
Supplementation with the Symbiotic Formulation Prodefen® Increases Neuronal Nitric Oxide Synthase and Decreases Oxidative Stress in Superior Mesenteric Artery from Spontaneously Hypertensive Rats


 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Blood Pressure Measurements
2.3. Animal Euthanasia and Sample Collection
2.4. Circulating Endotoxin Levels
2.5. Short-Chain Fatty Acids (SCFA) Analysis
2.6. Vascular Reactivity
2.7. Nitric Oxide Release
2.8. Detection of Superoxide Anions
2.9. Peroxynitrite Detection
2.10. Superoxide Dismutase Activity
2.11. PKA and PKC Activity Assays
2.12. Western Blot Analysis
2.13. Drugs and Antibodies Used
2.14. Data Analysis
3. Results
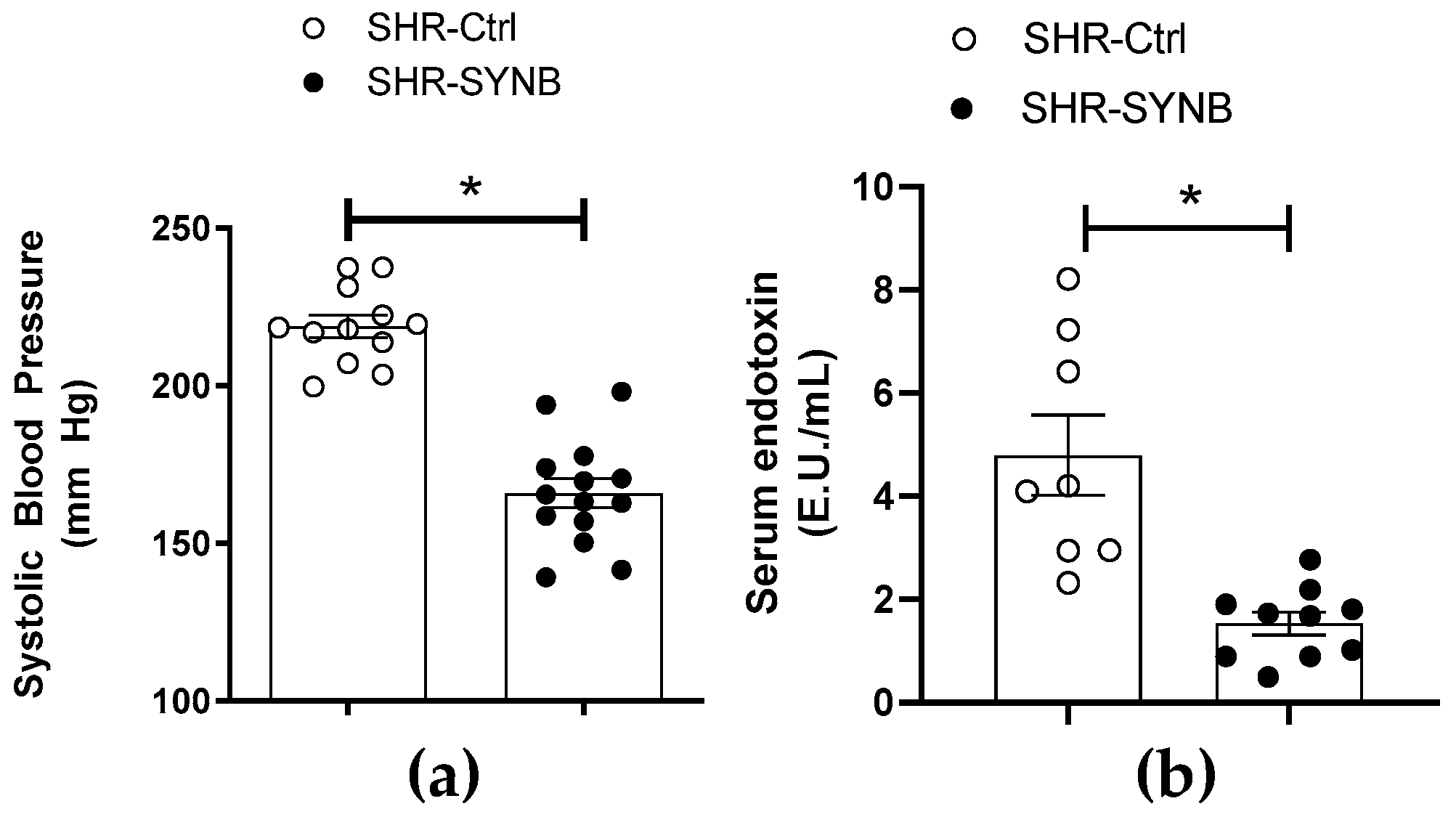
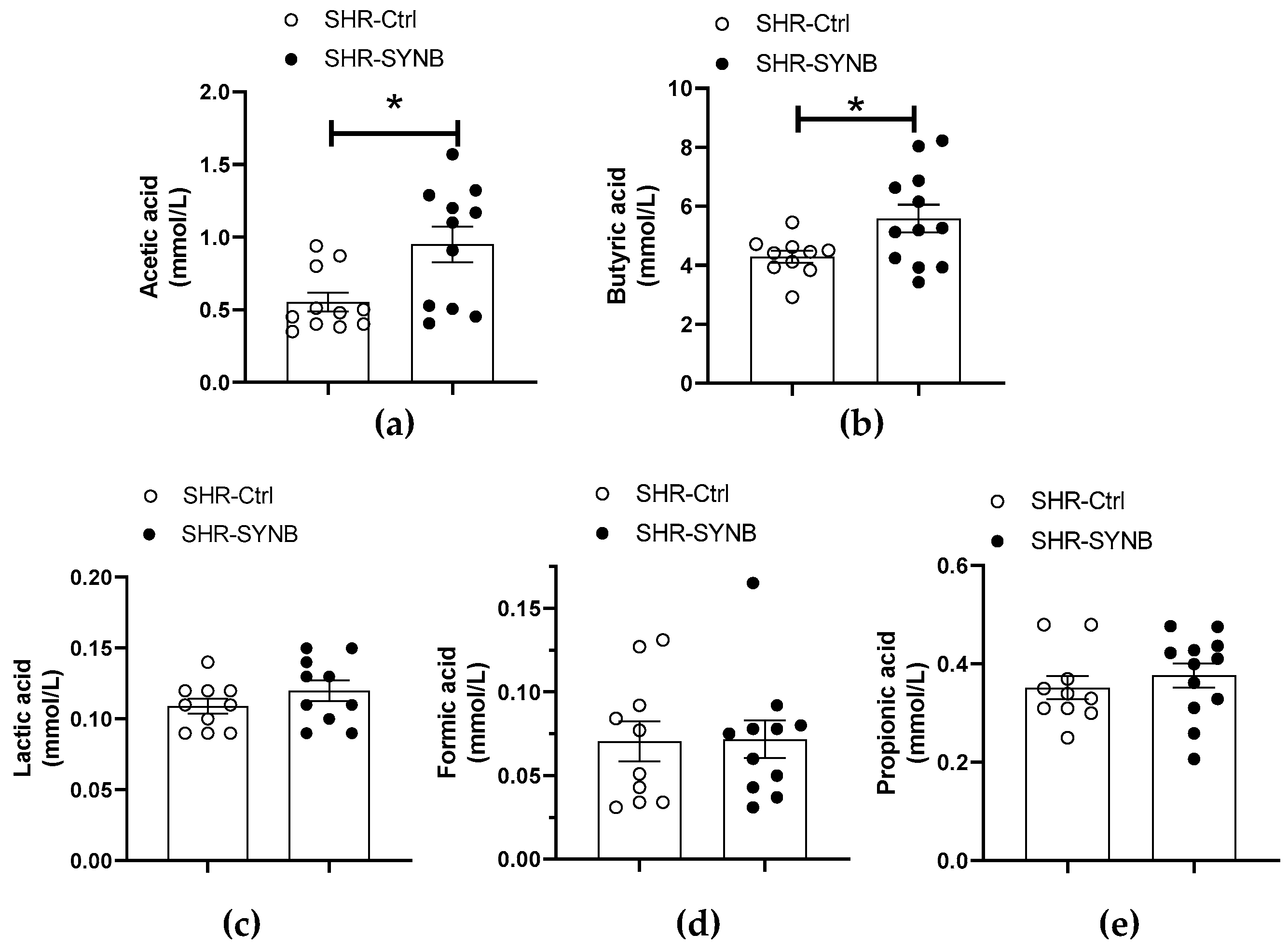
3.1. Systemic Effects of Supplementation with Prodefen®
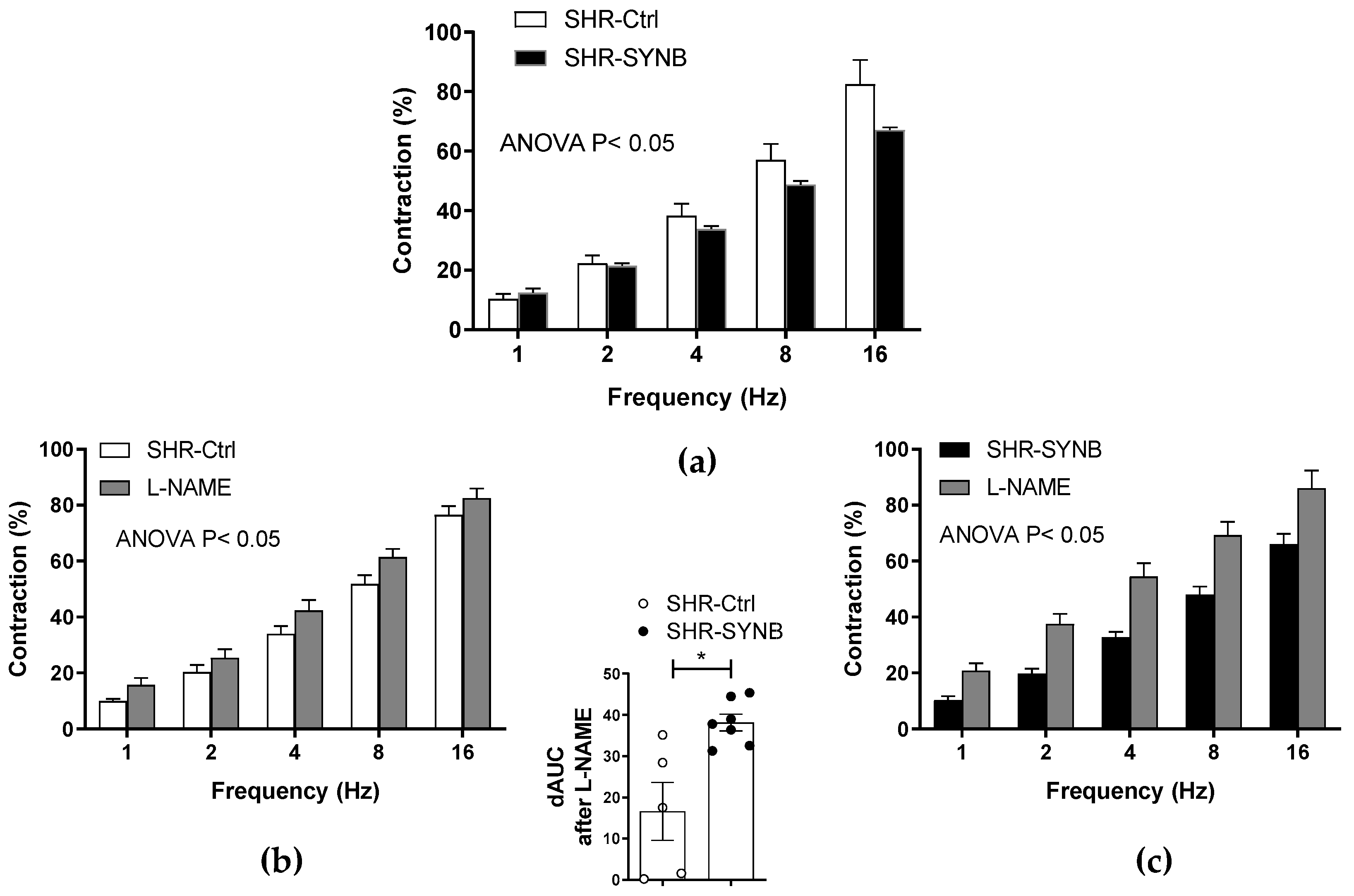
3.2. Contractile Response to Electrical Field Stimulation
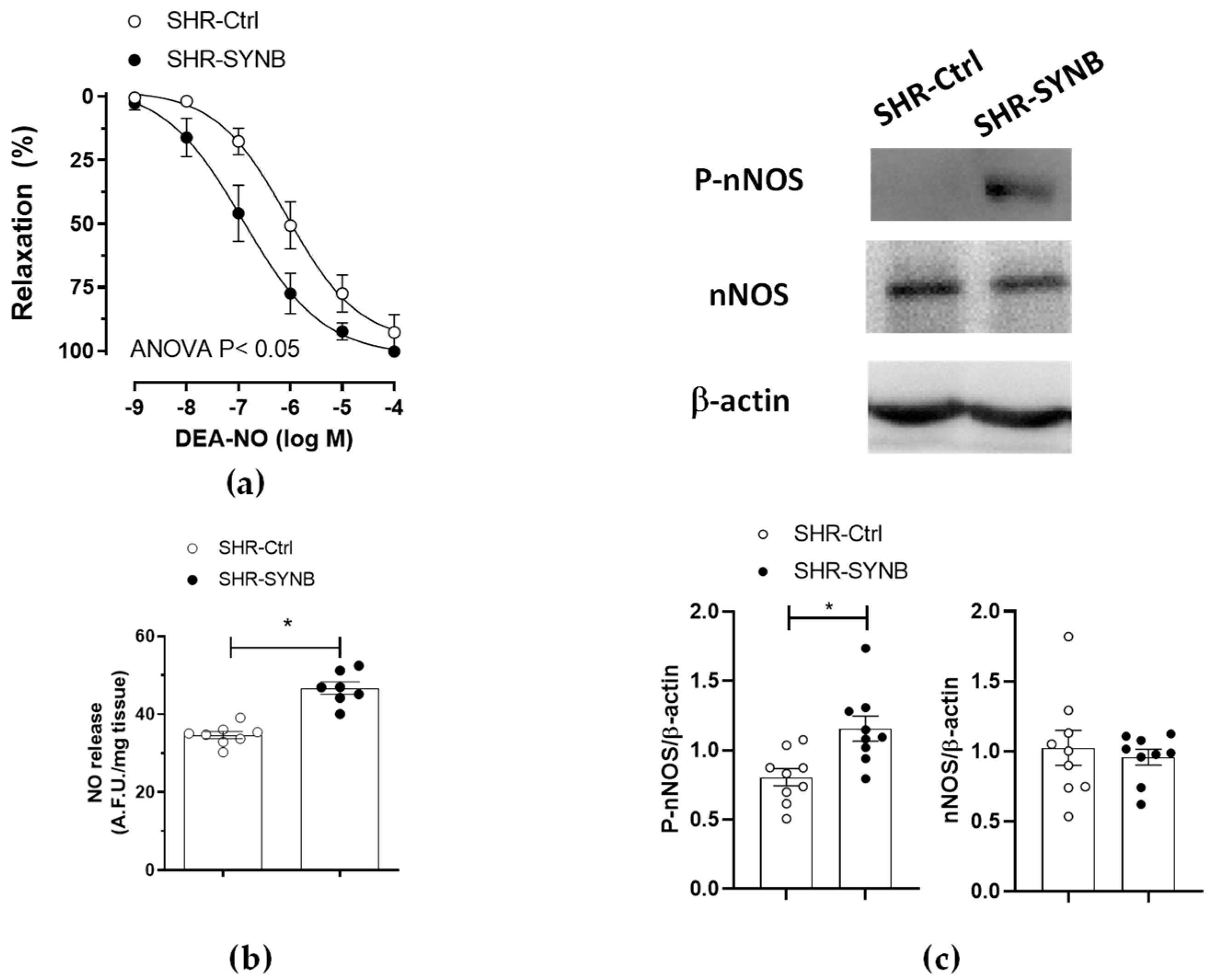
3.3. Modifications on Nitrergic Component of Mesenteric Innervation
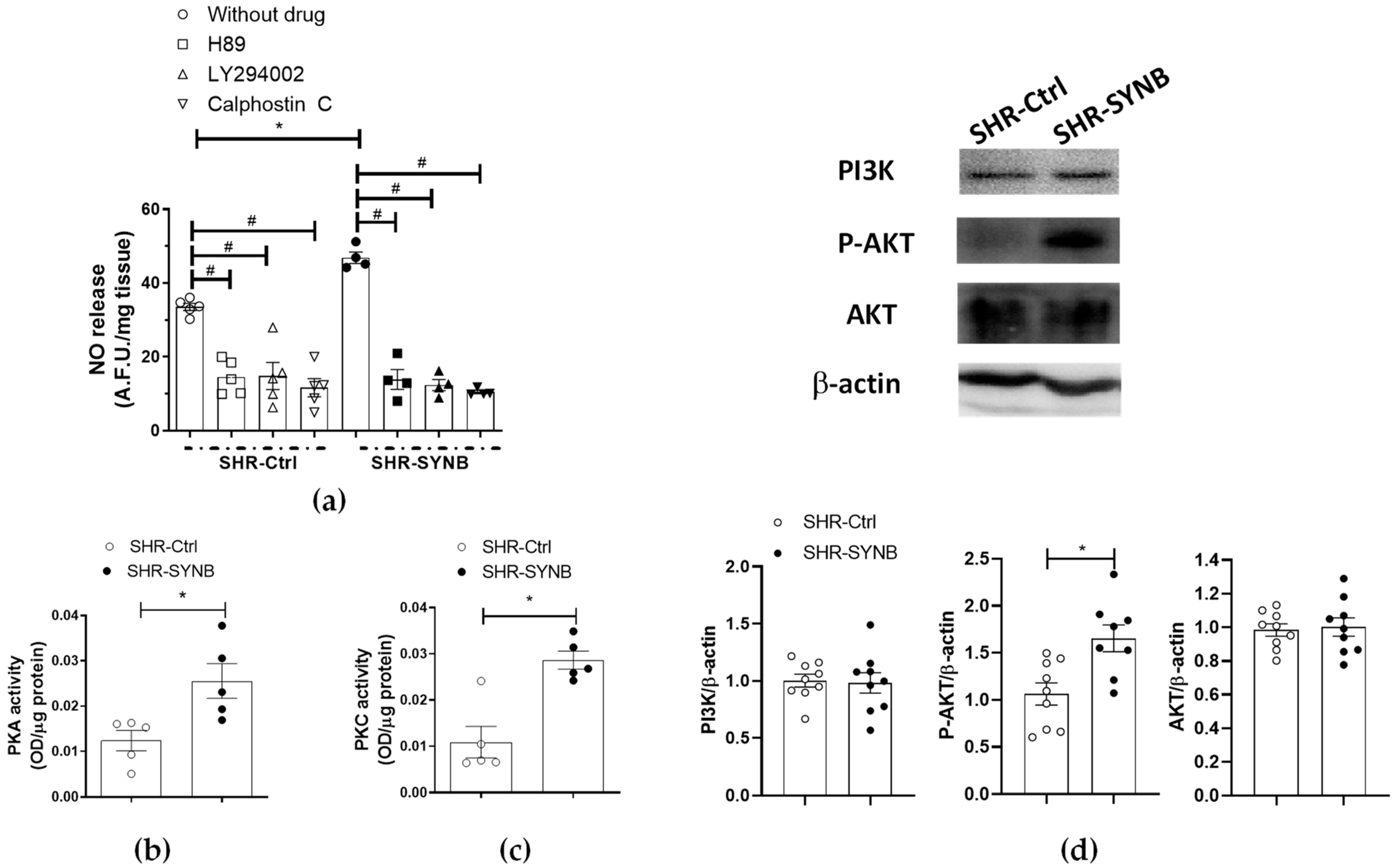
3.4. Mechanisms Implicated in Neuronal Nitric Oxide Release
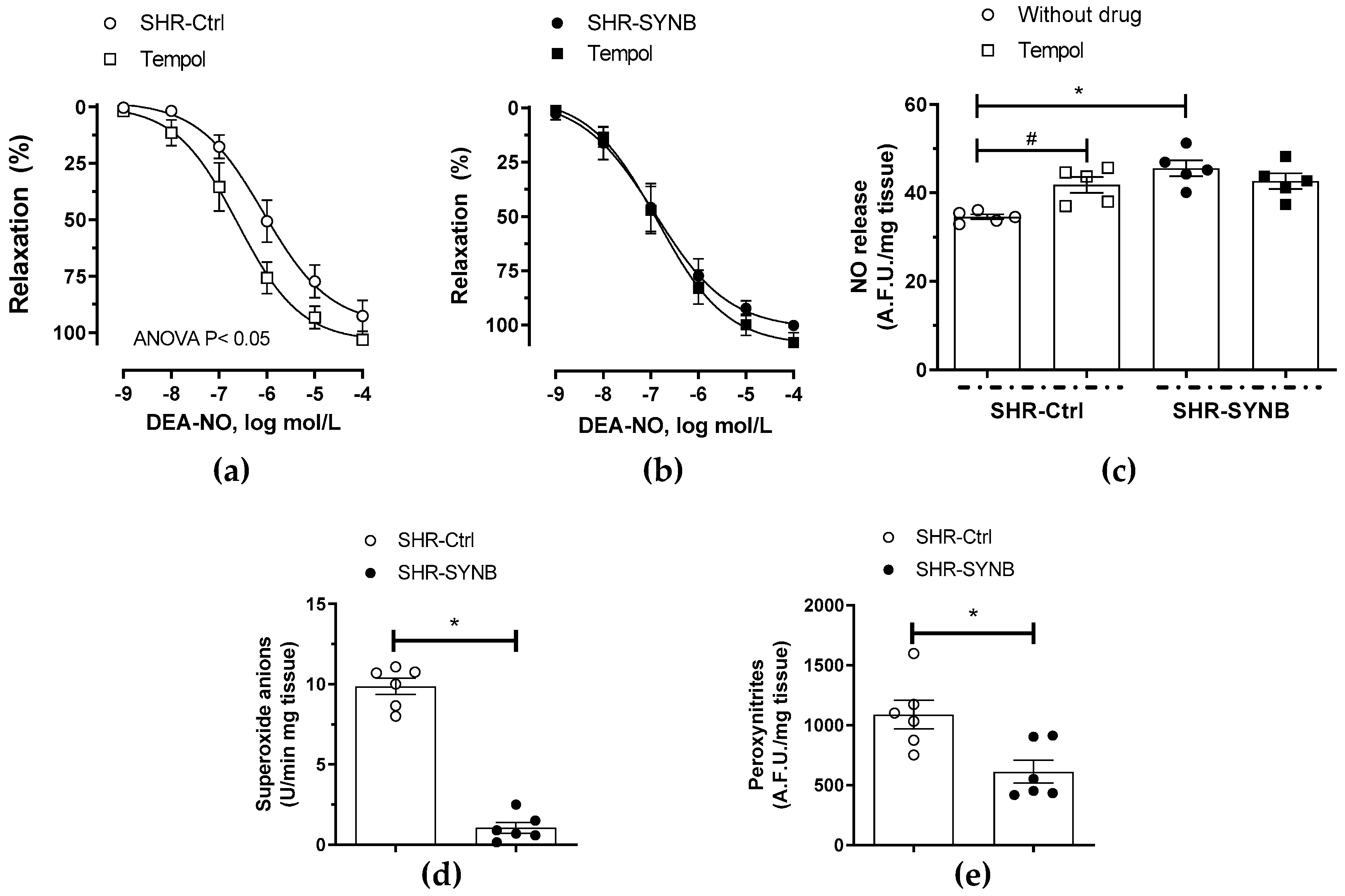
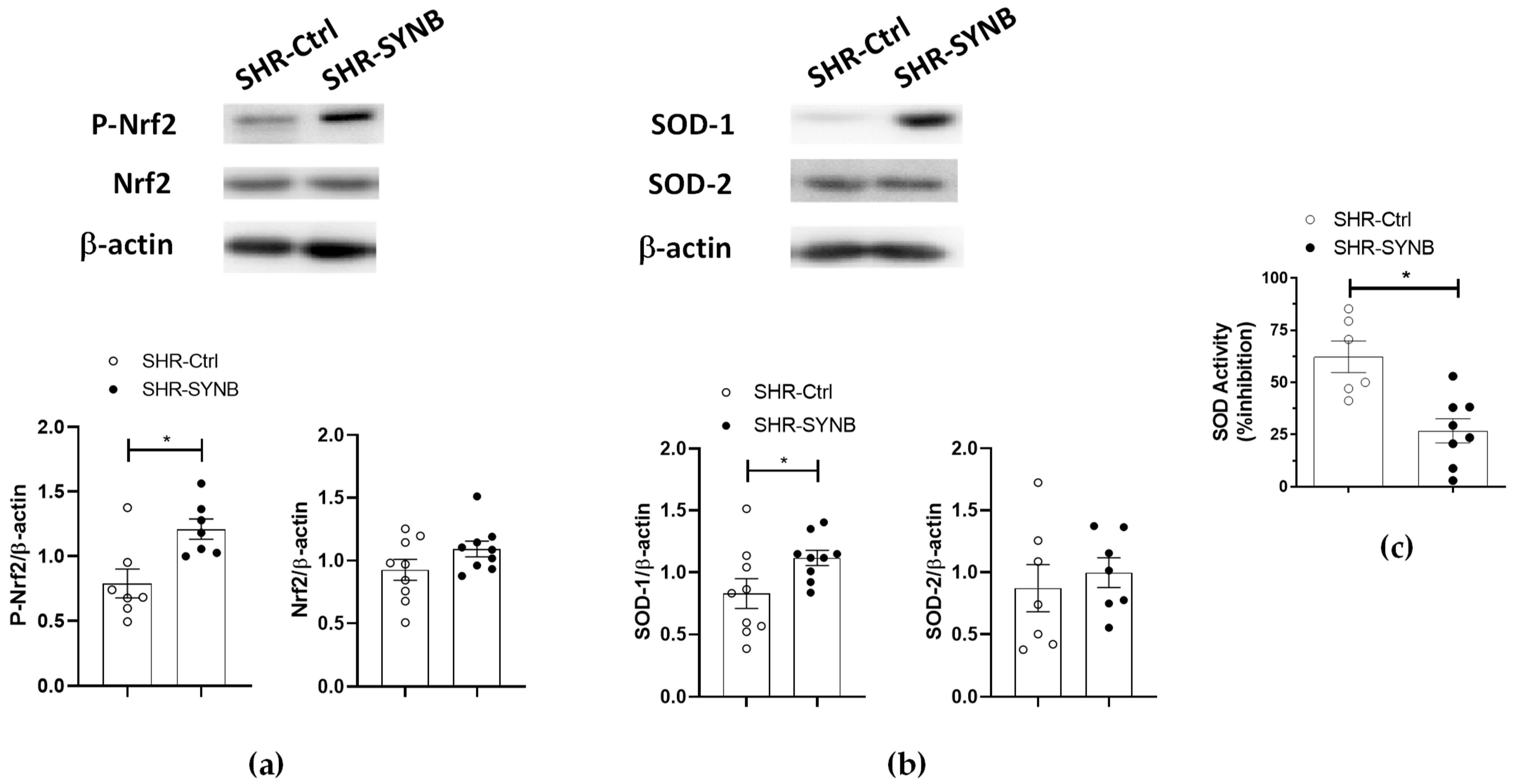
3.5. Neuronal Nitric Oxide Bioavailability: Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Egan, B.M.; Kjeldsen, S.E.; Grassi, G.; Esler, M.; Mancia, G. The global burden of hypertension exceeds 1.4 billion people: Should a systolic blood pressure target below 130 become the universal standard? J. Hypertens. 2019, 37, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Mackay, C.R.; Kaye, D.M. Beyond gut feelings: How the gut microbiota regulates blood pressure. Nat. Rev. Cardiol. 2018, 15, 20–32. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/es/ (accessed on 24 May 2018).
