
Nitric Oxide Production and Regulation in the Teleost Cardiovascular System




Abstract
1. Introduction
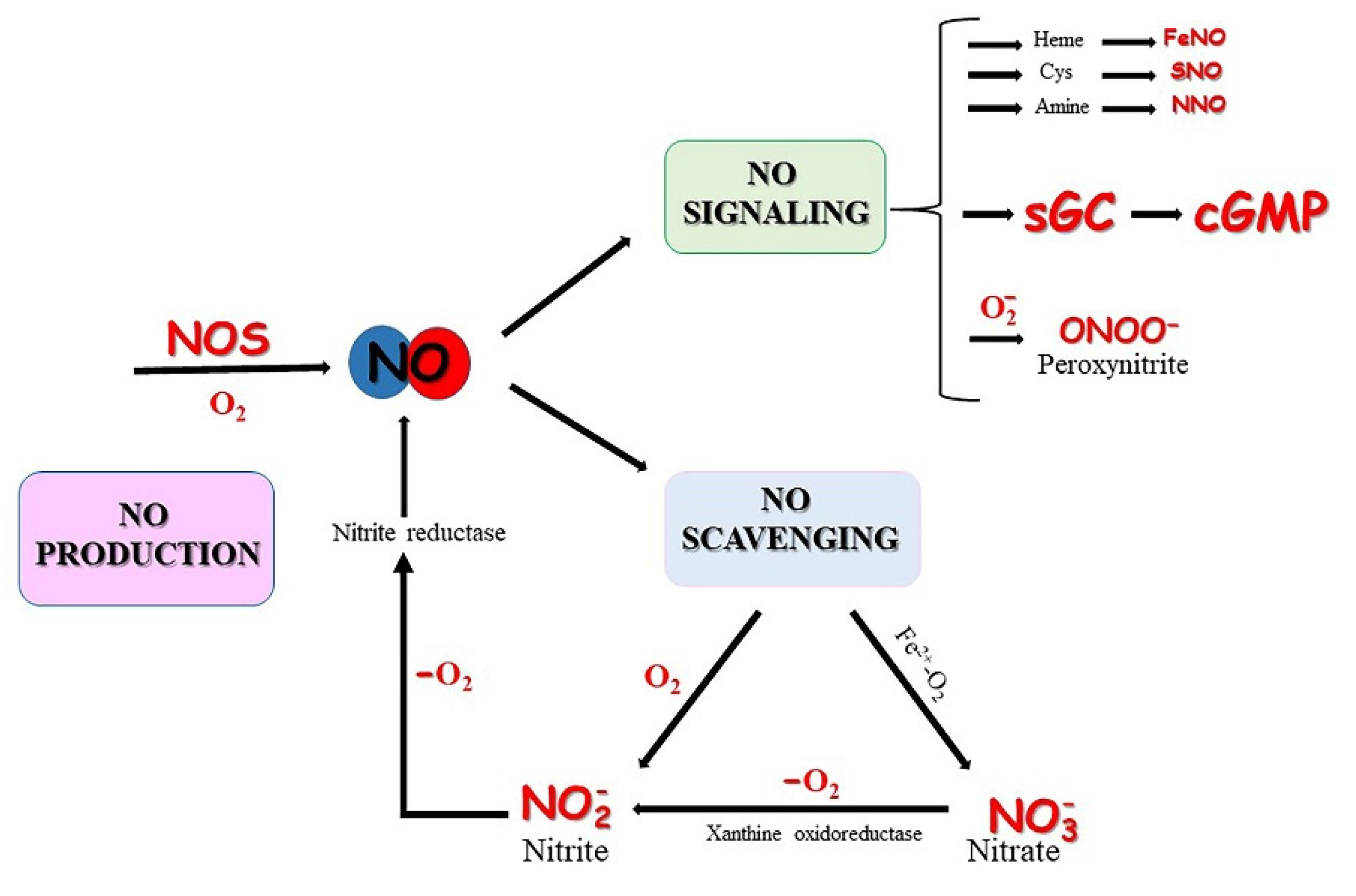
2. Oxygen-Dependent NO Synthesis
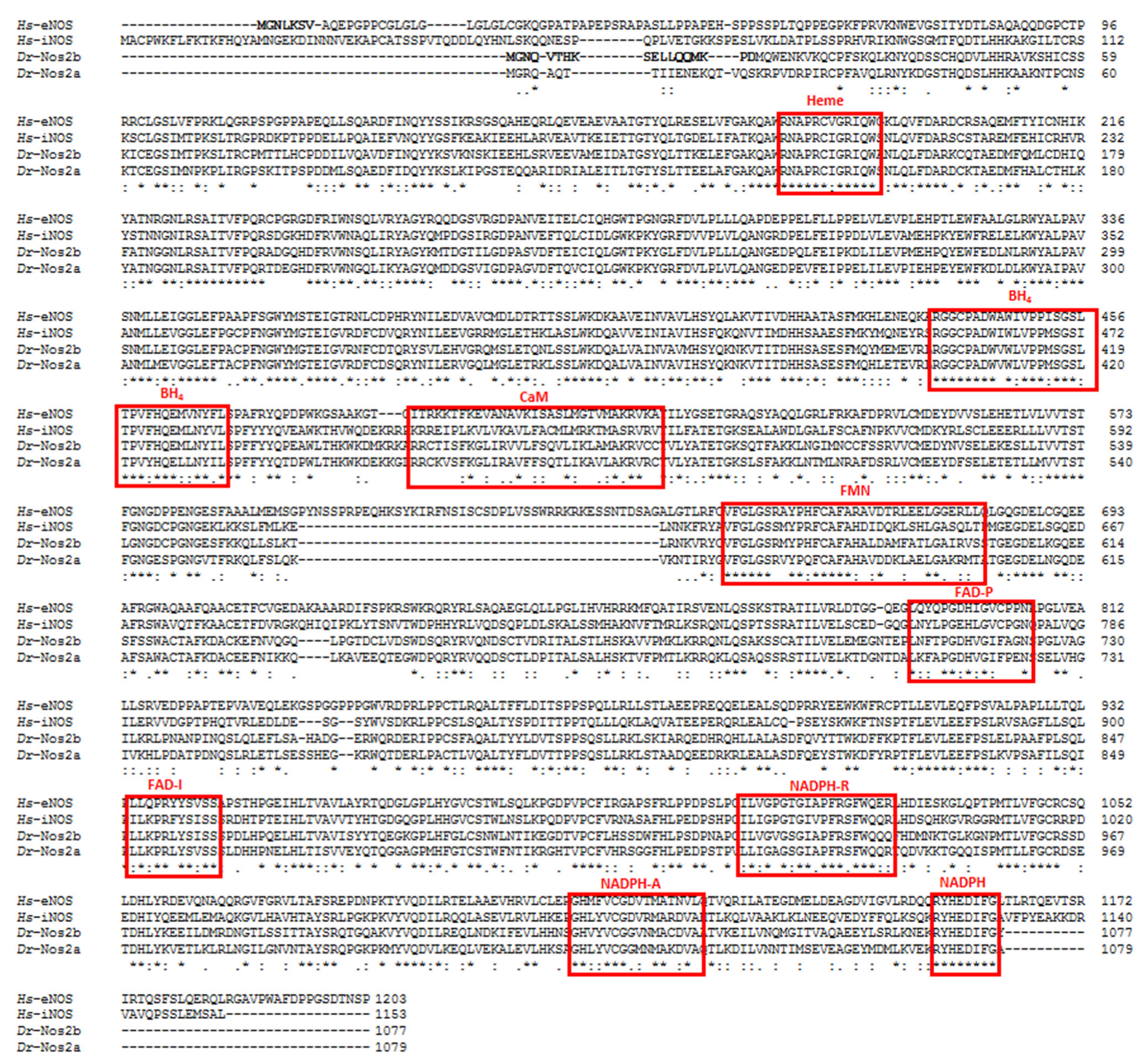
Evolution and Expression of NOSs
3. Nitrite-Dependent NO Formation
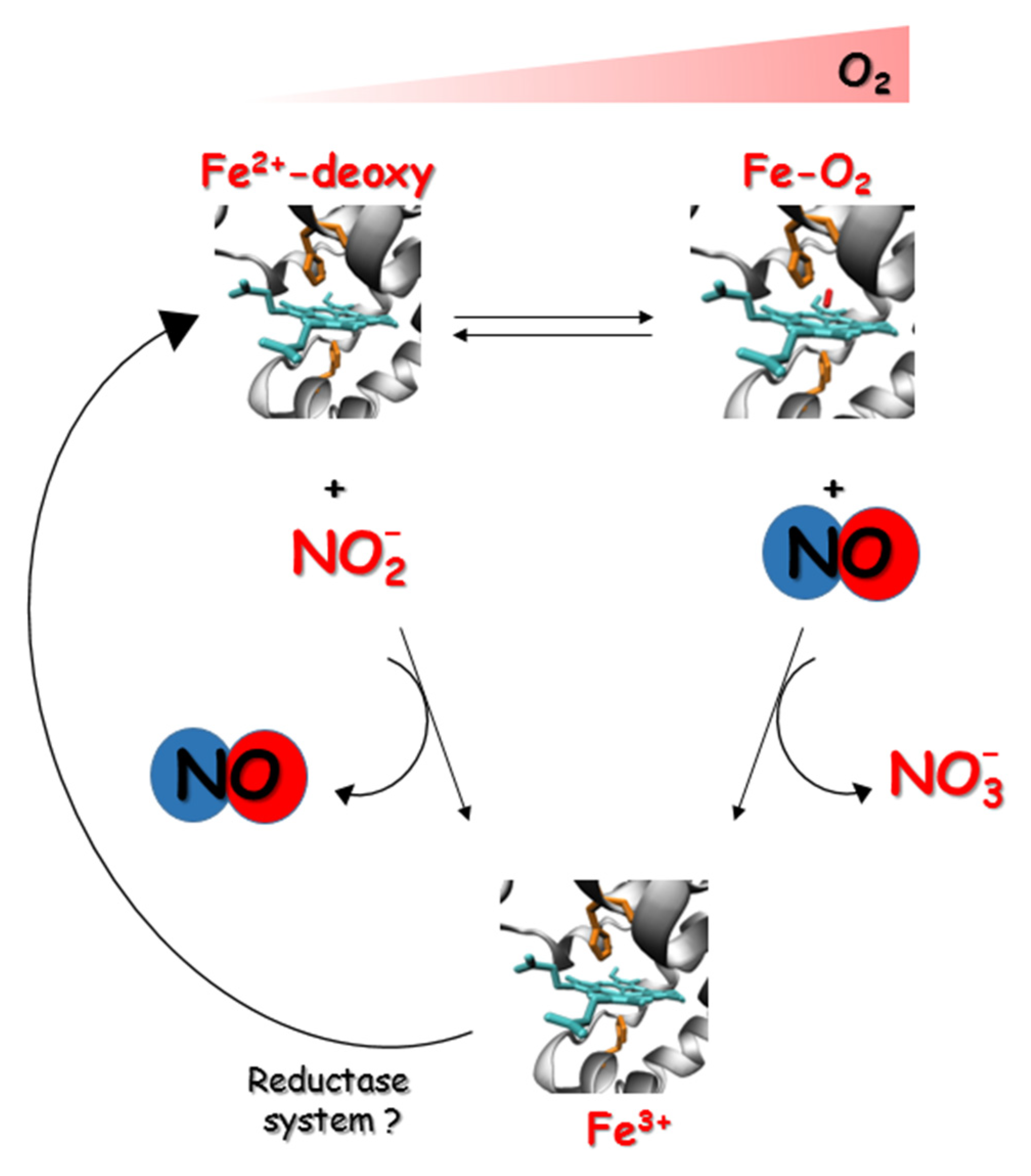
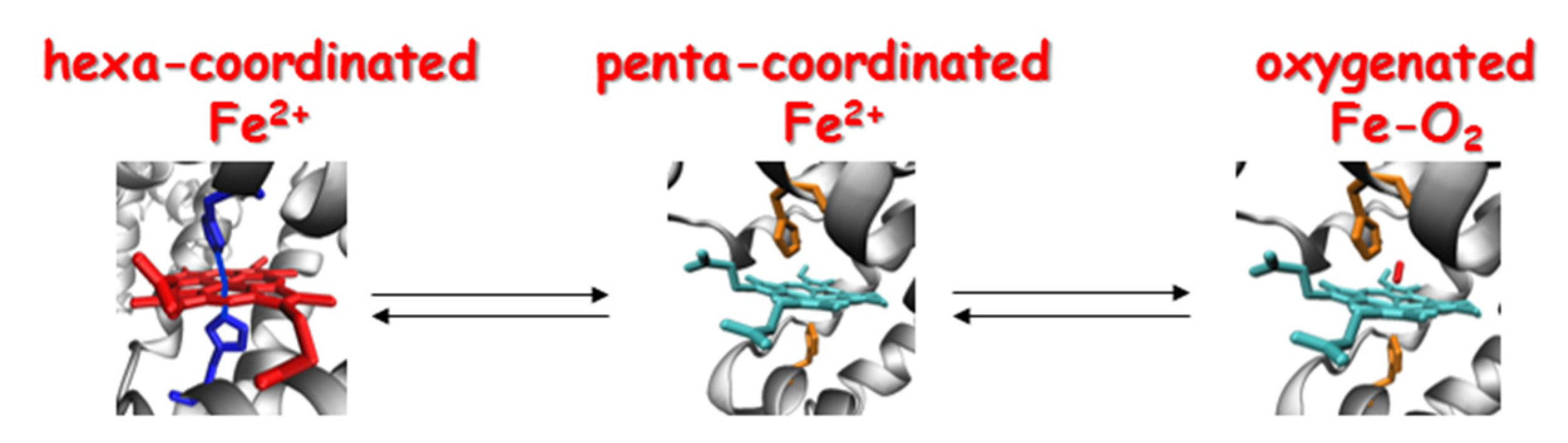
3.1. NO Formation by Globins
3.2. NO Scavenging by Globins
4. NO Signaling in Teleost
5. S-Nitrosylation of Globins
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feelisch, M.; Martin, J.F. The early role of nitric oxide in evolution. Trends Ecol. Evol. 1995, 10, 496–499. [Google Scholar] [CrossRef]
- Brüne, B.; Mohr, S.; Messmer, U.K. Protein thiol modification and apoptotic cell death as cGMP-independent nitric oxide (NO) signaling pathways. Rev. Physiol. Biochem. Pharmacol. 1996, 127, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.M.J.; Ferrige, A.G.; Moncada, S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 1987, 327, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Buga, G.M.; Wood, K.S.; Byrns, R.E.; Chaudhuri, G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc. Natl. Acad. Sci. USA 1987, 84, 9265–9269. [Google Scholar] [CrossRef]
- Arnold, W.P.; Mittal, C.K.; Katsuki, S.; Murad, F. Nitric oxide activates guanylate cyclase and increases guanosine 3′:5′-cyclic monophosphate levels in various tissue preparations. Proc. Natl. Acad. Sci. USA 1977, 74, 3203–3207. [Google Scholar] [CrossRef]
- Furchgott, R.F.; Zawadzki, J.V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Schmidt, H.H.H.W.; Walter, U. NO at work. Cell 1994, 78, 919–925. [Google Scholar] [CrossRef]
- Moroz, L.L.; Kohn, A.B. On the comparative biology of Nitric Oxide (NO) synthetic pathways: Parallel evolution of NO-mediated signaling. In Nitric Oxide; Advances in Experimental Biology; Tota, B., Trimmer, B.B.T.-A., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 1–44. ISBN 1872-2423. [Google Scholar]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef]
- Gladwin, M.T.; Kim-Shapiro, D.B. The functional nitrite reductase activity of the heme-globins. Blood 2008, 112, 2636–2647. [Google Scholar] [CrossRef]
- Shikama, K. Nature of the FeO2 bonding in myoglobin and hemoglobin: A new molecular paradigm. Prog. Biophys. Mol. Biol. 2006, 91, 83–162. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate–nitrite–nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Gladwin, M.T.; Schechter, A.N.; Kim-Shapiro, D.B.; Patel, R.P.; Hogg, N.; Shiva, S.; Cannon, R.O.; Kelm, M.; Wink, D.A.; Espey, M.G.; et al. The emerging biology of the nitrite anion. Nat. Chem. Biol. 2005, 1, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Cosby, K.; Partovi, K.S.; Crawford, J.H.; Patel, R.P.; Reiter, C.D.; Martyr, S.; Yang, B.K.; Waclawiw, M.A.; Zalos, G.; Xu, X.; et al. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat. Med. 2003, 9, 1498–1505. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, J.; Stark, K.; Esslinger, U.B.; Rumpf, P.M.; Koesling, D.; de Wit, C.; Kaiser, F.J.; Braunholz, D.; Medack, A.; Fischer, M.; et al. Dysfunctional nitric oxide signalling increases risk of myocardial infarction. Nature 2013, 504, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Wobst, J.; Kessler, T.; Dang, T.A.; Erdmann, J.; Schunkert, H. Role of sGC-dependent NO signalling and myocardial infarction risk. J. Mol. Med. 2015, 93, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Gladwin, M.T.; Weitzberg, E. Strategies to increase nitric oxide signalling in cardiovascular disease. Nat. Rev. Drug Discov. 2015, 14, 623–641. [Google Scholar] [CrossRef]
- Hill, B.G.; Dranka, B.P.; Bailey, S.M.; Lancaster, J.R., Jr.; Darley-Usmar, V.M. What Part of NO Don’t You Understand? Some Answers to the Cardinal Questions in Nitric Oxide Biology. J. Biol. Chem. 2010, 285, 19699–19704. [Google Scholar] [CrossRef]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef]
- Farah, C.; Michel, L.Y.M.; Balligand, J.-L. Nitric oxide signalling in cardiovascular health and disease. Nat. Rev. Cardiol. 2018, 15, 292–316. [Google Scholar] [CrossRef]
- Toda, N.; Ayajiki, K. Phylogenesis of constitutively formed nitric oxide in non-mammals. Rev. Physiol. Biochem. Pharmacol. 2006, 157, 31–80. [Google Scholar] [PubMed]
- Rochon, E.R.; Corti, P. Globins and nitric oxide homeostasis in fish embryonic development. Mar. Genom. 2020, 49, 100721. [Google Scholar] [CrossRef] [PubMed]
- Grayfer, L.; Kerimoglu, B.; Yaparla, A.; Hodgkinson, J.W.; Xie, J.; Belosevic, M. Mechanisms of Fish Macrophage Antimicrobial Immunity. Front. Immunol. 2018, 9, 1105. [Google Scholar] [CrossRef] [PubMed]
- Bickler, P.E.; Buck, L.T. Hypoxia tolerance in reptiles, amphibians, and fishes: Life with variable oxygen availability. Annu. Rev. Physiol. 2007, 69, 145–170. [Google Scholar] [CrossRef]
- Pedersen, C.L.; Faggiano, S.; Helbo, S.; Gesser, H.; Fago, A. Roles of nitric oxide, nitrite and myoglobin on myocardial efficiency in trout (Oncorhynchus mykiss) and goldfish (Carassius auratus): Implications for hypoxia tolerance. J. Exp. Biol. 2010, 213, 2755–2762. [Google Scholar] [CrossRef][Green Version]
- Padilla, P.A.; Roth, M.B. Oxygen deprivation causes suspended animation in the zebrafish embryo. Proc. Natl. Acad. Sci. USA 2001, 98, 7331–7335. [Google Scholar] [CrossRef]
- Sakai, C.; Ijaz, S.; Hoffman, E.J. Zebrafish Models of Neurodevelopmental Disorders: Past, Present, and Future. Front. Mol. Neurosci. 2018, 11, 294. [Google Scholar] [CrossRef]
- Rees, B.B.; Sudradjat, F.A.; Love, J.W. Acclimation to hypoxia increases survival time of zebrafish, Danio rerio, during lethal hypoxia. J. Exp. Zool. 2001, 289, 266–272. [Google Scholar] [CrossRef]
