
Potential Neurotoxic Effects of Glioblastoma-Derived Exosomes in Primary Cultures of Cerebellar Neurons via Oxidant Stress and Glutathione Depletion

 , , ,
, , ,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Glioblastoma Cell Lines and Primary Neuron Cell Culture
2.2. Exosome Isolation and Primary Neuron Cell Treatment
2.3. MTT Assay
2.4. Total Oxidant Status (TOS) and Total Antioxidant Status (TAS)
2.5. Lactate Dehydrogenase (LDH) Assay
2.6. Glutathione (GSH) Assay
2.7. Statistical Analyses
3. Results
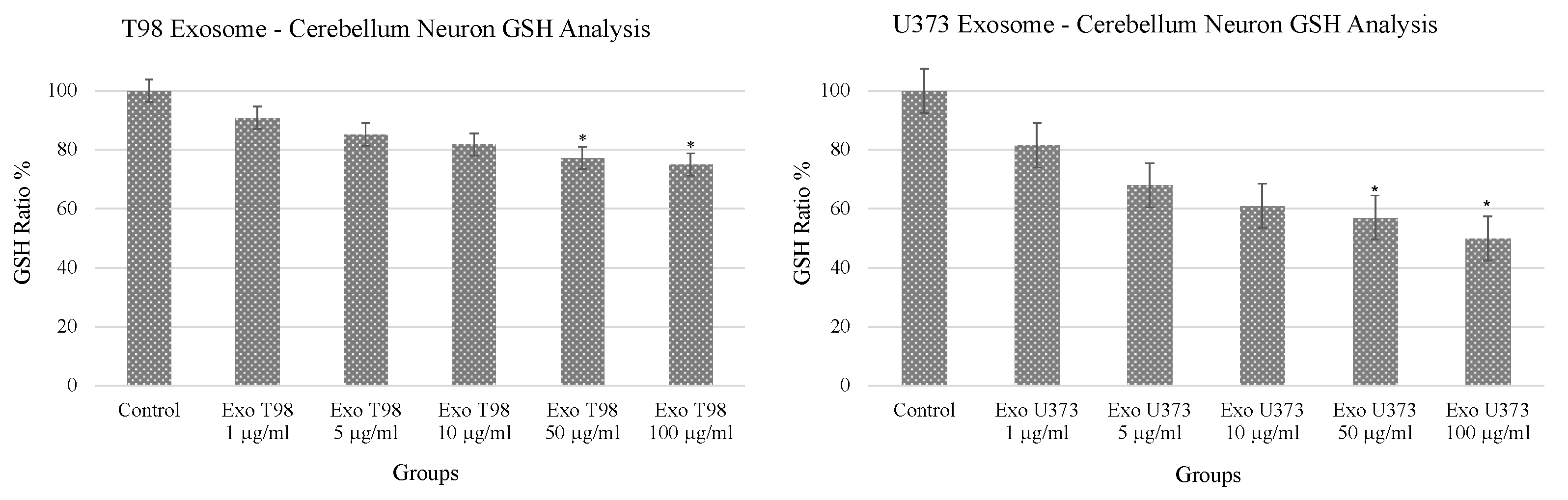
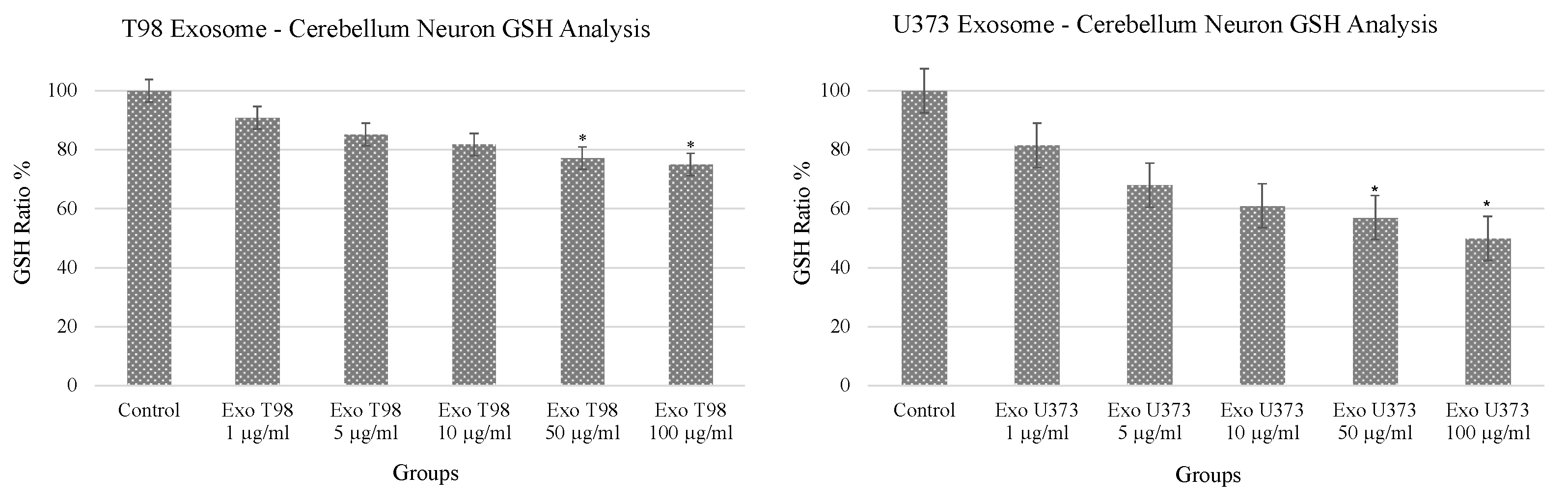
3.1. Alteration of Glutathione (GSH) Levels in Cerebellar Neurons after GBM-Derived Exosomes Treatment
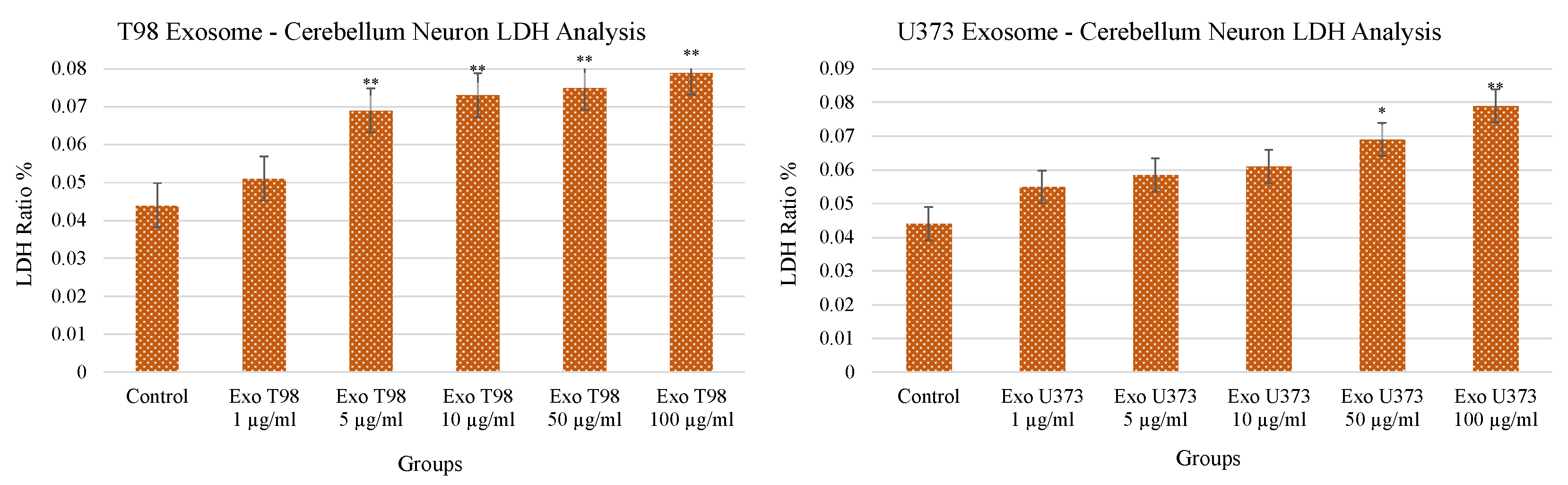
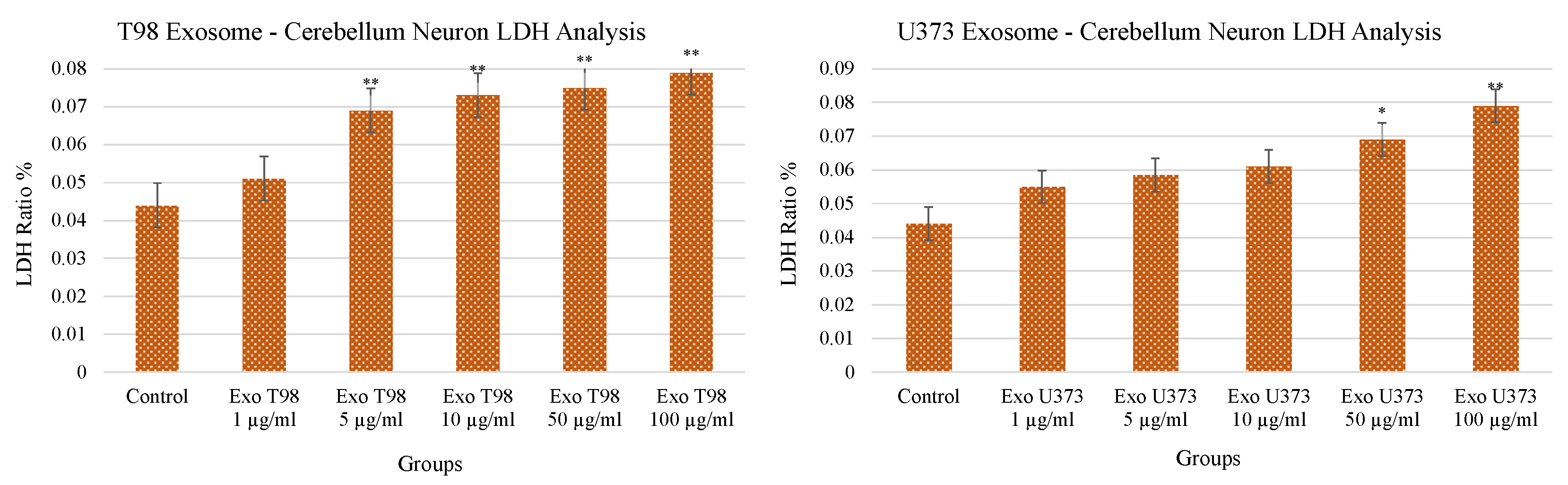
3.2. Lactate Dehydrogenase (LDH) Levels in Cerebellar Neurons after GBM-Derived Exosomes Treatment
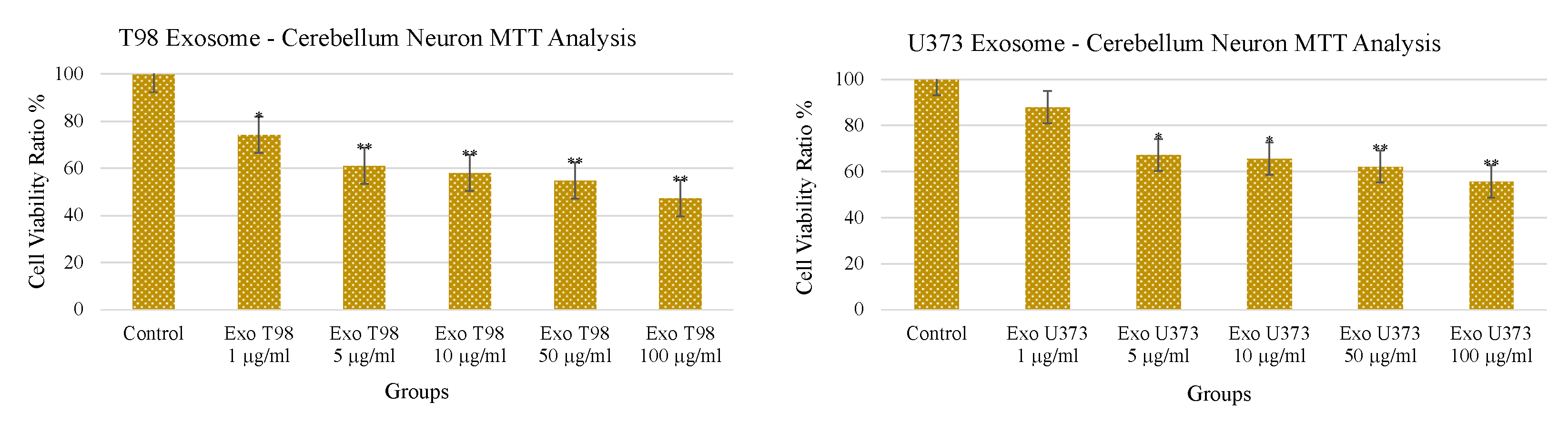
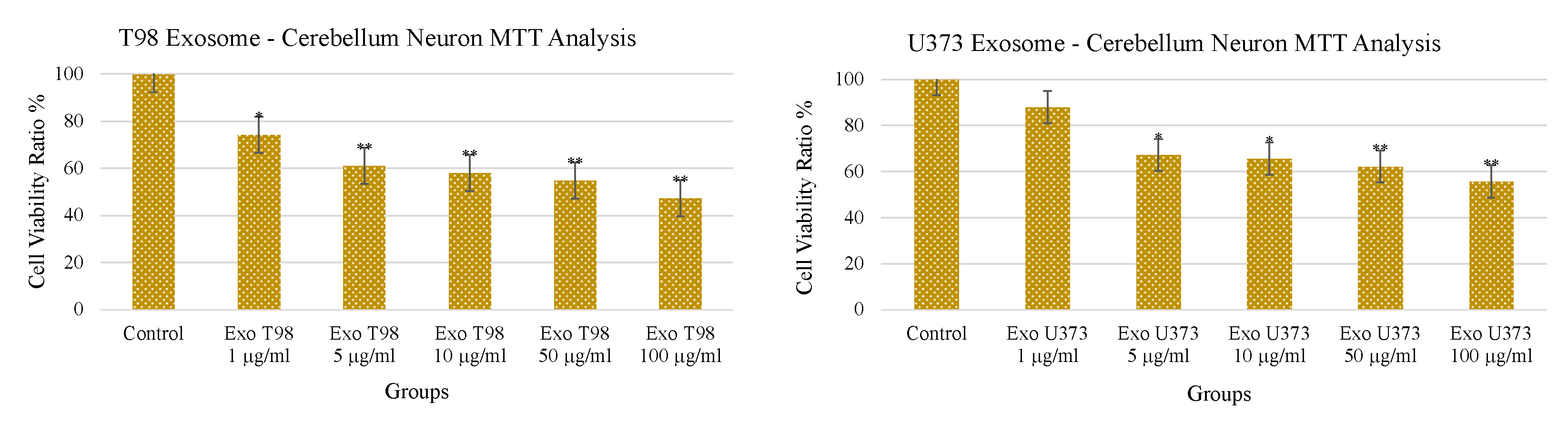
3.3. Treatment with GBM-Derived Exosomes Reduces Cell Viability of Cerebellar Neurons In Vitro
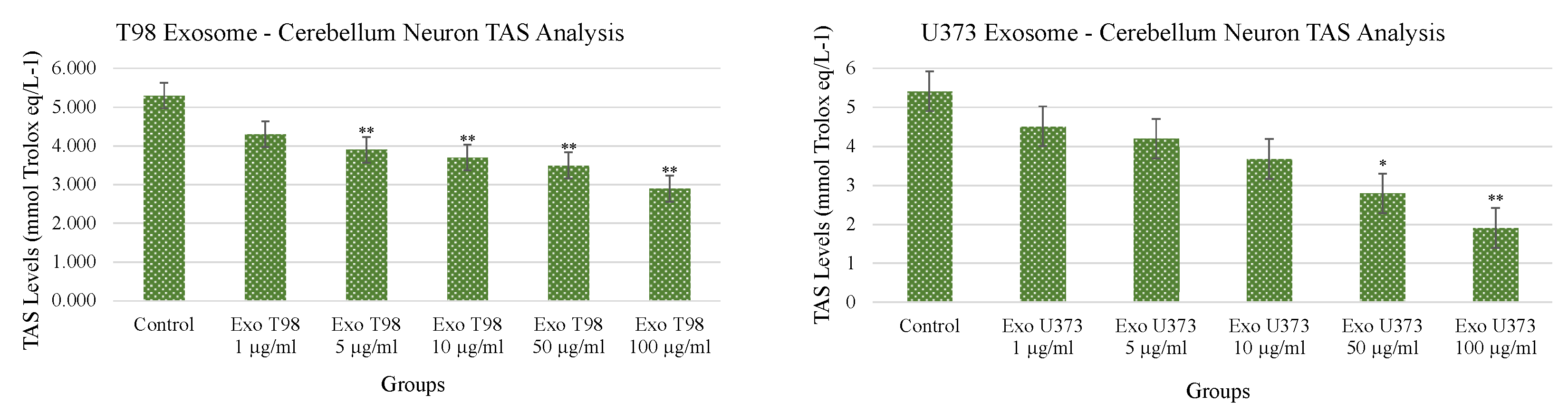
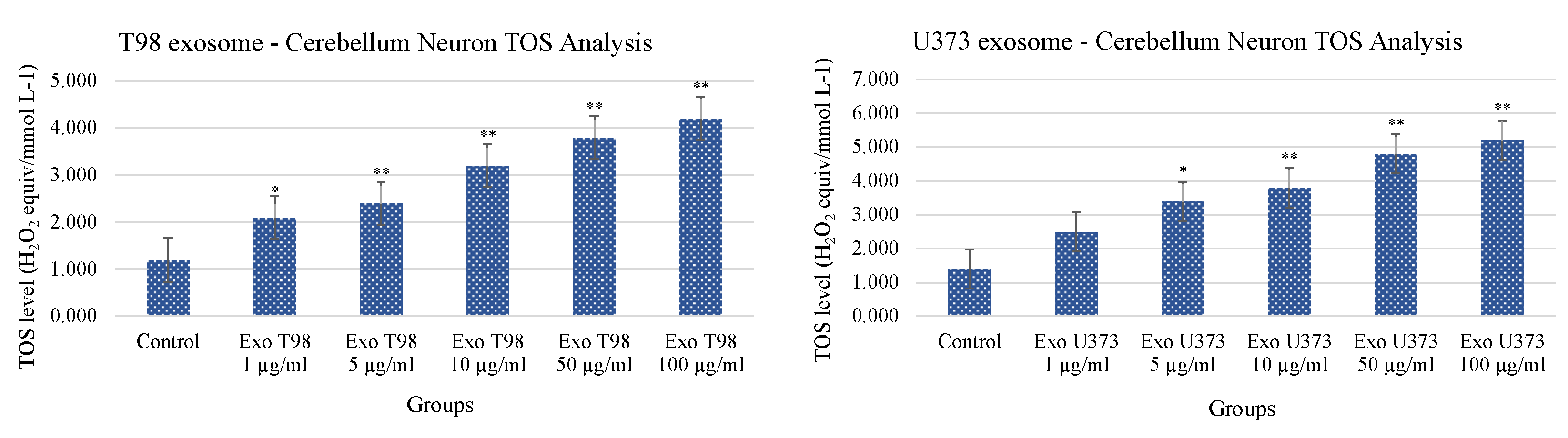
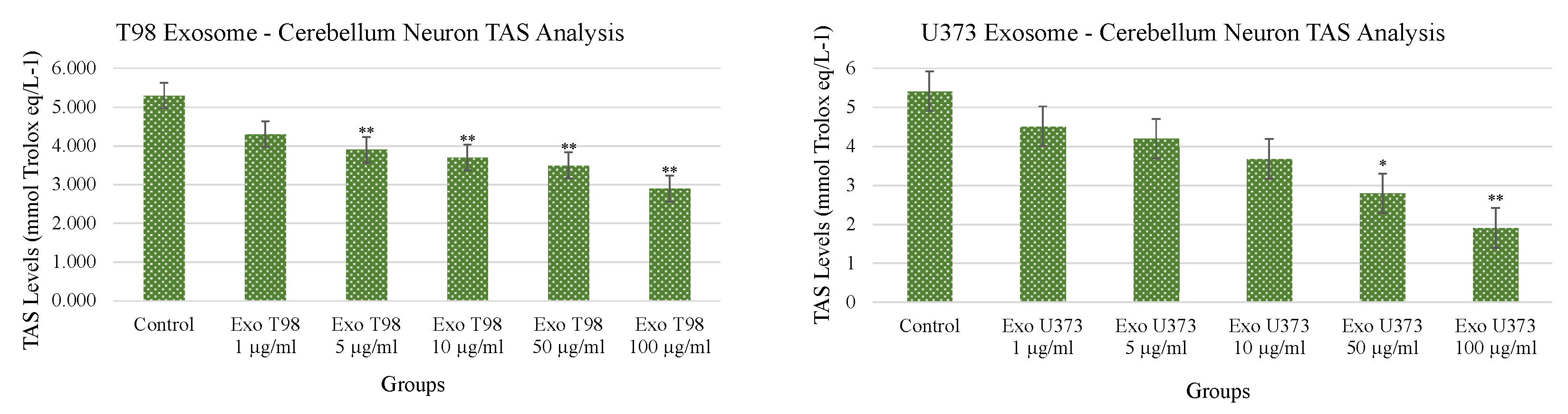
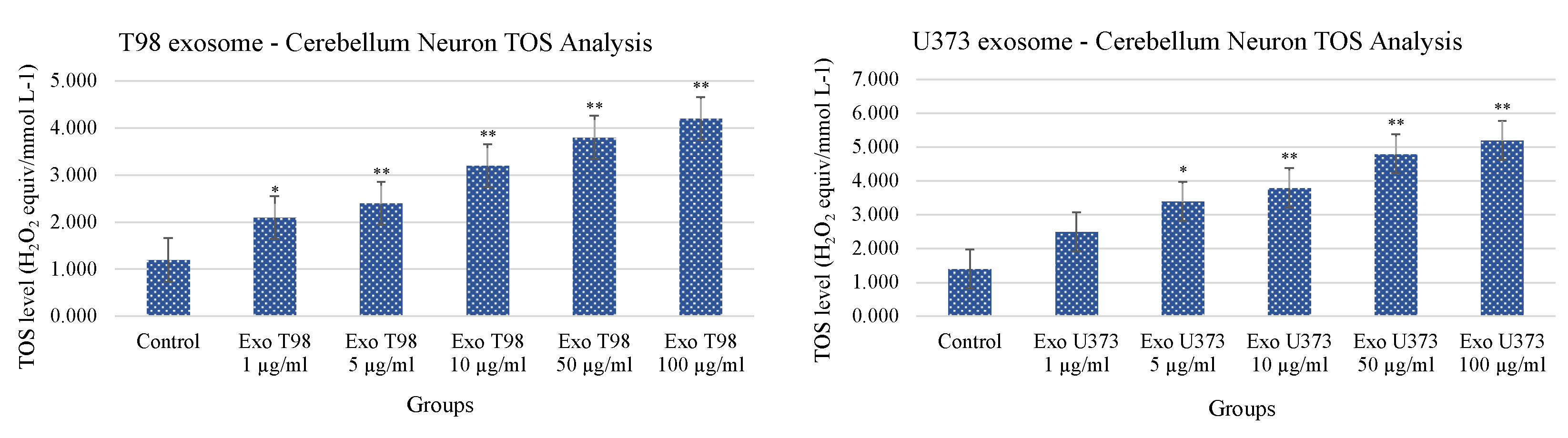
3.4. Total Antioxidant Status (TAS) and Total Oxidant Status (TOS) in Cerebellar Neurons Treated with T98G and U373 GBM-Derived Exosomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Altinoz, M.A.; Ozpinar, A.; Ozpinar, A.; Hacker, E.; Elmaci, I. Hypothesis: Could Hepatitis B vaccine act as an immune adjuvant in glioblastoma? Clues to conduct further epidemiological analyses. Int. Immunopharmacol. 2020, 81, 106038. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Liu, S.C.; Yang, Z.; Zhang, T. Impaired hippocampal synaptic plasticity in C6 glioma-bearing rats. J. Neuro-Oncol. 2011, 103, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Erturk Cetin, O.; Isler, C.; Uzan, M.; Ozkara, C. Epilepsy-related brain tumors. Seizure 2017, 44, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janelsins, M.C.; Kohli, S.; Mohile, S.G.; Usuki, K.; Ahles, T.A.; Morrow, G.R. An update on cancer- and chemotherapy-related cognitive dysfunction: Current status. Semin. Oncol. 2011, 38, 431–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, J.; Wu, J.Y.; Perucca, P.; Zelmann, R.; Mader, M.; Dubeau, F.; Mathern, G.W.; Schulze-Bonhage, A.; Gotman, J. Removing high-frequency oscillations: A prospective multicenter study on seizure outcome. Neurology 2018, 91, e1040–e1052. [Google Scholar] [CrossRef]