- Carretero, O.A.; Oparil, S. Essential hypertension. Part I: Definition and etiology. Circulation 2000, 101, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Gillis, E.E.; Sullivan, J.C. Sex Differences in Hypertension: Recent Advances. Hypertension 2016, 68, 1322–1327. [Google Scholar] [CrossRef]
- Takala, J. Determinants of splanchnic blood flow. Br. J. Anaesth. 1996, 77, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Sastre, E.; Márquez-Rodas, I.; Blanco-Rivero, J.; Balfagón, G. Inervación perivascular de la arteria mesentérica superior: Implicaciones fisiopatológicas—Perivascular innervation of the superior mesenteric artery: Pathophysiological implications. Rev. Neurol. 2010, 50, 727–737. [Google Scholar] [PubMed]
- Xavier, F. Nitrergic perivascular innervation in health and diseases: Focus on vascular tone regulation. Acta Physiol. 2020, 230, e13484. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhu, D.Y. Neuronal nitric oxide synthase: Structure, subcellular localization, regulation, and clinical implications. Nitric Oxide 2009, 20, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Marín, J.; Ferrer, M.; Balfagón, G. Role of protein kinase C in electrical-stimulation-induced neuronal nitric oxide release in mesenteric arteries from hypertensive rats. Clin. Sci. 2000, 99, 277–283. [Google Scholar] [CrossRef]
