
Effect of Exogenous Melatonin Application on the Grain Yield and Antioxidant Capacity in Aromatic Rice under Combined Lead–Cadmium Stress


 ,
, 
Abstract
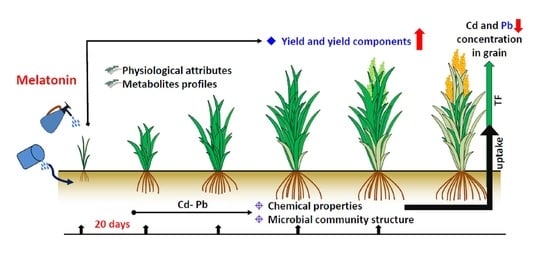
:
1. Introduction
2. Materials and Methods
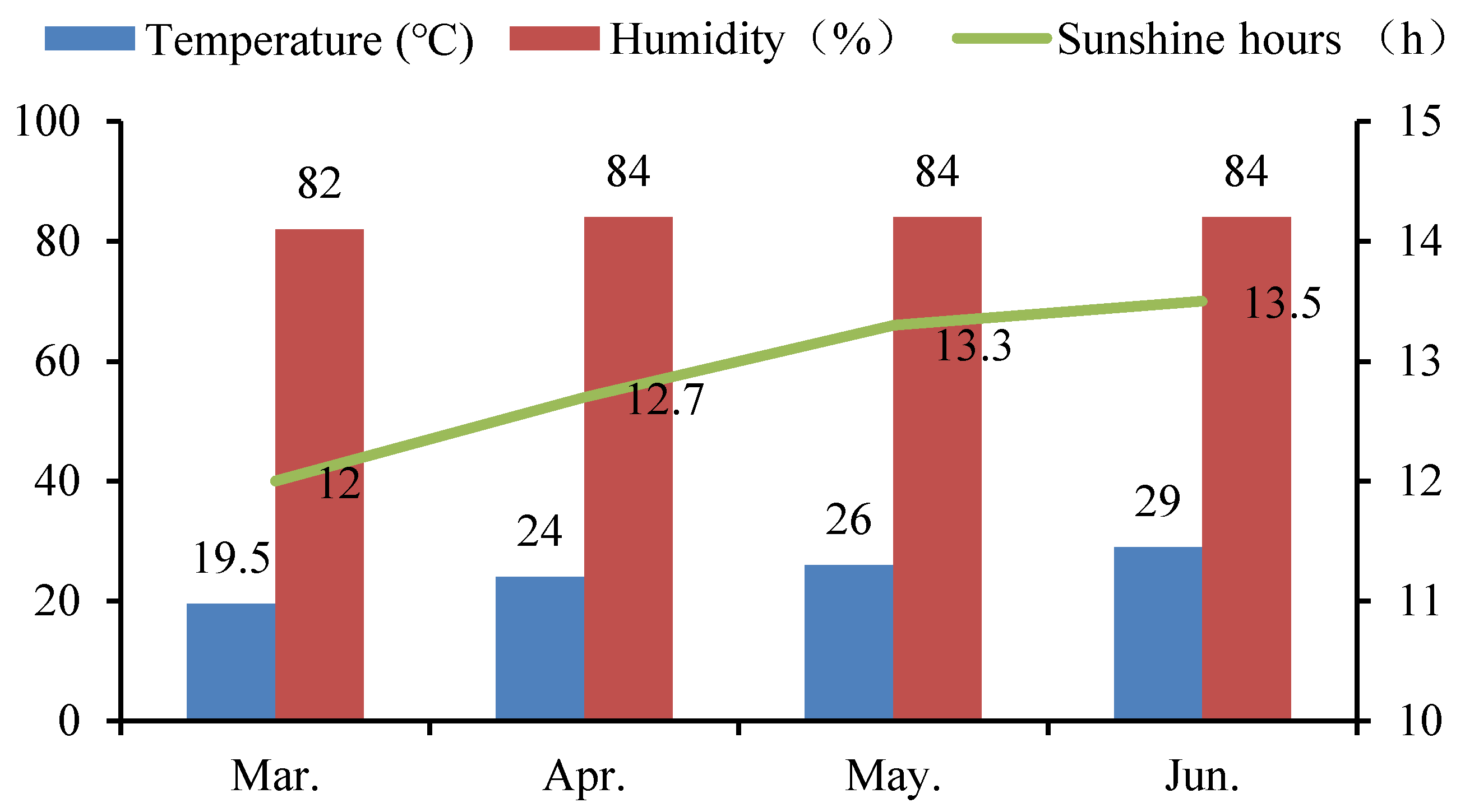
2.1. Experimental Design
2.2. Experimental Treatments
2.3. Sampling and Measurement
2.3.1. Determination of Cd and Pb Contents
2.3.2. Determination of Grain Yield and Yield Components
2.3.3. Determination of GSH, ASA, and MTs Contents
2.3.4. Determination of SOD, POD, and CAT Activities and MDA and H2O2 Contents
2.3.5. Determination of Soil Chemical Properties
2.3.6. Investigation of Microorganisms in the Rhizosphere
2.3.7. Determination of Primary Metabolites
2.4. Statistical Analyses
3. Results
3.1. Cd and Pb Accumulation and Translocation Factor
3.2. Grain Yield and Yield-Related Traits
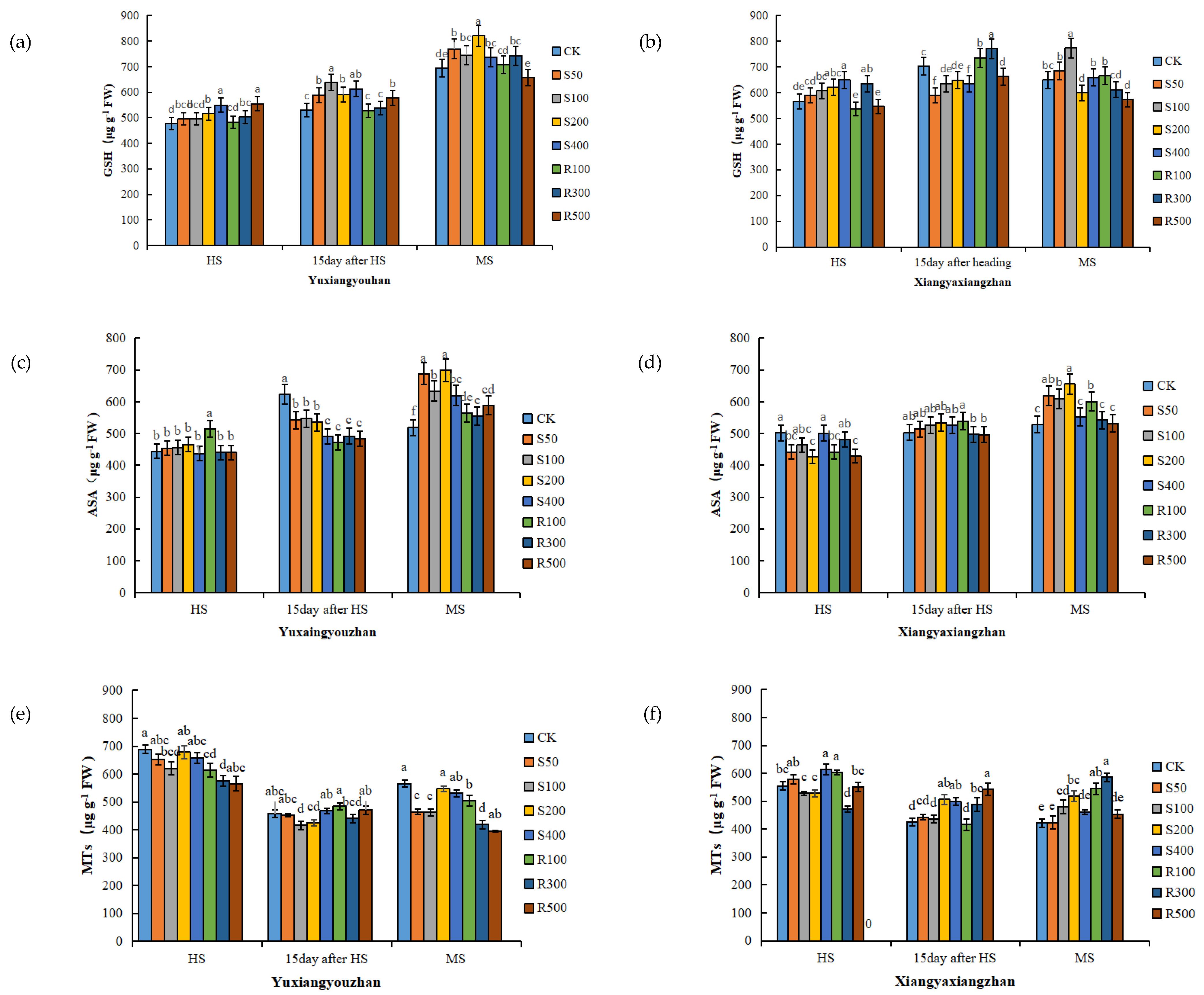
3.3. GSH, ASA, and MTs Contents in Leaves
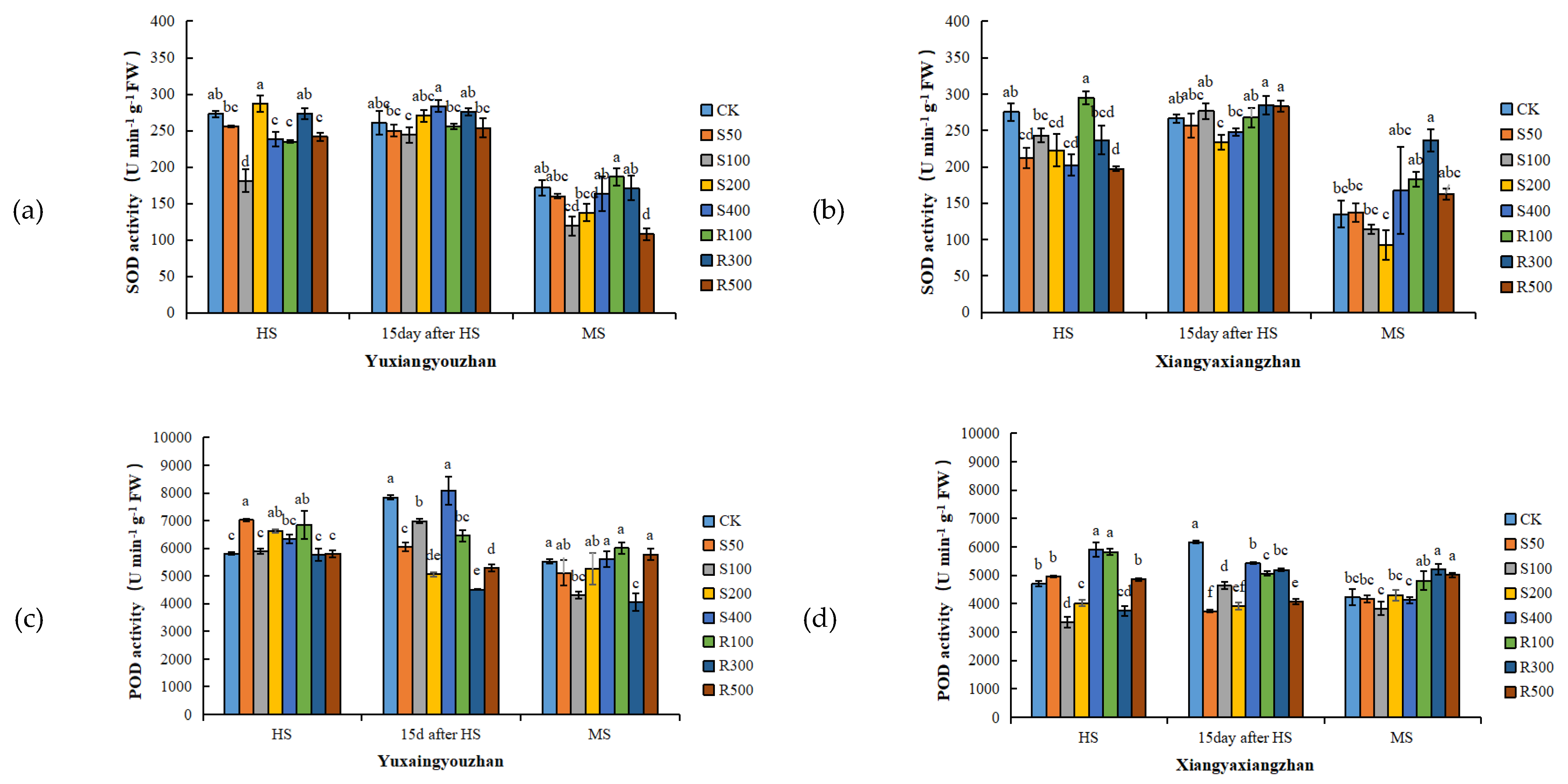
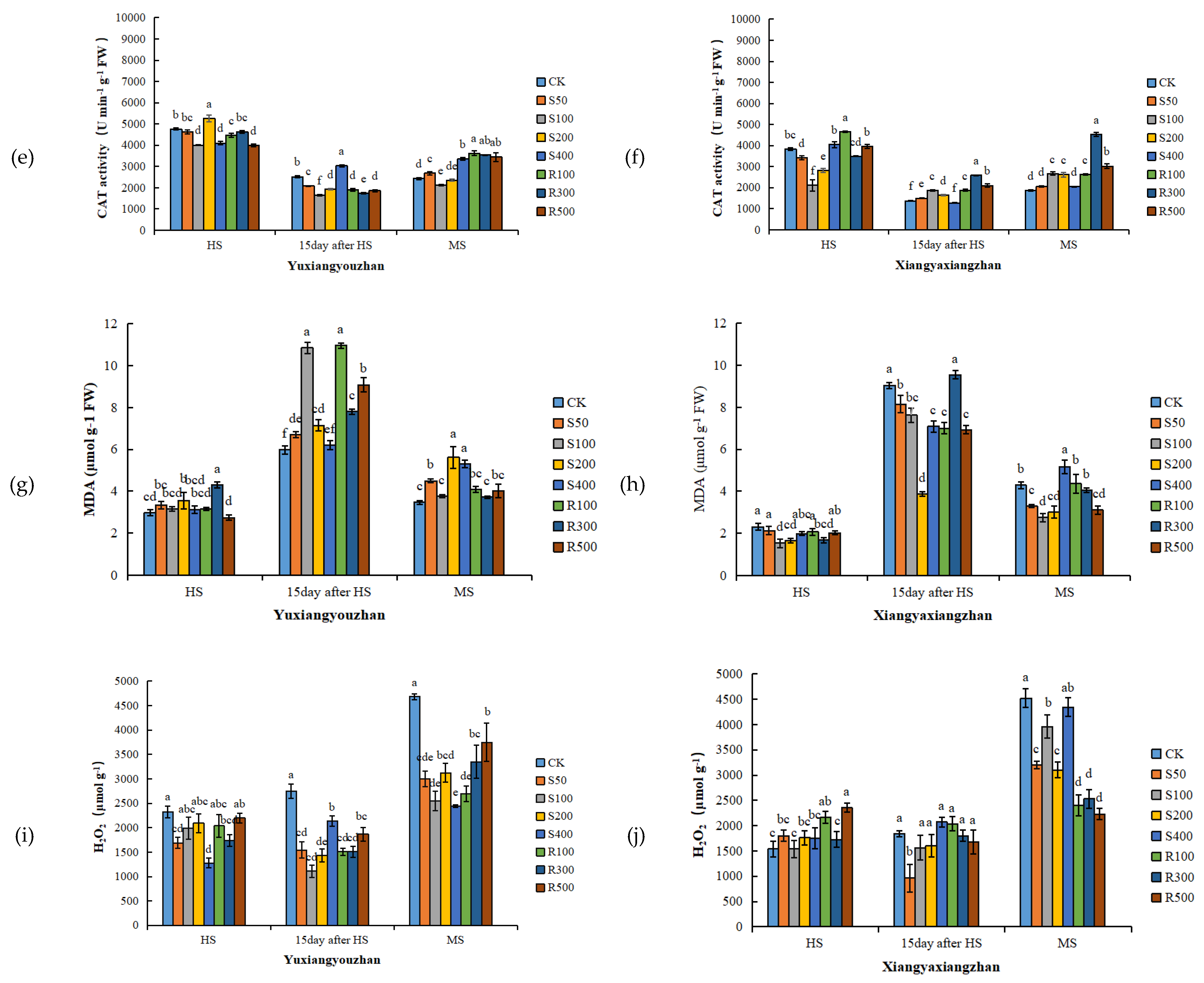
3.4. Antioxidant Enzyme Activities and MDA and H2O2 Contents in Leaves
3.5. Rhizosphere Microbial Community Structure
3.6. Investigation of Primary Metabolites
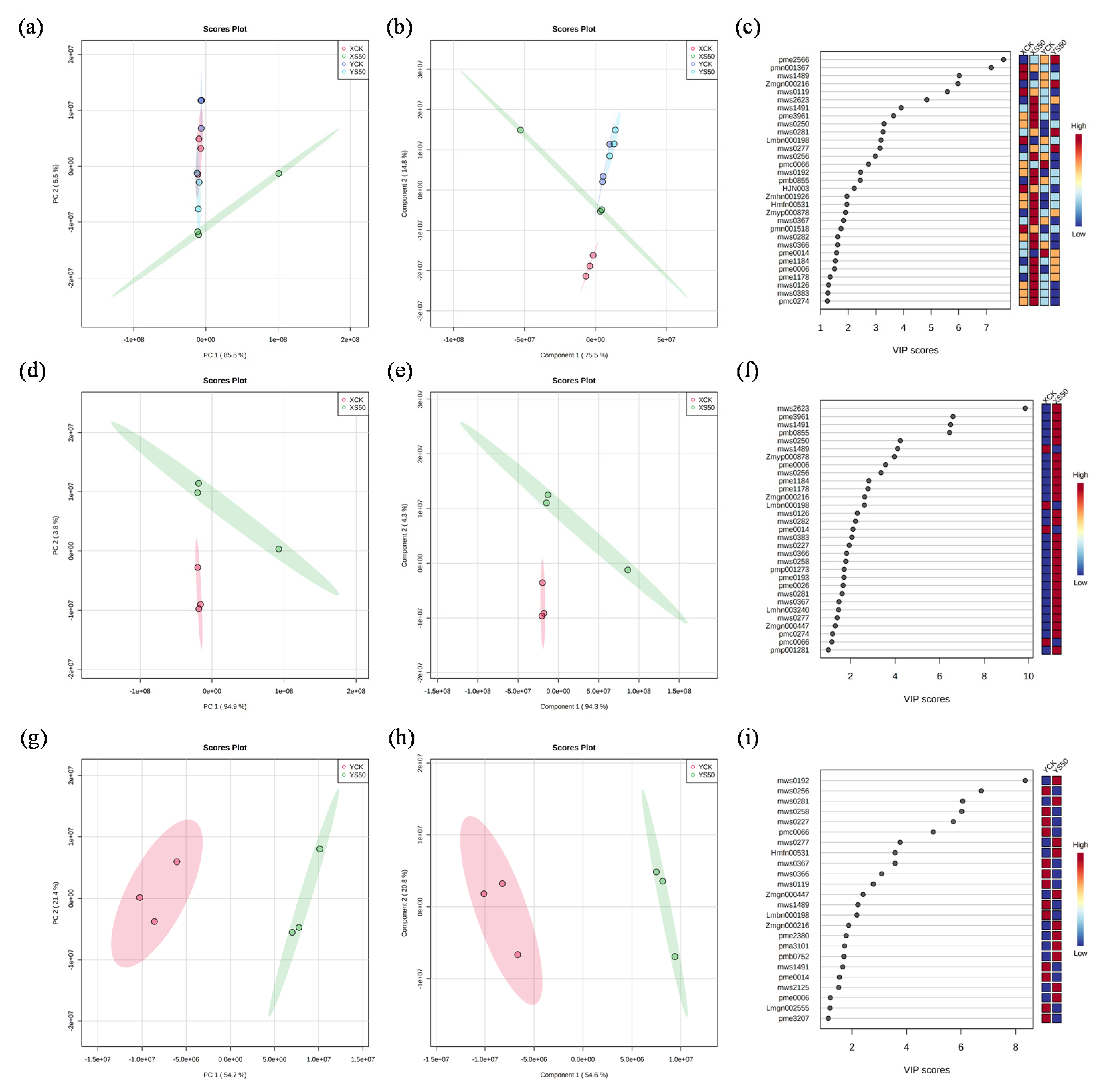
3.6.1. Metabolite Profiling
3.6.2. Differential Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, H.; He, L.; Du, B.; Pan, S.; Mo, Z.; Duan, M.; Tian, H.; Tang, X. Biofortification with chelating selenium in fragrant rice: Effects on photosynthetic rates, aroma, grain quality and yield formation. Field Crop. Res. 2020, 255, 107909. [Google Scholar] [CrossRef]
- Huang, G.; Ding, C.; Guo, F.; Zhang, T.; Wang, X. The optimum Se application time for reducing Cd uptake by rice (Oryza sativa L.) and its mechanism. Plant Soil 2018, 431, 231–243. [Google Scholar] [CrossRef]
- Su, N.; Wu, Q.; Chen, H.; Huang, Y.; Zhu, Z.; Chen, Y.; Cui, J. Hydrogen gas alleviates toxic effects of cadmium in Brassica campestris seedlings through up-regulation of the antioxidant capacities: Possible involvement of nitric oxide. Environ. Pollut. 2019, 251, 45–55. [Google Scholar] [CrossRef]
- Ashraf, U.; Mahmood, M.H.-U.; Hussain, S.; Abbas, F.; Anjum, S.A.; Tang, X. Lead (Pb) distribution and accumulation in different plant parts and its associations with grain Pb contents in fragrant rice. Chemosphere 2020, 248, 126003. [Google Scholar] [CrossRef]
- Sharafi, K.; Nodehi, R.N.; Mahvi, A.H.; Pirsaheb, M.; Nazmara, S.; Mahmoudi, B.; Yunesian, M. Bioaccessibility analysis of toxic metals in consumed rice through an in vitro human digestion model—Comparison of calculated human health risk from raw, cooked and digested rice. Food Chem. 2019, 299, 125126. [Google Scholar] [CrossRef]
- Gong, Z.Z.; Xiong, L.M.; Shi, H.Z.; Yang, S.H.; Herrera-Estrella, L.R.; Xu, G.H.; Chao, D.Y.; Li, J.R.; Wang, P.Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China-Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Xia, Q.; Lamb, D.; Peng, C.; Ng, J.C. Interaction effects of As, Cd and Pb on their respective bioaccessibility with time in co-contaminated soils assessed by the Unified BARGE Method. Environ. Sci. Pollut. Res. 2017, 24, 5585–5594. [Google Scholar] [CrossRef]
