
Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health
Abstract

1. Introduction
2. Dietary Intake and Composition of Quercetin Glycosides from Foods
3. Intestinal Absorption and Bioavailability of Dietary Quercetin Glycosides
3.1. General Behavior in the Digestive Tract
3.2. Absorption and Conjugation of Quercetin Glucosides in the Small Intestine
3.3. Metabolism of Quercetin Glucosides and Circulation of Their Metabolites
3.4. Catabolism of Quercetin Glycosides by the Microbiota in the Large Intestine
4. Modulation of Bioavailability
4.1. General Comments
4.2. Effects of the Food Matrix
4.3. Effect of Coexisting Ingredients
4.4. Effects of Cooking and Processing
5. Mechanisms of Onset of Atherosclerosis
5.1. Endothelial Dysfunction and Formation of oxLDL
5.2. Phagocytosis of oxLDL by Monocyte-Derived Macrophages and Foam Cell Formation
5.3. Proatherogenic Effect of oxLDL on Endothelial Cells and VSMCs
5.4. Fibrosis of Blood Vessels and Thrombus Formation
6. Proposed Mechanism for CVD Prevention by Quercetin Glycosides
6.1. Mechanism of Anti-Inflammatory Activity of Phenolic Compounds
- Phenolic compounds inhibit the NFκB signaling pathway, resulting in downregulated gene expression of inflammation-inducing enzymes such as cyclooxygenase-2 (COX-2), inducible NOS (iNOS), and 5-lipoxygenase (5-LOX) and inflammatory cytokines such as IL-1β, IL-6, and TNF-α.
- Phenolic compounds prevent the adhesion of monocytes to endothelial cells by suppressing the expression of E-selectin, MCP-1, VCAM-1, and ICAM-1.
- Phenolic compounds attenuate endothelial oxidative stress by inhibiting NOX-dependent ROS production. They also elevate levels of the endothelial anti-atherosclerotic factor NO by accelerating the phosphorylation of eNOS via an AMP-activated protein kinase (AMPK)-dependent mechanism. In addition, phenolic compounds elevate eNOS expression by activating the nuclear factor E2-related factor 2 (Nrf2) pathway.
- Phenolic compounds suppress the migration of VSMCs by inhibiting platelet-derived growth factor-related signaling molecules such as phosphoinositide-3-kinase (PI3K). They also suppress VSMC growth by inducing apoptosis via activation of p38 mitogen-activated protein kinase and P53 signaling pathways.
6.2. Insights from In Vivo Animal Experiments
6.2.1. Modulation of Inflammatory Cytokines and Oxidative Stress
6.2.2. Modulation of Plasma LDL Levels
6.3. In Vivo Animal Studies on the Anti-Atherogenic Effects of Onion Intake
7. Disadvantages of In Vitro Cultured Cell and In Vivo Animal Studies for Elucidating the Mechanism of Action of Quercetin Glycosides
7.1. Mechanisms of Action of Conjugated Metabolites
7.1.1. Modulation of Oxidative Enzymes and Inflammatory Enzymes
7.1.2. Protection of Endothelial Function
7.1.3. VSMCs as Cellular Targets
7.1.4. Vasodilation and Vasoconstriction
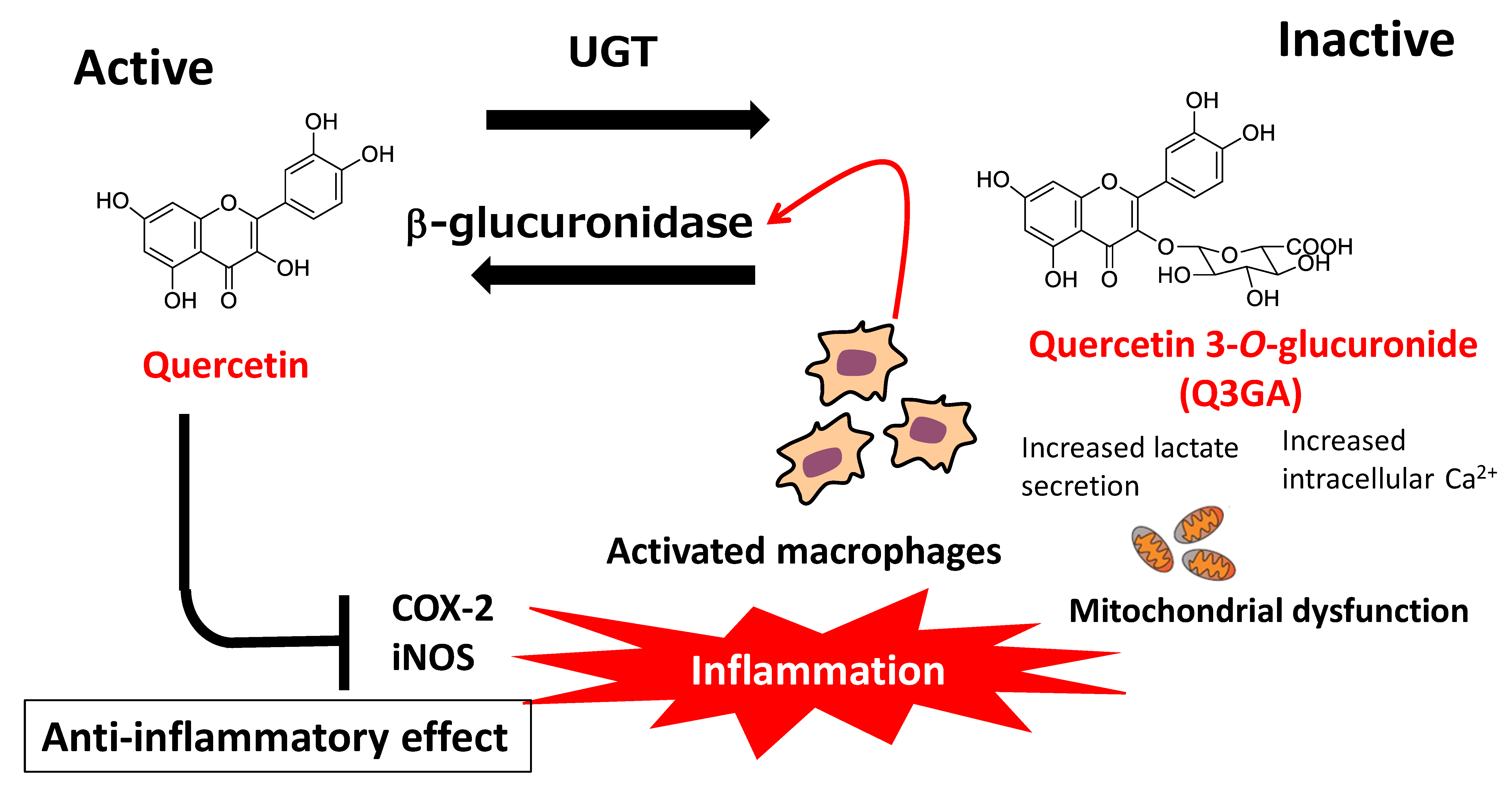
7.2. Deconjugation of Quercetin Glucuronides
8. Contribution of the Microbiota to the Anti-Atherosclerotic Effect of Quercetin Glycosides
8.1. Function of Catabolites Produced by the Action of the Microbiota
8.2. Modulation of Gut Microbiota Profiles
9. Quercetin Glucosides as Senolytics
10. Human Studies on the CVD-Preventive Effects of Quercetin Glycosides
10.1. Observational Studies
10.2. Intervention Studies
11. Conclusions and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mathesius, U. Flavonoid functions in plants and their interactions with other organisms. Plants 2018, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Ferreyra, M.L.F.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef]
- Biswas, A.; Thattai, M. Promiscuity and specificity of eukaryotic glycosyltransferases. Biochem. Soc. Transact. 2020, 48, 891–900. [Google Scholar] [CrossRef]
- Terao, J.; Piskula, M.K.; Yao, Q. Protective effect of epicatechin, epicatechin gallate and quercetin on lipid peroxidation in phospholipid bilayers. Arch. Biochem. Biophys. 1994, 308, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 1995, 4, 375–383. [Google Scholar] [CrossRef]
- Bors, W.; Michel, C.; Stettmaier, K. Antioxidant effects of flavonoids. Biofactors 1997, 6, 399–402. [Google Scholar] [CrossRef]
- Ferrali, M.; Signorini, C.; Caciotti, B.; Sugherini, L.; Ciccoli, L.; Giachetti, D.; Comporti, M. Protection against oxidative damage of erythrocyte membrane by the flavonoid quercetin and its relation to iron chelating activity. FEBS Lett. 1997, 416, 123–129. [Google Scholar] [CrossRef]
- Oboh, G.; Ademosun, A.O.; Ogunsuyi, O.B. Quercetin and its role in chronic diseases. Adv. Exp. Med. Biol. 2016, 929, 377–387. [Google Scholar] [CrossRef]
- Deepika; Maurya, P.K. Health benefits of quercetin in age-related diseases. Molecules 2022, 27, 2498. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef]
- Ferrenczyova, K.; Kalocayova, B.; Bartekova, M. Potential implications of quercetin and its derivatives in cardioprotection. Int. J. Mol. Sci. 2020, 21, 1585. [Google Scholar] [CrossRef] [PubMed]
- WHO Fact Sheets Cardiovascular Diseases 11 June 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvd) (accessed on 14 September 2022).
