
Adding High-Intensity Interval Training to Classical Resistance Training Does Not Impede the Recovery from Inactivity-Induced Leg Muscle Weakness

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Volunteers and Interventions
2.2. Training after Unloading Period
2.3. Muscle Function
2.4. Ultrasonography
2.5. Blood Lipid Peroxidation and Total Antioxidant Capacity
2.6. Statistical Analysis
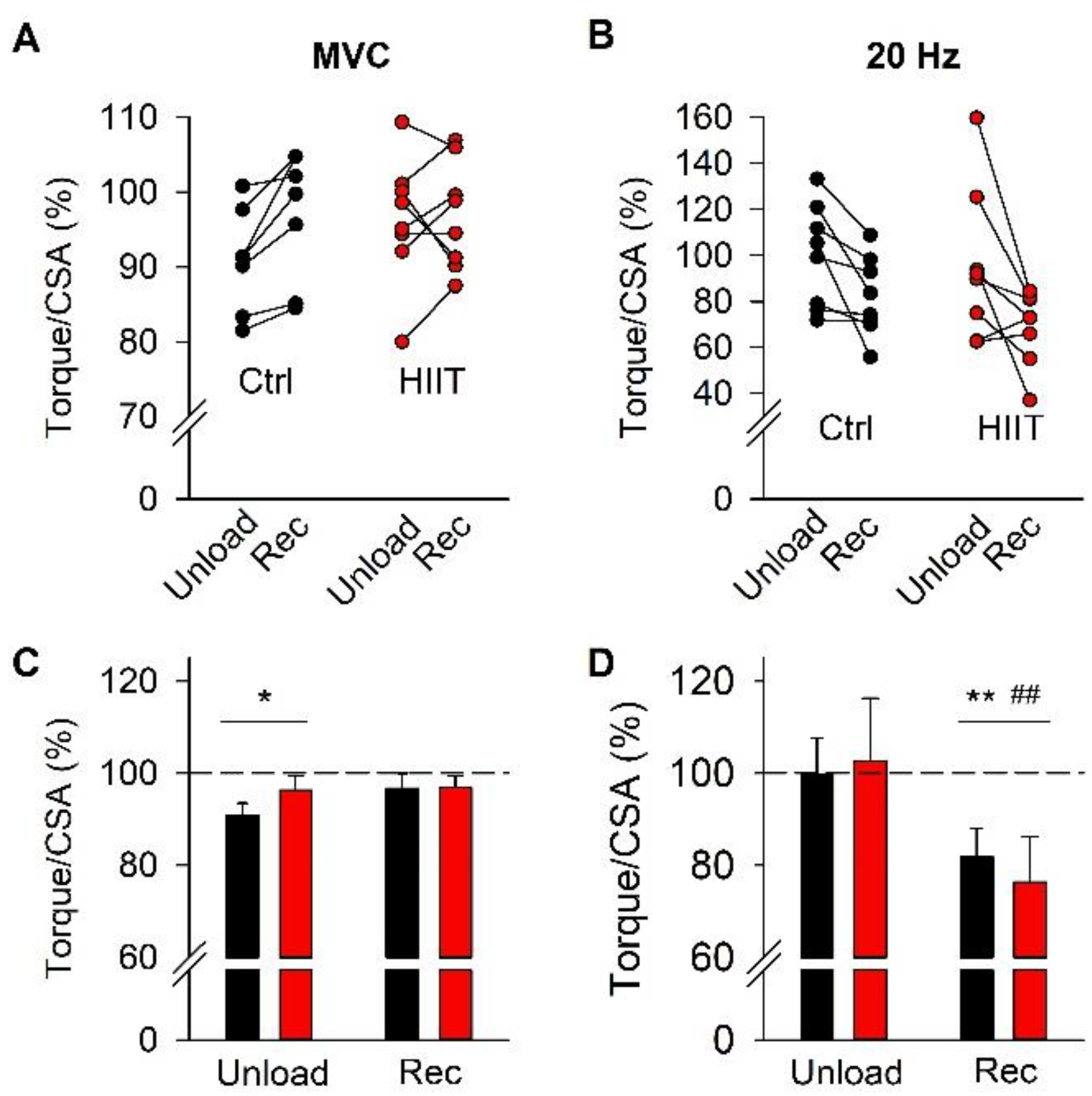
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swallow, E.B.; Reyes, D.; Hopkinson, N.S.; Man, W.D.-C.; Porcher, R.; Cetti, E.J.; Moore, A.J.; Moxham, J.; Polkey, M.I. Quadriceps Strength Predicts Mortality in Patients with Moderate to Severe Chronic Obstructive Pulmonary Disease. Thorax 2007, 62, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Dirks, M.L.; van Loon, L.J.C. Skeletal Muscle Atrophy during Short-Term Disuse: Implications for Age-Related Sarcopenia. Ageing Res. Rev. 2013, 12, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, H.; Deminice, R.; Yoshihara, T.; Powers, S.K. Mitochondrial Dysfunction Induces Muscle Atrophy during Prolonged Inactivity: A Review of the Causes and Effects. Arch. Biochem. Biophys. 2019, 662, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Memme, J.M.; Slavin, M.; Moradi, N.; Hood, D.A. Mitochondrial Bioenergetics and Turnover during Chronic Muscle Disuse. Int. J. Mol. Sci. 2021, 22, 5179. [Google Scholar] [CrossRef]
- Powers, S.K. Can Antioxidants Protect against Disuse Muscle Atrophy? Sports Med. 2014, 44 (Suppl. 2), S155–S165. [Google Scholar] [CrossRef]
- Cheng, A.J.; Yamada, T.; Rassier, D.E.; Andersson, D.C.; Westerblad, H.; Lanner, J.T. Reactive Oxygen/Nitrogen Species and Contractile Function in Skeletal Muscle during Fatigue and Recovery. J. Physiol. 2016, 594, 5149–5160. [Google Scholar] [CrossRef]
- Lamb, G.D.; Westerblad, H. Acute Effects of Reactive Oxygen and Nitrogen Species on the Contractile Function of Skeletal Muscle. J. Physiol. 2011, 589, 2119–2127. [Google Scholar] [CrossRef]
- Powers, S.K.; Deminice, R.; Ozdemir, M.; Yoshihara, T.; Bomkamp, M.P.; Hyatt, H. Exercise-Induced Oxidative Stress: Friend or Foe? J. Sport Health Sci. 2020, 9, 415–425. [Google Scholar] [CrossRef]
- Smuder, A.J.; Min, K.; Hudson, M.B.; Kavazis, A.N.; Kwon, O.-S.; Nelson, W.B.; Powers, S.K. Endurance Exercise Attenuates Ventilator-Induced Diaphragm Dysfunction. J. Appl. Physiol. (1985) 2012, 112, 501–510. [Google Scholar] [CrossRef]
- Paulsen, G.; Hamarsland, H.; Cumming, K.T.; Johansen, R.E.; Hulmi, J.J.; Børsheim, E.; Wiig, H.; Garthe, I.; Raastad, T. Vitamin C and E Supplementation Alters Protein Signalling after a Strength Training Session, but Not Muscle Growth during 10 Weeks of Training. J. Physiol. 2014, 592, 5391–5408. [Google Scholar] [CrossRef]