- Li, J.; Liu, M.; Gao, J.; Jiang, Y.; Wu, L.; Cheong, Y.K.; Ren, G.; Yang, Z. AVNP2 protects against cognitive impairments induced by C6 glioma by suppressing tumour associated inflammation in rats. Brain Behav. Immun. 2020, 87, 645–659. [Google Scholar] [CrossRef]
- Venugopal, C.; Wang, X.S.; Manoranjan, B.; McFarlane, N.; Nolte, S.; Li, M.; Murty, N.; Siu, K.W.; Singh, S.K. GBM secretome induces transient transformation of human neural precursor cells. J. Neurooncol. 2012, 109, 457–466. [Google Scholar] [CrossRef]
- Broekman, M.L.; Maas, S.L.N.; Abels, E.R.; Mempel, T.R.; Krichevsky, A.M.; Breakefield, X.O. Multidimensional communication in the microenvirons of glioblastoma. Nat. Rev. Neurol. 2018, 14, 482–495. [Google Scholar] [CrossRef]
- Lee, J.; Chaloner Winton Hall, R. The Impact of Gliomas on Cognition and Capacity. J. Am. Acad. Psychiatry Law. 2019, 47, 350–359. [Google Scholar] [CrossRef]
- Buckingham, S.C.; Campbell, S.L.; Haas, B.R.; Montana, V.; Robel, S.; Ogunrinu, T.; Sontheimer, H. Glutamate release by primary brain tumors induces epileptic activity. Nat. Med. 2011, 17, 1269–1274. [Google Scholar] [CrossRef]
- Wei, Z.; Batagov, A.O.; Schinelli, S.; Wang, J.; Wang, Y.; El Fatimy, R.; Rabinovsky, R.; Balaj, L.; Chen, C.C.; Hochberg, F.; et al. Coding and noncoding landscape of extracellular RNA released by human glioma stem cells. Nat. Commun. 2017, 8, 1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.D.; Schaal, D.; Kore, R.A.; Hamzah, R.N.; Pandanaboina, S.C.; Hayar, A.; Griffin, R.J.; Srivatsan, M.; Reyna, N.S.; Xie, J.Y. Glioma-derived exosomes drive the differentiation of neural stem cells to astrocytes. PLoS ONE 2020, 15, e0234614. