

Targeting Pro-Oxidant Iron with Exogenously Administered Apotransferrin Provides Benefits Associated with Changes in Crucial Cellular Iron Gate Protein TfR in a Model of Intracerebral Hemorrhagic Stroke in Mice

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Hypoxic/Ischemic Stroke Model (IS)
2.3. Intracerebral Hemorrhage Stroke Model (ICH)
2.4. Tape Removal Test
2.5. Tail Bleeding Test
2.6. Blood and Brain Sampling and Processing
2.7. Assessment of the Ischemic Infarct Volume in the IS Model, Hemorrhage Volume, and Brain Hemoglobin and Heme Concentration in the ICH Model
2.8. RT-qPCR
2.9. Western Blot (WB)
2.10. Determination of 4-Hydroxynonenal (4-HNE) Using Immunohistochemistry (IHC)
2.11. TSAT Assessment
2.12. Antibodies
2.13. Statistics
3. Results
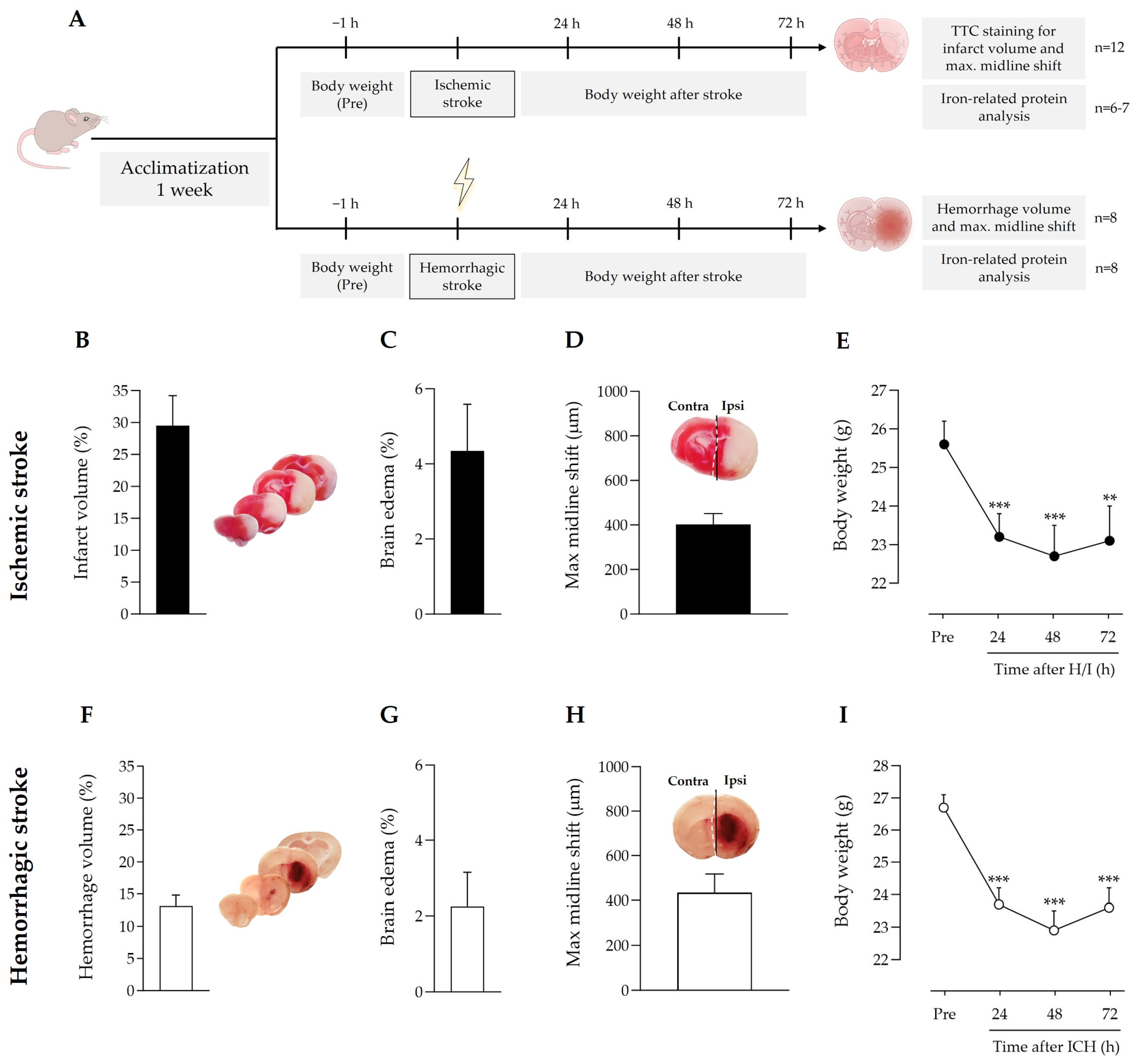
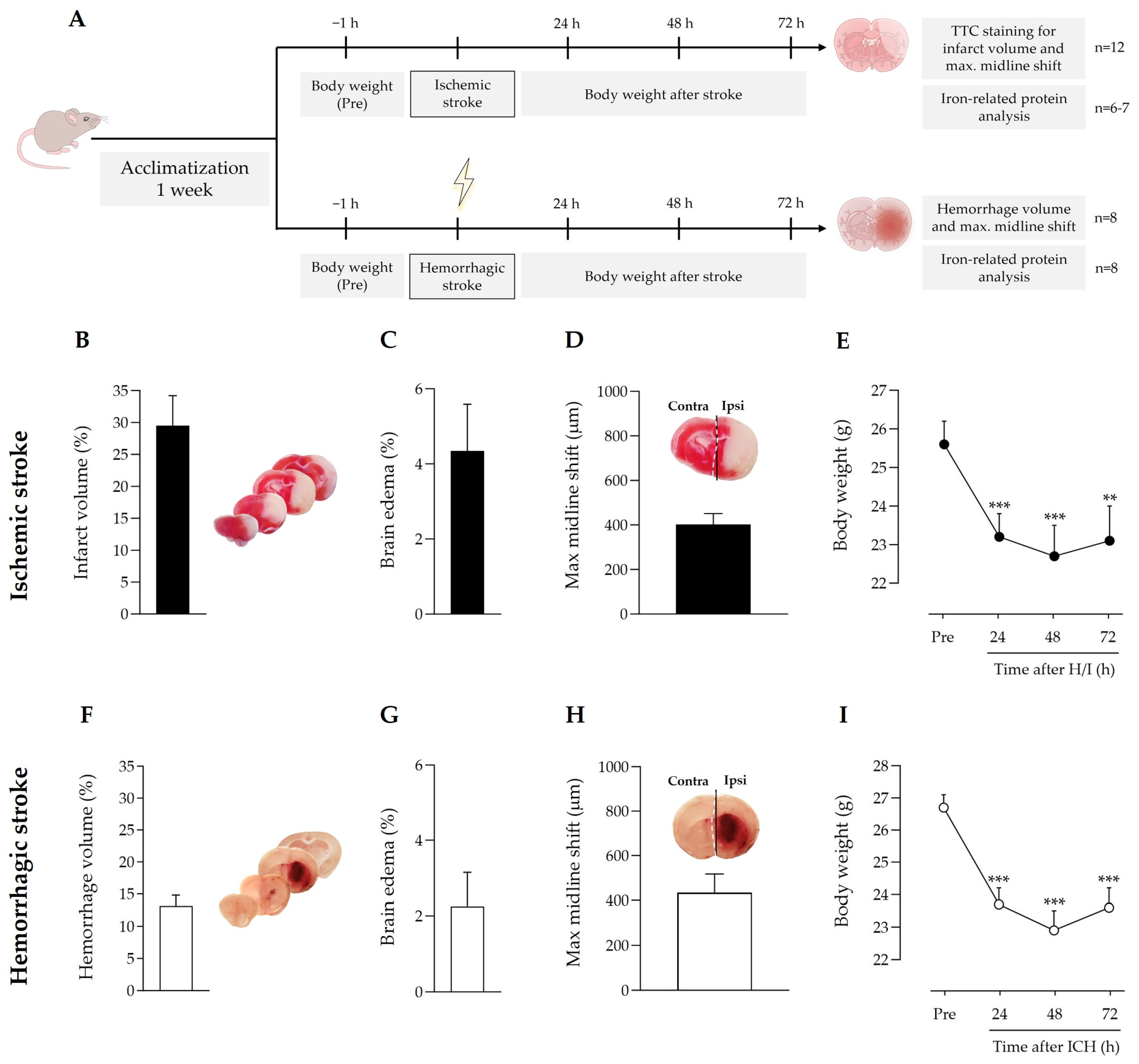
3.1. Gross Differences and Commonalities between Ischemic and Hemorrhagic Stroke Models
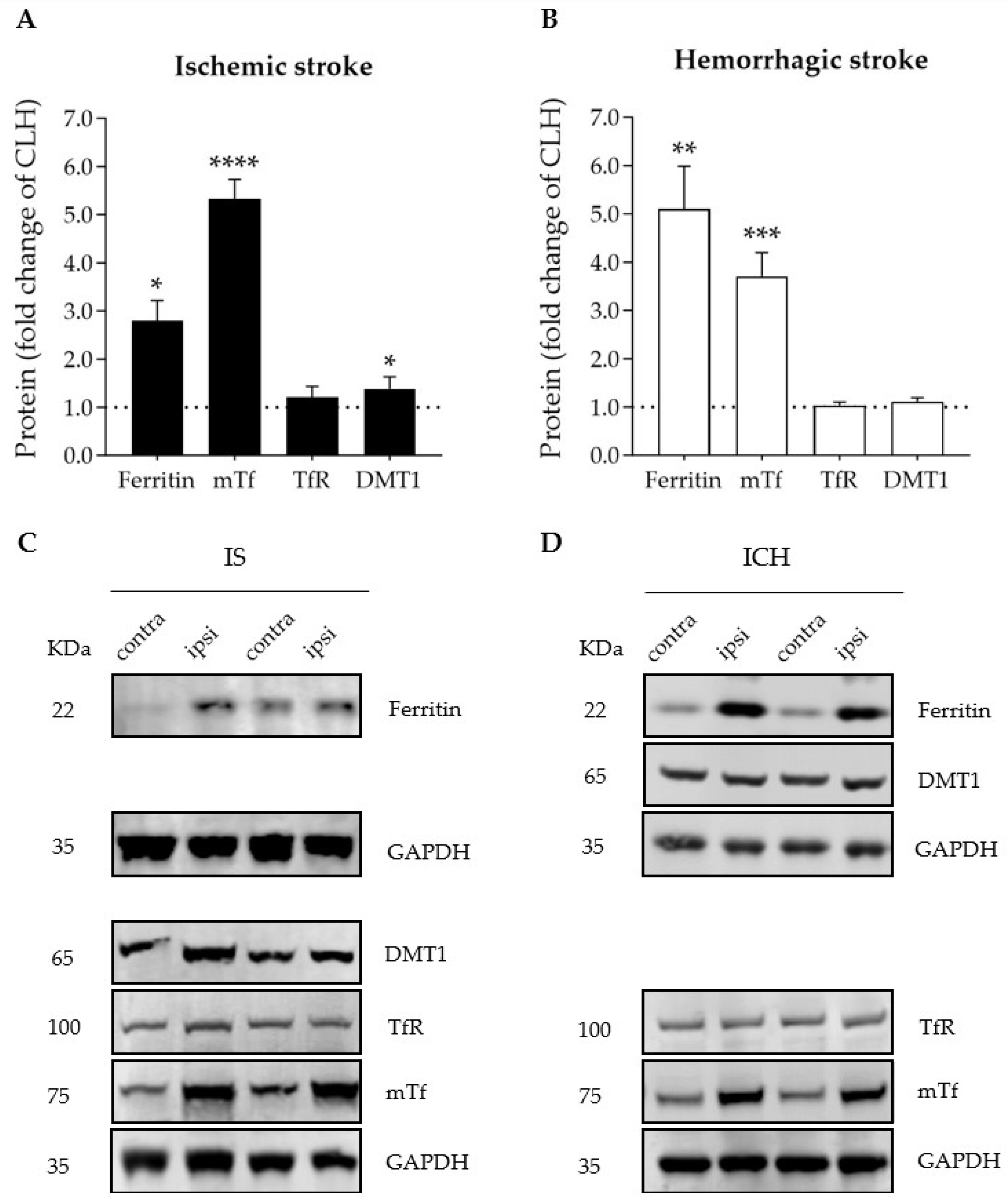
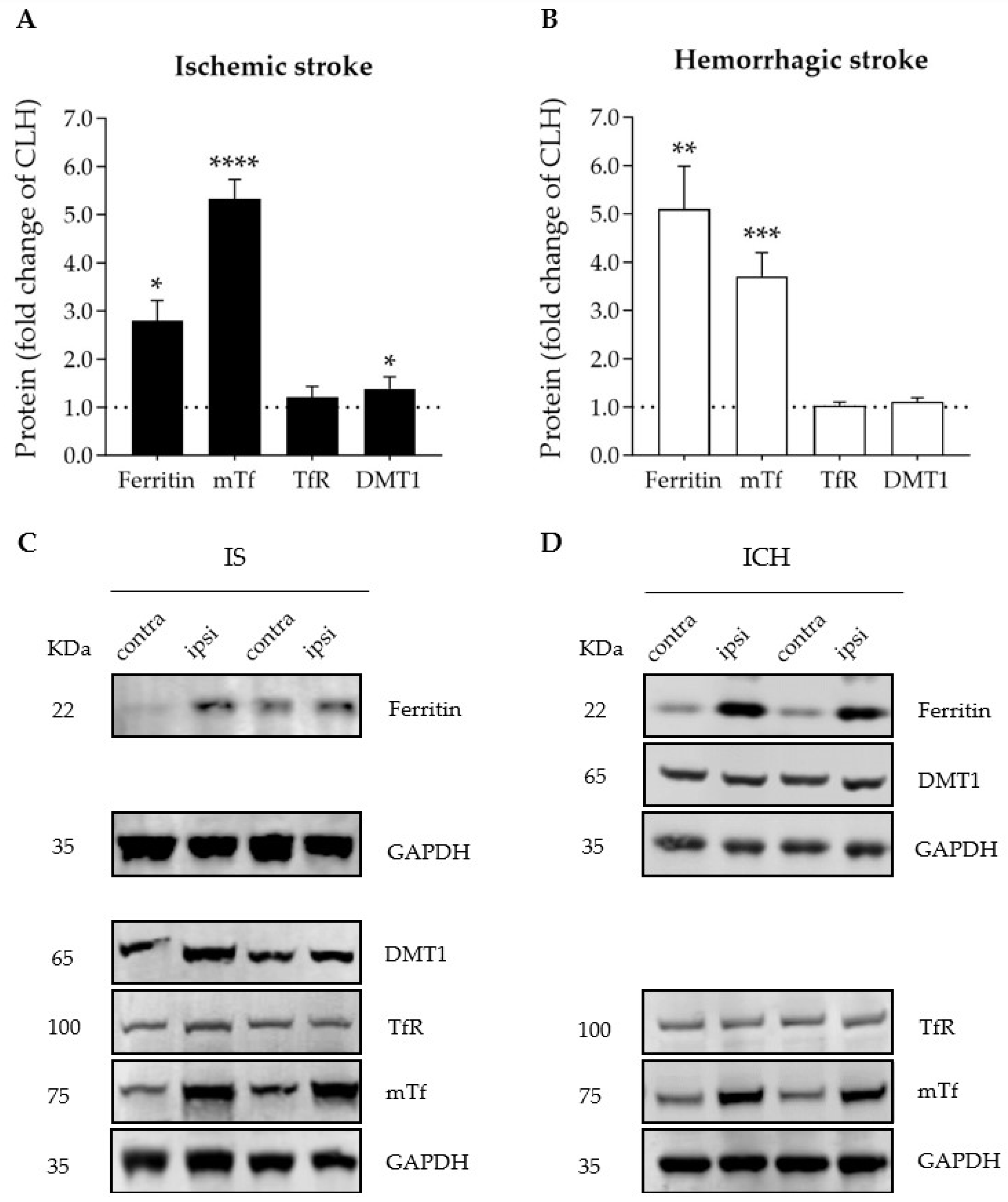
3.2. Ischemic and Hemorrhagic Stroke Share a Common Profile of Iron-Binding, Ferroptosis-Related Proteins in the Ipsilateral Brain Hemisphere 72 h after Stroke Induction
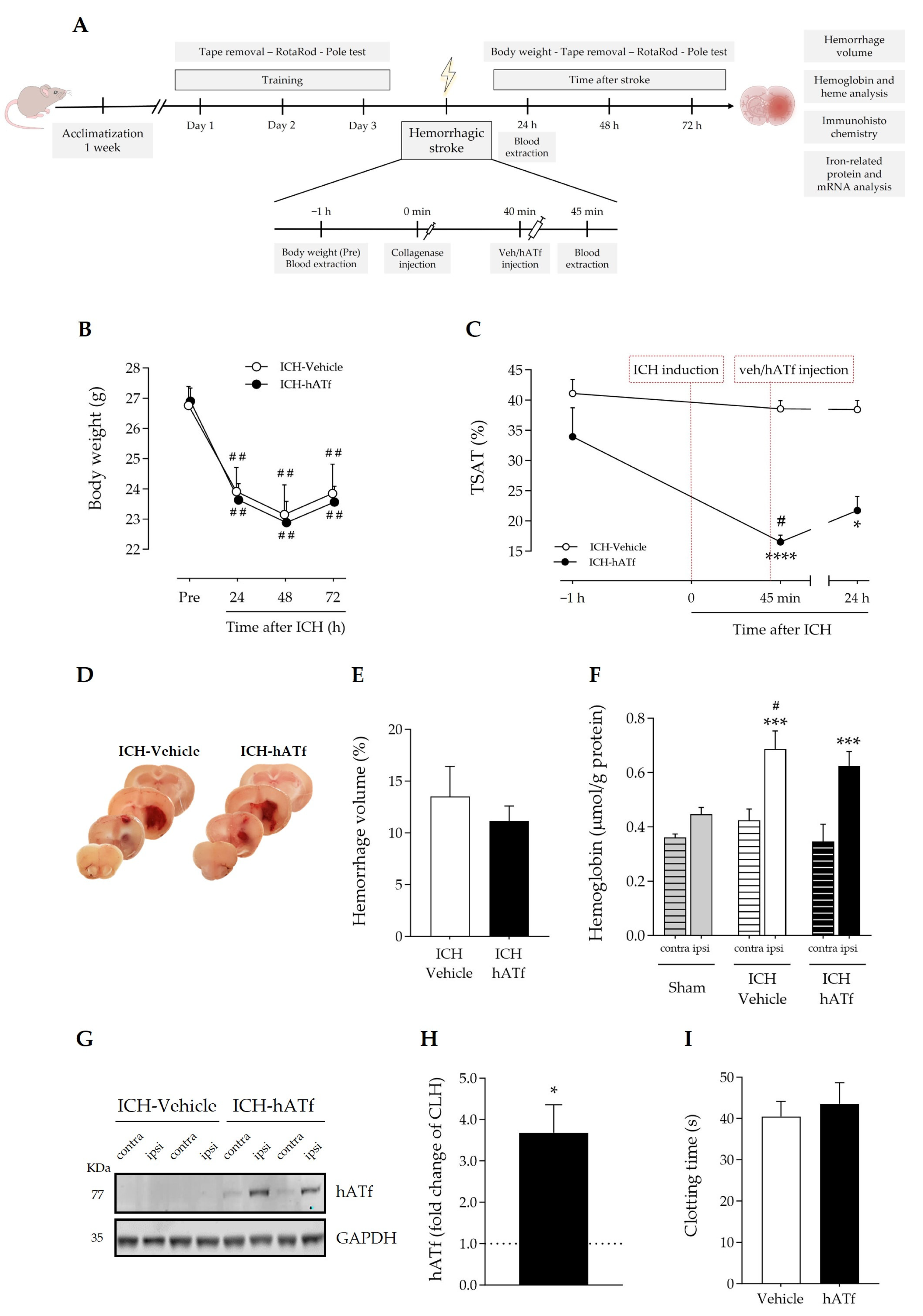
3.3. Early Peripheral Administration of hATf into the Bloodstream after ICH Induction Does Not Reduce Parenchymal Hemorrhage
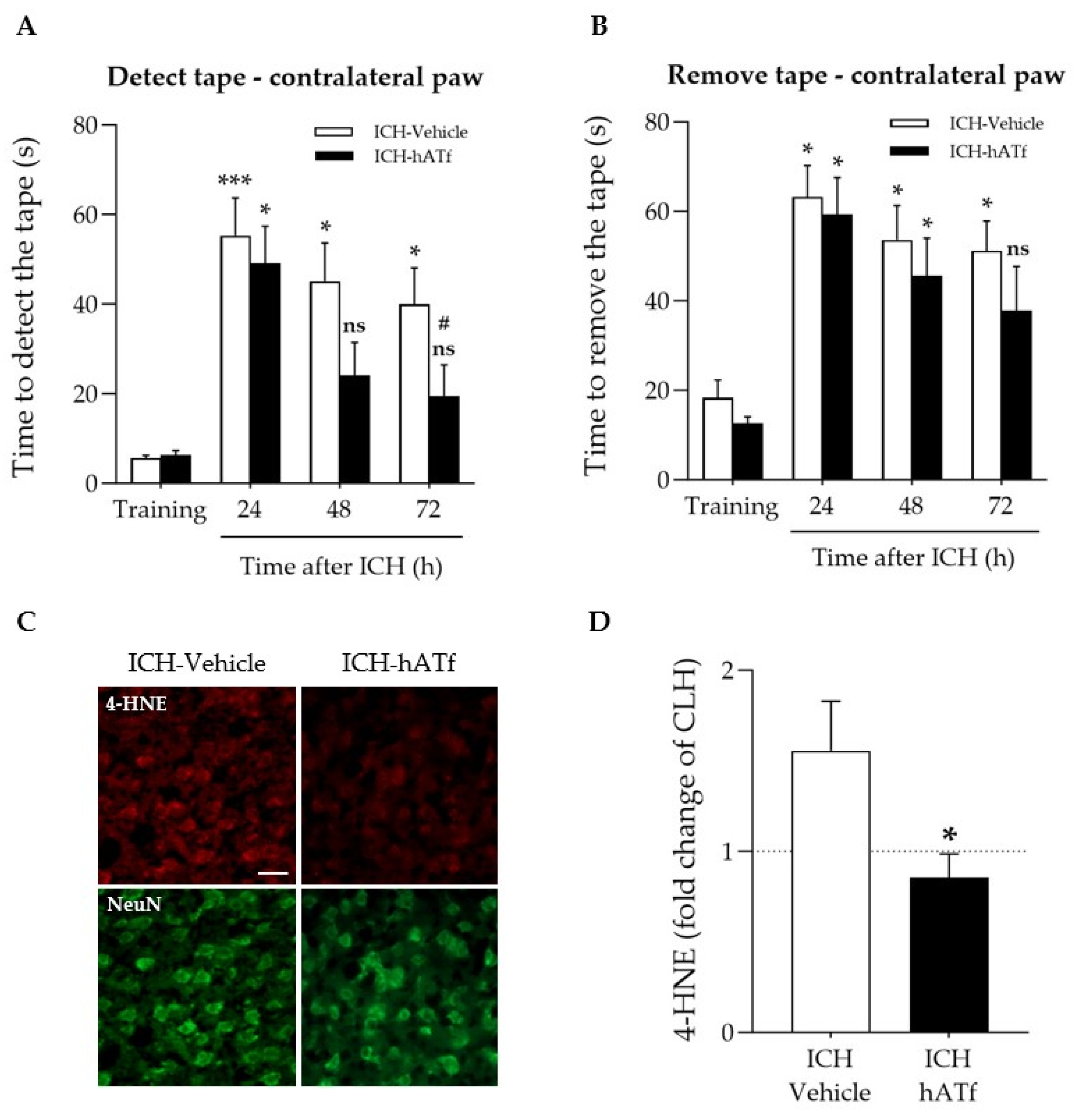
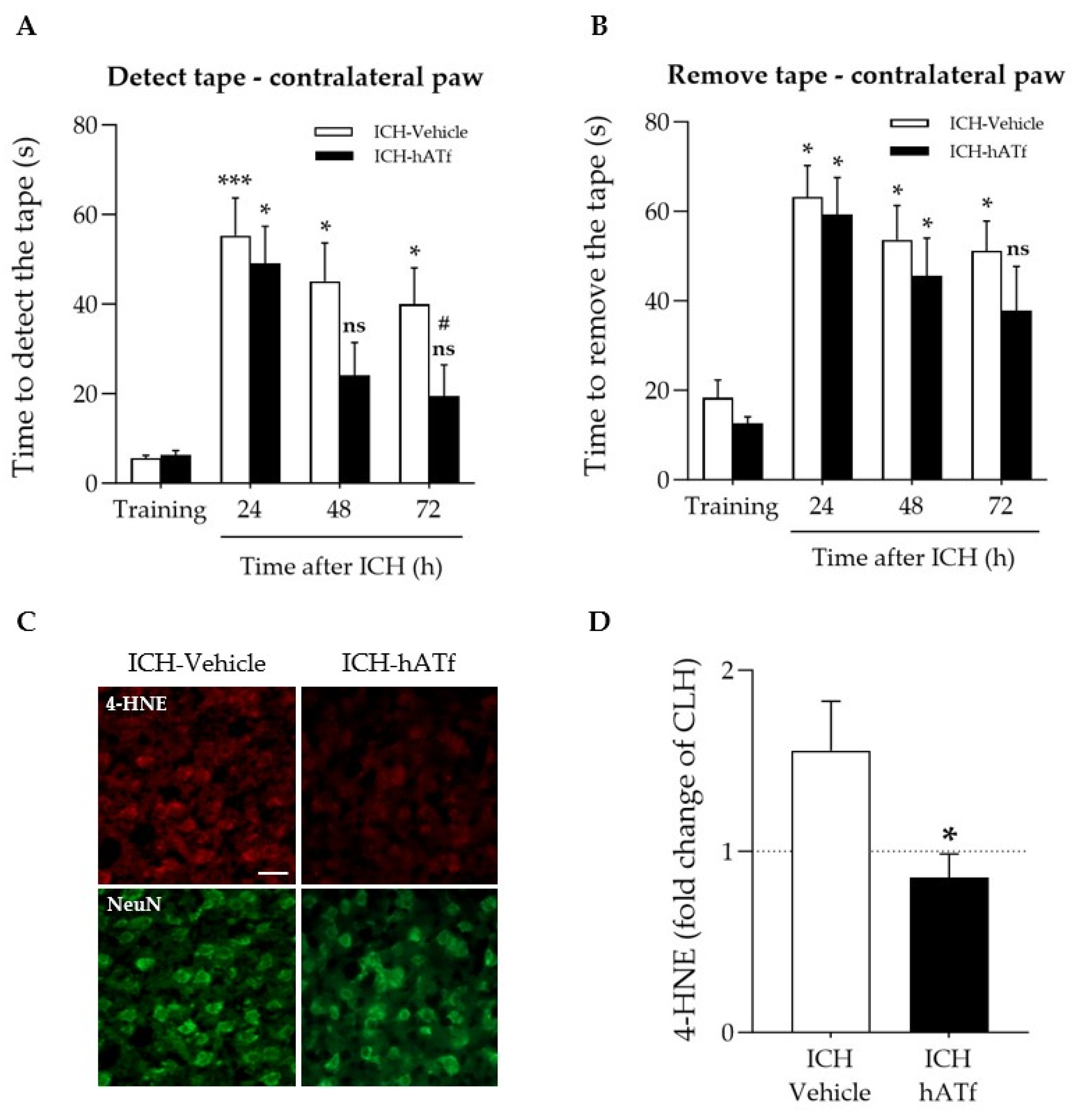
3.4. hATf Reduces ICH-Induced Sensorimotor Impairment in the Adhesive Tape Detection and Removal Test
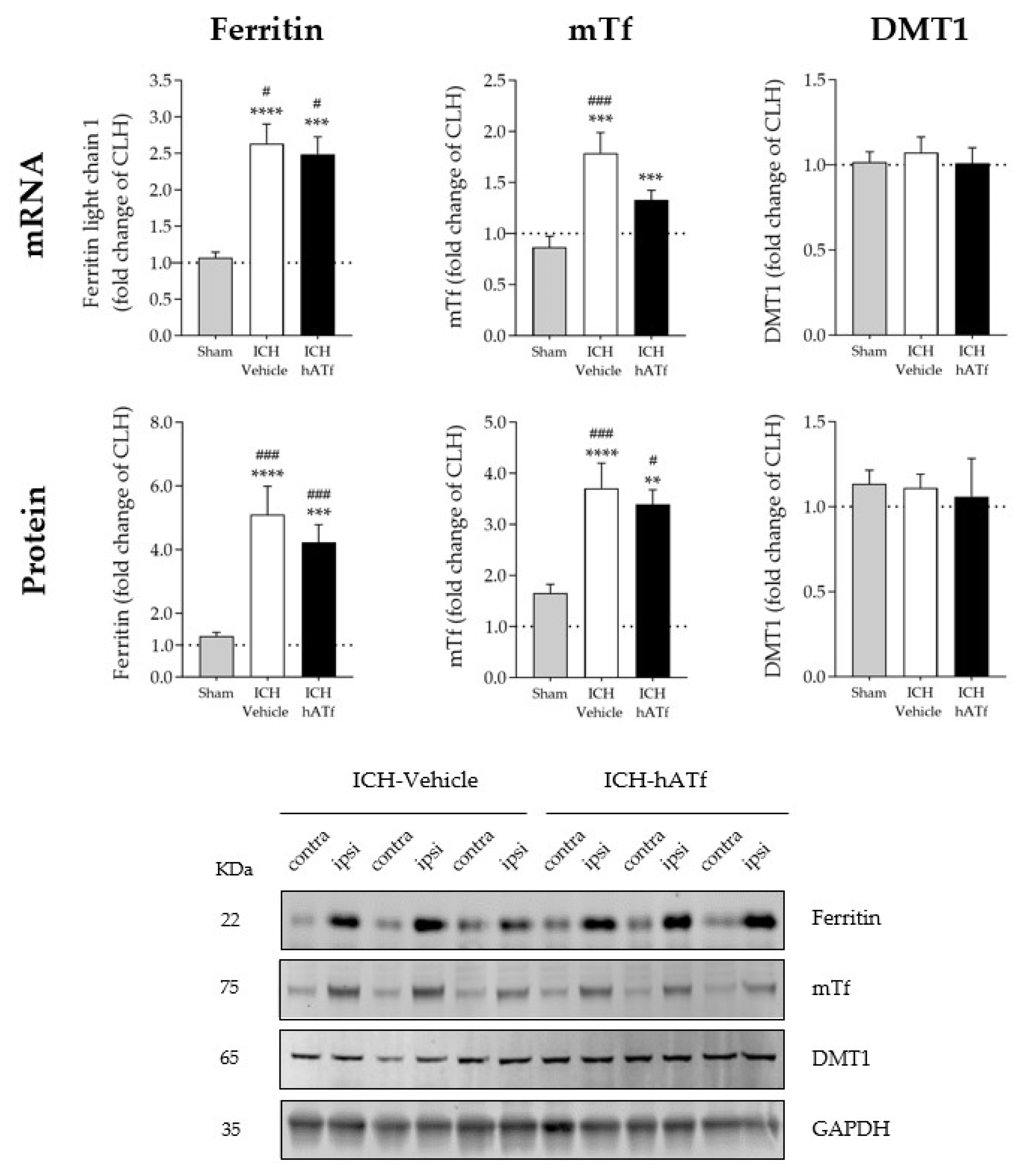
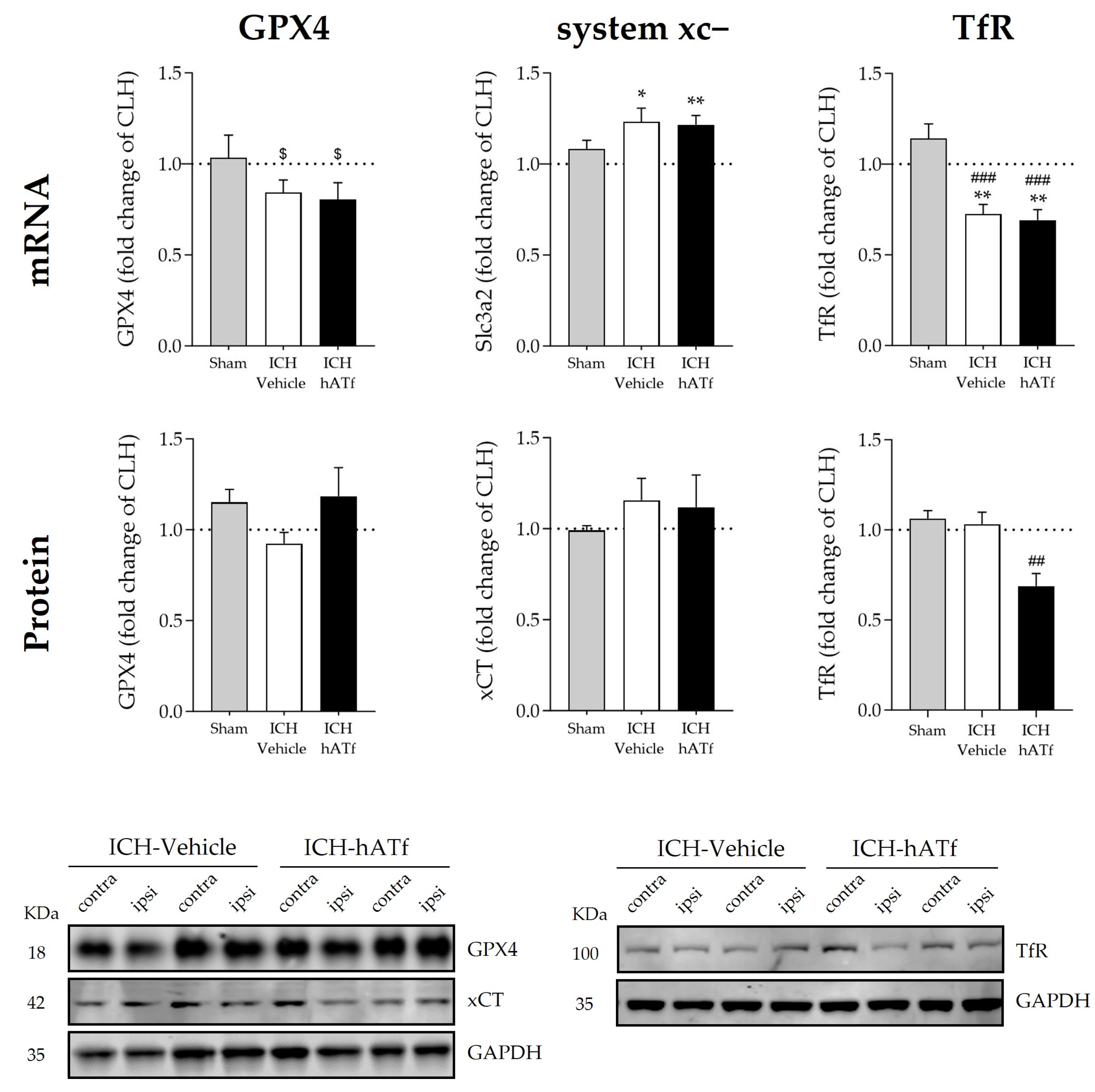
3.5. Effect of hATf on Iron Storage/Transport-Related mRNA and Protein Levels in ICH Mice
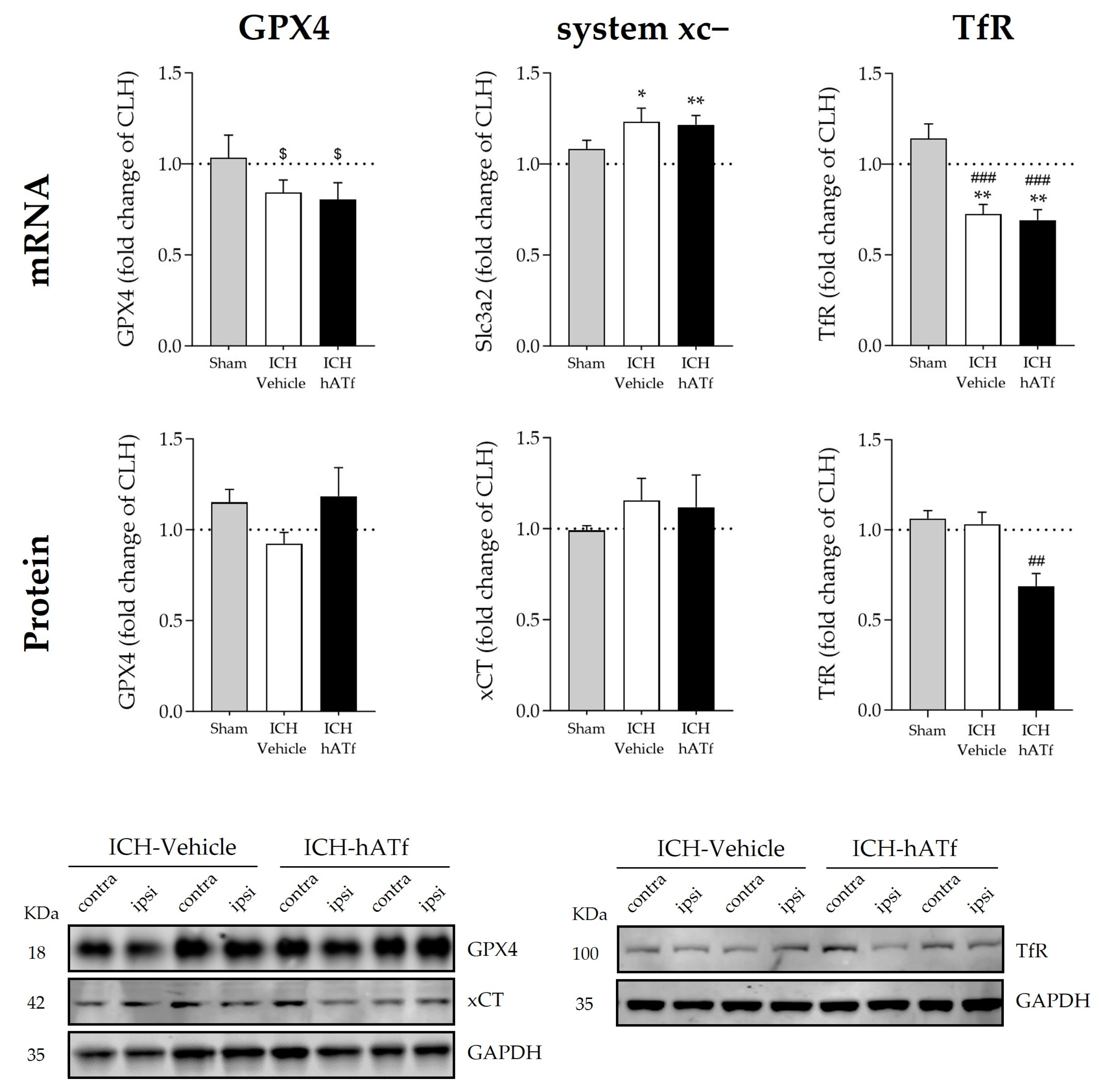
3.6. Effects of hATf on Classical Key Ferroptosis mRNA and Protein Levels in ICH Mice
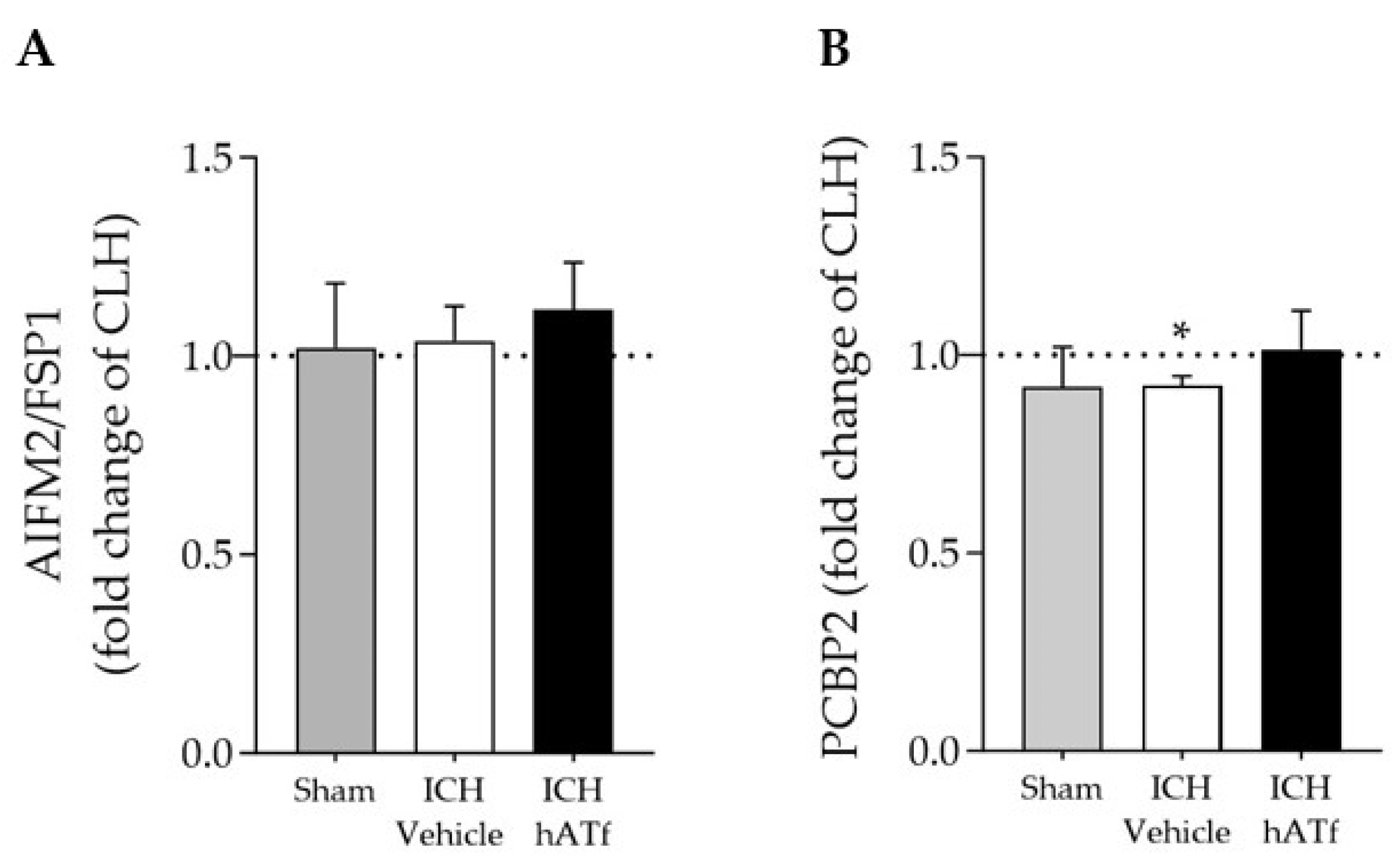
3.7. Effect of hATf on the mRNA Expression of Newly Reported Key Ferroptosis Players AIFM2/FSP1 and PCBP2 in ICH Mice
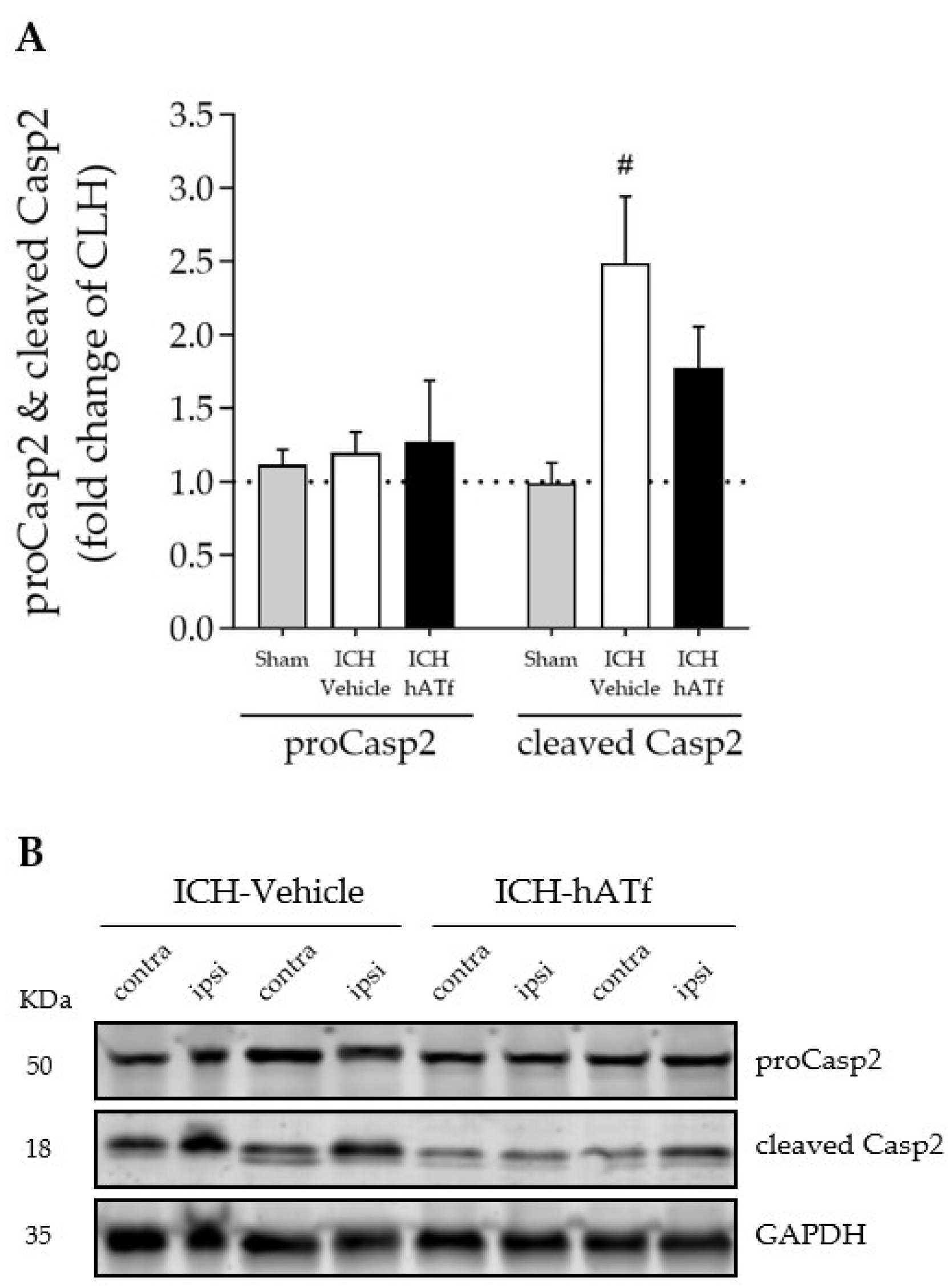
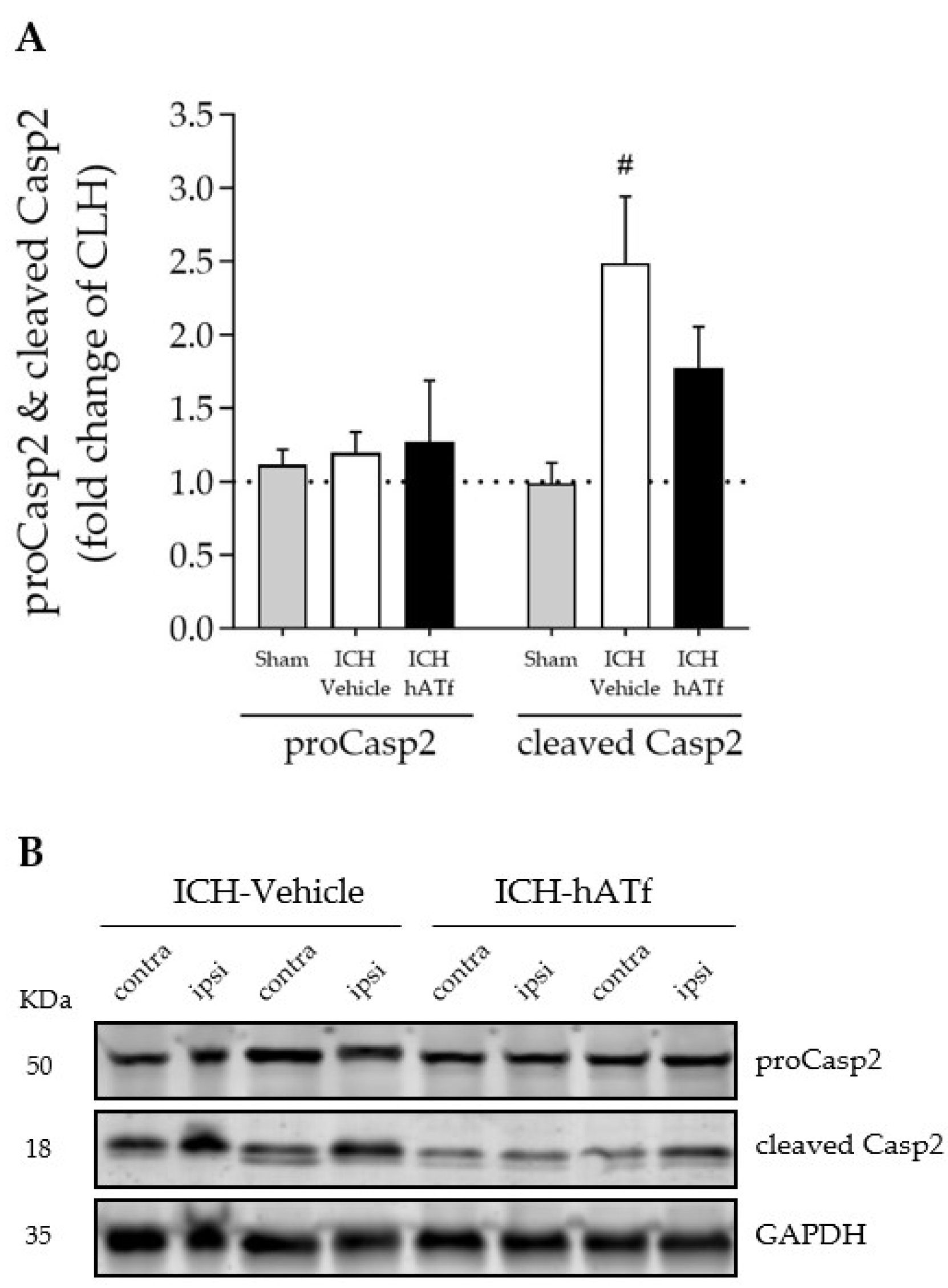