- Aras-López, R.; Xavier, F.E.; Ferrer, M.; Balfagón, G. Dexamethasone decreases neuronal nitric oxide release in mesenteric arteries from hypertensive rats through decreased protein kinase C activation. Clin. Sci. 2009, 117, 305–312. [Google Scholar] [CrossRef][Green Version]
- Llévenes, P.; Rodrigues-Díez, R.; Cros-Brunsó, L.; Prieto, M.I.; Casaní, L.; Balfagón, G.; Blanco-Rivero, J. Beneficial Effect of a Multistrain Synbiotic Prodefen® Plus on the Systemic and Vascular Alterations Associated with Metabolic Syndrome in Rats: The Role of the Neuronal Nitric Oxide Synthase and Protein Kinase, A. Nutrients 2020, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Cros-Brunsó, L.; Camacho-Rodríguez, L.; Martínez-González, Á.; Llévenes, P.; Salaices, M.; García-Redondo, A.B.; Blanco-Rivero, J. A Blunted Sympathetic Function and an Enhanced Nitrergic Activity Contribute to Reduce Mesenteric Resistance in Hyperthyroidism. Int. J. Mol. Sci. 2021, 22, 570. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Briones, A.M. Reactive oxygen species and vascular biology: Implications in human hypertension. Hypertens. Res. 2011, 34, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Rivero, J.; Roque, F.R.; Sastre, E.; Caracuel, L.; Couto, G.K.; Avendaño, M.S.; Paula, S.M.; Rossoni, L.V.; Salaices, M.; Balfagón, G. Aerobic exercise training increases neuronal nitric oxide release and bioavailability and decreases noradrenaline release in mesenteric artery from spontaneously hypertensive rats. J. Hypertens. 2013, 31, 916–926. [Google Scholar] [CrossRef]
- Dreger, H.; Westphal, K.; Weller, A.; Baumann, G.; Stangl, V.; Meiners, S.; Stangl, K. Nrf2-dependent upregulation of antioxidative enzymes: A novel pathway for proteasome inhibitor-mediated cardioprotection. Cardiovasc. Res. 2009, 83, 354–361. [Google Scholar] [CrossRef]