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.Q.; Tan, C.J.; Toh, Y.L.; Sze, S.K.; Ho, H.K.; Limoli, C.L.; Chan, A. Role of Exosomes in Cancer-Related Cognitive Impairment. Int. J. Mol. Sci. 2020, 21, 2755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Pierce, D.; Light, K.E.; Melchert, R.B.; Fu, Q.; Kumar, K.S.; Hauer-Jensen, M. Sublethal total body irradiation leads to early cerebellar damage and oxidative stress. Curr. Neurovasc. Res. 2010, 7, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amidei, C.; Kushner, D.S. Clinical implications of motor deficits related to brain tumors†. Neurooncol. Pr. 2015, 2, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packer, R.J.; Meadows, A.T.; Rorke, L.B.; Goldwein, J.L.; D’Angio, G. Long-term sequelae of cancer treatment on the central nervous system in childhood. Med. Pediatr. Oncol. 1987, 15, 241–253. [Google Scholar] [CrossRef]
- Cantelmi, D.; Schweizer, T.A.; Cusimano, M.D. Role of the cerebellum in the neurocognitive sequelae of treatment of tumours of the posterior fossa: An update. Lancet Oncol. 2008, 9, 569–576. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef]
- Takahashi, A.; Okada, R.; Nagao, K.; Kawamata, Y.; Hanyu, A.; Yoshimoto, S.; Takasugi, M.; Watanabe, S.; Kanemaki, M.T.; Obuse, C.; et al. Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat. Commun. 2017, 8, 15287. [Google Scholar] [CrossRef] [Green Version]
- Aires, I.D.; Ribeiro-Rodrigues, T.; Boia, R.; Catarino, S.; Girao, H.; Ambrosio, A.F.; Santiago, A.R. Exosomes derived from microglia exposed to elevated pressure amplify the neuroinflammatory response in retinal cells. Glia 2020, 68, 2705–2724. [Google Scholar] [CrossRef]
- D’Asti, E.; Chennakrishnaiah, S.; Lee, T.H.; Rak, J. Extracellular Vesicles in Brain Tumor Progression. Cell. Mol. Neurobiol. 2016, 36, 383–407. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.K.; Khan, M.A.; Zubair, H.; Srivastava, S.K.; Khushman, M.; Singh, S.; Singh, A.P. Comparative analysis of exosome isolation methods using culture supernatant for optimum yield, purity and downstream applications. Sci. Rep. 2019, 9, 5335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.T.; Huang, Y.Y.; Zheng, L.; Qin, S.H.; Xu, X.P.; An, T.X.; Xu, Y.; Wu, Y.S.; Hu, X.M. Comparison of isolation methods of exosomes and exosomal RNA from cell culture medium and serum. Int. J. Mol. Med. 2017, 40, 834–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Jiang, L.; Hu, H.; Wang, H.; Wang, X.; Jiang, J.; Ma, Y.; Yang, J.; Hou, Y.; Xie, D.; et al. Pretreatment of exosomes derived from hUCMSCs with TNF-α ameliorates acute liver failure by inhibiting the activation of NLRP3 in macrophage. Life Sci. 2020, 246, 117401. [Google Scholar] [CrossRef]