- He, H.H.; Wang, X.; Wu, M.M.; Guo, L.; Fan, C.B.; Peng, Q. Cadmium and lead affect the status of mineral nutrients in alfalfa grown on a calcareous soil. Soil Sci. Plant Nutr. 2020, 66, 506–514. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, M.; Chen, W. Joint Toxicity Assessment of Siduron and Heavy Metals on Cucumber and Wheat Seed Germination and Seedling Growth. Asian J. Ecotoxicol. 2017, 12, 544–555. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Pandey, P.; Rajpoot, R.; Rani, A.; Dubey, R.S. Cadmium and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings. Protoplasma 2014, 251, 1047–1065. [Google Scholar] [CrossRef]
- Wang, C.; Rong, H.; Zhang, X.; Shi, W.; Hong, X.; Liu, W.; Cao, T.; Yu, X.; Yu, Q. Effects and mechanisms of foliar application of silicon and selenium composite sols on diminishing cadmium and lead translocation and affiliated physiological and biochemical responses in hybrid rice (Oryza sativa L.) exposed to cadmium and lead. Chemosphere 2020, 251, 126347. [Google Scholar] [CrossRef]
- Dai, C.; Cui, W.; Pan, J.; Xie, Y.; Wang, J.; Shen, W. Proteomic analysis provides insights into the molecular bases of hydrogen gas-induced cadmium resistance in Medicago sativa. J. Proteom. 2017, 152, 109–120. [Google Scholar] [CrossRef]
- Huang, S.; Rao, G.; Ashraf, U.; He, L.; Zhang, Z.; Zhang, H.; Mo, Z.; Pan, S.; Tang, X. Application of inorganic passivators reduced Cd contents in brown rice in oilseed rape-rice rotation under Cd contaminated soil. Chemosphere 2020, 259, 127404. [Google Scholar] [CrossRef]
- Fan, D.; Sun, J.; Liu, C.; Wang, S.; Han, J.; Agathokleous, E.; Zhu, Y. Measurement and modeling of hormesis in soil bacteria and fungi under single and combined treatments of Cd and Pb. Sci. Total Environ. 2021, 783, 147494. [Google Scholar] [CrossRef]
- Hussain, B.; Lin, Q.; Hamid, Y.; Sanaullah, M.; Di, L.; Hashmi, M.L.U.R.; Khan, M.B.; He, Z.; Yang, X. Foliage application of selenium and silicon nanoparticles alleviates Cd and Pb toxicity in rice (Oryza sativa L.). Sci. Total Environ. 2020, 712, 136497. [Google Scholar] [CrossRef]
- Shida, C.; Castrucci, A.M.L.; Lamy-Freund, M.T. High melatonin solubility in aqueous medium. J. Pineal Res. 1994, 16, 198–201. [Google Scholar] [CrossRef]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar]
- Wang, P.; Sun, X.; Chang, C.; Feng, F.; Liang, D.; Cheng, L.; Ma, F. Delay in leaf senescence of Malus hupehensis by long-term melatonin application is associated with its regulation of metabolic status and protein degradation. J. Pineal Res. 2013, 55, 424–434. [Google Scholar] [CrossRef]
- Byeon, Y.; Back, K. An increase in melatonin in transgenic rice causes pleiotropic phenotypes, including enhanced seedling growth, delayed flowering, and low grain yield. J. Pineal Res. 2014, 56, 408–414. [Google Scholar] [CrossRef]
- Tal, O.; Haim, A.; Harel, O.; Gerchman, Y. Melatonin as an antioxidant and its semi-lunar rhythm in green macroalga Ulva sp. J. Exp. Bot. 2011, 62, 1903–1910. [Google Scholar] [CrossRef]
- Okazaki, M.; Higuchi, K.; Hanawa, Y.; Shiraiwa, Y.; Ezura, H. Cloning and characterization of a Chlamydomonas reinhardtii cDNA arylalkylamine N-acetyltransferase and its use in the genetic engineering of melatonin content in the Micro-Tom tomato. J. Pineal Res. 2009, 46, 373–382. [Google Scholar] [CrossRef]
- Hodžić, E.; Galijašević, S.; Balaban, M.; Rekanović, S.; Makić, H.; Kukavica, B.; Mihajlović, D. The protective role of melatonin under heavy metal-induced stress in Melissa Officinalis L. Turk. J. Chem. 2021, 45, 737–748. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Elkelish, A.; Dafea, M.; Khan, M.; Arnao, M.B.; Abdelhamid, M.T.; Abu El-Ezz, A.; Almoneafy, A.; Mahmoud, A.; Awad, M.; et al. Role of Melatonin in Plant Tolerance to Soil Stressors: Salinity, pH and Heavy Metals. Molecules 2020, 25, 5359. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Ali, S.; Refay, Y.; Rizwan, M.; Alhammad, B.A.; El-Hendawy, S.E. Chromium resistant microbes and melatonin reduced Cr uptake and toxicity, improved physio-biochemical traits and yield of wheat in contaminated soil. Chemosphere 2020, 250, 126239. [Google Scholar] [CrossRef]