- Al-Sawaf, O.; Clarner, T.; Fragoulis, A.; Kan, Y.W.; Pufe, T.; Streetz, K.; Wruck, C.J. Nrf2 in health and disease: Current and future clinical implications. Clin. Sci. 2015, 129, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Barančík, M.; Grešová, L.; Barteková, M.; Dovinová, I. Nrf2 as a key player of redox regulation in cardiovascular diseases. Physiol. Res. 2016, 65 (Suppl. S1), S1–S10. [Google Scholar] [CrossRef]
- Sastre, E.; Caracuel, L.; Balfagón, G.; Blanco-Rivero, J. Aerobic exercise training increases nitrergic innervation function and decreases sympathetic innervation function in mesenteric artery from rats fed a high-fat diet. J. Hypertens. 2015, 33, 1819–1830. [Google Scholar] [CrossRef]
- Javkhedkar, A.A.; Quiroz, Y.; Rodriguez-Iturbe, B.; Vaziri, N.D.; Lokhandwala, M.F.; Banday, A.A. Resveratrol restored Nrf2 function, reduced renal inflammation, and mitigated hypertension in spontaneously hypertensive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R840–R846. [Google Scholar] [CrossRef] [PubMed]
- Beale, A.L.; Kaye, D.M.; Marques, F.Z. The role of the gut microbiome in sex differences in arterial pressure. Biol. Sex Differ. 2019, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef]
- Icaza-Chávez, M.E. Microbiota intestinal en la salud y la enfermedad. Rev. Gastroenterol. Mex. 2013, 78, 240–248. [Google Scholar] [CrossRef]