- Daane, J.M.; Auvinet, J.; Stoebenau, A.; Yergeau, D.; Harris, M.P.; Detrich, H.W. Developmental constraint shaped genome evolution and erythrocyte loss in Antarctic fishes following paleoclimate change. PLoS Genet. 2020, 16, e1009173. [Google Scholar] [CrossRef]
- Sidell, B.D.; O’Brien, K.M. When bad things happen to good fish: The loss of hemoglobin and myoglobin expression in Antarctic icefishes. J. Exp. Biol. 2006, 209, 1791–1802. [Google Scholar] [CrossRef]
- Moylan, T.J.; Sidell, B.D. Concentrations of myoglobin and myoglobin mRNA in heart ventricles from Antarctic fishes. J. Exp. Biol. 2000, 203, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Corti, P.; Coppola, D.; Altomonte, G.; Xue, J.; Russo, R.; di Prisco, G.; Verde, C. Regulation of globin expression in Antarctic fish under thermal and hypoxic stress. Mar. Genom. 2021, 57, 100831. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Pesce, A.; Vermeylen, S.; Abbruzzetti, S.; Nardini, M.; Marchesani, F.; Berghmans, H.; Seira, C.; Bruno, S.; Luque, F.J.; et al. Structural and functional properties of Antarctic fish cytoglobins-1: Cold-reactivity in multi-ligand reactions. Comput. Struct. Biotechnol. J. 2020, 18, 2132–2144. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, B.; Vermeylen, S.; Hammerschmid, D.; Trashin, S.; Rahemi, V.; Konijnenberg, A.; De Schutter, A.; Cheng, C.-H.C.; Giordano, D.; Verde, C.; et al. Antarctic fish versus human cytoglobins—The same but yet so different. J. Inorg. Biochem. 2017, 173, 66–78. [Google Scholar] [CrossRef]
- Giordano, D.; Boron, I.; Abbruzzetti, S.; Van Leuven, W.; Nicoletti, F.P.; Forti, F.; Bruno, S.; Cheng, C.-H.C.; Moens, L.; di Prisco, G.; et al. Biophysical characterisation of neuroglobin of the icefish, a natural knockout for hemoglobin and myoglobin. Comparison with human neuroglobin. PLoS ONE 2012, 7, e44508. [Google Scholar] [CrossRef]
- Cheng, C.-H.C.; di Prisco, G.; Verde, C. The “icefish paradox.” Which is the task of neuroglobin in Antarctic hemoglobin-less icefish? IUBMB Life 2009, 61, 184–188. [Google Scholar] [CrossRef]
- Beers, J.M.; Jayasundara, N. Antarctic notothenioid fish: What are the future consequences of “losses” and “gains” acquired during long-term evolution at cold and stable temperatures? J. Exp. Biol. 2015, 218, 1834–1845. [Google Scholar] [CrossRef]
- Daane, J.M.; Giordano, D.; Coppola, D.; di Prisco, G.; Detrich, H.W., 3rd; Verde, C. Adaptations to environmental change: Globin superfamily evolution in Antarctic fishes. Mar. Genom. 2020, 49, 100724. [Google Scholar] [CrossRef]
- Cheng, C.-H.C.; Detrich, H.W., 3rd. Molecular ecophysiology of Antarctic notothenioid fishes. Philos. Trans. R. Soc. London Ser. B Biol. Sci. 2007, 362, 2215–2232. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.M.; Sidell, B.D. The interplay among cardiac ultrastructure, metabolism and the expression of oxygen-binding proteins in Antarctic fishes. J. Exp. Biol. 2000, 203, 1287–1297. [Google Scholar] [CrossRef] [PubMed]
- Andreakis, N.; D’Aniello, S.; Albalat, R.; Patti, F.P.; Garcia-Fernndez, J.; Procaccini, G.; Sordino, P.; Palumbo, A. Evolution of the nitric oxide synthase family in metazoans. Mol. Biol. Evol. 2011, 28, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Mattila, J.T.; Thomas, A.C. Nitric oxide synthase: Non-canonical expression patterns. Front. Immunol. 2014, 5, 478. [Google Scholar] [CrossRef] [PubMed]
- Victor, V.M.; Nuñez, C.; D’Ocón, P.; Taylor, C.T.; Esplugues, J.V.; Moncada, S. Regulation of oxygen distribution in tissues by endothelial nitric oxide. Circ. Res. 2009, 104, 1178–1183. [Google Scholar] [CrossRef] [PubMed]
- Nisoli, E.; Tonello, C.; Cardile, A.; Cozzi, V.; Bracale, R.; Tedesco, L.; Falcone, S.; Valerio, A.; Cantoni, O.; Clementi, E.; et al. Calorie restriction promotes mitochondrial biogenesis by inducing the expression of eNOS. Science 2005, 310, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Ghafourifar, P.; Cadenas, E. Mitochondrial nitric oxide synthase. Trends Pharmacol. Sci. 2005, 26, 190–195. [Google Scholar] [CrossRef]
- Lacza, Z.; Puskar, M.; Figueroa, J.P.; Zhang, J.; Rajapakse, N.; Busija, D.W. Mitochondrial nitric oxide synthase is constitutively active and is functionally upregulated in hypoxia. Free Radic. Biol. Med. 2001, 31, 1609–1615. [Google Scholar] [CrossRef]
- Bredt, D.S. Nitric oxide signaling in brain: Potentiating the gain with YC-1. Mol. Pharmacol. 2003, 63, 1206–1208. [Google Scholar] [CrossRef]
- Barouch, L.A.; Harrison, R.W.; Skaf, M.W.; Rosas, G.O.; Cappola, T.P.; Kobeissi, Z.A.; Hobai, I.A.; Lemmon, C.A.; Burnett, A.L.; O’Rourke, B.; et al. Nitric oxide regulates the heart by spatial confinement of nitric oxide synthase isoforms. Nature 2002, 416, 337–339. [Google Scholar] [CrossRef]
- Daff, S.; Sagami, I.; Shimizu, T. The 42-amino acid insert in the FMN domain of neuronal nitric-oxide synthase exerts control over Ca(2+)/calmodulin-dependent electron transfer. J. Biol. Chem. 1999, 274, 30589–30595. [Google Scholar] [CrossRef]
- Salerno, J.C.; Harris, D.E.; Irizarry, K.; Patel, B.; Morales, A.J.; Smith, S.M.; Martasek, P.; Roman, L.J.; Masters, B.S.; Jones, C.L.; et al. An autoinhibitory control element defines calcium-regulated isoforms of nitric oxide synthase. J. Biol. Chem. 1997, 272, 29769–29777. [Google Scholar] [CrossRef] [PubMed]
- Prolo, C.; Alvarez, M.N.; Radi, R. Peroxynitrite, a potent macrophage-derived oxidizing cytotoxin to combat invading pathogens. Biofactors 2014, 40, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Loughran, P.A.; Stolz, D.B.; Barrick, S.R.; Wheeler, D.S.; Friedman, P.A.; Rachubinski, R.A.; Watkins, S.C.; Billiar, T.R. PEX7 and EBP50 target iNOS to the peroxisome in hepatocytes. Nitric Oxide Biol. Chem. 2013, 31, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.K.; Salerno, J.C. Nitric oxide synthases: Domain structure and alignment in enzyme function and control. Front. Biosci. 2003, 8, 193–209. [Google Scholar] [CrossRef]