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencout, M.S.; Tokgozoglu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers. 2019, 5, 56. [Google Scholar] [CrossRef]
- Nanri, A.; Mizoue, T.; Shimazu, T.; Ishihara, J.; Takachi, R.; Noda, M.; Iso, H.; Sasazuki, S.; Sawada, N.; Tugane, S. Dietary patterns and all-cause, cancer, and cardiovascular disease mortality in Japanese men and women: The Japan public health center-based prospective study. PLoS ONE 2017, 12, e0174848. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.; Feskens, E.J.; Hollman, P.C.; Katan, M.B.; Kromhout, D. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.; Kromhout, D.; Aravanis, C.; Blackburn, H.; Buzina, R.; Fidanza, F.; Giampaoli, S.; Jansen, A.; Menotti, A.; Nedeljkovic, S. Flavonoid intake and long-term risk of coronary heart disease and cancer in the seven countries study. Arch. Intern. Med. 1995, 155, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Mink, P.J.; Scrafford, C.G.; Barraj, L.M.; Harnack, L.; Hong, C.-P.; Nettleton, J.A.; Jacobs, D.R., Jr. Flavonoid intake and cardiovascular disease mortality: A prospective study in postmenopausal women. Am. J. Clin. Nutr. 2007, 85, 895–909. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Dalgaard, F.; Kyro, C.; Murray, K.; Bondonno, C.P.; Lewis, J.R.; Croft, K.D.; Gislason, G.; Scalbert, A.; Cassidy, A.; et al. Flavonoid intake is associated with lower mortality in the Daish Diet Cancer and Health Cohort. Nat. Commun. 2019, 10, 3651. [Google Scholar] [CrossRef]
- Kozlowska, A.; Szostak-Wegierek, D. Targeting cardiovascular diseases by flavonols: An update. Nutrients 2022, 14, 1439. [Google Scholar] [CrossRef]
- Grosso, G.J.; Godos, J.; Currenti, W.; Micek, A.; Falzone, L.; Libra, M.; Giampieri, F.; Forbes-Hernandez, T.Y.; Quiles, J.L.; Battino, M.; et al. The effect of dietary polyphenols on vascular health and hypertension: Current evidence and mechanisms of action. Nutrients 2022, 14, 545. [Google Scholar] [CrossRef]
- Jaganath, I.B.; Crozier, A. Dietary flavonoids and phenolic compounds. In Plant Phenolics and Human Health; Fraga, C.G., Ed.; Wiley &Sons Inc: Hoboken, NJ, USA, 2010; pp. 1–51. [Google Scholar]
- Rothwell, J.A.; Pérez-Jiménez, J.; Neveu, V.; Medina-Ramon, A.; M’Hiri, N.; Garcia Lobato, P.; Manach, C.; Knox, K.; Eisner, R.; Wishart, D.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef]
- USDA: USDA Special Interest Databases on Flavonoids. 2015. Available online: https://data.nal.usda.gov/dataset/usda-special-interest-databases-flavonoids (accessed on 14 September 2022).
- Griffith, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions—A global benefit to health. Phytother. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.M.; Barraj, L.M.; Spungen, J.H.; Herman, D.R.; Randolph, K. Global assessment of select phytonutrient intakes by level of fruit and vegetable consumption. Br. J. Nutr. 2014, 112, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
- Walle, T.; Browning, A.M.; Steed, L.L.; Reed, S.G.; Walle, U.K. Flavonoid glucosides are hydrolyzed and thus activated in the oral cavity in humans. J. Nutr. 2005, 135, 48–52. [Google Scholar] [CrossRef]
- Crespy, V.; Morand, C.; Manach, C.; Besson, C.; Demigne, C.; Remesy, C. Part of quercetin absorbed in the small intestine is conjugated and further secreted in the intestinal lumen. Am. J. Physiol. 1999, 277, G120–G126. [Google Scholar] [CrossRef] [PubMed]
- Murota, K.; Terao, J. Antioxidative flavonoid quercetin: Implication of its intestinal absorption and metabolism. Arch. Biochem. Biophys. 2003, 417, 12–17. [Google Scholar] [CrossRef]
- Kawabata, K.; Mukai, R.; Ishisaka, A. Quercetin and related polyphenols: New insights and implications for their bioactivity and bioavailability. Food Funct. 2015, 6, 1399–1417. [Google Scholar] [CrossRef]
- Donovan, C.; Manach, R.M.; Faulks, P.A.; Kroon, P.A. Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; Crozier, A., Clifford, M.N., Ashihara, H., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 303–351. [Google Scholar]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotech. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Day, A.J.; Morgan, M.R.A. Experimental determination of octanol-water partition coefficients of quercetin and related flavonoids. J. Agric. Food Chem. 2005, 53, 4355–4360. [Google Scholar] [CrossRef]
- Wolffram, S.; Block, M.; Ader, P. Quercetin-3-glucoside is transported by the glucose carrier SGLT1 across the brush border membrane of rat small intestine. J. Nutr. 2002, 132, 630–635. [Google Scholar] [CrossRef]
- Day, A.J.; Canada, F.J.; Diaz, J.C.; Kroon, P.A.; Mclauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.A.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 46, 166–170. [Google Scholar] [CrossRef]
- Day, A.J.; Dupont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.C.; Morgan, M.R.A.; Williamson, G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver β-glucosidase activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Ioku, K.; Pongpiriyadacha, Y.; Konishi, Y.; Takei, Y.; Nakatani, N.; Terao, J. β-Glucosidase activity in the rat small intestine toward quercetin monoglucosides. Biosci. Biotechnol. Biochem. 1998, 62, 1428–1431. [Google Scholar] [CrossRef] [PubMed]
- Olthof, M.R.; Hollman, P.C.H.; Vree, T.B.; Katan, M.B. Bioavailability of quercetin-3-glucoside and quercetin-4′-glucoside do not differ in humans. J. Nutr. 2000, 130, 1200–1203. [Google Scholar] [CrossRef]
- Murota, K.; Matsuda, N.; Kashino, Y.; Fujikura, Y.; Nakamura, T.; Kato, Y.; Shimizu, R.; Okuyama, S.; Tanaka, H.; Koda, T.; et al. α-Oligoglucoylation of a sugar moiety enhances the bioavailability of quercetin glucosides in humans. Arch. Biochem. Biophys. 2010, 501, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.C.W.; Sesink, A.L.A.; Faassen-Peters, M.; Hollman, P.C.H. The type of sugar moiety is a determinant of the small intestinal uptake and subsequent biliary excretion of dietary quercetin glycosides. Br. J. Nutr. 2004, 91, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Morand, C.; Demigne, C.; Texier, O.; Regerat, F.; Remesy, C. Bioavailability of rutin and quercetin in rats. FEBS Lett. 1997, 409, 12–16. [Google Scholar] [CrossRef]
- Carbonaro, M.; Grant, G. Absorption of quercetin and rutin in rat small intestine. Ann. Nutr. Metab. 2005, 49, 178–182. [Google Scholar] [CrossRef]
- De Vries, J.H.M.; Hollman, P.C.H.; Meyboom, S.; Buysman, M.N.C.P.; Zock, P.L.; van Staveren, W.A.; Katan, M.B. Plasma concentrations and urinary excretion of the antioxidant flavonols quercetin and kaempferol as biomarkers for dietary intake. Am. J. Clin. Nutr. 1998, 68, 60–65. [Google Scholar] [CrossRef]
- Erlund, I.; Kosonen, T.; Alfthan, G.; Maenpaa, J.; Perttunen, K.; Kenraali, J.; Parantainen, J.; Aro, A. Pharmacokinetics of quercetin from quercetin aglycone and rutin in healthy volunteers. Eur. J. Clin. Pharmacol. 2000, 56, 545–553. [Google Scholar] [CrossRef]