- Gómez-Guzmán, M.; Toral, M.; Romero, M.; Jiménez, R.; Galindo, P.; Sánchez, M.; Zarzuelo, M.J.; Olivares, M.; Gálvez, J.; Duarte, J. Antihypertensive effects of probiotics Lactobacillus strains in spontaneously hypertensive rats. Mol. Nutr. Food Res. 2015, 59, 2326–2336. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, H.M.; Koning, C.J.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Diplock, A.T.; Aggett, P.J.; Ashwel1, M.; Bornet, F.; Fern, E.B.; Roberfroid, M.B. Scientific concepts of functional foods in Europe: Consensus Document. Br. J. Nutr. 1999, 81 (Suppl. S1), S1–S27. [Google Scholar] [CrossRef]
- Sanz, M.L.; Polemis, N.; Morales, V.; Corzo, N.; Drakoularakou, A.; Gibson, G.R.; Rastall, R.A. In vitro investigation into the potential prebiotic activity of honey oligosaccharides. J. Agric. Food Chem. 2005, 53, 2914–2921. [Google Scholar] [CrossRef]
- Nielsen, K.C.; Owman, C.; Sporrong, B. Ultrastructure of the autonomic innervation apparatus in the main pial arteries of rats and cats. Brain Res. 1971, 27, 25–32. [Google Scholar] [CrossRef]
- Sastre, E.; Blanco-Rivero, J.; Caracuel, L.; Lahera, V.; Balfagón, G. Effects of lipopolysaccharide on the neuronal control of mesenteric vascular tone in rats: Mechanisms involved. Shock 2012, 38, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Sastre, E.; Caracuel, L.; Blanco-Rivero, J.; Callejo, M.; Xavier, F.E.; Balfagón, G. Biphasic Effect of Diabetes on Neuronal Nitric Oxide Release in Rat Mesenteric Arteries. PLoS ONE 2016, 11, e0156793. [Google Scholar] [CrossRef]