- Griffith, O.W.; Stuehr, D.J. Nitric oxide synthases: Properties and catalytic mechanism. Annu. Rev. Physiol. 1995, 57, 707–736. [Google Scholar] [CrossRef] [PubMed]
- Crane, B.R.; Sudhamsu, J.; Patel, B.A. Bacterial Nitric Oxide Synthases. Annu. Rev. Biochem. 2010, 79, 445–470. [Google Scholar] [CrossRef]
- Hutfless, E.H.; Chaudhari, S.S.; Thomas, V.C. Chapter Five—Emerging Roles of Nitric Oxide Synthase in Bacterial Physiology. In Nitric Oxide and Other Small Signalling Molecules; Advances in Microbial Physiology; Poole, R.K.B.T.-A., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 72, pp. 147–191. ISBN 0065-2911. [Google Scholar]
- Agapie, T.; Suseno, S.; Woodward, J.J.; Stoll, S.; Britt, R.D.; Marletta, M.A. NO formation by a catalytically self-sufficient bacterial nitric oxide synthase from Sorangium cellulosum. Proc. Natl. Acad. Sci. USA 2009, 106, 16221–16226. [Google Scholar] [CrossRef]
- Lepiller, S.; Franche, N.; Solary, E.; Chluba, J.; Laurens, V. Comparative analysis of zebrafish nos2a and nos2b genes. Gene 2009, 445, 58–65. [Google Scholar] [CrossRef]
- Annona, G.; Sato, I.; Pascual-Anaya, J.; Braasch, I.; Voss, R.; Stundl, J.; Soukup, V.; Kuratani, S.; Postlethwait, J.H.; D’Aniello, S. Evolution of the Nitric Oxide Synthase Family in Vertebrates and Novel Insights in Gill Development. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lamas, S.; Marsden, P.A.; Li, G.K.; Tempst, P.; Michel, T. Endothelial nitric oxide synthase: Molecular cloning and characterization of a distinct constitutive enzyme isoform. Proc. Natl. Acad. Sci. USA 1992, 89, 6348–6352. [Google Scholar] [CrossRef]
- Laing, K.J.; Grabowski, P.S.; Belosevic, M.; Secombes, C.J. A partial sequence for nitric oxide synthase from a goldfish (Carassius auratus) macrophage cell line. Immunol. Cell Biol. 1996, 74, 374–379. [Google Scholar] [CrossRef]
- Wang, T.; Ward, M.; Grabowski, P.; Secombes, C.J. Molecular cloning, gene organization and expression of rainbow trout (Oncorhynchus mykiss) inducible nitric oxide synthase (iNOS) gene. Biochem. J. 2001, 358, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Saeij, J.P.; Stet, R.J.; Groeneveld, A.; Verburg-van Kemenade, L.B.; van Muiswinkel, W.B.; Wiegertjes, G.F. Molecular and functional characterization of a fish inducible-type nitric oxide synthase. Immunogenetics 2000, 51, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Oyan, A.M.; Nilsen, F.; Goksøyr, A.; Holmqvist, B. Partial cloning of constitutive and inducible nitric oxide synthases and detailed neuronal expression of NOS mRNA in the cerebellum and optic tectum of adult Atlantic salmon (Salmo salar). Brain Res. Mol. Brain Res. 2000, 78, 38–49. [Google Scholar] [CrossRef]
- Carriero, M.M.; Henrique-Silva, F.; Caetano, A.R.; Lobo, F.P.; Alves, A.L.; Varela, E.S.; Del Collado, M.; Moreira, G.S.A.; Maia, A.A.M. Characterization and gene expression analysis of pacu (Piaractus mesopotamicus) inducible nitric oxide synthase (iNOS) following Aeromonas dhakensis infection. Fish Shellfish. Immunol. 2018, 74, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Li, C.; Zhang, J.; Liu, S.; Feng, J.; Wang, R.; Li, Y.; Jiang, C.; Song, L.; Chen, A.; et al. Expression of nitric oxide synthase (NOS) genes in channel catfish is highly regulated and time dependent after bacterial challenges. Dev. Comp. Immunol. 2014, 45, 74–86. [Google Scholar] [CrossRef]
- Donald, J.A.; Forgan, L.G.; Cameron, M.S. The evolution of nitric oxide signalling in vertebrate blood vessels. J. Comp. Physiol. B 2015, 185, 153–171. [Google Scholar] [CrossRef]
- Trajanovska, S.; Donald, J.A. Endothelial nitric oxide synthase in the amphibian, Xenopus tropicalis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 274–281. [Google Scholar] [CrossRef]
- González-Domenech, C.M.; Muñoz-Chápuli, R. Molecular evolution of nitric oxide synthases in metazoans. Comp. Biochem. Physiol. Part D Genom. Proteom. 2010, 5, 295–301. [Google Scholar] [CrossRef]
- Olson, K.; Donald, J. Nervous control of circulation—The role of gasotransmitters, NO, CO, and H2S. Acta Histochem. 2009, 111, 244–256. [Google Scholar] [CrossRef]
- Holmqvist, B.; Ebbesson, L.; Alm, P. Nitric oxide and the zebrafish (Danio rerio): Developmental neurobiology and brain neurogenesis. Adv. Exp. Biol. 2007, 1, 229–465. [Google Scholar] [CrossRef]
- Amelio, D.; Garofalo, F.; Pellegrino, D.; Giordano, F.; Tota, B.; Cerra, M.C. Cardiac expression and distribution of nitric oxide synthases in the ventricle of the cold-adapted Antarctic teleosts, the hemoglobinless Chionodraco hamatus and the red-blooded Trematomus bernacchii. Nitric Oxide Biol. Chem. 2006, 15, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Imbrogno, S.; Garofalo, F.; Cerra, M.C.; Mahata, S.K.; Tota, B. The catecholamine release-inhibitory peptide catestatin (chromogranin A344-363) modulates myocardial function in fish. J. Exp. Biol. 2010, 213, 3636–3643. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tota, B.; Amelio, D.; Cerra, M.C.; Garofalo, F. The morphological and functional significance of the NOS/NO system in the respiratory, osmoregulatory, and contractile organs of the African lungfish. Acta Histochem. 2018, 120, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Amelio, D.; Garofalo, F.; Wong, W.P.; Chew, S.F.; Ip, Y.K.; Cerra, M.C.; Tota, B. Nitric oxide synthase-dependent “on/off” switch and apoptosis in freshwater and aestivating lungfish, Protopterus annectens: Skeletal muscle versus cardiac muscle. Nitric Oxide Biol. Chem. 2013, 32, 1–12. [Google Scholar] [CrossRef]