- Jaganath, I.B.; Mullen, W.; Edwards, C.A.; Crozier, A. The relative contribution of the small and large intestine to the absorption and metabolism of rutin in man. Free Radic. Res. 2006, 40, 1035–1046. [Google Scholar] [CrossRef]
- Boonpawa, R.; Spenkelink, A.; Rietjens, I.M.C.M.; Punt, A. A physiologically based kinetic (PBK) model describing plasma concentrations of quercetin and its metabolites in rats. Biochem. Pharmacol. 2014, 89, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Kinjo, C.; Nakamura, Y.; Kato, Y.; Nishikawa, M.; Hamada, M.; Nakajima, N.; Ikushiro, S.; Murota, K. Lymphatic metabolites of quercetin after intestinal administration of quercetin-3-glucoside and its aglycone in rats. Arch. Biochem. Biophys. 2018, 645, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Boitier, A.; Stewart, A.J.; Crozier, A. Flavonoid metabolites in human plasma and urine after the consumption of red onions: Analysis by liquid chromatography with photodiode array and full scan tandem mass spectrometric detection. J. Chromatogr. A 2004, 1058, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Mitchell, A.E. Pharmacokinetics of quercetin absorption from apples and onions in healthy humans. J. Agric. Food Chem. 2012, 60, 3874–3881. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ebeler, S.E.; Zweigenbaum, J.A.Z.; Mitchell, A.E. UHPLC-(ESI)QTOF MS/MS profiling of quercetin metabolites in human plasma postconsumption of applesauce enriched with apple peel and onion. J. Agric. Food Chem. 2012, 60, 8510–8520. [Google Scholar] [CrossRef] [PubMed]
- Boonpawa, R.; Moradi, N.; Spenkelink, A.; Rietjens, I.M.C.M.; Punt, A. Use of physiologically based kinetic (PBK) modeling to study interindividual human variation and species differences in plasma concentrations of quercetin and its metabolites. Biochem. Pharmacol. 2015, 98, 690–702. [Google Scholar] [CrossRef]
- Olthof, M.R.; Hollman, P.C.H.; Buijsman, M.N.C.P.; van Amelsvoort, M.M.; Katan, M.B. Chlorogenic acid, quercetin-3-rutinoside and black tea phenols are extensively metabolized in humans. J. Nutr. 2003, 133, 1806–1814. [Google Scholar] [CrossRef]
- Braune, A.; Gutschow, M.; Engst, W.; Blaut, M. Degradation of quercetin and luteolin by Eubacterium ramulus. Appl. Environ. Microbiol. 2001, 67, 5558–5567. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, S.Y.; Park, S.Y.; Han, M.J. Metabolism of quercitrin by human intestinal bacteria and its relation to some biological activities. Biol. Pharm. Bull. 1999, 22, 749–751. [Google Scholar] [CrossRef]
- Jaganath, I.B.; Mullen, W.; Lean, M.E.J.; Edwards, C.A.; Crozier, A. In vitro catabolism of rutin by human fecal bacteria and the antioxidant capacity of its catabolites. Free Radic. Biol. Med. 2009, 47, 1180–1189. [Google Scholar] [CrossRef]
- Yang, J.; Qian, D.; Guo, J.; Jiang, S.; Shang, E.; Duan, J.; Xu, J. Identification of the major metabolites of hyperoxide produced by the human intestinal bacteria using the ultra performance liquid chromatography/quadrupole-time-of-flight mass spectrometry. J. Ethnopharmacol. 2013, 147, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; Liu, X.; Bolling, B. Flavonoids and gut health. Curr. Opin. Biotechnol. 2020, 61, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Ludwig, I.A.; Polyviou, T.; Malkova, D.; Garcia, A.; Moreno-Rojas, J.M.; Clozier, A. Identification of plasma and urinary metabolites and catabolites derived from orange juice (poly)phenols: Analysis by high-performance liquid chromatography-high resolution mass spectrometry. J Agric. Food Chem. 2016, 64, 5724–5735. [Google Scholar] [CrossRef]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 7, 429–452. [Google Scholar] [CrossRef]
- Jin, F.; Nieman, D.C.; Shanely, R.A.; Knab, A.M.; Austin, M.D.; Sha, W. The variable plasma quercetin response to 12-week quercetin supplementation in humans. Eur. J. Clin. Nutr. 2010, 64, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Bruno, R.S. Endogenous and exogenous mediators of quercetin bioavailability. J. Nutr. Biochem. 2015, 26, 201–210. [Google Scholar] [CrossRef]
- Egert, S.; Wolffram, S.; Schulze, B.; Langguth, P.; Hubbermann, E.M.; Schwarz, K.; Adolphi, B.; Bosy-Westphal, A.; Rimbach, G.; Muller, M.J. Enriched cereal bars are more effective in increasing plasma quercetin compared with quercetin from powder-filled hard capsules. Br. J. Nutr. 2012, 107, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Burak, C.; Brull, V.; Langguth, P.; Zimmerman, B.F.; Stoffel-Wagner, B.; Sausen, U.; Stehle, P.; Wolffram, S.; Egert, S. Higher plasma quercetin levels following oral administration of an onion skin extract compared with pure quercetin dihydrate in humans. Eur. J. Nutr. 2017, 56, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Williamson, G. Comparison of the urinary excretion of quercetin glycosides from red onion and aglycone from dietary supplements in healthy subjects: A randomized single-blinded cross-over study. Food Funct. 2015, 6, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Wiczkowski, W.; Romaszko, J.; Bucinski, A.; Szawara-Nowak, D.; Honke, J.; Zielinski, H.; Piskula, M.K. Quercetin from shallots (Allium cepa L. var. aggregatum) is more bioavailable than its glucosides. J. Nutr. 2008, 138, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Kashino, Y.; Murota, K.; Matsuda, N.; Tomotake, M.; Hamano, T.; Mukai, R.; Terao, J. Effect of processed onions on the plasma concentration of quercetin in rats and humans. J. Food Sci. 2015, 80, H2597–H2602. [Google Scholar] [CrossRef]
- Guo, Y.; Mah, E.; Davis, C.G.; Jalili, T.; Ferruzzi, M.G.; Chun, O.K.; Bruno, R.S. Dietary fat increases quercetin bioavailability in overweight adults. Mol. Nutr. Food Res. 2013, 57, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ippoushi, K.; Ito, H.; Horie, H.; Terao, J. Enhancing effect of lipids and emulsifiers on the accumulation of quercetin metabolites in blood plasma after the short-term ingestion of onion by rats. Biosci. Biotechnol. Biochem. 2003, 67, 2548–2555. [Google Scholar] [CrossRef] [PubMed]
- Nuka, E.; Takahashi, M.; Okitsu, M.; Nayama, C.; Nishijima, H.; Sogawa, R.; Kawabata, K.; Terao, J.; Mukai, R. Lowering effect of combined sweet potato and onion intake on plasma quercetin concentration and underlying mechanism involving intestinal b-glucosidase activity. Biosci. Biotech. Biochem. 2022, 86, 1695–1698. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Murota, K.; Kumamoto, S.; Misumi, K.; Bando, N.; Ikushiro, S.; Takahashi, N.; Sekido, K.; Kato, Y.; Terao, J. Plasma metabolites of dietary flavonoids after combination meal consumption with onion and tofu in humans. Mol. Nutr. Food Res. 2014, 58, 310–317. [Google Scholar] [CrossRef]
- Crozier, A.; Lean, M.E.J.; McDonald, M.S.; Black, C. Quantitative analysis of the flavonoid content of commercial tomatoes, onions, lettuce and celery. J. Agric. Food Chem. 1997, 45, 590–595. [Google Scholar] [CrossRef]
- Ioku, K.; Aoyama, Y.; Tokuno, A.; Terao, J.; Nakatani, N.; Takei, Y. Various cooking methods and the flavonoid content in onion. J. Nutr. Sci. Vitaminol. 2001, 47, 78–83. [Google Scholar] [CrossRef]