- Li, Y.; Liang, L.; Huang, S.; Li, W.; Ashraf, U.; Ma, L.; Mo, Z. Exogenous Melatonin and Catechol Application Modulate Physio-Biochemical Attributes and Early Growth of Fragrant Rice Under Cd Toxicity. J. Soil Sci. Plant Nutr. 2021, 21, 2285–2296. [Google Scholar] [CrossRef]
- Mo, Z.; Li, W.; Pan, S.; Fitzgerald, T.L.; Xiao, F.; Tang, Y.; Wang, Y.; Duan, M.; Tian, H.; Tang, X. Shading during the grain filling period increases 2-acetyl-1-pyrroline content in fragrant rice. Rice 2015, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Huang, Z.; Li, Y.; Xie, W.; Li, W.; Tang, X.; Ashraf, U.; Kong, L.; Wu, L.; Wang, S.; et al. Selenium-silicon (Se-Si) induced modulations in physio-biochemical responses, grain yield, quality, aroma formation and lodging in fragrant rice. Ecotoxicol. Environ. Saf. 2020, 196, 110525. [Google Scholar] [CrossRef]
- Xie, W.J.; Ashraf, U.; Zhong, D.T.; Lin, R.B.; Xian, P.Q.; Zhao, T.; Feng, H.Y.; Wang, S.L.; Duan, M.Y.; Tang, X.R.; et al. Application of gamma-aminobutyric acid (GABA) and nitrogen regulates aroma biochemistry in fragrant rice. Food Sci. Nutr. 2019, 7, 3784–3796. [Google Scholar] [CrossRef]
- Xie, W.J.; Kong, L.L.; Ma, L.; Ashraf, U.; Pan, S.G.; Duan, M.Y.; Tian, H.; Wu, L.M.; Tang, X.R.; Mo, Z.W. Enhancement of 2-acetyl-1-pyrroline (2AP) concentration, total yield, and quality in fragrant rice through exogenous gamma-aminobutyric acid (GABA) application. J. Cereal Sci. 2020, 91, 102900. [Google Scholar] [CrossRef]
- Xie, W.J.; Li, Y.H.; Li, Y.Z.; Ma, L.; Ashraf, U.; Tang, X.R.; Pan, S.G.; Tian, H.; Mo, Z.W. Application of gamma-aminobutyric acid under low light conditions: Effects on yield, aroma, element status, and physiological attributes of fragrant rice. Ecotoxicol. Environ. Saf. 2021, 213, 111941. [Google Scholar] [CrossRef]
- Kanu, A.S.; Ashraf, U.; Mo, Z.; Fuseini, I.; Mansaray, L.R.; Duan, M.; Pan, S.; Tang, X. Cadmium Uptake and Distribution in Fragrant Rice Genotypes and Related Consequences on Yield and Grain Quality Traits. J. Chem. 2017, 2017, 1405878. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Jiao, Y.; Chen, C.; Shireen, F.; Zheng, Z.; Imtiaz, M.; Bie, Z.; Huang, Y. Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression. J. Plant Physiol. 2018, 220, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Banerjee, A.; Roychoudhury, A. Arsenic Toxicity is Counteracted by Exogenous Application of Melatonin to Different Extents in Arsenic-susceptible and Arsenic-tolerant Rice Cultivars. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Liu, J.-N.; Zhou, Q.-X.; Sun, T.; Ma, L.Q.; Wang, S. Growth responses of three ornamental plants to Cd and Cd–Pb stress and their metal accumulation characteristics. J. Hazard. Mater. 2008, 151, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, H.; Wang, J.; Wang, Y.; Pan, S.; Tian, H.; Duan, M.; Wang, S.; Tang, X.; Mo, Z. Responses of plant growth, physiological, gas exchange parameters of super and non-super rice to rhizosphere temperature at the tillering stage. Sci. Rep. 2019, 9, 10618. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ni, T.; Li, J.; Lu, Q.; Fang, Z.Y.; Huang, Q.W.; Zhang, R.F.; Li, R.; Shen, B.; Shen, Q.R. Effects of organic-inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice-wheat cropping system. Appl. Soil Ecol. 2016, 99, 1–12. [Google Scholar] [CrossRef]