- Amelio, D.; Garofalo, F.; Brunelli, E.; Loong, A.M.; Wong, W.P.; Ip, Y.K.; Tota, B.; Cerra, M.C. Differential NOS expression in freshwater and aestivating Protopterus dolloi (lungfish): Heart vs kidney readjustments. Nitric Oxide Biol. Chem. 2008, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, R.; Schwerte, T.; Pelster, B. Nitric oxide and vascular reactivity in developing zebrafish, Danio rerio. Am. J. Physiol. Integr. Comp. Physiol. 2000, 279, R2200–R2207. [Google Scholar] [CrossRef]
- Imbrogno, S.; Capria, C.; Tota, B.; Jensen, F.B. Nitric oxide improves the hemodynamic performance of the hypoxic goldfish (Carassius auratus) heart. Nitric Oxide Biol. Chem. 2014, 42, 24–31. [Google Scholar] [CrossRef]
- Pellegrino, D.; Palmerini, C.A.; Tota, B. No hemoglobin but NO: The icefish (Chionodraco hamatus) heart as a paradigm. J. Exp. Biol. 2004, 207, 3855–3864. [Google Scholar] [CrossRef][Green Version]
- Garofalo, F.; Amelio, D.; Cerra, M.C.; Tota, B.; Sidell, B.D.; Pellegrino, D. Morphological and physiological study of the cardiac NOS/NO system in the Antarctic (Hb-/Mb-) icefish Chaenocephalus aceratus and in the red-blooded Trematomus bernacchii. Nitric Oxide Biol. Chem. 2009, 20, 69–78. [Google Scholar] [CrossRef]
- Morlà, M.; Agustì, G.N.A.; Rahman, I.; Motterlini, R.; Saus, C.; Morales-Nin, B.; Company, J.B.; Busquets, X. Nitric oxide synthase type I (nNOS), vascular endothelial growth factor (VEGF) and myoglobin-like expression in skeletal muscle of Antarctic icefishes (Notothenioidei: Channichthyidae). Polar Biol. 2003, 26, 458–462. [Google Scholar] [CrossRef]
- Garofalo, F.; Pellegrino, D.; Amelio, D.; Tota, B. The Antarctic hemoglobinless icefish, fifty five years later: A unique cardiocirculatory interplay of disaptation and phenotypic plasticity. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 154, 10–28. [Google Scholar] [CrossRef] [PubMed]
- Giulivi, C. Characterization and function of mitochondrial nitric-oxide synthase. Free Radic. Biol. Med. 2003, 34, 397–408. [Google Scholar] [CrossRef]
- Tatsumi, T.; Matoba, S.; Kawahara, A.; Keira, N.; Shiraishi, J.; Akashi, K.; Kobara, M.; Tanaka, T.; Katamura, M.; Nakagawa, C.; et al. Cytokine-induced nitric oxide production inhibits mitochondrial energy production and impairs contractile function in rat cardiac myocytes. J. Am. Coll. Cardiol. 2000, 35, 1338–1346. [Google Scholar] [CrossRef]
- Garofalo, F.; Santovito, G.; Amelio, D. Morpho-functional effects of heat stress on the gills of Antarctic T. bernacchii and C. hamatus. Mar. Pollut. Bull. 2019, 141, 194–204. [Google Scholar] [CrossRef]
- Amelio, D.; Garofalo, F.; Capria, C.; Tota, B.; Imbrogno, S. Effects of temperature on the nitric oxide-dependent modulation of the Frank-Starling mechanism: The fish heart as a case study. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 164, 356–362. [Google Scholar] [CrossRef]
- Jørgensen, S.M.; Castro, V.; Krasnov, A.; Torgersen, J.; Timmerhaus, G.; Hevrøy, E.M.; Hansen, T.J.; Susort, S.; Breck, O.; Takle, H. Cardiac responses to elevated seawater temperature in Atlantic salmon. BMC Physiol. 2014, 14, 2. [Google Scholar] [CrossRef]
- Palmer, L.A.; Semenza, G.L.; Stoler, M.H.; Johns, R.A. Hypoxia induces type II NOS gene expression in pulmonary artery endothelial cells via HIF-1. Am. J. Physiol. 1998, 274, L212–L219. [Google Scholar] [CrossRef]
- Melillo, G.; Musso, T.; Sica, A.; Taylor, L.S.; Cox, G.W.; Varesio, L. A hypoxia-responsive element mediates a novel pathway of activation of the inducible nitric oxide synthase promoter. J. Exp. Med. 1995, 182, 1683–1693. [Google Scholar] [CrossRef]
- Coulet, F.; Nadaud, S.; Agrapart, M.; Soubrier, F. Identification of hypoxia-response element in the human endothelial nitric-oxide synthase gene promoter. J. Biol. Chem. 2003, 278, 46230–46240. [Google Scholar] [CrossRef]
- Angelone, T.; Quintieri, A.M.; Pasqua, T.; Filice, E.; Cantafio, P.; Scavello, F.; Rocca, C.; Mahata, S.K.; Gattuso, A.; Cerra, M.C. The NO stimulator, Catestatin, improves the Frank-Starling response in normotensive and hypertensive rat hearts. Nitric Oxide Biol. Chem. 2015, 50, 10–19. [Google Scholar] [CrossRef]
- McNeill, B.; Perry, S.F. The interactive effects of hypoxia and nitric oxide on catecholamine secretion in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 2006, 209, 4214–4223. [Google Scholar] [CrossRef] [PubMed]
- Shiva, S.; Gladwin, M.T. Nitrite mediates cytoprotection after ischemia/reperfusion by modulating mitochondrial function. Basic Res. Cardiol. 2009, 104, 113. [Google Scholar] [CrossRef] [PubMed]
- Gladwin, M.T. Haldane, hot dogs, halitosis, and hypoxic vasodilation: The emerging biology of the nitrite anion. J. Clin. Investig. 2004, 113, 19–21. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Corti, P.; Xue, J.; Tejero, J.; Wajih, N.; Sun, M.; Stolz, D.B.; Tsang, M.; Kim-Shapiro, D.B.; Gladwin, M.T. Globin X is a six-coordinate globin that reduces nitrite to nitric oxide in fish red blood cells. Proc. Natl. Acad. Sci. USA 2016, 113, 8538–8543. [Google Scholar] [CrossRef] [PubMed]
- Gladwin, M.T.; Shelhamer, J.H.; Schechter, A.N.; Pease-Fye, M.E.; Waclawiw, M.A.; Panza, J.A.; Ognibene, F.P.; Cannon, R.O. Role of circulating nitrite and S-nitrosohemoglobin in the regulation of regional blood flow in humans. Proc. Natl. Acad. Sci. USA 2000, 97, 11482–11487. [Google Scholar] [CrossRef]
- Dejam, A.; Hunter, C.J.; Schechter, A.N.; Gladwin, M.T. Emerging role of nitrite in human biology. Blood Cells. Mol. Dis. 2004, 32, 423–429. [Google Scholar] [CrossRef]
- Tejero, J.; Gladwin, M.T. The globin superfamily: Functions in nitric oxide formation and decay. Biol. Chem. 2014, 395, 631–639. [Google Scholar] [CrossRef]