- Polia, F.; Pastor-Belda, M.; Martinez-Blazquez, A.; Horcajada, M.-N.; Tomas-Barberan, F.A.; Garcia-Villalba, R. Technological and biotechnological process to enhance the bioavailability of dietary (poly)phenols in humans. J. Agric. Food Chem. 2022, 70, 2092–2107. [Google Scholar] [CrossRef]
- Jebari-Benslaiman, S.; Galicia-Garcia, U.; Larrea-Sebal, A.; Olaetxea, J.R.; Alloza, I.; Vandenbroeck, K.; Benito-Vicente, A.; Martin, C. Pathophysiology of atherosclerosis. Int. J. Mol. Sci. 2022, 23, 3346. [Google Scholar] [CrossRef]
- Li, R.-L.; Wang, L.-Y.; Liu, S.; Duan, H.-X.; Zhang, Q.; Zhang, T.; Peng, W.; Huang, Y.; Wu, C. Natural Flavonoids derived from fruits are potential agents against atherosclerosis. Front. Nutr. 2022, 9, 862277. [Google Scholar] [CrossRef]
- Terao, J. Factors affecting bioavailability of quercetin-related flavonoids and the consequences of their vascular function. Biochem. Pharmacol. 2017, 139, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Rahaman, S.; Islam, R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, S.; et al. Role of phenolic compounds in human disease: Current knowledge and future prospects. Molecules 2022, 27, 233. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jiang, H.; Wu, X.; Fang, J. Therapeutic effects of quercetin on inflammation, obesity, and type 2 diabetes. Mediat. Inflamm. 2016, 2016, 9340637. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Li, C.; Zhang, W.-J.; Frei, B. Quercetin inhibits LPS-induced adhesion molecule expression and oxidant production in human aortic endothelial cells by p38-mediated Nrf2 activation and antioxidant enzyme induction. Redox Biol. 2016, 9, 104–113. [Google Scholar] [CrossRef]
- Morikawa, K.; Nonaka, M.; Narahara, M.; Torii, I.; Kawaguchi, K.; Yoshikawa, T.; Kumazawa, Y.; Morikawa, S. Inhibitory effect of quercetin on carrageenan-induced inflammation in rats. Life Sci. 2003, 74, 709–721. [Google Scholar] [CrossRef]
- Mamani-Matsuda, M.; Kauss, T.; Al-Kharrat, A.; Rambert, J.; Fawaz, F.; Thiolat, D.; Moynet, D.; Coves, S.; Malvy, D.; Mossalayi, M.D. Therapeutic and preventive properties of quercetin in experimental arthritis correlate with decreased macrophage inflammatory mediators. Biochem. Pharmacol. 2006, 72, 1304–1310. [Google Scholar] [CrossRef]
- Milenkovic, M.; Arsenovic-Ranin, N.; Stojic-Vukanic, Z.; Bufan, B.; Vucicevic, D.; Jancic, I. Quercetin ameliorates experimental autoimmune myocarditis in rats. J. Pharm. Pharmaceut. Sci. 2010, 13, 311–319. [Google Scholar] [CrossRef]
- Zheng, J.Y.; Wu, J.H.; Chen, J.; Liu, J.; Lu, Y.Y.; Huang, C.L.; Hu, G.Y.; Wang, X.P.; Zeng, Y. Therapeutic effects of quercetin on early inflammation in hypertriglyceridemia-related acute pancreatitis and its mechanism. Pancreatology 2016, 16, 200–210. [Google Scholar] [CrossRef]
- Annapurna, A.; Reddy, C.S.; Akondi, R.B.; Rao, S.R.C. Cardioprotective actions of two bioflavonoids, quercetin and rutin, in experimental myocardial infarction in both normal and streptozotocin-induced type 1 diabetic rats. J. Pharm. Pharmacol. 2009, 61, 1365–1374. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, J.; Feng, D.; Qin, H.; Wen, H.; Yin, Z.; Gao, G.; Li, C. Protective effect of quercetin against oxidative stress and brain edema in an experimental rat model of subarachnoid hemorrhage. Int. J. Med. Sci. 2014, 11, 282–290. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, L.; Lu, S. Evaluation of antioxidant and immunity activities of quercetin in isoproterenol-treated rats. Molecules 2012, 17, 4281–4291. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Jiang, Y.; Jing, W.; Sun, B.; Miao, C.; Ren, L. Quercetin provides greater cardioprotective effect than its glycoside derivative rutin on isoproterenol-induced cardiac fibrosis in the rat. Can. J. Physiol. Pharmacol. 2013, 91, 951–959. [Google Scholar] [CrossRef]
- Sun, P.; Zhao, L.; Zhang, N.; Zhou, J.; Zhang, L.; Wu, W.; Ji, B.; Zhou, F. Bioactivity of dietary polyphenols: The role in LDL-C lowering. Foods 2021, 10, 2666. [Google Scholar] [CrossRef]
- Nekohashi, M.; Ogawa, M.; Ogihara, T.; Nakazawa, K.; Kato, H.; Misaka, T.; Abe, K.; Kobayashi, S. Luteolin and quercetin affect the cholesterol absorption mediated by epithelial cholesterol transporter Niemann-Pick C1-Like 1 in Caco-2 cells and rats. PLoS ONE 2014, 9, e97901. [Google Scholar] [CrossRef]
- Kamada, C.; da Silva, E.L.; Ohnishi-Kameyama, M.; Moon, J.-H.; Terao, J. Attenuation of lipid peroxidation and hyperlipidemia by quercetin glucoside in the Aorta of high cholesterol-fed rabbit. Free Radic. Res. 2005, 39, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mbikay, M.; Mayne, J.; Sirois, F.; Fedoryak, O.; Raymond, A.; Noad, J.; Chretien, M. Mice fed a high-cholesterol diet supplemented with quercetin-3-glucoside show attenuated hyperlipidemia and hyperinsulinemia associated with differential regulation of PCSK9 and LDLR in their liver and pancreas. Mol. Nutr. Food Res. 2018, 62, e1700729. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cao, H.; Shen, D.; Chen, C.; Xing, S.; Dou, F.; Jia, Q. Effect of quercetin on atherosclerosis based on expressions of ABCA1, LXR-α, and PCSK9 in ApoE-/- mice. Chin. J. Integr. Med. 2020, 26, 114–121. [Google Scholar] [CrossRef]
- Zhang, M.; Xie, Z.; Gao, W.; Pu, L.; Wei, J.; Guo, C. Quercetin regulates hepatic cholesterol metabolism by promoting cholesterol-to-bile acid conversion and cholesterol efflux in rats. Nutr. Res. 2016, 36, 271–279. [Google Scholar] [CrossRef]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Granados, O.; Torre-Villalvazo, I.; Serna-Saldivar, S.O.; Torres, N.; Palacios-Gonzalez, B.; Tovar, A.R. Flavonoids and saponins extracted from black bean (Phaseolus vulgaris L.) seed coats modulate lipid metabolism and biliary cholesterol secretion in C57BL/6 mice. Br. J. Nutr. 2014, 112, 886–899. [Google Scholar] [CrossRef]
- Chambers, K.E.; Day, P.E.; Aboufarrag, H.T.; Kroon, P.A. Polyphenol effects of cholesterol metabolism via bile acid biosynthesis, CYP7A1: A review. Nutrients 2019, 11, 2588. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-M.; Moon, J.; Do, H.J.; Chung, J.H.; Lee, K.-H.; Cha, Y.-J.; Shin, M.-J. Onion peel extract increases hepatic low-density lipoprotein receptor and ATP-binding cassette transporter A1 messenger RNA expression in Sprague-Dawley rats fed a high fat diet. Nutr. Res. 2012, 32, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Pena, D.; Angulo, J.; Vallejo, S.; Colina-Coca, C.; de Ancos, B.; Sanchez-Ferrer, C.F.; Peiro, C.; Sanchez-Moreno, C. High-cholesterol diet enriched with onion affects endothelium-dependent relaxation and NADPH oxidase activity in mesenteric microvessels from Wistar rats. Nutr. Metab. 2014, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Pena, D.; Checa, A.; de Ancos, B.; Wheelock, C.E. New insights into the effects of onion consumption on lipid mediators using a diet-induced model of hypercholesterolemia. Redox Biol. 2017, 11, 205–212. [Google Scholar] [CrossRef]