- Reth, S.; Reichstein, M.; Falge, E. The effect of soil water content, soil temperature, soil pH-value and the root mass on soil CO2 efflux—A modified model. Plant Soil. 2005, 268, 21–33. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [Green Version]
- Demecsová, L.; Tamás, L. Reactive oxygen species, auxin and nitric oxide in metal-stressed roots: Toxicity or defence. BioMetals 2019, 32, 717–744. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Guo, X.Q.; Mo, G.L.; Qin, X.F. Migration and transformation characteristics of heavy metal cadmium in soil-rice. Guizhou Agric. Sci. 2021, 49, 143–149. [Google Scholar]
- Posmyk, M.M.; Kuran, H.; Marciniak, K.; Janas, K.M. Presowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations. J. Pineal Res. 2008, 45, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Chen, K.M.; Gong, H.J.; Wang, S.M. Glutathione metabolism and environmental stresses in plants. Xibei Zhiwu Xuebao 2004, 24, 1119–1130. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Hasan, M.K.; Ahammed, G.J.; Yin, L.L.; Shi, K.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q.; Zhou, J. Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthesis, vacuolar sequestration, and antioxidant potential in Solanum lycopersicum L. Front. Plant Sci. 2015, 6, 601. [Google Scholar] [CrossRef] [PubMed]
- Somashekaraiah, B.V.; Padmaja, K.; Prasad, A.R.K. Phytotoxicity of cadmium ions on germinating seedlings of mung bean (Phaseolus vulgaris): Involvement of lipid peroxides in chlorophyll degradation. Physiol. Plant. 1992, 85, 85–89. [Google Scholar] [CrossRef]
- Sandalio, L.; Dalurzo, H.; Gómez, M.; Romero-Puertas, M.; del Río, L. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Yang, Y.-J.; Cheng, L.-M.; Liu, Z.-H. Rapid effect of cadmium on lignin biosynthesis in soybean roots. Plant Sci. 2007, 172, 632–639. [Google Scholar] [CrossRef]
- Corticeiro, S.C.; Lima, A.I.G.; Figueira, E.M.D.A.P. The importance of glutathione in oxidative status of Rhizobium leguminosarum biovar viciae under Cd exposure. Enzym. Microb. Technol. 2006, 40, 132–137. [Google Scholar] [CrossRef]
- Zhang, H.-J.; Zhang, N.; Yang, R.-C.; Wang, L.; Sun, Q.-Q.; Li, D.-B.; Cao, Y.-Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Plummer, B.F.; Hardies, L.J.; Weintraub, S.T.; Shepherd, A.M. A novel melatonin metabolite, cyclic 3-hydroxymelatonin: A biomarker of in vivo hydroxyl radical generation. Biochem. Biophys. Res. Commun. 1998, 253, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.C.; Reiter, R.J.; Plummer, B.F.; Limson, J.; Weintraub, S.T.; Qi, W. Melatonin directly scavenges hydrogen peroxide: A potentially new metabolic pathway of melatonin biotransformation. Free Radic. Biol. Med. 2000, 29, 1177–1185. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Wei, Z.; Liang, D.; Liu, C.; Yin, L.; Jia, D.; Fu, M.; Ma, F. The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef] [PubMed]
- García, J.J.; López-Pingarrón, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.-X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, A.; Kamrava, S.K.; Joghataei, M.T.; Darabi, R.; Shakeri-Zadeh, A.; Shahriari, M.; Reiter, R.J.; Ghaznavi, H.; Mehrzadi, S. Apoptosis signaling pathways in osteoarthritis and possible protective role of melatonin. J. Pineal Res. 2016, 61, 411–425. [Google Scholar] [CrossRef]