- Liu, X.; Tong, J.; Zweier, J.R.; Follmer, D.; Hemann, C.; Ismail, R.S.; Zweier, J.L. Differences in oxygen-dependent nitric oxide metabolism by cytoglobin and myoglobin account for their differing functional roles. FEBS J. 2013, 280, 3621–3631. [Google Scholar] [CrossRef]
- Li, H.; Samouilov, A.; Liu, X.; Zweier, J.L. Characterization of the Magnitude and Kinetics of Xanthine Oxidase-catalyzed Nitrite Reduction: Evaluation of Its role in nitric oxide generation in anoxic tissues. J. Biol. Chem. 2001, 276, 24482–24489. [Google Scholar] [CrossRef]
- Jensen, F.B. The role of nitrite in nitric oxide homeostasis: A comparative perspective. Biochim. Biophys. Acta Bioenerg. 2009, 1787, 841–848. [Google Scholar] [CrossRef]
- Jensen, F.B. Nitrite disrupts multiple physiological functions in aquatic animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 135, 9–24. [Google Scholar] [CrossRef]
- Hansen, M.N.; Gerber, L.; Jensen, F.B. Nitric oxide availability in deeply hypoxic crucian carp: Acute and chronic changes and utilization of ambient nitrite reservoirs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R532–R540. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.N.; Lundberg, J.O.; Filice, M.; Fago, A.; Christensen, N.M.G.; Jensen, F.B. The roles of tissue nitrate reductase activity and myoglobin in securing nitric oxide availability in deeply hypoxic crucian carp. J. Exp. Biol. 2016, 219, 3875–3883. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.N.; Jensen, F.B. Nitric oxide metabolites in goldfish under normoxic and hypoxic conditions. J. Exp. Biol. 2010, 213, 3593–3602. [Google Scholar] [CrossRef] [PubMed]
- Fago, A.; Jensen, F.B. Hypoxia tolerance, nitric oxide, and nitrite: Lessons from extreme animals. Physiology 2015, 30, 116–126. [Google Scholar] [CrossRef]
- Sandvik, G.K.; Nilsson, G.E.; Jensen, F.B. Dramatic increase of nitrite levels in hearts of anoxia-exposed crucian carp supporting a role in cardioprotection. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R468–R477. [Google Scholar] [CrossRef][Green Version]
- Gattuso, A.; Garofalo, F.; Cerra, M.C.; Imbrogno, S. Hypoxia tolerance in teleosts: Implications of cardiac nitrosative signals. Front. Physiol. 2018, 9, 366. [Google Scholar] [CrossRef]
- Garofalo, F.; Amelio, D.; Gattuso, A.; Cerra, M.C.; Pellegrino, D. Cardiac contractility in Antarctic teleost is modulated by nitrite through xanthine oxidase and cytochrome p-450 nitrite reductase. Nitric Oxide Biol. Chem. 2015, 49, 1–7. [Google Scholar] [CrossRef]
- Rochon, E.R.; Missinato, M.A.; Xue, J.; Tejero, J.; Tsang, M.; Gladwin, M.T.; Corti, P. Nitrite Improves Heart Regeneration in Zebrafish. Antioxid. Redox Signal. 2020, 32, 363–377. [Google Scholar] [CrossRef]
- Keppner, A.; Maric, D.; Correia, M.; Koay, T.W.; Orlando, I.M.C.; Vinogradov, S.N.; Hoogewijs, D. Lessons from the post-genomic era: Globin diversity beyond oxygen binding and transport. Redox Biol. 2020, 37, 101687. [Google Scholar] [CrossRef]
- Rassaf, T.; Flögel, U.; Drexhage, C.; Hendgen-Cotta, U.; Kelm, M.; Schrader, J. Nitrite reductase function of deoxymyoglobin: Oxygen sensor and regulator of cardiac energetics and function. Circ. Res. 2007, 100, 1749–1754. [Google Scholar] [CrossRef] [PubMed]
- Grubina, R.; Huang, Z.; Shiva, S.; Joshi, M.S.; Azarov, I.; Basu, S.; Ringwood, L.A.; Jiang, A.; Hogg, N.; Kim-Shapiro, D.B.; et al. Concerted nitric oxide formation and release from the simultaneous reactions of nitrite with deoxy- and oxyhemoglobin. J. Biol. Chem. 2007, 282, 12916–12927. [Google Scholar] [CrossRef] [PubMed]
- Keszler, A.; Brandal, G.; Baumgardt, S.; Ge, Z.-D.; Pratt, P.F.; Riess, M.L.; Bienengraeber, M. Far red/near infrared light-induced protection against cardiac ischemia and reperfusion injury remains intact under diabetic conditions and is independent of nitric oxide synthase. Front. Physiol. 2014, 5, 305. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Shiva, S.; Kim-Shapiro, D.B.; Patel, R.P.; Ringwood, L.A.; Irby, C.E.; Huang, K.T.; Ho, C.; Hogg, N.; Schechter, A.N.; et al. Enzymatic function of hemoglobin as a nitrite reductase that produces NO under allosteric control. J. Clin. Investig. 2005, 115, 2099–2107. [Google Scholar] [CrossRef] [PubMed]
- Hendgen-Cotta, U.B.; Merx, M.W.; Shiva, S.; Schmitz, J.; Becher, S.; Klare, J.P.; Steinhoff, H.-J.; Goedecke, A.; Schrader, J.; Gladwin, M.T.; et al. Nitrite reductase activity of myoglobin regulates respiration and cellular viability in myocardial ischemia-reperfusion injury. Proc. Natl. Acad. Sci. USA 2008, 105, 10256–10261. [Google Scholar] [CrossRef]
- Reeder, B.J.; Ukeri, J. Strong modulation of nitrite reductase activity of cytoglobin by disulfide bond oxidation: Implications for nitric oxide homeostasis. Nitric Oxide Biol. Chem. 2018, 72, 16–23. [Google Scholar] [CrossRef]
- Tiso, M.; Tejero, J.; Basu, S.; Azarov, I.; Wang, X.; Simplaceanu, V.; Frizzell, S.; Jayaraman, T.; Geary, L.; Shapiro, C.; et al. Human neuroglobin functions as a redox-regulated nitrite reductase. J. Biol. Chem. 2011, 286, 18277–18289. [Google Scholar] [CrossRef]
- Götting, M.; Nikinmaa, M. More than hemoglobin—The unexpected diversity of globins in vertebrate red blood cells. Physiol. Rep. 2015, 3, e12284. [Google Scholar] [CrossRef]
- Corti, P.; Ieraci, M.; Tejero, J. Characterization of zebrafish neuroglobin and cytoglobins 1 and 2: Zebrafish cytoglobins provide insights into the transition from six-coordinate to five-coordinate globins. Nitric Oxide 2016, 53, 22–34. [Google Scholar] [CrossRef]
- Amdahl, M.B.; Petersen, E.E.; Bocian, K.; Kaliszuk, S.J.; DeMartino, A.W.; Tiwari, S.; Sparacino-Watkins, C.E.; Corti, P.; Rose, J.J.; Gladwin, M.T.; et al. The Zebrafish Cytochrome b(5)/Cytochrome b(5) Reductase/NADH System Efficiently Reduces Cytoglobins 1 and 2: Conserved Activity of Cytochrome b(5)/Cytochrome b(5) Reductases during Vertebrate Evolution. Biochemistry 2019, 58, 3212–3223. [Google Scholar] [CrossRef]