- Blanco-Rivero, J.; Cachofeiro, V.; Lahera, V.; Aras-Lopez, R.; Márquez-Rodas, I.; Salaices, M.; Xavier, F.E.; Ferrer, M.; Balfagón, G. Participation of prostacyclin in endothelial dysfunction induced by aldosterone in normotensive and hypertensive rats. Hypertension 2005, 46, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Chen, K.; Quan, D.; Kang, L.; Yang, D.; Wu, H.; Yan, M.; Wu, S.; Lv, L.; Zhang, G. The Combination of Scutellaria baicalensis Georgi and Sophora japonica, L. ameliorate Renal Function by Regulating Gut Microbiota in Spontaneously Hypertensive Rats. Front. Pharmacol. 2021, 11, 575294. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.A.; Shafiq, M.; Reza, M.I.; Bharati, P.; Husain, A.; Singh, P.; Hanif, K.; Gayen, J.R. Ethanolic extract of Cissus quadrangularis improves vasoreactivity by modulation of eNOS expression and oxidative stress in spontaneously hypertensive rats. Clin. Exp. Hypertens. 2022, 44, 63–71. [Google Scholar] [CrossRef]
- Khalesi, S.; Sun, J.; Buys, N.; Jayasinghe, R. Effect of probiotics on blood pressure: A systematic review and meta-analysis of randomized, controlled trials. Hypertension 2014, 64, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Hadi, A.; Pourmasoumi, M.; Kazemi, M.; Najafgholizadeh, A.; Marx, W. Efficacy of synbiotic interventions on blood pressure: A systematic review and meta-analysis of clinical trials. Crit. Rev. Food Sci. Nutr. 2021, 1–11. [Google Scholar] [CrossRef]
- Arabi, S.M.; Bahrami, L.S.; Rahnama, I.; Sahebkar, A. Impact of synbiotic supplementation on cardiometabolic and anthropometric indices in patients with metabolic syndrome: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2022, 176, 106061. [Google Scholar] [CrossRef] [PubMed]