3.8. hATf Reduces ICH-Induced Caspase 2 Activation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanson, L.R.; Roeytenberg, A.; Martinez, P.M.; Coppes, V.G.; Sweet, D.C.; Rao, R.J.; Marti, D.L.; Hoekman, J.D.; Matthews, R.B.; Frey, W.H., 2nd; et al. Intranasal deferoxamine provides increased brain exposure and significant protection in rat ischemic stroke. J. Pharmacol. Exp. Ther. 2009, 330, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Freret, T.; Valable, S.; Chazalviel, L.; Saulnier, R.; Mackenzie, E.T.; Petit, E.; Bernaudin, M.; Boulouard, M.; Schumann-Bard, P. Delayed administration of deferoxamine reduces brain damage and promotes functional recovery after transient focal cerebral ischemia in the rat. Eur. J. Neurosci. 2006, 23, 1757–1765. [Google Scholar] [CrossRef] [PubMed]
- DeGregorio-Rocasolano, N.; Martí-Sistac, O.; Ponce, J.; Castelló-Ruiz, M.; Millán, M.; Guirao, V.; García-Yébenes, I.; Salom, J.B.; Ramos-Cabrer, E.A.; Lizasoain, I.; et al. Iron-Loaded transferrin (Tf) is detrimental whereas iron-free Tf confers protection against brain ischemia by modifying blood Tf saturation and subsequent neuronal damage. Redox Biol. 2018, 15, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Millán, M.; DeGregorio-Rocasolano, N.; Pérez de la Ossa, N.; Reverté, S.; Costa, J.; Giner, P.; Silva, Y.; Sobrino, T.; Rodríguez-Yáñez, M.; Nombela, F.; et al. Targeting Pro-Oxidant Iron with Deferoxamine as a Treatment for Ischemic Stroke: Safety and Optimal Dose Selection in a Randomized Clinical Trial. Antioxidants 2021, 10, 1270. [Google Scholar] [CrossRef]
- Chen-Roetling, J.; Chen, L.; Regan, R.F. Apotransferrin protects cortical neurons from hemoglobin toxicity. Neuropharmacology 2011, 60, 423–431. [Google Scholar] [CrossRef]
- Foster, L.; Robinson, L.; Yeatts, S.D.; Conwit, R.A.; Shehadah, A.; Lioutas, V.; Selim, M.; i-DEF Investigators. Effect of Deferoxamine on Trajectory of Recovery After Intracerebral Hemorrhage: A Post Hoc Analysis of the i-DEF Trial. Stroke 2022, 53, 2204–2210. [Google Scholar] [CrossRef]
