 a | 7.51 ± 0.03 fgh | 3.60 ± 0.11 cd | 71.89 ± 0.43 i | 35.20 ± 0.28 c | 33.78 ± 0.32 g | 12.26 ± 1.45 gh |
| Mn + CA | 19.54 ± 0.65 gh | 19.98 ± 0.14 b | 8.39 ± 0.08 ef | 3.90 ± 0.01 b | 76.77 ± 0.20 g | 34.18 ± 0.28 c | 41.40 ± 0.22 d | 20.86 ± 0.22 ab |
| Mn + CA + ALA1 | 25.93 ± 0.17 bc | 18.64 ± 0.16 cd | 7.94 ± 0.03 fg | 3.57 ± 0.02 cd | 72.16 ± 0.16 i | 28.61 ± 0.19 g | 37.10 ± 0.31 f | 18.49 ± 0.11 cd |
| Mn + CA + ALA2 | 17.89 ± 0.47 hi | 16.75 ± 0.31 e | 6.91 ± 0.09 gh | 2.96 ± 0.07 g | 66.11 ± 0.28 j | 31.02 ± 0.28 e | 32.68 ± 0.38 g | 16.68 ± 0.78 de |
| Mn + CA + SA1 | 23.28 ± 0.60 de | 17.36 ± 0.02 e | 7.41 ± 0.08 fgh | 3.12 ± 0.04 ef | 81.00 ± 0.93 f | 40.39 ± 0.21 a | 40.32 ± 0.54 de | 14.25 ± 0.38 f |
| Mn + CA + SA2 | 27.36 ± 0.53 ab | 15.69 ± 0.07 f | 8.54 ± 0.21 def | 3.68 ± 0.05 c | 74.79 ± 0.57 h | 29.89 ± 0.19 f | 45.16 ± 0.543 b | 12.10 ± 0.48 h |
| Treatments | Shoot Mn Concentration (mg kg−1) | Root Mn Concentration (mg kg−1) | BCF of Shoots | BCF of Roots | TF |
|---|---|---|---|---|---|
| Mn | 355.15 ± 29.10 d | 246.04 ± 21.92 ef | 0.44 ± 0.036 d | 0.31 ± 0.03 ef | 1.44 ± 0.12 f |
| Mn + ALA1 | 397.88 ± 29.05 d | 391.23 ± 34.17 b | 0.50 ± 0.036 d | 0.49 ± 0.04 b | 1.02 ± 0.07 g |
| Mn + ALA2 | 578.25 ± 34.98 c | 391.37 ± 5.94 b | 0.72 ± 0.04 c | 0.49 ± 0.01 b | 1.48 ± 0.09 f |
| Mn + SA1 | 679.07 ± 35.97 bc | 383.20 ± 17.88 b | 0.85 ± 0.04 bc | 0.48 ± 0.02 b | 1.77 ± 0.09 e |
| Mn + SA2 | 688.34 ± 19.45 b | 277.88 ± 18.81 de | 0.86 ± 0.02 b | 0.35 ± 0.02 de | 2.48 ± 0.07 b |
| Mn + CA | 8.94 ± 1.22 e | 477.94 ± 12.30 a | 0.01 ± 0.01 e | 0.60 ± 0.02 a | 0.02 ± 0.01 h |
| Mn + CA + ALA1 | 819.34 ± 39.49 a | 348.64 ± 7.40 bc | 1.02 ± 0.05 a | 0.44 ± 0.01 bc | 2.35 ± 0.11 bc |
| Mn + CA + ALA2 | 603.73 ± 30.06 bc | 331.41 ± 6.76 c | 0.75 ± 0.03 bc | 0.41 ± 0.01 c | 1.82 ± 0.09 de |
| Mn + CA + SA1 | 624.59 ± 48.26 bc | 298.57 ± 3.26 cd | 0.78 ± 0.06 bc | 0.37 ± 0.01 cd | 2.09 ± 0.16 cd |
| Mn + CA + SA2 | 602.74 ± 21.84 bc | 216.54 ± 6.57 f | 0.75 ± 0.03 bc | 0.27 ± 0.01 f | 2.78 ± 0.10 a |
| Treatments | Root Mn BCQ (μg) | Shoot Mn BCQ (μg) | Total Mn BCQ (μg) | Remove Efficiency (%) |
|---|---|---|---|---|