- Gonzalez-Pena, D.; Gimenez, L.; de Ancos, B.; Sanchez-Moreno, C. Role of dietary onion in modifying the faecal bile acid content in rats fed a high-cholesterol diet. Food Func. 2017, 8, 2184–2192. [Google Scholar] [CrossRef]
- Kang, H.-J.; Balasubramanian, P.; Pichiah, T.; Abinaya, R.V.; Sohn, H.-S.; Cha, Y.-S. Hypocholesterolemic effect of quercetin-rich onion peel extract in C57BL/6J mice fed with high cholesterol diet. Food Sci. Biotechnol. 2016, 25, 855–860. [Google Scholar] [CrossRef]
- Li, W.; Yang, C.; Mei, X.; Huang, R.; Zhang, S.; Tang, Y.; Dong, Q.; Zhou, C. Effect of the polyphenol-rich extract from Allium cepa on hyperlipidemic spraque-dawley rats. J. Food Biochem. 2020, 45, e13565. [Google Scholar] [CrossRef]
- Day, A.J.; Bao, Y.; Morgan, M.R.; Williamson, G. Conjugation position of quercetin glucuronides and effect on biological activity. Free Radic. Biol. Med. 2000, 29, 1234–1243. [Google Scholar] [CrossRef]
- Loke, W.M.; Proudfoot, J.M.; Stewart, S.; Mckinley, A.J.; Needs, P.W.; Kroon, P.A.; Hodgson, J.M.; Croft, K.D. Metabolic transformation has a profound effect on anti-inflammatory activity of flavonoids such as quercetin: Lack of association between antioxidant and lipoxygenase inhibitory activity. Biochem. Pharmacol. 2008, 75, 1045–1053. [Google Scholar] [CrossRef]
- Shiba, Y.; Kinoshita, T.; Chuman, H.; Taketani, Y.; Takeda, E.; Kato, Y.; Naito, M.; Kawabata, K.; Ishisaka, A.; Terao, J.; et al. Flavonoids as substrates and inhibitors of myeloperoxidase: Molecular actions of aglycone and metabolites. Chem Res. Toxicol. 2008, 21, 1600–1609. [Google Scholar] [CrossRef]
- Loke, W.M.; Proudfoot, J.M.; Mckinley, A.J.; Needs, P.W.; Kroon, P.A.; Hodgson, J.M.; Croft, K.D. Quercetin and its in vivo metabolites inhibit neutrophil-mediated low-density lipoprotein oxidation. J. Agric. Food Chem. 2008, 56, 3609–3615. [Google Scholar] [CrossRef] [PubMed]
- De Pascual-Teresa, S.; Johnston, K.L.; Dupont, M.S.; O’Leary, K.A.; Needs, P.W.; Morgan, L.M.; Clifford, M.N.; Bao, Y.; Williamson, G. Quercetin metabolites downregulate cyclooxygenase-2 transcription in human lymphocytes ex vivo but not in vivo. J. Nutr. 2004, 134, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Croft, K.D.; Hodgson, J.M.; Kyle, R.; Lee, I.-L.E.; Wang, Y.; Stocker, R.; Ward, N.C. Quercetin and its metabolites improve vessel function by inducing eNOS activity via phosphorylation of AMPK. Biochem. Pharmacol. 2012, 84, 1036–1044. [Google Scholar] [CrossRef]
- Guo, X.-D.; Zhang, D.-Y.; Gao, X.-J.; Parry, J.; Liu, K.; Liu, B.-L.; Wang, M. Quercetin and quercetin-3-glucuronide are equally effective in ameliorating endothelial insulin resistance through inhibition of reactive oxygen species-associated inflammation. Mol. Nutr. Food Res. 2013, 57, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, M.; Kajiya, K.; Terao, J.; Kaji, K.; Kumazawa, S.; Nakayama, T.; Shimoi, K. Effect of quercetin conjugates on vascular permeability and expression of adhesion molecules. Biofactors 2004, 22, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Tribolo, S.; Lodi, F.; Connor, C.; Suri, S.; Wilson, V.G.; Taylor, M.A.; Needs, P.W.; Kroon, P.A.; Hughes, D.A. Comparative effects of quercetin and its predominant human metabolites on adhesion molecule expression in activated human vascular endothelial cells. Atherosclerosis 2008, 197, 50–56. [Google Scholar] [CrossRef]
- Suri, S.; Taylor, M.A.; Verity, A.; Tribolo, S.; Needs, P.W.; Kroon, P.A.; Hughes, D.A.; Wilson, V.G. A comparative study of the effects of quercetin and its glucuronide and sulfate metabolites on human neutrophil function in vitro. Biochem. Pharmacol. 2008, 76, 645–653. [Google Scholar] [CrossRef]
- Filippini, A.; Sica, G.; D’Alessio, A. The caveolar membrane system in endothelium: From cell signaling to vascular pathology. J. Cell Biochem. 2018, 119, 5060–5071. [Google Scholar] [CrossRef]
- Kamada, C.; Mukai, R.; Kondo, A.; Sato, S.; Terao, J. Effect of quercetin and its metabolite on caveolin-1 expression induced by oxidized LDL and lysophosphatidylcholine in endothelial cells. J. Clin. Biochem. Nutr. 2016, 58, 193–201. [Google Scholar] [CrossRef]
- Kondo-Kawai, A.; Sakai, T.; Terao, J.; Mukai, R. Suppressive effects of quercetin on hydrogen peroxide-induced caveolin-1 phosphorylation in endothelial cells. J. Clin. Biochem. Nutr. 2021, 69, 28–36. [Google Scholar] [CrossRef]
- Winterbone, M.S.; Tribolo, S.; Needs, P.W.; Kroon, P.A.; Hughes, D.A. Physiologically relevant metabolites of quercetin have no effect on adhesion molecule or chemokine expression in human vascular smooth muscle cells. Atherosclerosis 2009, 202, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Yoshizumi, M.; Tsuchiya, K.; Suzaki, Y.; Kirima, K.; Kyaw, M.; Moon, J.-H.; Terao, J.; Tamaki, T. Quercetin glucuronide prevents VSMC hypertrophy by angiotensin II via the inhibition of JNK and AP-1 signaling pathway. Biochem. Biophys. Res. Commun. 2002, 293, 1458–1465. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, K.; Izawa-Ishizawa, Y.; Ohnishi, S.; Motobayashi, Y.; Kawazoe, K.; Hamano, S.; Tsuchiya, K.; Tomita, S.; Minakuchi, K.; Tamaki, T. Quercetin glucuronide inhibits cell migration and proliferation by platelet-derived growth factor in vascular smooth muscle cells. J. Pharmacol. Sci. 2009, 109, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Suri, S.; Rayment, S.; Hughes, D.A.; Kroon, P.A.; Needs, P.W.; Taylor, M.A.; Tribolo, S.; Wilson, V.G. Quercetin and its major metabolites selectively modulate cyclic GMP-dependent relaxations and associated tolerance in pig isolated coronary artery. Br. J. Pharmacol. 2010, 159, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Lodi, F.; Winterbone, M.S.; Tribolo, S.; Needs, P.W.; Hughes, D.A.; Kroon, P.A. Human quercetin conjugated metabolites attenuate TNF-α induced changes in vasomodulatory molecules in an HUASMCs/HUVECs co-culture model. Planta Med. 2012, 78, 1571–1573. [Google Scholar] [CrossRef]
- Terao, J.; Murota, K.; Kawai, Y. Conjugated quercetin glucuronides as bioactive metabolites and precursors of aglycone in vivo. Food Func. 2011, 2, 11–17. [Google Scholar] [CrossRef]
- Perez-Vizcaino, F.; Duarte, J.; Santos-Buelga, C. The Flavonoid paradox: Conjugation and deconjugation as key steps for the biological activity of flavonoids. J. Sci. Food Agric. 2012, 92, 1822–1825. [Google Scholar] [CrossRef]
- Shimoi, K.; Saka, N.; Nozawa, R.; Sato, M.; Amano, I.; Nakayama, T.; Kinae, N. Deglucuronidation of a flavonoid, luteolin monoglucuronide, during inflammation. Drug Metab. Dispos. 2001, 29, 1521–1524. [Google Scholar]
- Kawai, Y.; Nishikawa, T.; Shiba, Y.; Saito, S.; Murota, K.; Shibata, N.; Kobayashi, M.; Kanayama, M.; Uchida, K.; Terao, J. Macrophage as a target of quercetin glucuronides in human atherosclerotic arteries. J. Biol. Chem. 2008, 283, 9424–9434. [Google Scholar] [CrossRef]