- Wei, C.; Wang, J.; Foster, L.D.; Yeatts, S.D.; Moy, C.; Mocco, J.; Selim, M.; i-DEF Investigators. Effect of Deferoxamine on Outcome According to Baseline Hematoma Volume: A Post Hoc Analysis of the i-DEF Trial. Stroke 2022, 53, 1149–1156. [Google Scholar] [CrossRef]
- Xi, G.; Keep, R.F.; Hoff, J.T. Mechanisms of brain injury after intracerebral haemorrhage. Lancet Neurol. 2006, 5, 53–63. [Google Scholar] [CrossRef]
- Koeppen, A.H.; Dickson, A.C.; McEvoy, J.A. The cellular reactions to experimental intracerebral hemorrhage. J. Neurol. Sci. 1995, 134, 102–112. [Google Scholar] [CrossRef]
- Xi, G.; Keep, R.F.; Hoff, J.T. Erythrocytes and delayed brain edema formation following intracerebral hemorrhage in rats. J. Neurosurg. 1998, 89, 991–996. [Google Scholar] [CrossRef]
- Wu, J.; Hua, Y.; Keep, R.F.; Nakamura, T.; Hoff, J.T.; Xi, G. Iron and iron-handling proteins in the brain after intracerebral hemorrhage. Stroke 2003, 34, 2964–2969. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.P.; Xi, G.; Keep, R.F.; Hua, Y.; Nemoianu, A.; Hoff, J.T. Brain edema after experimental intracerebral hemorrhage: Role of hemoglobin degradation products. J. Neurosurg. 2002, 96, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Zille, M.; Karuppagounder, S.S.; Chen, Y.; Gough, P.J.; Bertin, J.; Finger, J.; Milner, T.A.; Jonas, E.A.; Ratan, R.R. Neuronal Death After Hemorrhagic Stroke In Vitro And In Vivo Shares Features of Ferroptosis and Necroptosis. Stroke 2017, 48, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Xi, G.; Hua, Y.; Bhasin, R.R.; Ennis, S.R.; Keep, R.F.; Hoff, J.T. Mechanisms of edema formation after intracerebral hemorrhage: Effects of extravasated red blood cells on blood flow and blood-brain barrier integrity. Stroke 2001, 32, 2932–2938. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, S.; Li, Q.; Sun, H.; Wang, H. Pharmacological Inhibition of Ferroptosis as a Therapeutic Target for Neurodegenerative Diseases and Strokes. Adv. Sci. 2023, 10, e2300325. [Google Scholar] [CrossRef]
- Duan, L.; Zhang, Y.; Yang, Y.; Su, S.; Zhou, L.; Lo, P.C.; Cai, J.; Qiao, Y.; Li, M.; Huang, S.; et al. Baicalin Inhibits Ferroptosis in Intracerebral Hemorrhage. Front. Pharmacol. 2021, 12, 629379. [Google Scholar] [CrossRef]