- Sharma, P.; Mesci, P.; Carromeu, C.; McClatchy, D.R.; Schiapparelli, L.; Yates, J.R., 3rd; Muotri, A.R.; Cline, H.T. Exosomes regulate neurogenesis and circuit assembly. Proc. Natl. Acad. Sci. USA 2019, 116, 16086–16094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monfared, H.; Jahangard, Y.; Nikkhah, M.; Mirnajafi-Zadeh, J.; Mowla, S.J. Potential Therapeutic Effects of Exosomes Packed with a miR-21-Sponge Construct in a Rat Model of Glioblastoma. Front. Oncol. 2019, 9, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Wang, J.; Li, H.; Gao, S.; Shi, R.; Yang, D.; Wang, X.; Wang, X.; Zhu, L.; Wang, X.; et al. Extracellular Vesicles Secreted by Human Adipose-derived Stem Cells (hASCs) Improve Survival Rate of Rats with Acute Liver Failure by Releasing lncRNA H19. EBioMedicine 2018, 34, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Christofi, T.; Baritaki, S.; Falzone, L.; Libra, M.; Zaravinos, A. Current Perspectives in Cancer Immunotherapy. Cancers 2019, 11, 1472. [Google Scholar] [CrossRef] [Green Version]
- Falzone, L.; Salomone, S.; Libra, M. Evolution of Cancer Pharmacological Treatments at the Turn of the Third Millennium. Front. Pharmacol. 2018, 9, 1300. [Google Scholar] [CrossRef] [Green Version]
- Silantyev, A.S.; Falzone, L.; Libra, M.; Gurina, O.I.; Kardashova, K.S.; Nikolouzakis, T.K.; Nosyrev, A.E.; Sutton, C.W.; Mitsias, P.D.; Tsatsakis, A. Current and Future Trends on Diagnosis and Prognosis of Glioblastoma: From Molecular Biology to Proteomics. Cells 2019, 8, 863. [Google Scholar] [CrossRef] [Green Version]
- Candido, S.; Lupo, G.; Pennisi, M.; Basile, M.S.; Anfuso, C.D.; Petralia, M.C.; Gattuso, G.; Vivarelli, S.; Spandidos, D.A.; Libra, M.; et al. The analysis of miRNA expression profiling datasets reveals inverse microRNA patterns in glioblastoma and Alzheimer’s disease. Oncol. Rep. 2019, 42, 911–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuaeva, N.O.; Falzone, L.; Porozov, Y.B.; Nosyrev, A.E.; Trukhan, V.M.; Kovatsi, L.; Spandidos, D.A.; Drakoulis, N.; Kalogeraki, A.; Mamoulakis, C.; et