- Paulsen, G.; Cumming, K.T.; Holden, G.; Hallén, J.; Rønnestad, B.R.; Sveen, O.; Skaug, A.; Paur, I.; Bastani, N.E.; Østgaard, H.N.; et al. Vitamin C and E Supplementation Hampers Cellular Adaptation to Endurance Training in Humans: A Double-Blind, Randomised, Controlled Trial. J. Physiol. 2014, 592, 1887–1901. [Google Scholar] [CrossRef]
- Wyckelsma, V.L.; Venckunas, T.; Brazaitis, M.; Gastaldello, S.; Snieckus, A.; Eimantas, N.; Baranauskiene, N.; Subocius, A.; Skurvydas, A.; Pääsuke, M.; et al. Vitamin C and E Treatment Blunts Sprint Interval Training-Induced Changes in Inflammatory Mediator-, Calcium-, and Mitochondria-Related Signaling in Recreationally Active Elderly Humans. Antioxidants 2020, 9, 879. [Google Scholar] [CrossRef] [PubMed]
- Larsen, F.J.; Schiffer, T.A.; Ørtenblad, N.; Zinner, C.; Morales-Alamo, D.; Willis, S.J.; Calbet, J.A.; Holmberg, H.-C.; Boushel, R. High-Intensity Sprint Training Inhibits Mitochondrial Respiration through Aconitase Inactivation. FASEB J. 2016, 30, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Place, N.; Ivarsson, N.; Venckunas, T.; Neyroud, D.; Brazaitis, M.; Cheng, A.J.; Ochala, J.; Kamandulis, S.; Girard, S.; Volungevičius, G.; et al. Ryanodine Receptor Fragmentation and Sarcoplasmic Reticulum Ca2+ Leak after One Session of High-Intensity Interval Exercise. Proc. Natl. Acad. Sci. USA 2015, 112, 15492–15497. [Google Scholar] [CrossRef] [PubMed]
- Gibala, M.J.; Little, J.P.; Macdonald, M.J.; Hawley, J.A. Physiological Adaptations to Low-Volume, High-Intensity Interval Training in Health and Disease. J. Physiol. 2012, 590, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Kessler, H.S.; Sisson, S.B.; Short, K.R. The Potential for High-Intensity Interval Training to Reduce Cardiometabolic Disease Risk. Sports Med. 2012, 42, 489–509. [Google Scholar] [CrossRef]
- Ramos, J.S.; Dalleck, L.C.; Tjonna, A.E.; Beetham, K.S.; Coombes, J.S. The Impact of High-Intensity Interval Training versus Moderate-Intensity Continuous Training on Vascular Function: A Systematic Review and Meta-Analysis. Sports Med. 2015, 45, 679–692. [Google Scholar] [CrossRef]
- Steinz, M.M.; Santos-Alves, E.; Lanner, J.T. Skeletal Muscle Redox Signaling in Rheumatoid Arthritis. Clin. Sci. (Lond.) 2020, 134, 2835–2850. [Google Scholar] [CrossRef]
- Murlasits, Z.; Kneffel, Z.; Thalib, L. The Physiological Effects of Concurrent Strength and Endurance Training Sequence: A Systematic Review and Meta-Analysis. J. Sports Sci. 2018, 36, 1212–1219. [Google Scholar] [CrossRef]
- Ribeiro, J.; Afonso, J.; Camões, M.; Sarmento, H.; Sá, M.; Lima, R.; Oliveira, R.; Clemente, F.M. Methodological Characteristics, Physiological and Physical Effects, and Future Directions for Combined Training in Soccer: A Systematic Review. Healthcare 2021, 9, 1075. [Google Scholar] [CrossRef]