| Mn | 29.40 ± 2.62 e | 203.85 ± 16.70 f | 233.25 ± 19.32 f | 0.19 ± 0.016 e |
| Mn+ALA1 | 54.97 ± 4.80 c | 260.37 ± 19.00 e | 315.34 ± 23.80 e | 0.26 ± 0.019 d |
| Mn+ALA2 | 42.82 ± 0.65 d | 380.72 ± 23.03 abc | 423.54 ± 23.68 bc | 0.35 ± 0.020 b |
| Mn+SA1 | 64.72 ± 3.02 b | 428.77 ± 22.71 a | 493.49 ± 25.73 a | 0.41 ± 0.019 a |
| Mn+SA2 | 33.98 ± 2.30 e | 361.93 ± 10.22 bcd | 395.91 ± 12.52 cd | 0.33 ± 0.008 b |
| Mn + CA | 83.45 ± 2.15 a | 6.96 ± 0.95 g | 90.41 ± 3.1 g | 0.08 ± 0.001 f |
| Mn+CA+ALA1 | 42.53 ± 0.90 d | 421.63 ± 20.32 ab | 464.16 ± 21.22 ab | 0.39 ± 0.017 ab |
| Mn+CA+ALA2 | 47.62 ± 0.97 d | 354.27 ± 17.64 cd | 401.89 ± 18.61 bc | 0.33 ± 0.015 bc |
| Mn+CA+SA1 | 42.54 ± 0.46 d | 392.99 ± 30.36 abc | 435.53 ± 30.82 abc | 0.36 ± 0.026 ab |
| Mn+CA+SA2 | 18.56 ± 0.56 f | 320.23 ± 11.60 d | 338.79 ± 12.16 de | 0.28 ± 0.010 cd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Pan, J.; Najeeb, U.; El-Beltagi, H.S.; Huang, Q.; Lu, H.; Xu, L.; Shi, B.; Zhou, W. Promotive Role of 5-Aminolevulinic Acid or Salicylic Acid Combined with Citric Acid on Sunflower Growth by Regulating Manganese Absorption. Antioxidants 2023, 12, 580. https://doi.org/10.3390/antiox12030580
Li J, Pan J, Najeeb U, El-Beltagi HS, Huang Q, Lu H, Xu L, Shi B, Zhou W. Promotive Role of 5-Aminolevulinic Acid or Salicylic Acid Combined with Citric Acid on Sunflower Growth by Regulating Manganese Absorption. Antioxidants. 2023; 12(3):580. https://doi.org/10.3390/antiox12030580
Chicago/Turabian StyleLi, Juanjuan, Jianmin Pan, Ullah Najeeb, Hossam S. El-Beltagi, Qian Huang, Huaijian Lu, Ling Xu, Bixian Shi, and Weijun Zhou. 2023. "Promotive Role of 5-Aminolevulinic Acid or Salicylic Acid Combined with Citric Acid on Sunflower Growth by Regulating Manganese Absorption" Antioxidants 12, no. 3: 580. https://doi.org/10.3390/antiox12030580
APA StyleLi, J., Pan, J., Najeeb, U., El-Beltagi, H. S., Huang, Q., Lu, H., Xu, L., Shi, B., & Zhou, W. (2023). Promotive Role of 5-Aminolevulinic Acid or Salicylic Acid Combined with Citric Acid on Sunflower Growth by Regulating Manganese Absorption. Antioxidants, 12(3), 580. https://doi.org/10.3390/antiox12030580