al. Translational Application of Circulating DNA in Oncology: Review of the Last Decades Achievements. Cells 2019, 8, 1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Certo, F.; Altieri, R.; Maione, M.; Schonauer, C.; Sortino, G.; Fiumanò, G.; Tirrò, E.; Massimino, M.; Broggi, G.; Vigneri, P.; et al. FLAIRectomy in Supramarginal Resection of Glioblastoma Correlates with Clinical Outcome and Survival Analysis: Prospective, Single Institution, Case Series. Oper. Neurosurg. 2021, 20, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Falzone, L.; Gattuso, G.; Tsatsakis, A.; Spandidos, D.A.; Libra, M. Current and innovative methods for the diagnosis of COVID-19 infection (Review). Int. J. Mol. Med. 2021, 47, 100. [Google Scholar] [CrossRef]
- Barbagallo, D.; Caponnetto, A.; Barbagallo, C.; Battaglia, R.; Mirabella, F.; Brex, D.; Stella, M.; Broggi, G.; Altieri, R.; Certo, F.; et al. The GAUGAA Motif Is Responsible for the Binding between circSMARCA5 and SRSF1 and Related Downstream Effects on Glioblastoma Multiforme Cell Migration and Angiogenic Potential. Int. J. Mol. Sci. 2021, 22, 1678. [Google Scholar] [CrossRef]
- Broggi, G.; Salvatorelli, L.; Barbagallo, D.; Certo, F.; Altieri, R.; Tirrò, E.; Massimino, M.; Vigneri, P.; Guadagno, E.; Maugeri, G.; et al. Diagnostic Utility of the Immunohistochemical Expression of Serine and Arginine Rich Splicing Factor 1 (SRSF1) in the Differential Diagnosis of Adult Gliomas. Cancers 2021, 13, 2086. [Google Scholar] [CrossRef]
- Stella, M.; Falzone, L.; Caponnetto, A.; Gattuso, G.; Barbagallo, C.; Battaglia, R.; Mirabella, F.; Broggi, G.; Altieri, R.; Certo, F.; et al. Serum Extracellular Vesicle-Derived circHIPK3 and circSMARCA5 Are Two Novel Diagnostic Biomarkers for Glioblastoma Multiforme. Pharmaceuticals 2021, 14, 618. [Google Scholar] [CrossRef]
- Balasa, A.; Serban, G.; Chinezu, R.; Hurghis, C.; Tamas, F.; Manu, D. The Involvement of Exosomes in Glioblastoma Development, Diagnosis, Prognosis, and Treatment. Brain Sci. 2020, 10, 553. [Google Scholar] [CrossRef]