- Wang, P.; Yin, L.; Liang, D.; Li, C.; Ma, F.; Yue, Z. Delayed senescence of apple leaves by exogenous melatonin treatment: Toward regulating the ascorbate-glutathione cycle. J. Pineal Res. 2012, 53, 11–20. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin promotes adventitious- and lateral root regeneration in etiolated hypocotyls of Lupinus albus L. J. Pineal Res. 2007, 42, 147–152. [Google Scholar] [CrossRef]
- Navarro-León, E.; Albacete, A.; Romero, L.; Blasco, B.; Ruiz, J.M. Response of carboxylate metabolism to zinc deficiency in Lactuca sativa and Brassica oleracea plants. J. Plant Nutr. Soil Sci. 2016, 179, 758–764. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J.; Wang, G.; Fan, X.; Sun, X.; Qin, H.; Xu, N.; Zhong, M.; Qiao, Z.; Tang, Y.; et al. Proline responding1Plays a Critical Role in Regulating General Protein Synthesis and the Cell Cycle in Maize. Plant Cell 2014, 26, 2582–2600. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liang, L.; Li, W.; Ashraf, U.; Ma, L.; Tang, X.; Pan, S.; Tian, H.; Mo, Z. ZnO nanoparticle-based seed priming modulates early growth and enhances physio-biochemical and metabolic profiles of fragrant rice against cadmium toxicity. J. Nanobiotechnol. 2021, 19, 75. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Cd Content at the HS (mg kg−1) | Cd Content at MS (mg kg−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Roots | Shoots | Leaves | Roots | Shoots | Leaves | Grains | ||
| Yuxiangyouzhan | CK | 0.6959 ± 0.0097 a | 0.3190 ± 0.0029 b | 0.2452 ± 0.0056 b | 0.6351 ± 0.0088 c | 0.4449 ± 0.0117 e | 0.3781 ± 0.0412 cd | 0.1101 ± 0.0019 f |
| S50 | 0.6391 ± 0.0067 b | 0.2964 ± 0.0116 c | 0.2174 ± 0.0041 bc | 0.5544 ± 0.0227 c | 0.5452 ± 0.0099 b | 0.3882 ± 0.0026 cd | 0.1404 ± 0.0047 e | |
| S100 | 0.6337 ± 0.0023 b | 0.2946 ± 0.0058 c | 0.1542 ± 0.0199 d | 0.8774 ± 0.0384 b | 0.4299 ± 0.0037 e | 0.350 ± 0.0039 cd | 0.1444 ± 0.0012 de | |
| S200 | 0.6520 ± 0.0005 b | 0.3284 ± 0.0006 ab | 0.1920 ± 0.0041 c | 0.6587 ± 0.0074 c | 0.5146 ± 0.0038 cd | 0.4646 ± 0.0141 a | 0.1539 ± 0.0034 c | |
| S400 | 0.6352 ± 0.0123 b | 0.2964 ± 0.0015 c | 0.2942 ± 0.0174 a | 1.0646 ± 0.107 a | 0.5317 ± 0.0097 bc | 0.3724 ± 0.0146 cd | 0.1512 ± 0.0008 cd | |
| R100 | 0.6532 ± 0.0085 b | 0.345 ± 0.0122 b | 0.3203 ± 0.0026 a | 0.9645 ± 0.0402 ab | 0.6179 ± 0.0147 a | 0.4501 ± 0.0042 ab | 0.1754 ± 0.0015 b | |
| R300 | 0.6398 ± 0.0039 b | 0.3264 ± 0.0051 ab | 0.2473 ± 0.0109 b | 0.9879 ± 0.019 ab | 0.5421 ± 0.0017 bc | 0.3395 ± 0.0052 d | 0.1687 ± 0.0006 b | |
| R500 | 0.6541 ± 0.0032 b | 0.3463 ± 0.005 b | 0.3234 ± 0.0038 a | 1.0074 ± 0.0199 ab | 0.4944 ± 0.0105 d | 0.3993 ± 0.0055 bc | 0.1844 ± 0.0017 a | |
| Xiangyaxiangzhan | CK | 0.7083 ± 0.0054 a | 0.3518 ± 0.0063 d | 0.3285 ± 0.0025 bc | 1.0652 ± 0.0135 b | 0.5453 ± 0.0114 a | 0.4977 ± 0.005 a | 0.1753 ± 0.0012 e |
| S50 | 0.6390 ± 0.0018 d | 0.3251 ± 0.0002 g | 0.2756 ± 0.0062 e | 1.0205 ± 0.0105 c | 0.5003 ± 0.0047 b | 0.4131 ± 0.002 d | 0.1919 ± 0.0002 b | |
| S100 | 0.6308 ± 0.0002 d | 0.3470 ± 0.0013 de | 0.3202 ± 0.0015 bcd | 1.1254 ± 0.0073 a | 0.4860 ± 0.0076 b | 0.4583 ± 0.0167 bc | 0.1749 ± 0.0001 e | |
| S200 | 0.5923 ± 0.0014 e | 0.3631 ± 0.0007 c | 0.2985 ± 0.0029 de | 0.9774 ± 0.0073 d | 0.3441 ± 0.0025 c | 0.3596 ± 0.0065 e | 0.1431 ± 0.0017 f | |