- Baar, K. Training for Endurance and Strength: Lessons from Cell Signaling. Med. Sci. Sports Exerc. 2006, 38, 1939–1944. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Lundby, C.; Cotter, J.D.; Burke, L.M. Maximizing Cellular Adaptation to Endurance Exercise in Skeletal Muscle. Cell Metab. 2018, 27, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, D.L.; Silva, L.A.; Tromm, C.B.; da Rosa, G.L.; Silveira, P.C.L.; de Souza, C.T.; Latini, A.; Pinho, R.A. Impact of Different Resistance Training Protocols on Muscular Oxidative Stress Parameters. Appl. Physiol. Nutr. Metab. 2012, 37, 1239–1246. [Google Scholar] [CrossRef]
- Hendrickse, P.W.; Venckunas, T.; Platkevicius, J.; Kairaitis, R.; Kamandulis, S.; Snieckus, A.; Stasiulis, A.; Vitkiene, J.; Subocius, A.; Degens, H. Endurance Training-Induced Increase in Muscle Oxidative Capacity without Loss of Muscle Mass in Younger and Older Resistance-Trained Men. Eur. J. Appl. Physiol. 2021, 121, 3161–3172. [Google Scholar] [CrossRef]
- Schumann, M.; Feuerbacher, J.F.; Sünkeler, M.; Freitag, N.; Rønnestad, B.R.; Doma, K.; Lundberg, T.R. Compatibility of Concurrent Aerobic and Strength Training for Skeletal Muscle Size and Function: An Updated Systematic Review and Meta-Analysis. Sports Med. 2022, 52, 601–612. [Google Scholar] [CrossRef]
- Tsitkanou, S.; Spengos, K.; Stasinaki, A.-N.; Zaras, N.; Bogdanis, G.; Papadimas, G.; Terzis, G. Effects of High-Intensity Interval Cycling Performed after Resistance Training on Muscle Strength and Hypertrophy. Scand. J. Med. Sci. Sports 2017, 27, 1317–1327. [Google Scholar] [CrossRef]
- Perry, B.D.; Wyckelsma, V.L.; Murphy, R.M.; Steward, C.H.; Anderson, M.; Levinger, I.; Petersen, A.C.; McKenna, M.J. Dissociation between Short-Term Unloading and Resistance Training Effects on Skeletal Muscle Na+,K+-ATPase, Muscle Function, and Fatigue in Humans. J. Appl. Physiol. (1985) 2016, 121, 1074–1086. [Google Scholar] [CrossRef]
- Craig, C.L.; Marshall, A.L.; Sjöström, M.; Bauman, A.E.; Booth, M.L.; Ainsworth, B.E.; Pratt, M.; Ekelund, U.; Yngve, A.; Sallis, J.F.; et al. International Physical Activity Questionnaire: 12-Country Reliability and Validity. Med. Sci. Sports Exerc. 2003, 35, 1381–1395. [Google Scholar] [CrossRef]
- Earle, T.B.; Earle, R.W. Essentials of Strength Training and Conditionin, 3rd ed.; Human Kinetics: Champaign, IL, USA, 2008; ISBN 978-0-7360-5803-2. [Google Scholar]
- Tang, Q.; Su, Y.-W.; Xian, C.J. Determining Oxidative Damage by Lipid Peroxidation Assay in Rat Serum. Bio Protoc. 2019, 9, e3263. [Google Scholar] [CrossRef]
- Franchi, M.V.; Reeves, N.D.; Narici, M.V. Skeletal Muscle Remodeling in Response to Eccentric vs. Concentric Loading: Morphological, Molecular, and Metabolic Adaptations. Front. Physiol. 2017, 8, 447. [Google Scholar] [CrossRef]