- Ciregia, F.; Urbani, A.; Palmisano, G. Extracellular Vesicles in Brain Tumors and Neurodegenerative Diseases. Front. Mol. Neurosci. 2017, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Katrib, A.; Jeong, H.H.; Fransen, N.L.; Henzel, K.S.; Miller, J.A. An Inflammatory Landscape for Preoperative Neurologic Deficits in Glioblastoma. Front. Genet. 2019, 10, 488. [Google Scholar] [CrossRef] [Green Version]
- Lange, F.; Hörnschemeyer, J.; Kirschstein, T. Glutamatergic Mechanisms in Glioblastoma and Tumor-Associated Epilepsy. Cells 2021, 10, 1226. [Google Scholar] [CrossRef] [PubMed]
- Șovrea, A.S.; Boșca, B.; Melincovici, C.S.; Constantin, A.M.; Crintea, A.; Mărginean, M.; Dronca, E.; Jianu, M.E.; Suflețel, R.; Gonciar, D.; et al. Multiple Faces of the Glioblastoma Microenvironment. Int. J. Mol. Sci. 2022, 23, 595. [Google Scholar] [CrossRef]
- Wefel, J.S.; Noll, K.R.; Scheurer, M.E. Neurocognitive functioning and genetic variation in patients with primary brain tumours. Lancet Oncol. 2016, 17, e97–e108. [Google Scholar] [CrossRef] [Green Version]
- Facecchia, K.; Fochesato, L.A.; Ray, S.D.; Stohs, S.J.; Pandey, S. Oxidative toxicity in neurodegenerative diseases: Role of mitochondrial dysfunction and therapeutic strategies. J. Toxicol. 2011, 2011, 683728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, J.M.; Öberg, J.; Brené, S.; Coppotelli, G.; Terzioglu, M.; Pernold, K.; Goiny, M.; Sitnikov, R.; Kehr, J.; Trifunovic, A.; et al. High brain lactate is a hallmark of aging and caused by a shift in the lactate dehydrogenase A/B ratio. Proc. Natl. Acad. Sci. USA 2010, 107, 20087–20092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backos, D.S.; Fritz, K.S.; McArthur, D.G.; Kepa, J.K.; Donson, A.M.; Petersen, D.R.; Foreman, N.K.; Franklin, C.C.; Reigan, P. Glycation of glutamate cysteine ligase by 2-deoxy-d-ribose and its potential impact on chemoresistance in glioblastoma. Neurochem. Res. 2013, 38, 1838–1849. [Google Scholar] [CrossRef] [PubMed]
- Valvona, C.J.; Fillmore, H.L.; Nunn, P.B.; Pilkington, G.J. The Regulation and Function of Lactate Dehydrogenase A: Therapeutic Potential in Brain Tumor. Brain Pathol. 2016, 26, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Daniele, S.; Giacomelli, C.; Zappelli, E.; Granchi, C.; Trincavelli, M.L.; Minutolo, F.; Martini, C. Lactate dehydrogenase-A inhibition induces human glioblastoma multiforme stem cell differentiation and death. Sci. Rep. 2015, 5, 15556. [Google Scholar] [CrossRef] [Green Version]