- Amdahl, M.B.; Sparacino-Watkins, C.E.; Corti, P.; Gladwin, M.T.; Tejero, J. Efficient Reduction of Vertebrate Cytoglobins by the Cytochrome b(5)/Cytochrome b(5) Reductase/NADH System. Biochemistry 2017, 56, 3993–4004. [Google Scholar] [CrossRef] [PubMed]
- Hoogewijs, D.; Ebner, B.; Germani, F.; Hoffmann, F.G.; Fabrizius, A.; Moens, L.; Burmester, T.; Dewilde, S.; Storz, J.F.; Vinogradov, S.N.; et al. Androglobin: A chimeric globin in metazoans that is preferentially expressed in Mammalian testes. Mol. Biol. Evol. 2012, 29, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Burmester, T.; Ebner, B.; Weich, B.; Hankeln, T. Cytoglobin: A novel globin type ubiquitously expressed in vertebrate tissues. Mol. Biol. Evol. 2002, 19, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Kugelstadt, D.; Haberkamp, M.; Hankeln, T.; Burmester, T. Neuroglobin, cytoglobin, and a novel, eye-specific globin from chicken. Biochem. Biophys. Res. Commun. 2004, 325, 719–725. [Google Scholar] [CrossRef]
- Fuchs, C.; Burmester, T.; Hankeln, T. The amphibian globin gene repertoire as revealed by the Xenopus genome. Cytogenet. Genome Res. 2006, 112, 296–306. [Google Scholar] [CrossRef]
- Roesner, A.; Fuchs, C.; Hankeln, T.; Burmester, T. A globin gene of ancient evolutionary origin in lower vertebrates: Evidence for two distinct globin families in animals. Mol. Biol. Evol. 2005, 22, 12–20. [Google Scholar] [CrossRef]
- Burmester, T.; Weich, B.; Reinhardt, S.; Hankeln, T. A vertebrate globin expressed in the brain. Nature 2000, 407, 520–523. [Google Scholar] [CrossRef]
- Gallagher, M.; Macqueen, D. Evolution and Expression of Tissue Globins in Ray-Finned Fishes. Genome Biol. Evol. 2017, 9, 32–47. [Google Scholar] [CrossRef]
- Trent, J.T.; Hvitved, A.N.; Hargrove, M.S. A Model for Ligand Binding to Hexacoordinate Hemoglobins. Biochemistry 2001, 40, 6155–6163. [Google Scholar] [CrossRef]
- de Sanctis, D.; Dewilde, S.; Pesce, A.; Moens, L.; Ascenzi, P.; Hankeln, T.; Burmester, T.; Bolognesi, M. Crystal Structure of Cytoglobin: The Fourth Globin Type Discovered in Man Displays Heme Hexa-coordination. J. Mol. Biol. 2004, 336, 917–927. [Google Scholar] [CrossRef]
- Dewilde, S.; Kiger, L.; Burmester, T.; Hankeln, T.; Baudin-Creuza, V.; Aerts, T.; Marden, M.C.; Caubergs, R.; Moens, L. Biochemical characterization and ligand binding properties of neuroglobin, a novel member of the globin family. J. Biol. Chem. 2001, 276, 38949–38955. [Google Scholar] [CrossRef] [PubMed]
- Vitagliano, L.; Vergara, A.; Bonomi, G.; Merlino, A.; Verde, C.; di Prisco, G.; Howes, B.D.; Smulevich, G.; Mazzarella, L. Spectroscopic and Crystallographic Characterization of a Tetrameric Hemoglobin Oxidation Reveals Structural Features of the Functional Intermediate Relaxed/Tense State. J. Am. Chem. Soc. 2008, 130, 10527–10535. [Google Scholar] [CrossRef] [PubMed]
- Antonio, R.; Luigi, V.; di Guido, P.; Adriana, Z.; Lelio, M. The crystal structure of a tetrameric hemoglobin in a partial hemichrome state. Proc. Natl. Acad. Sci. USA 2002, 99, 9801–9806. [Google Scholar] [CrossRef]
- Jensen, F.B. Red blood cell pH, the Bohr effect, and other oxygenation-linked phenomena in blood O2 and CO2 transport. Acta Physiol. Scand. 2004, 182, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Jensen, F.B.; Kolind, R.A.H.; Jensen, N.S.; Montesanti, G.; Wang, T. Interspecific variation and plasticity in hemoglobin nitrite reductase activity and its correlation with oxygen affinity in vertebrates. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 206, 47–53. [Google Scholar] [CrossRef]
- Jensen, F.B. Nitric oxide formation from the reaction of nitrite with carp and rabbit hemoglobin at intermediate oxygen saturations. FEBS J. 2008, 275, 3375–3387. [Google Scholar] [CrossRef]
- Suzuki, T.; Imai, K. Evolution of myoglobin. Cell. Mol. Life Sci. 1998, 54, 979–1004. [Google Scholar] [CrossRef]
- Fraser, J.; de Mello, L.V.; Ward, D.; Rees, H.H.; Williams, D.R.; Fang, Y.; Brass, A.; Gracey, A.Y.; Cossins, A.R. Hypoxia-inducible myoglobin expression in nonmuscle tissues. Proc. Natl. Acad. Sci. USA 2006, 103, 2977–2981. [Google Scholar] [CrossRef]
- Roesner, A.; Mitz, S.A.; Hankeln, T.; Burmester, T. Globins and hypoxia adaptation in the goldfish, Carassius auratus. FEBS J. 2008, 275, 3633–3643. [Google Scholar] [CrossRef]
- Cossins, A.R.; Williams, D.R.; Foulkes, N.S.; Berenbrink, M.; Kipar, A. Diverse cell-specific expression of myoglobin isoforms in brain, kidney, gill and liver of the hypoxia-tolerant carp and zebrafish. J. Exp. Biol. 2009, 212, 627–638. [Google Scholar] [CrossRef]
- Brunori, M. Nitric oxide, cytochrome-c oxidase and myoglobin. Trends Biochem. Sci. 2001, 26, 21–23. [Google Scholar] [CrossRef]
- Moncada, S.; Rees, D.D.; Schulz, R.; Palmer, R.M. Development and mechanism of a specific supersensitivity to nitrovasodilators after inhibition of vascular nitric oxide synthesis in vivo. Proc. Natl. Acad. Sci. USA 1991, 88, 2166–2170. [Google Scholar] [CrossRef]
- Lowenstein, C.J.; Dinerman, J.L.; Snyder, S.H. Nitric Oxide: A Physiologic Messenger. Ann. Intern. Med. 1994, 120, 227–237. [Google Scholar] [CrossRef]
- Shiva, S.; Huang, Z.; Grubina, R.; Sun, J.; Ringwood, L.A.; MacArthur, P.H.; Xu, X.; Murphy, E.; Darley-Usmar, V.M.; Gladwin, M.T. Deoxymyoglobin is a nitrite reductase that generates nitric oxide and regulates mitochondrial respiration. Circ. Res. 2007, 100, 654–661. [Google Scholar] [CrossRef]
- Beers, J.M.; Borley, K.A.; Sidell, B.D. Relationship among circulating hemoglobin, nitric oxide synthase activities and angiogenic poise in red- and white-blooded Antarctic notothenioid fishes. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 422–429. [Google Scholar] [CrossRef]