- Reeves, N.D.; Maganaris, C.N.; Narici, M.V. Ultrasonographic Assessment of Human Skeletal Muscle Size. Eur. J. Appl. Physiol. 2004, 91, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Appell, H.J.; Duarte, J.A.; Soares, J.M. Supplementation of Vitamin E May Attenuate Skeletal Muscle Immobilization Atrophy. Int. J. Sports Med. 1997, 18, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Dalla Libera, L.; Ravara, B.; Gobbo, V.; Tarricone, E.; Vitadello, M.; Biolo, G.; Vescovo, G.; Gorza, L. A Transient Antioxidant Stress Response Accompanies the Onset of Disuse Atrophy in Human Skeletal Muscle. J. Appl. Physiol. (1985) 2009, 107, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Miura, M.; Itokawa, Y. Oxidative Stress in Skeletal Muscle Atrophied by Immobilization. Acta Physiol. Scand. 1991, 142, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Fang, Q.; Xu, T.; Wu, C.; Xu, L.; Wang, L.; Yang, X.; Yu, S.; Zhang, Q.; Ding, F.; et al. Mechanistic Role of Reactive Oxygen Species and Therapeutic Potential of Antioxidants in Denervation- or Fasting-Induced Skeletal Muscle Atrophy. Front. Physiol. 2018, 9, 215. [Google Scholar] [CrossRef]
- Kamandulis, S.; de Souza Leite, F.; Hernández, A.; Katz, A.; Brazaitis, M.; Bruton, J.D.; Venckunas, T.; Masiulis, N.; Mickeviciene, D.; Eimantas, N.; et al. Prolonged Force Depression after Mechanically Demanding Contractions Is Largely Independent of Ca2+ and Reactive Oxygen Species. FASEB J. 2017, 31, 4809–4820. [Google Scholar] [CrossRef]
- Muanjai, P.; Mickevicius, M.; Sniečkus, A.; Sipavičienė, S.; Satkunskiene, D.; Kamandulis, S.; Jones, D.A. Low Frequency Fatigue and Changes in Muscle Fascicle Length Following Eccentric Exercise of the Knee Extensors. Exp. Physiol. 2020, 105, 502–510. [Google Scholar] [CrossRef]
- Schlittler, M.; Neyroud, D.; Tanga, C.; Zanou, N.; Kamandulis, S.; Skurvydas, A.; Kayser, B.; Westerblad, H.; Place, N.; Andersson, D.C. Three Weeks of Sprint Interval Training Improved High-Intensity Cycling Performance and Limited Ryanodine Receptor Modifications in Recreationally Active Human Subjects. Eur. J. Appl. Physiol. 2019, 119, 1951–1958. [Google Scholar] [CrossRef]
- Skurvydas, A.; Mamkus, G.; Kamandulis, S.; Dudoniene, V.; Valanciene, D.; Westerblad, H. Mechanisms of Force Depression Caused by Different Types of Physical Exercise Studied by Direct Electrical Stimulation of Human Quadriceps Muscle. Eur. J. Appl. Physiol. 2016, 116, 2215–2224. [Google Scholar] [CrossRef]
- Vera-Ibañez, A.; Colomer-Poveda, D.; Romero-Arenas, S.; Viñuela-García, M.; Márquez, G. Neural Adaptations after Short-Term Wingate-Based High-Intensity Interval Training. J. Musculoskelet Neuronal. Interact. 2017, 17, 275–282. [Google Scholar]