- Long, D.M.; Frame, A.K.; Reardon, P.N.; Cumming, R.C.; Hendrix, D.A.; Kretzschmar, D.; Giebultowicz, J.M. Lactate dehydrogenase expression modulates longevity and neurodegeneration in Drosophila melanogaster. Aging 2020, 12, 10041–10058. [Google Scholar] [CrossRef]
- Spinelli, C.; Montermini, L.; Meehan, B.; Brisson, A.R.; Tan, S.; Choi, D.; Nakano, I.; Rak, J. Molecular subtypes and differentiation programmes of glioma stem cells as determinants of extracellular vesicle profiles and endothelial cell-stimulating activities. J. Extracell. Vesicles 2018, 7, 1490144. [Google Scholar] [CrossRef]
- Spinelli, C.; Tawil, N.; Adnani, L.; Rak, J.; Choi, D. Extracellular Vesicle Mediated Vascular Pathology in Glioblastoma. Subcell. Biochem. 2021, 97, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Matarredona, E.R.; Pastor, A.M. Extracellular Vesicle-Mediated Communication between the Glioblastoma and Its Microenvironment. Cells 2019, 9, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yekula, A.; Yekula, A.; Muralidharan, K.; Kang, K.; Carter, B.S.; Balaj, L. Extracellular Vesicles in Glioblastoma Tumor Microenvironment. Front. Immunol. 2020, 10, 3137. [Google Scholar] [CrossRef] [PubMed]
- Guescini, M.; Genedani, S.; Stocchi, V.; Agnati, L.F. Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. J. Neural. Transm. 2010, 117, 1. [Google Scholar] [CrossRef] [PubMed]
- Agnati, L.F.; Guidolin, D.; Baluska, F.; Leo, G.; Barlow, P.W.; Carone, C.; Genedani, S. A new hypothesis of pathogenesis based on the divorce between mitochondria and their host cells: Possible relevance for Alzheimer’s disease. Curr. Alzheimer Res. 2010, 7, 307–322. [Google Scholar] [CrossRef]
- Singh, B.; Modica-Napolitano, J.S.; Singh, K.K. Defining the momiome: Promiscuous information transfer by mobile mitochondria and the mitochondrial genome. Semin. Cancer Biol. 2017, 47, 1–17. [Google Scholar] [CrossRef]
- Jin, Q.; Wu, P.; Zhou, X.; Qian, H.; Xu, W. Extracellular Vesicles: Novel Roles in Neurological Disorders. Stem. Cells Int. 2021, 2021, 6640836. [Google Scholar] [CrossRef]