- Kooyman, G.L.; Ponganis, P.J. The physiological basis of diving to depth: Birds and mammals. Annu. Rev. Physiol. 1998, 60, 19–32. [Google Scholar] [CrossRef]
- Kawada, N.; Kristensen, D.B.; Asahina, K.; Nakatani, K.; Minamiyama, Y.; Seki, S.; Yoshizato, K. Characterization of a stellate cell activation-associated protein (STAP) with peroxidase activity found in rat hepatic stellate cells. J. Biol. Chem. 2001, 276, 25318–25323. [Google Scholar] [CrossRef]
- Hankeln, T.; Ebner, B.; Fuchs, C.; Gerlach, F.; Haberkamp, M.; Laufs, T.L.; Roesner, A.; Schmidt, M.; Weich, B.; Wystub, S.; et al. Neuroglobin and cytoglobin in search of their role in the vertebrate globin family. J. Inorg. Biochem. 2005, 99, 110–119. [Google Scholar] [CrossRef]
- Halligan, K.E.; Jourd’heuil, F.L.; Jourd’heuil, D. Cytoglobin Is Expressed in the Vasculature and Regulates Cell Respiration and Proliferation via Nitric Oxide Dioxygenation. J. Biol. Chem. 2009, 284, 8539–8547. [Google Scholar] [CrossRef]
- Li, H.; Hemann, C.; Abdelghany, T.M.; El-Mahdy, M.A.; Zweier, J.L. Characterization of the Mechanism and Magnitude of Cytoglobin-mediated Nitrite Reduction and Nitric Oxide Generation under Anaerobic Conditions. J. Biol. Chem. 2012, 287, 36623–36633. [Google Scholar] [CrossRef]
- Liu, X.; Follmer, D.; Zweier, J.R.; Huang, X.; Hemann, C.; Liu, K.; Druhan, L.J.; Zweier, J.L. Characterization of the function of cytoglobin as an oxygen-dependent regulator of nitric oxide concentration. Biochemistry 2012, 51, 5072–5082. [Google Scholar] [CrossRef]
- Li, D.; Chen, X.Q.; Li, W.-J.; Yang, Y.-H.; Wang, J.-Z.; Yu, A.C.H. Cytoglobin Up-regulated by Hydrogen Peroxide Plays a Protective Role in Oxidative Stress. Neurochem. Res. 2007, 32, 1375–1380. [Google Scholar] [CrossRef]
- Fuchs, C.; Luckhardt, A.; Gerlach, F.; Burmester, T.; Hankeln, T. Duplicated cytoglobin genes in teleost fishes. Biochem. Biophys. Res. Commun. 2005, 337, 216–223. [Google Scholar] [CrossRef]
- Montfort, W.R.; Wales, J.A.; Weichsel, A. Structure and Activation of Soluble Guanylyl Cyclase, the Nitric Oxide Sensor. Antioxid. Redox Signal. 2017, 26, 107–121. [Google Scholar] [CrossRef]
- Poulos, T.L. Soluble guanylate cyclase. Curr. Opin. Struct. Biol. 2006, 16, 736–743. [Google Scholar] [CrossRef]
- Padayatti, P.S.; Pattanaik, P.; Ma, X.; van den Akker, F. Structural insights into the regulation and the activation mechanism of mammalian guanylyl cyclases. Pharmacol. Ther. 2004, 104, 83–99. [Google Scholar] [CrossRef]
- Derbyshire, E.R.; Marletta, M.A. Structure and regulation of soluble guanylate cyclase. Annu. Rev. Biochem. 2012, 81, 533–559. [Google Scholar] [CrossRef]
- Francis, S.H.; Busch, J.L.; Corbin, J.D. cGMP-Dependent Protein Kinases and cGMP Phosphodiesterases in Nitric Oxide and cGMP Action. Pharmacol. Rev. 2010, 62, 525–563. [Google Scholar] [CrossRef]
- Rippe, C.; Zhu, B.; Krawczyk, K.K.; Van Bavel, E.; Albinsson, S.; Sjölund, J.; Bakker, E.N.T.P.; Swärd, K. Hypertension reduces soluble guanylyl cyclase expression in the mouse aorta via the Notch signaling pathway. Sci. Rep. 2017, 7, 1334. [Google Scholar] [CrossRef]
- Harumi, T.; Watanabe, T.; Yamamoto, T.; Tanabe, Y.; Suzuki, N. Expression of membrane-bound and soluble guanylyl cyclase mRNAs in embryonic and adult retina of the medaka fish Oryzias latipes. Zool. Sci. 2003, 20, 133–140. [Google Scholar] [CrossRef][Green Version]
- Saraiva, L.R.; Ahuja, G.; Ivandic, I.; Syed, A.S.; Marioni, J.C.; Korsching, S.I.; Logan, D.W. Molecular and neuronal homology between the olfactory systems of zebrafish and mouse. Sci. Rep. 2015, 5, 11487. [Google Scholar] [CrossRef]
- Wittmann, C.; Reischl, M.; Shah, A.H.; Kronfuss, E.; Mikut, R.; Liebel, U.; Grabher, C. A Zebrafish Drug-Repurposing Screen Reveals sGC-Dependent and sGC-Independent Pro-Inflammatory Activities of Nitric Oxide. PLoS ONE 2015, 10, e0137286. [Google Scholar] [CrossRef]
- Zhao, Y.; Brandish, P.E.; DiValentin, M.; Schelvis, J.P.M.; Babcock, G.T.; Marletta, M.A. Inhibition of Soluble Guanylate Cyclase by ODQ. Biochemistry 2000, 39, 10848–10854. [Google Scholar] [CrossRef]
- Vishnolia, K.K.; Rakovic, A.; Hoene, C.; Tarhbalouti, K.; Aherrahrou, Z.; Erdmann, J. sGC Activity and Regulation of Blood Flow in a Zebrafish Model System. Front. Physiol. 2021, 12, 633171. [Google Scholar] [CrossRef]
- Xiyuan, Z.; Fink, R.H.A.; Mosqueira, M. NO-sGC Pathway Modulates Ca2+ Release and Muscle Contraction in Zebrafish Skeletal Muscle. Front. Physiol. 2017, 8, 607. [Google Scholar] [CrossRef]
- Hoenicka, M.; Becker, E.M.; Apeler, H.; Sirichoke, T.; Schröder, H.; Gerzer, R.; Stasch, J.P. Purified soluble guanylyl cyclase expressed in a baculovirus/Sf9 system: Stimulation by YC-1, nitric oxide, and carbon monoxide. J. Mol. Med. 1999, 77, 14–23. [Google Scholar] [CrossRef]
- Friebe, A.; Schultz, G.; Koesling, D. Sensitizing soluble guanylyl cyclase to become a highly CO-sensitive enzyme. EMBO J. 1996, 15, 6863–6868. [Google Scholar] [CrossRef]
- Stone, J.R.; Marletta, M.A. Synergistic activation of soluble guanylate cyclase by YC-1 and carbon monoxide: Implications for the role of cleavage of the iron-histidine bond during activation by nitric oxide. Chem. Biol. 1998, 5, 255–261. [Google Scholar] [CrossRef][Green Version]
- Follmann, M.; Griebenow, N.; Hahn, M.G.; Hartung, I.; Mais, F.-J.; Mittendorf, J.; Schäfer, M.; Schirok, H.; Stasch, J.-P.; Stoll, F.; et al. The chemistry and biology of soluble guanylate cyclase stimulators and activators. Angew. Chem. Int. Ed. Engl. 2013, 52, 9442–9462. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Zhou, W.; Li, X.; Chen, H. The role of PKG in oocyte maturation of zebrafish. Biochem. Biophys. Res. Commun. 2018, 505, 530–535. [Google Scholar] [CrossRef]