- Han, R.; Wan, J.; Han, X.; Ren, H.; Falck, J.R.; Munnuri, S.; Yang, Z.J.; Koehler, R.C. 20-HETE Participates in Intracerebral Hemorrhage-Induced Acute Injury by Promoting Cell Ferroptosis. Front. Neurol. 2021, 12, 763419. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, Y.; Yuan, S.; Zhang, P.; Zhang, J.; Li, H.; Li, X.; Shen, H.; Wang, Z.; Chen, G. Glutathione peroxidase 4 participates in secondary brain injury through mediating ferroptosis in a rat model of intracerebral hemorrhage. Brain Res. 2018, 1701, 112–125. [Google Scholar] [CrossRef]
- Guan, J.; Williams, C.; Gunning, M.; Mallard, C.; Gluckman, P. The effects of IGF-1 treatment after hypoxic-ischemic brain injury in adult rats. J. Cereb. Blood Flow Metab. 1993, 13, 609–616. [Google Scholar] [CrossRef]
- Klebe, D.; Iniaghe, L.; Burchell, S.; Reis, C.; Akyol, O.; Tang, J.; Zhang, J.H. Intracerebral hemorrhage in mice. Methods Mol. Biol. 2018, 1717, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Aliena-Valero, A.; Rius-Pérez, S.; Pérez, S.; Torregrosa, G.; Salom, J.B. Optimised lyophilisation-based method for different biomolecule single-extractions from the same rat brain sample: Suitability for RNA and protein expression analyses after ischemic stroke. J. Neurosci. Methods. 2019, 327, 108402. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Wee Yong, V.; Xue, M. Vildagliptin improves neurological function by inhibiting apoptosis and ferroptosis following intracerebral hemorrhage in mice. Neurosci. Lett. 2022, 776, 136579. [Google Scholar] [CrossRef] [PubMed]
- Byrne, S.L.; Mason, A.B. Human serum transferrin: A tale of two lobes. Urea gel and steady state fluorescence analysis of recombinant transferrins as a function of pH, time, and the soluble portion of the transferrin receptor. J. Biol. Inorg. Chem. 2009, 14, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, M.H.; Maitani, T. Differed preferential iron-binding lobe in human transferrin depending on the presence of bicarbonate detected by HPLC/high-resolution inductively coupled plasma mass spectrometry. Biochim. Biophys. Acta 2000, 1523, 182–188. [Google Scholar] [CrossRef]