- Ishisaka, A.; Kawabata, K.; Miki, S.; Shiba, Y.; Minekawa, S.; Nishikawa, T.; Mukai, R.; Terao, J.; Kawai, Y. Mitochondrial dysfunction leads to deconjugation of quercetin glucuronides in inflammatory macrophages. PLoS ONE 2013, 8, e80843. [Google Scholar] [CrossRef]
- Menendez, C.; Duenas, M.; Galindo, P.; Gonzalez-Manzano, S.; Jimenez, R.; Moreno, L.; Zarzuelo, M.J.; Rodoriguez-Gomez, I.; Duarte, J.; Santos-Buelga, C.; et al. Vascular deconjugation of quercetin glucuronide: The flavonoid paradox revealed? Mol. Nutr. Food Res. 2011, 55, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Galindo, P.; Rodoriguez-Gomez, I.; Gonzalez-Manzano, S.; Duenas, M.; Jimenez, R.; Menendez, C.; Vargas, F.; Tamargo, J.; Santos-Buelga, C.; Perez-Vizcaino, F.; et al. Glucuronidated quercetin lowers blood pressure in spontaneously hypertensive rats via deconjugation. PLoS ONE 2012, 7, e32673. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, R.; Lopez-Sepulveda, R.; Romero, M.; Toral, M.; Cogolludo, A.; Perez-Vizcaino, F.; Duarte, J. Quercetin and its metabolites inhibit the membrane NADPH activity in vascular smooth muscle cells from normotenstive and spontaneously hypertensive rats. Food Funct. 2015, 6, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.; Gonzalez-Manzano, S.; Jimenez, R.; Perez-Abud, R.; Haro, J.M.; Osuna, A.; Santos-Buelga, C.; Duarte, J. The flavonoid quercetin induces acute vasodilator effects in healthy volunteers: Correlation with beta-glucuronidase activity. Pharm. Res. 2014, 89, 11–18. [Google Scholar] [CrossRef]
- Nishikawa, M.; Kada, Y.; Kimata, M.; Sakaki, T.; Ikushiro, S. Comparison of metabolism and biological properties among positional isomers of quercetin glucuronide in LPS- and RANKL-challenged RAW264.7 cells. Biosci. Biotechnol. Biochem. 2022, 86, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Minihane, A.-M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef]
- Feng, J.; Ge, C.; Li, W.; Li, R. 3-(3-hydroxyphenyl)propionic acid, a microbial metabolite of quercetin, inhibits monocyte binding to endothelial cells via modulating E-selectin expression. Fitoterapia 2022, 156, 105071. [Google Scholar] [CrossRef]
- Monagas, M.; Khan, N.; Andres-Lacueva, C.; Urpi-Sarda, M.; Vazquez-Agell, M.; Lamuela-Raventos, R.M.; Estruch, R. Dihydroxylated phenolic acids derived from microbial metabolism reduce lipopolysaccharide-stimulated cytokine secretion by human peripheral blood mononuclear cells. Br. J. Nutr. 2009, 102, 201–206. [Google Scholar] [CrossRef]
- Xue, H.; Xie, W.; Jiang, Z.; Wang, M.; Wang, J.; Zhao, H.; Zhang, X. 3,4-Dihydroxyphenylacetic acid, a microbiota-derived metabolite of quercetin, attenuates acetoaminophen (APAP)-induced liver injury through activation of Nrf-2. Xenobiotica 2016, 46, 931–939. [Google Scholar] [CrossRef]
- Tang, Y.; Nakashima, S.; Saiki, S.; Myoi, Y.; Abe, N.; Kuwazuru, S.; Zhu, B.; Ashida, H.; Murata, Y.; Nakamura, Y. 3,4-Dihydroxyphenylacetic acid is a predominant biologically-active catabolite of quercetin glycosides. Food Res. Int. 2016, 89, 716–723. [Google Scholar] [CrossRef]
- Su, K.-Y.; Yu, C.Y.; Chen, Y.-P.; Hua, K.-F.; Chen, Y.-L.S. 3,4-Dihydroxytoluene, a metabolite of rutin, inhibits inflammatory responses in lipopolysaccharide-activated macrophages by reducing the activation of NF-kB signaling. BMC Complement. Altern. Med. 2014, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Glasser, G.; Graefe, E.U.; Struck, G.; Veit, M.; Gebhardt, R. Comparison of antioxidative capacities and inhibitory effect on cholesterol biosynthesis of quercetin and potential metabolites. Phytomedicine 2002, 9, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Pourova, J.; Najmanova, I.; Voprsalova, M.; Migkos, T.; Pilarova, V.; Applova, L.; Novakova, L.; Mladenk, P. Two flavonoid metabolites, 3,4-dihydroxyphenylacetic acid and 4-methylcatechol, relax arteries ex vivo and decrease blood pressure in vivo. Vascul. Pharmacol. 2018, 111, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Emoto, T.; Sasaki, N.; Hirata, K. Gut microbiota and coronary artery disease. Int. Heart J. 2016, 57, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Koudoufio, M.; Desjardins, Y.; Feldman, F.; Spahis, S.; Delvin, E.; Levy, E. Insight into polyphenol and gut microbiota crosstalk: Are their metabolites the key to understand protective effects against metabolic disorders? Antioxidants 2020, 9, 982. [Google Scholar] [CrossRef]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar]
- Nie, J.; Zhang, L.; Zhao, G.; Du, X. Quercetin reduces atherosclerotic lesions by altering the gut microbiota and reducing atherogenic lipid metabolites. J. App. Microbiol. 2019, 127, 1824–1834. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef]
- Ekbatan, S.S.; Sleno, L.; Sabally, K.; Khairallah, J.; Azadi, B.; Rodes, L.; Prakash, S.; Donnelly, D.J.; Kubow, S. Biotransformation of polyphenols in a dynamic multistage gastrointestinal model. Food Chem. 2016, 204, 453–462. [Google Scholar] [CrossRef]
- Fernandez-Jalao, I.; Balderas, C.; Calvo, M.V.; Fontecha, J.; Sanchez-Moreno, C.; De Ancos, B. Impact of high-pressure processed onion on colonic metabolism using a dynamic gastrointestinal digestion simulator. Metabolites 2021, 11, 262. [Google Scholar] [CrossRef]
- Dos Santos, A.S.; de Albuquerque, T.M.R.; de Brito Alves, J.L.; de Souza, E.L. Effects of quercetin and resveratrol on in vitro properties related to the functionality of potentially probiotic lactobacillus strains. Front. Microbiol. 2019, 10, 2229. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Sugiyama, Y.; Sakano, T.; Ohigashi, H. Flavonols enhanced production of anti-inflammatory substances by Bifidobacterium adolescentis: Prebiotic actions of galangin, quercetin, and fisetin. Biofactors 2013, 39, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Kato, Y.; Sakano, T.; Baba, N.; Hagiwara, K.; Tamura, A.; Baba, S.; Natsume, M.; Ohigashi, H. Effects of phytochemicals on in vitro anti-inflammatory activity of Bifidobacterium adolescentis. Biosci. Biotechnol. Biochem. 2015, 79, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Volstatova, T.; Marchica, A.; Hroncova, Z.; Bernardi, R.; Doskocil, I.; Havlik, J. Effects of chlorogenic acid, epicatechin gallate, and quercetin on mucin expression and secretion in the Caco-2/HT29-MTX cell model. Food Sci. Nutr. 2019, 7, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Eng. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef]
- Hu, Y.; Zhao, Y.; Yuan, L.; Yang, X. Protective effects of tartary buckwheat flavonoids on high TMAO diet-induced vascular dysfunction and liver injury in mice. Food Funct. 2015, 6, 3359–3372. [Google Scholar] [CrossRef]
- Hua, F.; Zhou, P.; Bao, G.-H.; Ling, T.-J. Flavonoids in Lu’an Guapian tea as potential inhibitors of TMA-lyase in acute myocardial infarction. J. Food Biochem. 2022, 46, e14110. [Google Scholar] [CrossRef]
- Bloom, S.I.; Islam, M.T.; Lesniewski, L.A.; Donato, A.J. Mechanisms and consequences of endothelial cell senescence. Nat. Rev. Cardiol. 2023, 20, 38–51. [Google Scholar] [CrossRef]