- Hernanz, R.; Briones, A.M.; Salaices, M.; Alonso, M.J. New roles for old pathways? A circuitous relationship between reactive oxygen species and cyclo-oxygenase in hypertension. Clin. Sci. 2014, 126, 111–121. [Google Scholar] [CrossRef] [PubMed]
- González-Amor, M.; García-Redondo, A.B.; Jorge, I.; Zalba, G.; Becares, M.; Ruiz-Rodríguez, M.J.; Rodríguez, C.; Bermeo, H.; Rodrigues-Díez, R.; Rios, F.J.; et al. Interferon stimulated gene 15 pathway is a novel mediator of endothelial dysfunction and aneurysms development in angiotensin II infused mice through increased oxidative stress. Cardiovasc. Res. 2021, cvab321. [Google Scholar] [CrossRef] [PubMed]
- Rizzoni, D.; De Ciuceis, C.; Szczepaniak, P.; Paradis, P.; Schiffrin, E.L.; Guzik, T.J. Immune System and Microvascular Remodeling in Humans. Hypertension 2022, 79, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Bomfim, G.F.; Dos Santos, R.A.; Oliveira, M.A.; Giachini, F.R.; Akamine, E.H.; Tostes, R.C.; Fortes, Z.B.; Webb, R.C.; Carvalho, M.H. Toll-like receptor 4 contributes to blood pressure regulation and vascular contraction in spontaneously hypertensive rats. Clin. Sci. 2012, 122, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Diez-Echave, P.; Vezza, T.; Rodríguez-Nogales, A.; Hidalgo-Garcia, L.; Garrido-Mesa, J.; Ruiz-Malagon, A.; Molina-Tijeras, J.A.; Romero, M.; Robles-Vera, I.; Leyva-Jiménez, F.J.; et al. The Beneficial Effects of Lippia Citriodora Extract on Diet-Induced Obesity in Mice Are Associated with Modulation in the Gut Microbiota Composition. Mol. Nutr. Food Res. 2020, 64, e2000005. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Muralitharan, R.R.; Marques, F.Z. Diet-related gut microbial metabolites and sensing in hypertension. J. Hum. Hypertens. 