- Agarwal, R. Transferrin saturation with intravenous irons: An in vitro study. Kidney Int. 2004, 66, 1139–1144. [Google Scholar] [CrossRef]
- Feng, H.; Schorpp, K.; Jin, J.; Yozwiak, C.E.; Hoffstrom, B.G.; Decker, A.M.; Rajbhandari, P.; Stokes, M.E.; Bender, H.G.; Csuka, J.M.; et al. Transferrin Receptor Is a Specific Ferroptosis Marker. Cell Rep. 2020, 30, 3411–3423.e7. [Google Scholar] [CrossRef]
- Wang, G.; Hu, W.; Tang, Q.; Wang, L.; Sun, X.G.; Chen, Y.; Yin, Y.; Xue, F.; Sun, Z. Effect Comparison of Both Iron Chelators on Outcomes, Iron Deposit, and Iron Transporters After Intracerebral Hemorrhage in Rats. Mol. Neurobiol. 2016, 53, 3576–3585. [Google Scholar] [CrossRef]
- Yang, G.; Qian, C.; Zhang, C.; Bao, Y.; Liu, M.Y.; Jiang, F.; Li, W.; Liu, Y.; Ke, Y.; Qian, Z.M. Hepcidin attenuates the iron-mediated secondary neuronal injury after intracerebral hemorrhage in rats. Transl. Res. 2021, 229, 53–68. [Google Scholar] [CrossRef]
- Hatakeyama, T.; Okauchi, M.; Hua, Y.; Keep, R.F.; Xi, G. Deferoxamine reduces neuronal death and hematoma lysis after intracerebral hemorrhage in aged rats. Transl. Stroke Res. 2013, 4, 546–553. [Google Scholar] [CrossRef]
- Ding, H.; Yan, C.Z.; Shi, H.; Zhao, Y.S.; Chang, S.Y.; Yu, P.; Wu, W.S.; Zhao, C.Y.; Chang, Y.Z.; Duan, X.L. Hepcidin is involved in iron regulation in the ischemic brain. PLoS ONE 2011, 6, e25324. [Google Scholar] [CrossRef] [PubMed]
- Selim, M.; Foster, L.D.; Moy, C.S.; Xi, G.; Hill, M.D.; Morgenstern, L.B.; Greenberg, S.M.; James, M.L.; Singh, V.; Clark, W.M.; et al. Deferoxamine mesylate in patients with intracerebral haemorrhage (i-DEF): A multicentre, randomised, placebo-controlled, double-blind phase 2 trial. Lancet Neurol. 2019, 18, 428–438. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, Z.; Fang, M.; Han, Y.; Wang, G.; Wang, S.; Xue, M.; Li, Y.; Zhang, L.; Wu, J.; et al. Transferrin plays a central role in coagulation balance by interacting with clotting factors. Cell Res. 2020, 30, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Fang, M.; Cheng, R.; Zhang, Z.; Wang, Y.; Shen, C.; Han, Y.; Lu, Q.; Du, Y.; Liu, Y.; et al. Iron-deficiency and estrogen are associated with ischemic stroke by up-regulating transferrin to induce hypercoagulability. Circ. Res. 2020, 127, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.N.; Zhang, X.; Sergueeva, A.I.; Miasnikova, G.Y.; Ganz, T.; Prchal, J.T.; Gordeuk, V.R. Increased transferrin protects from thrombosis in Chuvash erythrocytosis. Am. J. Hematol. 2023, 98, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Doll, S.; Freitas, F.P.; Shah, R.; Aldrovandi, M.; da Silva, M.C.; Ingold, I.; Goya Grocin, A.; Xavier da Silva, T.N.; Panzilius, E.; Scheel, C.H.; et al. FSP1 is a glutathione-independent ferroptosis suppressor. Nature 2019, 575, 693–698. [Google Scholar] [CrossRef]
- Bersuker, K.; Hendricks, J.M.; Li, Z.; Magtanong, L.; Ford, B.; Tang, P.H.; Roberts, M.A.; Tong, B.; Maimone, T.J.; Zoncu, R.; et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature 2019, 575, 688–692. [Google Scholar] [CrossRef]
- Yue, L.; Luo, Y.; Jiang, L.; Sekido, Y.; Toyokuni, S. PCBP2 knockdown promotes ferroptosis in malignant mesothelioma. Pathol. Int. 2022, 72, 242–251. [Google Scholar] [CrossRef]
- Jin, J.; Schorpp, K.; Samaga, D.; Unger, K.; Hadian, K.; Stockwell, B.R. Machine Learning Classifies Ferroptosis and Apoptosis Cell Death Modalities with TfR1 Immunostaining. ACS Chem. Biol. 2022, 17, 654–660. [Google Scholar] [CrossRef]
- Tang, L.J.; Zhou, Y.J.; Xiong, X.M.; Li, N.S.; Zhang, J.J.; Luo, X.J.; Peng, J. Ubiquitin-specific protease 7 promotes ferroptosis via activation of the p53/TfR1 pathway in the rat hearts after ischemia/reperfusion. Free Radic. Biol. Med. 2021, 162, 339–352. [Google Scholar] [CrossRef]
- Du, L.; Wu, Y.; Jia, Q.; Li, J.; Li, Y.; Ma, H.; Fan, Z.; Guo, X.; Li, L.; Peng, Y.; et al. Autophagy Suppresses Ferroptosis by Degrading TFR1 to Alleviate Cognitive Dysfunction in Mice with SAE. Cell Mol. Neurobiol. 2023, 43, 3605–3622. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Hu, L.; Sun, T.; Wang, X.; Xiao, Z.; Shen, D.; Wu, W.; Luo, Z.; Wei, C.; Wang, X.; et al. H3K9 trimethylation dictates neuronal ferroptosis through repressing Tfr1. J. Cereb. Blood Flow Metab. 2023, 43, 1365–1381. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, W.L.; Shen, L.J.; Jiang, T.T.; Xia, J.J.; You, D.L.; Hu, S.Y.; Wang, L.; Wu, X. Crocin Alleviates Intracerebral Hemorrhage-Induced Neuronal Ferroptosis by Facilitating Nrf2 Nuclear Translocation. Neurotox. Res. 2022, 40, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Wang, L.; Wu, G. Mechanism of RNA-binding protein Lin28 in neuronal ferroptosis after intracerebral haemorrhage. Folia Neuropathol. 2022, 60, 35–47. [Google Scholar] [CrossRef]
- Zhang, H.; Wen, M.; Chen, J.; Yao, C.; Lin, X.; Lin, Z.; Ru, J.; Zhuge, Q.; Yang, S. Pyridoxal Isonicotinoyl Hydrazone Improves Neurological Recovery by Attenuating Ferroptosis and Inflammation in Cerebral Hemorrhagic Mice. Biomed. Res. Int. 2021, 2021, 9916328. [Google Scholar] [CrossRef]
- Neag, M.-A.; Mitre, A.-O.; Catinean, A.; Mitre, C.-I. An Overview on the Mechanisms of Neuroprotection and Neurotoxicity of Isoflurane and Sevoflurane in Experimental Studies. Brain Res. Bull. 2020, 165, 281–289. [Google Scholar] [CrossRef]
- Liu, P.; Yuan, J.; Feng, Y.; Chen, X.; Wang, G.; Zhao, L. Ferroptosis contributes to isoflurane-induced neurotoxicity and learning and memory impairment. Cell Death Discov. 2021, 7, 72. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, X.; Zhong, J.; Wang, S.; Zhang, C.; Li, M.; Hu, Q.; Wang, S.; Chen, L.; Chen, W.; et al. Dexpramipexole Attenuates White Matter Injury to Facilitate Locomotion and Motor Coordination Recovery via Reducing Ferroptosis after Intracerebral Hemorrhage. Oxid. Med. Cell. Longev. 2022, 2022, 6160701. [Google Scholar] [CrossRef]
- Iwai, M.; Kajino, T.; Nakatochi, M.; Yanagisawa, K.; Hosono, Y.; Isomura, H.; Shimada, Y.; Suzuki, M.; Taguchi, A.; Takahashi, T. Long non-coding RNA TILR constitutively represses TP53 and apoptosis in lung cancer. Oncogene 2023, 42, 364–373. [Google Scholar] [CrossRef]