- Shimizu, I.; Minamino, T. Cellular senescence in arterial diseases. J. Lipid Atheroscler. 2020, 9, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Gasek, N.; Kuchel, G.A.; Kirkland, J.L.; Xu, M. Strategies for targeting senescent cells in human disease. Nat. Aging 2021, 1, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, J.L.; Tchkonia, T. Selolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef]
- Zhu, Y.; Tchkonia, T.; Pirskhalava, T.; Gower, A.C.; Ding, H.; Giogadeze, N.; Palmer, A.K.; Ikeno, Y.; Hubbard, G.B.; Lenburg, M.; et al. The Achilles’ heel of senescent cells: From transcriptome to senolytic drugs. Aging Cell 2015, 14, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Hickson, L.J.; Prata, L.G.P.L.; Bobart, S.A.; Evans, T.K.; Giorgadze, N.; Hashmi, S.K.; Herrmann, S.M.; Jensen, M.D.; Jia, Q.; Jordan, K.L.; et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine 2019, 47, 446–456. [Google Scholar] [CrossRef]
- Mennen, L.I.; Sapinho, D.; de Bree, A.; Arnault, N.; Bertrais, S.; Galan, P.; Hercberg, S. Consumption of foods rich in flavonoids is related to a decreased cardiovascular risk in apparently healthy French women. J. Nutr. 2004, 134, 923–926. [Google Scholar] [CrossRef]
- Ivey, K.L.; Hodgson, J.M.; Croft, K.D.; Lewis, J.R.; Prince, R.L. Flavonoid intake and all-cause mortality. Am. J. Clin. Nutr. 2015, 101, 1012–1020. [Google Scholar] [CrossRef]
- Samieri, C.; Sun, Q.; Townsend, M.K.; Rimm, E.B.; Grodstein, F. Dietary flavonoid intake at midlife healthy aging in women. Am. J. Clin. Nutr. 2014, 100, 1489–1497. [Google Scholar] [CrossRef]
- Micek, A.; Godos, J.; Del Rio, D.; Galvano, F.; Grosso, G. Dietary flavonoids and cardiovascular disease: A comprehensive dose-response meta-analysis. Mol. Nutr. Food Res. 2021, 65, e2001019. [Google Scholar] [CrossRef]
- Galeone, C.; Tavani, A.; Pelucchi, C.; Negri, E.; La Vecchia, C. Allium vegetable intake and risk of acute myocardial infarction in Italy. Eur. J. Nutr. 2009, 48, 120–123. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Momenan, A.A.; Azizi, F. Allium vegetable intakes and the incidence of cardiovascular disease, hypertension, chronic kidney disease, and type 2 diabetes in adults: A longitudinal follow-up study. J. Hypertens. 2017, 35, 1909–1916. [Google Scholar] [CrossRef] [PubMed]
- Serban, M.-C.; Sahebkar, A.; Zanchetti, A.; Mikhailidis, D.P.; Howard, G.; Antal, D.; Andrica, F.; Ahmed, A.; Aronow, W.S.; Muntner, P.; et al. Effects of quercetin on blood pressure: A systematic review and meta-analysis of randomized controlled trials. J. Am. Heart Assoc. 2016, 5, e002713. [Google Scholar] [CrossRef]
- Kay, C.D.; Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cassidy, A. Relative impact of flavonoid composition, dose and structure on vascular function: A systematic review of randomized controlled trials of flavonoid-rich food products. Mol. Nutr. Food Res. 2012, 56, 1605–1616. [Google Scholar] [CrossRef] [PubMed]
- Loke, W.M.; Hodgson, J.M.; Proudfoot, J.M.; McKinley, A.J.; Puddey, I.B.; Croft, K.D. Pure dietary flavonoids quercetin and (−)-epicatechin augment nitric oxide products and reduce endothelin-1 acutely in healthy men. Am. J. Clin. Nutr. 2008, 88, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Bosy-Westphal, A.; Seiberl, J.; Kurbitz, C.; Settler, U.; Plachta-Danielzik, S.; Wagner, A.E.; Frank, J.; Schrezenmeir, J.; Rimbach, G.; et al. Quercetin reduces systolic blood pressure and plasma oxidized low-density lipoprotein concentrations in overweight subjects with a high-cardiovascular disease risk phenotype: A double-blinded, placebo-controlled cross-over study. Br. J. Nutr. 2009, 102, 1065–1074. [Google Scholar] [CrossRef]
- Lee, K.-H.; Park, E.; Lee, H.-J.; Kim, M.-O.; Cha, Y.-J.; Kim, J.-M.; Lee, H.; Shin, M.-J. Effects of daily quercetin-rich supplementation on cardiometabolic risks in male smokers. Nutr. Res. Pract. 2011, 5, 28–33. [Google Scholar] [CrossRef]
- Dower, J.I.; Geleijnse, J.M.; Gijsbers, L.; Schalkwijk, C.; Kromhout, D.; Hollman, P.C.H. Supplementation of the pure flavonoids epicatechin and quercetin affects some biomarkers of endothelial dysfunction and inflammation in (pre)hypertensive adults: A randomized double-blind, placebo-controlled, crossover trial. J. Nutr. 2015, 145, 1459–1463. [Google Scholar] [CrossRef]
- Dower, J.I.; Geleijnse, J.M.; Gijsbers, L.; Zock, P.L.; Kromhout, D.; Hollman, P.C.H. Effect of the pure flavonoids epicatechin and quercetin on vascular function and cardiometabolic health: A randomized, double-blind placebo-controlled, crossover trial. Am. J. Clin. Nutr. 2015, 101, 914–921. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Bondonno, C.P.; Rich, L.; Mas, E.; Shinde, S.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. Acute effects of quercetin-3-O-glucoside on endothelial function and blood pressure: A randomized dose-response study. Am. J. Clin. Nutr. 2016, 104, 97–103. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Bondonno, C.P.; Ward, N.C.; Woodman, R.J.; Hodgson, J.M.; Croft, K.D. Enzymatically modified isoquercitrin improves endothelial function in volunteers at risk of cardiovascular disease. Br. J. Nutr. 2020, 123, 182–189. [Google Scholar] [CrossRef]
- Ou, Q.; Zheng, Z.; Zhao, Y.; Lin, W. Impact of quercetin on systemic levels of inflammation: A meta-analysis of randomized controlled human trials. Int. J. Food Sci. Nutr. 2020, 71, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Janssen, K.; Mensink, R.P.; Cox, F.J.; Harryvan, J.L.; Hovenier, R.; Hollman, P.C.; Katan, M.B. Effects of the flavonoids quercetin and apigenin on hemostasis in healthy volunteers: Results from an in vitro and dietary supplement study. Am. J. Clin. Nutr. 1998, 67, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Lean, M.E.J.; Noroozi, M.; Kelly, I.; Burns, J.; Talwar, D.; Sattar, N.; Crozier, A. Dietary flavonols protect diabetic human lymphocytes against oxidative damage to DNA. Diabetes 1999, 48, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.P.; Dobson, V.L.; Duthie, S.J.; Kyle, J.A.; Collins, A.R. Absorption and DNA protective effect of flavonoid glycosides from an onion meal. Eur. J. Nutr. 2000, 39, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, H.; Tsuge, N.; Sawada, H.; Higashi, Y. Chronic intake of onion extract containing quercetin improved postprandial endothelial dysfunction in healthy men. J. Am. Coll. Nutr. 2013, 32, 160–164. [Google Scholar] [CrossRef]
- Ebrahimi-Mamaghani, M.; Saghafi-Asl, M.; Pirouzpanah, S.; Asghari-Jafarabadi, M. Effects of raw red onion consumption on metabolic features in overweight or obese women with polycystic ovary syndrome: A randomized controlled clinical trial. J. Obstet. Gynaecol. Res. 2014, 40, 1067–1076. [Google Scholar] [CrossRef]
- Brull, V.; Burak, C.; Stoffel-Wagner, B.; Wolffram, S.; Nickenig, G.; Muller, C.; Langguth, P.; Alteheld, B.; Fimmers, R.; Naaf, S.; et al. Effects of a quercetin-rich onion skin extract on 24h ambulatory blood pressure and endothelial function in overweight-to-obese patients with (pre-)hypertension: A randomized double blinded placebo-controlled cross over trial. Br. J. Nutr. 2015, 114, 1263–1277. [Google Scholar] [CrossRef]