| S400 | 0.6608 ± 0.0012 c | 0.3957 ± 0.004 b | 0.3596 ± 0.0029 a | 1.0145 ± 0.0024 c | 0.4987 ± 0.0013 b | 0.4249 ± 0.0037 cd | 0.1858 ± 0.0011 c | |
| R100 | 0.6383 ± 0.0056 d | 0.3329 ± 0.0021 fg | 0.3027 ± 0.0007 cde | 1.0829 ± 0.0167 b | 0.4973 ± 0.0045 b | 0.4029 ± 0.0049 bc | 0.1813 ± 0.0005 d | |
| R300 | 0.6782 ± 0.0092 b | 0.3377 ± 0.0026 ef | 0.2815 ± 0.0265 e | 1.0676 ± 0.005 b | 0.536 ± 0.0169 a | 0.4386 ± 0.0063 bc | 0.1758 ± 0.0013 e | |
| R500 | 0.5869 ± 0.0027 e | 0.4063 ± 0.0059 a | 0.3335 ± 0.0072 ab | 1.0186 ± 0.0149 c | 0.4874 ± 0.0074 b | 0.4189 ± 0.0078 cd | 0.2074 ± 0.0002 c | |
| Cultivars | Treatments | TF of HS(Cd) | TF of MS(Cd) | TF of HS(Pb) | TF of MS(Pb) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R to S | S to L | R to S | S to L | L to G | R to S | S to L | R to S | S to L | L to G | ||
| Yuxiangyouzhan | CK | 0.4583 | 0.7687 | 0.7005 | 0.8500 | 0.2912 | 0.1631 | 0.7646 | 0.1177 | 0.9260 | 0.0487 |
| S50 | 0.4637 | 0.7335 | 0.9835 | 0.7120 | 0.3617 | 0.1849 | 0.6960 | 0.1986 | 0.6255 | 0.0122 | |
| S100 | 0.4648 | 0.5236 | 0.4899 | 0.8142 | 0.4127 | 0.1879 | 0.5988 | 0.1166 | 0.7683 | 0.0411 | |
| S200 | 0.5036 | 0.5846 | 0.7812 | 0.9029 | 0.3312 | 0.1456 | 0.5175 | 0.2066 | 0.5529 | 0.0299 | |
| S400 | 0.4666 | 0.9927 | 0.4994 | 0.7004 | 0.4060 | 0.1945 | 0.6210 | 0.1505 | 0.6794 | 0.0473 | |
| R100 | 0.5282 | 0.9283 | 0.6406 | 0.7284 | 0.3898 | 0.1478 | 0.7407 | 0.1705 | 0.9445 | 0.0254 | |
| R300 | 0.5101 | 0.7578 | 0.5488 | 0.6262 | 0.4969 | 0.1928 | 0.4544 | 0.1167 | 0.8652 | 0.0539 | |
| R500 | 0.5294 | 0.9339 | 0.4908 | 0.8076 | 0.4617 | 0.1880 | 0.9723 | 0.1420 | 0.4224 | 0.1423 | |
| Xiangyaxiangzhan | CK | 0.4967 | 0.9337 | 0.5119 | 0.9127 | 0.3523 | 0.1523 | 0.7218 | 0.1426 | 0.7639 | 0.0844 |
| S50 | 0.5088 | 0.8478 | 0.4903 | 0.8256 | 0.4646 | 0.1469 | 0.6150 | 0.1542 | 0.6247 | 0.0729 | |
| S100 | 0.5501 | 0.9228 | 0.4319 | 0.9429 | 0.3817 | 0.1660 | 0.5222 | 0.1978 | 0.4477 | 0.1010 | |
| S200 | 0.6130 | 0.8220 | 0.3521 | 1.0450 | 0.3980 | 0.1630 | 0.6848 | 0.1514 | 0.6047 | 0.1251 | |
| S400 | 0.5988 | 0.9088 | 0.4916 | 0.8520 | 0.4373 | 0.1459 | 0.8065 | 0.2444 | 0.7118 | 0.0517 | |
| R100 | 0.5215 | 0.9095 | 0.4593 | 0.8102 | 0.4499 | 0.1293 | 0.7340 | 0.1474 | 0.6758 | 0.0644 | |
| R300 | 0.4980 | 0.8334 | 0.5021 | 0.8183 | 0.4007 | 0.1123 | 0.7577 | 0.1613 | 0.5962 | 0.0613 | |
| R500 | 0.6922 | 0.8209 | 0.4785 | 0.8595 | 0.4950 | 0.2372 | 0.8762 | 0.1646 | 0.7682 | 0.0972 | |
| Treatments | Pb Content at the HS (mg kg−1) | Pb Content at MS (mg kg−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Roots | Shoots | Leaves | Roots | Shoots | Leaves | Grains | ||
| Yuxiangyouzhan | CK | 2.2953 ± 0.0317 b | 0.3744 ± 0.0031 ab | 0.2863 ± 0.0026 b | 2.3747 ± 0.0126 ef | 0.2795 ± 0.0086 c | 0.2588 ± 0.0179 d | 0.0126 ± 0.0002 cd |
| S50 | 2.2107 ± 0.0046 c | 0.4087 ± 0.0103 ab | 0.2844 ± 0.0021 b | 2.2534 ± 0.0156 f | 0.4474 ± 0.0160 b | 0.2799 ± 0.0094 cd | 0.0034 ± 0.0004 f | |
| S100 | 2.2429 ± 0.0091 c | 0.4215 ± 0.0133 ab | 0.2524 ± 0.0059 c | 2.6869 ± 0.0284 d | 0.3133 ± 0.0081 c | 0.2407 ± 0.0058 de | 0.0099 ± 0.0008 de | |
| S200 | 2.3688 ± 0.0052 a | 0.3448 ± 0.0278 b | 0.1784 ± 0.0017 e | 2.6092 ± 0.027 de | 0.5392 ± 0.0284 a | 0.2981 ± 0.0186 cd | 0.0089 ± 0.0006 e | |
| S400 | 2.3190 ± 0.0067 b | 0.4511 ± 0.0657 a | 0.2801 ± 0.0006 b | 3.1400 ± 0.1116 c | 0.4726 ± 0.0235 b | 0.3211 ± 0.0049 bc | 0.0152 ± 0.0015 c | |
| R100 | 2.3833 ± 0.0167 a | 0.3522 ± 0.0051 b | 0.2609 ± 0.0095 c | 3.4200 ± 0.0556 ab | 0.5832 ± 0.0374 a | 0.5508 ± 0.0305 a | 0.0140 ± 0.0014 c | |