- Kristoffersen, M.; Sandbakk, Ø.; Rønnestad, B.R.; Gundersen, H. Comparison of Short-Sprint and Heavy Strength Training on Cycling Performance. Front. Physiol. 2019, 10, 1132. [Google Scholar] [CrossRef] [PubMed]
- MacInnis, M.J.; Zacharewicz, E.; Martin, B.J.; Haikalis, M.E.; Skelly, L.E.; Tarnopolsky, M.A.; Murphy, R.M.; Gibala, M.J. Superior Mitochondrial Adaptations in Human Skeletal Muscle after Interval Compared to Continuous Single-Leg Cycling Matched for Total Work. J. Physiol. 2017, 595, 2955–2968. [Google Scholar] [CrossRef] [PubMed]
- Andersen, G.; Heje, K.; Buch, A.E.; Vissing, J. High-Intensity Interval Training in Facioscapulohumeral Muscular Dystrophy Type 1: A Randomized Clinical Trial. J. Neurol. 2017, 264, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Andonian, B.J.; Bartlett, D.B.; Huebner, J.L.; Willis, L.; Hoselton, A.; Kraus, V.B.; Kraus, W.E.; Huffman, K.M. Effect of High-Intensity Interval Training on Muscle Remodeling in Rheumatoid. Arthritis Res. Ther. 2018, 20, 283. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ctrl Group (n = 8) | HIIT Group (n = 8) | |
|---|---|---|
| Age (years) | 25.0 ± 1.6 | 26.9 ± 2.2 |
| Height (cm) | 180.9 ± 1.9 | 184.5 ± 3.2 |
| Body mass (kg) | 81.9 ± 3.5 | 84.4 ± 4.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venckunas, T.; Brazaitis, M.; Snieckus, A.; Mickevicius, M.; Eimantas, N.; Subocius, A.; Mickeviciene, D.; Westerblad, H.; Kamandulis, S. Adding High-Intensity Interval Training to Classical Resistance Training Does Not Impede the Recovery from Inactivity-Induced Leg Muscle Weakness. Antioxidants 2023, 12, 16. https://doi.org/10.3390/antiox12010016
Venckunas T, Brazaitis M, Snieckus A, Mickevicius M, Eimantas N, Subocius A, Mickeviciene D, Westerblad H, Kamandulis S. Adding High-Intensity Interval Training to Classical Resistance Training Does Not Impede the Recovery from Inactivity-Induced Leg Muscle Weakness. Antioxidants. 2023; 12(1):16. https://doi.org/10.3390/antiox12010016
Chicago/Turabian StyleVenckunas, Tomas, Marius Brazaitis, Audrius Snieckus, Mantas Mickevicius, Nerijus Eimantas, Andrejus Subocius, Dalia Mickeviciene, Håkan Westerblad, and Sigitas Kamandulis. 2023. "Adding High-Intensity Interval Training to Classical Resistance Training Does Not Impede the Recovery from Inactivity-Induced Leg Muscle Weakness" Antioxidants 12, no. 1: 16. https://doi.org/10.3390/antiox12010016
APA StyleVenckunas, T., Brazaitis, M., Snieckus, A., Mickevicius, M., Eimantas, N., Subocius, A., Mickeviciene, D., Westerblad, H., & Kamandulis, S. (2023). Adding High-Intensity Interval Training to Classical Resistance Training Does Not Impede the Recovery from Inactivity-Induced Leg Muscle Weakness. Antioxidants, 12(1), 16. https://doi.org/10.3390/antiox12010016