- Choi, E.-Y.; Lee, H.; Woom, J.S.; Jang, H.H.; Hwang, S.J.; Kim, H.S.; Kim, W.-S.; Kim, Y.-S.; Choue, R.; Cha, Y.-J.; et al. Effect of onion peel extract on endothelial function and endothelial progenitor cells in overweight and obese individuals. Nutrition 2015, 31, 1131–1135. [Google Scholar] [CrossRef]
- Chiu, H.-F.; Shen, Y.-C.; Huang, T.-Y.; Venkatakrishnan, K.; Wang, C.-K. Cardioprotective efficacy of red wine extract of onion in healthy hypercholesterolemic subjects. Phytother. Res. 2016, 30, 380–385. [Google Scholar] [CrossRef]
- Brull, V.; Burak, C.; Stoffel-Wagner, B.; Wolffram, S.; Nickenig, G.; Muller, C.; Langguth, P.; Alteheld, B.; Fimmers, R.; Stehle, P.; et al. No effects of quercetin from onion skin extract on serum leptin and adiponectin concentrations in overweight-to-obese patients with (pre-)hypertension: A randomized double-blinded, placebo-controlled crossover trial. Eur. J. Nur. 2017, 56, 2265–2275. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodoriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Sesso, H.D.; Manson, J.E.; Aragaki, A.K.; Rist, P.M.; Johnson, L.G.; Friedenberg, G.; Copeland, T.; Clar, A.; Mora, S.; Moorthy, M.V.; et al. Effect of cocoa flavanol supplementation for the prevention of cardiovascular disease events: The Cocoa Supplement and Multivitamin Outcomes Study (COSMOS) randomized clinical trial. Am. J. Clin. Nutr. 2022, 115, 1490–1500. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.X.; Lin, F.-J.; Li, H.; Li, H.-B.; Wu, D.-T.; Geng, F.; Ma, W.; Wang, Y.; Miao, B.-H.; Gan, R.-Y. Recent advances in bioactive compounds, health functions, and safety concerns of onion (Allium cepa L.). Front. Nutr. 2021, 8, 669805. [Google Scholar] [CrossRef] [PubMed]
- Landerberg, R.; Manach, C.; Kerckhof, F.-M.; Minihane, A.-M.; Noureldin, R.; Saleh, M.; De Roos, B.; Tomas-Barberan, F.; Morand, C.; Van de Wiele, T. Future prospects for dissecting inter-individual variability in the absorption, distribtution and elimination of plant bioactives of relevance for cardiometabolic endpoints. Eur. J. Nutr. 2019, 58, S21–S36. [Google Scholar] [CrossRef]
- Alotaibi, B.S.; Ijaz, M.; Buabeid, M.; Kharaba, Z.J.; Yaseen, H.S.; Murtaza, G. Therapeutic effects of plant-derived polyphenolic compounds in cardiovascular diseases: A review. Drug Des. Devel. Ther. 2021, 15, 4713–4732. [Google Scholar] [CrossRef]
- Kumar, M.; Barbhai, M.D.; Hasan, M.; Punia, S.; Dhumal, S.; Radha; Rais, N.; Chandran, D.; Pandiselvam, R.; Kothakota, A.; et al. Onion (Allium cepa L.) peels: A review on bioactive compounds and biomedical activities. Biomed. Pharmacother. 2022, 146, 112498. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Vegetables | Mean Content ± SD * (mg/100 g) | Minimum~Maximum Content (mg/100 g) |
|---|---|---|---|
| isorhamnetin 4′-O-glucoside | onion [red] | 6 | --- |
| onion [yellow] | 2.89 ± 0.88 | 1.90~3.80 | |
| quercetin 3,4′-O-diglucoside | onion [red] | 77.08 ± 64.44 | 20.22~207.49 |
| onion [white] | 3.12 ± 2.50 | 0~5.00 | |
| onion [yellow] | 26.58± 31.11 | 11.37~111.7 | |
| Shallot | 74.62 | --- | |
| quercetin 3-O-(6′′-malonyl-glucoside) | lettuce [green] | 1.85 ± 2.79 | 0.03~7.33 |
| lettuce [red] | 10.44 ±6.29 | 5.99~14.89 | |
| quercetin 3-O-garactoside (hyperoside) | lettuce [green] | 0.10 ± 0.11 | 0~0.26 |
| lettuce [red] | 0.14 ± 0.19 | 0~0.27 | |
| quercein 3-O-glucoside (isoquercitrin) | broccoli | 1.80 | --- |
| lettuce [green] | 0.37± 0.60 | 0.01~1.56 | |
| lettuce [red] | 2.19 ±1.91 | 0.84~3.54 | |
| onion [red] | 1.80 ± 2.55 | 0~3.60 | |
| onion [yellow] | 0.70 ± 0.45 | 0~1.50 | |
| quercetin 3-O-glucuronide (miquelianin) | lettuce [green] | 1.34 ± 1.44 | 0.01~3.70 |
| lettuce [red] | 2.65 ±1.82 | 1.36~3.94 | |
| green bean | 0.80 ± 0.44 | 0.35~1.51 | |
| quercetin 3-O-rhamnoside (quercitrin) | lettuce [green] | 0.13 ± 0.22 | 0~0.58 |
| lettuce [red] | 0.85 ± 0.58 | 0.44~1.26 | |
| green bean | 1.99 ± 1.07 | 0.93~3.50 | |
| quercetin 3-O-rutioside (rutin) | olive [black] | 45.36 ± 28.93 | 11.12~78.70 |
| tomato [cherry] whole | 3.33 ± 1.77 | 1.79~6.61 | |
| tomato whole | 0.14 ± 0.06 | 0.04~0.22 | |
| zucchini | 1.32 ± 0.57 | 0.80~2.26 | |
| lettuce green | 0.04 ± 0.10 | 0~0.24 | |
| onion [red] | 0.21 ±0.06 | 0.17~0.27 | |
| onion [yellow] | 0.68 ±0.40 | 0.18~1.36 | |
| green bean | 2.49± 2.37 | 0.02~6.66 | |
| asparagus | 23.19 ± 10.70 | 0~28.65 | |
| quercetin 3-O-sophoroside (biamaside) | broccoli | 6.5 | --- |
| Quercetin 4′-O-glucusode (spiraeoside) | onion [red] | 38.00 ± 29.51 | 30.01~114.30 |
| onion [white] | 2.25 ± 1.80 | 0~3.60 | |
| onion [yellow] | 21.55 ±16.86 | 13.77~83.03 | |
| shallot | 35.60 | --- |
| Dose/Day | Period | Number of Subjects | Biomarkers Affected | Effect | Remarks | Reference |
|---|---|---|---|---|---|---|
| whole 220 g (114 mg quercetin equivalent) | 7 days | 18 healthy | platelet aggregation | no effect | randomized crossover trial | Jansen et al. [176] |
| whole 400 g (76~110 mg quercetin equivalent) | 14 days | 10 type2 diabetic | lymphocyte DNA damage | − | randomized crossover trial | Lean et al. [177] |
| fried onion 200 g | 1 day | 6 healthy | lymphocyte DNA damage 8-OHdG | − − | randomized two phase crossover trial | Boyle et al. [178] |
| extract 4.3 g (51 mg quercetin equivalent) | 1 day | 23 healthy | FMD | + | before and after study | Nakayama et al. [179] |
| 2 whole (each 50~60 g) | 8 weeks | 54 overweight and obese | total cholesterol LDL | − − | randomized control parallel trial | Mamaghani et al. [180] |
| skin extract (162 mg quercetin equivalent) | 6 weeks | 31 hypertensive | LDL ambulatory blood pressure | − | randomized crossover trial | Brull et al. [181] |
| peel extract (100 mg quercetin equivalent) | 12 weeks | 72 healthy overweight and obese | FMD endothelial progenitor cells | + | randomized, double blind placebo- controlled trial | Choi et al. [182] |
| red wine extract 250 mL | 10 weeks | 23 healthy hyper-cholesterolemic | total cholesterol LDL | − − | randomized placebo-controlled trial | Chiu et al. [183] |
| skin extract (162 mg quercetin equivalent) | 6 weeks | 68 overweight to obese | serum CRP | no effect | double blind placebo-controlled crossover trial | Brull et al. [184] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terao, J. Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health. Antioxidants 2023, 12, 258. https://doi.org/10.3390/antiox12020258
Terao J. Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health. Antioxidants. 2023; 12(2):258. https://doi.org/10.3390/antiox12020258
Chicago/Turabian StyleTerao, Junji. 2023. "Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health" Antioxidants 12, no. 2: 258. https://doi.org/10.3390/antiox12020258
APA StyleTerao, J. (2023). Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health. Antioxidants, 12(2), 258. https://doi.org/10.3390/antiox12020258