- Mao, X.; Liu, J.; Chen, C.; Zhang, W.; Qian, R.; Chen, X.; Lu, H.; Ge, J.; Zhao, C.; Zhang, D.; et al. PCBP2 Modulates Neural Apoptosis and Astrocyte Proliferation After Spinal Cord Injury. Neurochem. Res. 2016, 41, 2401–2414. [Google Scholar] [CrossRef]
- Rosato-Siri, M.V.; Marziali, L.N.; Mattera, V.; Correale, J.; Pasquini, J.M. Combination therapy of apo-transferrin and thyroid hormones enhances remyelination. Glia 2021, 69, 151–164. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Serran, A.; Ordoño, J.; DeGregorio-Rocasolano, N.; Melià-Sorolla, M.; Odendaal, K.; Martí-Sistac, O.; Gasull, T. Targeting Pro-Oxidant Iron with Exogenously Administered Apotransferrin Provides Benefits Associated with Changes in Crucial Cellular Iron Gate Protein TfR in a Model of Intracerebral Hemorrhagic Stroke in Mice. Antioxidants 2023, 12, 1945. https://doi.org/10.3390/antiox12111945
García-Serran A, Ordoño J, DeGregorio-Rocasolano N, Melià-Sorolla M, Odendaal K, Martí-Sistac O, Gasull T. Targeting Pro-Oxidant Iron with Exogenously Administered Apotransferrin Provides Benefits Associated with Changes in Crucial Cellular Iron Gate Protein TfR in a Model of Intracerebral Hemorrhagic Stroke in Mice. Antioxidants. 2023; 12(11):1945. https://doi.org/10.3390/antiox12111945
Chicago/Turabian StyleGarcía-Serran, Alexia, Jesús Ordoño, Núria DeGregorio-Rocasolano, Marc Melià-Sorolla, Karla Odendaal, Octavi Martí-Sistac, and Teresa Gasull. 2023. "Targeting Pro-Oxidant Iron with Exogenously Administered Apotransferrin Provides Benefits Associated with Changes in Crucial Cellular Iron Gate Protein TfR in a Model of Intracerebral Hemorrhagic Stroke in Mice" Antioxidants 12, no. 11: 1945. https://doi.org/10.3390/antiox12111945
APA StyleGarcía-Serran, A., Ordoño, J., DeGregorio-Rocasolano, N., Melià-Sorolla, M., Odendaal, K., Martí-Sistac, O., & Gasull, T. (2023). Targeting Pro-Oxidant Iron with Exogenously Administered Apotransferrin Provides Benefits Associated with Changes in Crucial Cellular Iron Gate Protein TfR in a Model of Intracerebral Hemorrhagic Stroke in Mice. Antioxidants, 12(11), 1945. https://doi.org/10.3390/antiox12111945