| R300 | 2.3221 ± 0.0059 b | 0.4476 ± 0.0397 a | 0.2034 ± 0.0035 d | 3.5829 ± 0.0932 a | 0.4182 ± 0.0103 b | 0.3618 ± 0.0179 b | 0.0195 ± 0.0018 b | |
| R500 | 2.3900 ± 0.0080 a | 0.4493 ± 0.0018 a | 0.4369 ± 0.0051 a | 3.1836 ± 0.166 bc | 0.4521 ± 0.0167 b | 0.1909 ± 0.0092 e | 0.0272 ± 0.0011 a | |
| Xiangyaxiangzhan | CK | 2.3847 ± 0.0035 bc | 0.3632 ± 0.0169 bc | 0.2621 ± 0.004 bc | 2.7164 ± 0.0263 a | 0.3873 ± 0.0378 b | 0.2958 ± 0.0125 b | 0.0250 ± 0.0012 b |
| S50 | 2.2789 ± 0.0065 e | 0.3348 ± 0.0211 bc | 0.2059 ± 0.0197 d | 2.0039 ± 0.1730 c | 0.3090 ± 0.0127 c | 0.1930 ± 0.0018 d | 0.0141 ± 0.0007 e | |
| S100 | 2.3133 ± 0.0020 de | 0.3839 ± 0.0017 b | 0.2005 ± 0.0071 d | 2.3002 ± 0.0713 b | 0.4588 ± 0.0141 a | 0.2054 ± 0.0105 d | 0.0207 ± 0.0005 bcd | |
| S200 | 2.2784 ± 0.0192 e | 0.3713 ± 0.0096 b | 0.2543 ± 0.0058 c | 1.9672 ± 0.0902 c | 0.2978 ± 0.0224 c | 0.1800 ± 0.0037 d | 0.0225 ± 0.0004 bc | |
| S400 | 2.4662 ± 0.0129 a | 0.3597 ± 0.0434 bc | 0.2901 ± 0.0056 b | 1.9096 ± 0.0877 c | 0.4666 ± 0.0066 a | 0.3321 ± 0.0075 a | 0.0172 ± 0.0013 cde | |
| R100 | 2.3322 ± 0.0149 cd | 0.3016 ± 0.0208 cd | 0.2214 ± 0.0048 d | 2.6686 ± 0.0395 a | 0.3933 ± 0.0135 b | 0.2658 ± 0.0033 bc | 0.0171 ± 0.0005 cde | |
| R300 | 2.4035 ± 0.0423 b | 0.2698 ± 0.0077 d | 0.2044 ± 0.0027 d | 2.7063 ± 0.1484 a | 0.4365 ± 0.0046 ab | 0.2602 ± 0.0043 cd | 0.0160 ± 0.0017 de | |
| R500 | 2.3678 ± 0.0034 bc | 0.5616 ± 0.0191 a | 0.4921 ± 0.0208 a | 2.8137 ± 0.0294 a | 0.4632 ± 0.004 a | 0.3559 ± 0.0265 a | 0.0346 ± 0.0057 a | |
| Cultivars | Treatment | Tiller Number Per Hill | Grain Number Per Panicle | Filled Grain Percentage (%) | 1000-Grain Weight (g) | Grain Yield (g/Pot) |
|---|---|---|---|---|---|---|
| Yuxiangyouzhan | CK | 6.42 ± 0.21 b | 117.05 ± 6.18 b | 65.01 ± 4.64 b | 19.28 ± 0.2 b | 45.34 ± 0.99 c |
| S50 | 7.53 ± 0.35 a | 122.81 ± 16.13 ab | 66.14 ± 3.75 b | 19.75 ± 0.23 ab | 52.32 ± 2.81 bc | |
| S100 | 6.89 ± 0.53 ab | 133.07 ± 3.61 ab | 71.37 ± 4.81 ab | 19.64 ± 0.25 ab | 61.72 ± 2.26 ab | |
| S200 | 6.83 ± 0.17 ab | 126.98 ± 2.02 ab | 75.73 ± 3.55 ab | 20.13 ± 0.32 a | 66.16 ± 3.68 a | |
| S400 | 6.46 ± 0.18 b | 120.24 ± 6.29 b | 68.35 ± 2.43 ab | 19.70 ± 0.33 ab | 49.92 ± 3.31 c | |
| R100 | 6.67 ± 0.14 ab | 134.52 ± 8.84 ab | 79.83 ± 0.59 a | 19.25 ± 0.25 b | 62.68 ± 5.13 ab | |
| R300 | 7.50 ± 0.40 a | 144.34 ± 8.41 a | 68.94 ± 5.20 ab | 18.97 ± 0.20 b | 64.95 ± 3.09 a | |
| R500 | 6.78 ± 0.30 ab | 128.03 ± 8.33 ab | 68.87 ± 4.07 ab | 19.32 ± 0.25 b | 53.86 ± 5.47 bc | |
| Xiangyaxiangzhan | CK | 7.48 ± 0.29 b | 75.58 ± 10.14 b | 80.78 ± 4.56 a | 19.30 ± 0.12 a | 44.84 ± 1.41 b |
| S50 | 8.00 ± 0.43 ab | 83.02 ± 6.32 ab | 80.92 ± 1.29 a | 19.05 ± 0.25 ab | 47.07 ± 1.96 b | |
| S100 | 7.50 ± 0.18 b | 81.59 ± 2.60 ab | 82.76 ± 0.64 a | 19.37 ± 0.35 a | 47.17 ± 1.17 b | |
| S200 | 8.69 ± 0.47 a | 81.04 ± 7.60 ab | 82.36 ± 2.27 a | 18.91 ± 0.49 ab | 54.18 ± 2.26 a | |
| S400 | 7.58 ± 0.75 ab | 97.10 ± 6.73 a | 76.60 ± 2.47 a | 18.96 ± 0.47 ab | 54.76 ± 1.90 a | |
| R100 | 8.08 ± 0.55 ab | 89.12 ± 2.81 ab | 78.77 ± 2.40 a | 18.09 ± 0.26 b | 49.38 ± 0.80 ab | |
| R300 | 7.69 ± 0.18 ab | 96.13 ± 3.23 a | 76.83 ± 5.89 a | 18.46 ± 0.19 ab | 49.55 ± 1.82 ab | |
| R500 | 7.42 ± 0.25 b | 81.15 ± 7.56 ab | 76.27 ± 2.42 a | 18.75 ± 0.25 ab | 43.97 ± 4.24 b |
| Treatment | SOM (g kg−1) | pH | N (g kg−1) | P (g kg−1) | K (g kg−1) | Cd (mg kg−1) | Pb (mg kg−1) |
|---|---|---|---|---|---|---|---|