- Diesen, D.L.; Hess, D.T.; Stamler, J.S. Hypoxic Vasodilation by Red Blood Cells. Circ. Res. 2008, 103, 545–553. [Google Scholar] [CrossRef]
- Heinrich, T.A.; da Silva, R.S.; Miranda, K.M.; Switzer, C.H.; Wink, D.A.; Fukuto, J.M. Biological nitric oxide signalling: Chemistry and terminology. Br. J. Pharmacol. 2013, 169, 1417–1429. [Google Scholar] [CrossRef]
- Hess, D.T.; Matsumoto, A.; Kim, S.O.; Marshall, H.E.; Stamler, J.S. Protein S-nitrosylation: Purview and parameters. Nat. Rev. Mol. Cell Biol. 2005, 6, 150–166. [Google Scholar] [CrossRef]
- Jia, L.; Bonaventura, C.; Bonaventura, J.; Stamler, J.S. S-nitrosohaemoglobin: A dynamic activity of blood involved in vascular control. Nature 1996, 380, 221–226. [Google Scholar] [CrossRef]
- Stamler, J.S.; Simon, D.I.; Osborne, J.A.; Mullins, M.E.; Jaraki, O.; Michel, T.; Singel, D.J.; Loscalzo, J. S-nitrosylation of proteins with nitric oxide: Synthesis and characterization of biologically active compounds. Proc. Natl. Acad. Sci. USA 1992, 89, 444–448. [Google Scholar] [CrossRef]
- Pawloski, J.R.; Hess, D.T.; Stamler, J.S. Export by red blood cells of nitric oxide bioactivity. Nature 2001, 409, 622–626. [Google Scholar] [CrossRef]
- Reischl, E.; Dafre, A.L.; Franco, J.L.; Wilhelm Filho, D. Distribution, adaptation and physiological meaning of thiols from vertebrate hemoglobins. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 22–53. [Google Scholar] [CrossRef]
- Zhang, R.; Hess, D.T.; Qian, Z.; Hausladen, A.; Fonseca, F.; Chaube, R.; Reynolds, J.D.; Stamler, J.S. Hemoglobin βCys93 is essential for cardiovascular function and integrated response to hypoxia. Proc. Natl. Acad. Sci. USA 2015, 112, 6425–6430. [Google Scholar] [CrossRef]
- Sun, C.-W.; Yang, J.; Kleschyov, A.L.; Zhuge, Z.; Carlström, M.; Pernow, J.; Wajih, N.; Isbell, T.S.; Oh, J.-Y.; Cabrales, P.; et al. Hemoglobin β93 Cysteine Is Not Required for Export of Nitric Oxide Bioactivity from the Red Blood Cell. Circulation 2019, 139, 2654–2663. [Google Scholar] [CrossRef]
- Verde, C.; Vergara, A.; Giordano, D.; Mazzarella, L.; Di Prisco, G. The Root effect—A structural and evolutionary perspective. Antarct. Sci. 2007, 19, 271–278. [Google Scholar] [CrossRef]
- Bonaventura, J.; Lance, V.P. Nitric Oxide, Invertebrates and Hemoglobin1. Am. Zool. 2001, 41, 346–359. [Google Scholar] [CrossRef]
- Witting, P.K.; Douglas, D.J.; Mauk, A.G. Reaction of Human Myoglobin and Nitric Oxide: Heme iron or protein sulfhydryl (s) nitrosation dependence on the absence or presence of oxygen*. J. Biol. Chem. 2001, 276, 3991–3998. [Google Scholar] [CrossRef] [PubMed]
- Rayner, B.S.; Wu, B.-J.; Raftery, M.; Stocker, R.; Witting, P.K. Human S-Nitroso Oxymyoglobin Is a Store of Vasoactive Nitric Oxide. J. Biol. Chem. 2005, 280, 9985–9993. [Google Scholar] [CrossRef] [PubMed]
- Helbo, S.; Weber, R.E.; Fago, A. Expression patterns and adaptive functional diversity of vertebrate myoglobins. Biochim. Biophys. Acta Proteins Proteom. 2013, 1834, 1832–1839. [Google Scholar] [CrossRef]
- Schreiter, E.R.; Rodríguez, M.M.; Weichsel, A.; Montfort, W.R.; Bonaventura, J. S-Nitrosylation-induced Conformational Change in Blackfin Tuna Myoglobin *. J. Biol. Chem. 2007, 282, 19773–19780. [Google Scholar] [CrossRef]
- Helbo, S.; Gow, A.J.; Jamil, A.; Howes, B.D.; Smulevich, G.; Fago, A. Oxygen-linked S-nitrosation in fish myoglobins: A cysteine-specific tertiary allosteric effect. PLoS ONE 2014, 9, e97012. [Google Scholar] [CrossRef]
- Helbo, S.; Fago, A. Allosteric modulation by S-nitrosation in the low-O2 affinity myoglobin from rainbow trout. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R101–R108. [Google Scholar] [CrossRef]
- Petersen, M.G.; Dewilde, S.; Fago, A. Reactions of ferrous neuroglobin and cytoglobin with nitrite under anaerobic conditions. J. Inorg. Biochem. 2008, 102, 1777–1782. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | nos Genes Identified | Endothelial Activity | References |
|---|---|---|---|
| Danio rerio | nos1, nos2a, nos2b | + | [59,60,78] |
| Carassius auratus | nos1, nos2a, nos2ba, nos2bb | + | [59,60,79] |
| Cyprinus carpio | nos1, nos2a, nos2ba, nos2bb | nd 1 | [59,60] |
| Oncorhynchus mykiss | nos1, nos2a, nos2β | nd 1 | [59,60] |
| Salmo salar | nos1, nos2a, nos2β | nd 1 | [60] |
| Piaractus mesopotamicus | nos12, nos2 | nd 1 | [66] |
| Ictalurus punctatus | nos1, nos2.1, nos2.2 | nd 1 | [60,67] |
| Paramormyrops kingsleyae | nos1, nos2, nos3 | nd 1 | [60] |
| Takifugu poecilonotus | nos1 | nd 1 | [59] |
| Tetraodon nigroviridis | nos1 | nd 1 | [59] |
| Gasterosteus aculeatus | nos1 | nd 1 | [59] |
| Oryzias latipes | nos1 | nd 1 | [59] |
| Anguilla anguilla | nos1 | + | [74] |
| Thunnus thynnus thynnus | nos1 | + | [74] |
| Chionodraco hamatus | nos1 | + | [73] |
| Trematomus bernacchii | nos1 | + | [73] |
| Chaenocephalus aceratus | nos1 | + | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, D.; Verde, C.; Corti, P. Nitric Oxide Production and Regulation in the Teleost Cardiovascular System. Antioxidants 2022, 11, 957. https://doi.org/10.3390/antiox11050957
Giordano D, Verde C, Corti P. Nitric Oxide Production and Regulation in the Teleost Cardiovascular System. Antioxidants. 2022; 11(5):957. https://doi.org/10.3390/antiox11050957
Chicago/Turabian StyleGiordano, Daniela, Cinzia Verde, and Paola Corti. 2022. "Nitric Oxide Production and Regulation in the Teleost Cardiovascular System" Antioxidants 11, no. 5: 957. https://doi.org/10.3390/antiox11050957
APA StyleGiordano, D., Verde, C., & Corti, P. (2022). Nitric Oxide Production and Regulation in the Teleost Cardiovascular System. Antioxidants, 11(5), 957. https://doi.org/10.3390/antiox11050957