- Jiang, L.; Dong, H.; Cao, H.; Ji, X.; Luan, S.; Liu, J. Exosomes in Pathogenesis, Diagnosis, and Treatment of Alzheimer’s Disease. Med. Sci. Monit. 2019, 25, 3329–3335. [Google Scholar] [CrossRef]
- Porro, C.; Panaro, M.A.; Lofrumento, D.D.; Hasalla, E.; Trotta, T. The multiple roles of exosomes in Parkinson’s disease: An overview. Immunopharmacol. Immunotoxicol. 2019, 41, 469–476. [Google Scholar] [CrossRef]
- Karttunen, J.; Heiskanen, M.; Lipponen, A.; Poulsen, D.; Pitkänen, A. Extracellular Vesicles as Diagnostics and Therapeutics for Structural Epilepsies. Int. J. Mol. Sci. 2019, 20, 1259. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genc, S.; Pennisi, M.; Yeni, Y.; Yildirim, S.; Gattuso, G.; Altinoz, M.A.; Taghizadehghalehjoughi, A.; Bolat, I.; Tsatsakis, A.; Hacımüftüoğlu, A.; et al. Potential Neurotoxic Effects of Glioblastoma-Derived Exosomes in Primary Cultures of Cerebellar Neurons via Oxidant Stress and Glutathione Depletion. Antioxidants 2022, 11, 1225. https://doi.org/10.3390/antiox11071225
Genc S, Pennisi M, Yeni Y, Yildirim S, Gattuso G, Altinoz MA, Taghizadehghalehjoughi A, Bolat I, Tsatsakis A, Hacımüftüoğlu A, et al. Potential Neurotoxic Effects of Glioblastoma-Derived Exosomes in Primary Cultures of Cerebellar Neurons via Oxidant Stress and Glutathione Depletion. Antioxidants. 2022; 11(7):1225. https://doi.org/10.3390/antiox11071225
Chicago/Turabian StyleGenc, Sidika, Manuela Pennisi, Yesim Yeni, Serkan Yildirim, Giuseppe Gattuso, Meric A. Altinoz, Ali Taghizadehghalehjoughi, Ismail Bolat, Aristidis Tsatsakis, Ahmet Hacımüftüoğlu, and et al. 2022. "Potential Neurotoxic Effects of Glioblastoma-Derived Exosomes in Primary Cultures of Cerebellar Neurons via Oxidant Stress and Glutathione Depletion" Antioxidants 11, no. 7: 1225. https://doi.org/10.3390/antiox11071225
APA StyleGenc, S., Pennisi, M., Yeni, Y., Yildirim, S., Gattuso, G., Altinoz, M. A., Taghizadehghalehjoughi, A., Bolat, I., Tsatsakis, A., Hacımüftüoğlu, A., & Falzone, L. (2022). Potential Neurotoxic Effects of Glioblastoma-Derived Exosomes in Primary Cultures of Cerebellar Neurons via Oxidant Stress and Glutathione Depletion. Antioxidants, 11(7), 1225. https://doi.org/10.3390/antiox11071225