| XCK | 8.32 ± 0.08 | 6.42 ± 0.02 | 0.77 ± 0.11 | 1.52 ± 0.11 | 6.47 ± 0.29 | 1.15 ± 0.01 | 4.45 ± 0.06 |
| XR300 | 8.64 ± 0.13 | 6.25 ± 0.03 | 0.83 ± 0.01 | 0.95 ± 0.03 | 6.34 ± 0.11 | 1.18 ± 0.02 | 4.59 ± 0.08 |
| XS50 | 9.58 ± 0.16 | 6.18 ± 0.02 | 1.39 ± 0.05 | 1.64 ± 0.09 | 6.12 ± 0.28 | 1.18 ± 0.01 | 4.57 ± 0.04 |
| YCK | 9.82 ± 0.25 | 5.84 ± 0.03 | 1.12 ± 0.04 | 0.97 ± 0.12 | 6.81 ± 0.68 | 1.20 ± 0.01 | 3.51 ± 0.14 |
| YR300 | 8.97 ± 0.09 | 6.86 ± 0.02 | 0.68 ± 0.07 | 0.81 ± 0.02 | 5.57 ± 0.20 | 1.21 ± 0.02 | 4.49 ± 0.04 |
| YS50 | 9.96 ± 0.12 | 6.25 ± 0.03 | 1.47 ± 0.04 | 0.86 ± 0.04 | 6.35 ± 0.18 | 1.19 ± 0.01 | 3.97 ± 0.05 |
| Sample ID | OTU | ACE | Chao1 | Simpson | Shannon | Coverage |
|---|---|---|---|---|---|---|
| XCK | 1827.33 ± 3.93 ab | 1840.14 ± 4.16 a | 1842.1881 ± 5.4287 a | 0.0026 ± 0.0001 b | 6.6893 ± 0.0210 a | 0.9987 ± 0.0001 a |
| XR300 | 1822.67 ± 1.20 b | 1837.18 ± 2.64 a | 1839.7701 ± 4.3173 a | 0.0029 ± 0.0000 a | 6.6352 ± 0.0065 b | 0.9986 ± 0.0002 a |
| XS50 | 1836.33 ± 3.71 a | 1847.25 ± 3.07 a | 1851.9076 ± 4.0648 a | 0.0026 ± 0.0001 b | 6.7050 ± 0.0136 a | 0.9988 ± 0.0001 a |
| YCK | 1810.67 ± 7.26 b | 1830.41 ± 5.17 b | 1839.0321 ± 5.2287 a | 0.0030 ± 0.0001 a | 6.5805 ± 0.0227 b | 0.9984 ± 0.0002 a |
| YR300 | 1834.67 ± 3.18 a | 1847.36 ± 4.37 a | 1851.8427 ± 4.0325 a | 0.0026 ± 0.0001 b | 6.7054 ± 0.0187 a | 0.9987 ± 0.0001 a |
| YS50 | 1825.00 ± 4.16 ab | 1838.62 ± 2.97 ab | 1843.3003 ± 2.0112 a | 0.0027 ± 0.0001 b | 6.6682 ± 0.0056 a | 0.9986 ± 0.0002 a |
| Treatment | Acidobacteria | Actinobacteria | Chloroflexi | Gemmatimonadetes | Proteobacteria |
|---|---|---|---|---|---|
| XCK | 0.150 ± 0.004 a | 0.069 ± 0.001 b | 0.317 ± 0.014 a | 0.034 ± 0.001 a | 0.320 ± 0.011 a |
| XR300 | 0.149 ± 0.005 a | 0.082 ± 0.002 a | 0.344 ± 0.002 a | 0.035 ± 0.001 a | 0.276 ± 0.004 b |
| XS50 | 0.153 ± 0.002 a | 0.065 ± 0.004 b | 0.318 ± 0.010 a | 0.034 ± 0.003 a | 0.316 ± 0.004 a |
| YCK | 0.163 ± 0.005 a | 0.059 ± 0.007 a | 0.357 ± 0.009 a | 0.033 ± 0.002 a | 0.290 ± 0.011 b |
| YR300 | 0.134 ± 0.004 b | 0.063 ± 0.005 a | 0.323 ± 0.015 a | 0.028 ± 0.001 a | 0.325 ± 0.009 a |
| YS50 | 0.165 ± 0.005 a | 0.059 ± 0.005 a | 0.339 ± 0.005 a | 0.032 ± 0.001 a | 0.296 ± 0.002 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Huang, S.; Ma, L.; Kong, L.; Pan, S.; Tang, X.; Tian, H.; Duan, M.; Mo, Z. Effect of Exogenous Melatonin Application on the Grain Yield and Antioxidant Capacity in Aromatic Rice under Combined Lead–Cadmium Stress. Antioxidants 2022, 11, 776. https://doi.org/10.3390/antiox11040776
Jiang Y, Huang S, Ma L, Kong L, Pan S, Tang X, Tian H, Duan M, Mo Z. Effect of Exogenous Melatonin Application on the Grain Yield and Antioxidant Capacity in Aromatic Rice under Combined Lead–Cadmium Stress. Antioxidants. 2022; 11(4):776. https://doi.org/10.3390/antiox11040776
Chicago/Turabian StyleJiang, Ye, Suihua Huang, Lin Ma, Leilei Kong, Shenggang Pan, Xiangru Tang, Hua Tian, Meiyang Duan, and Zhaowen Mo. 2022. "Effect of Exogenous Melatonin Application on the Grain Yield and Antioxidant Capacity in Aromatic Rice under Combined Lead–Cadmium Stress" Antioxidants 11, no. 4: 776. https://doi.org/10.3390/antiox11040776
APA StyleJiang, Y., Huang, S., Ma, L., Kong, L., Pan, S., Tang, X., Tian, H., Duan, M., & Mo, Z. (2022). Effect of Exogenous Melatonin Application on the Grain Yield and Antioxidant Capacity in Aromatic Rice under Combined Lead–Cadmium Stress. Antioxidants, 11(4), 776. https://doi.org/10.3390/antiox11040776