2021, 35, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.K.; McKenzie, C.; Mariño, E.; Macia, L.; Mackay, C.R. Metabolite-Sensing G Protein-Coupled Receptors-Facilitators of Diet-Related Immune Regulation. Annu. Rev. Immunol. 2017, 35, 371–402. [Google Scholar] [CrossRef] [PubMed]
- Norlander, A.E.; Madhur, M.S.; Harrison, D.G. Correction: The immunology of hypertension. J. Exp. Med. 2018, 215, 719. [Google Scholar] [CrossRef] [PubMed]
- Drummond, G.R.; Vinh, A.; Guzik, T.J.; Sobey, C.G. Immune mechanisms of hypertension. Nat. Rev. Immunol. 2019, 19, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, J.; Kasubuchi, M.; Nakajima, A.; Irie, J.; Itoh, H.; Kimura, I. The role of short-chain fatty acid on blood pressure regulation. Curr. Opin. Nephrol. Hypertens. 2016, 25, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Bartolomaeus, H.; Avery, E.G.; Bartolomaeus, T.; Kozhakhmetov, S.; Zhumadilov, Z.; Müller, D.N.; Wilck, N.; Kushugulova, A.; Forslund, S.K. Blood pressure changes correlate with short-chain fatty acid production potential shifts under a synbiotic intervention. Cardiovasc. Res. 2020, 116, 1252–1253. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Sánchez, M.; Gómez-Guzmán, M.; Romero, M.; Yang, T.; Izquierdo-Garcia, J.L.; Jiménez, R.; Ruiz-Cabello, J.; et al. Probiotics Prevent Dysbiosis and the Rise in Blood Pressure in Genetic Hypertension: Role of Short-Chain Fatty Acids. Mol. Nutr. Food Res. 2020, 64, e1900616. [Google Scholar] [CrossRef] [PubMed]
- Le Blay, G.; Michel, C.; Blottière, H.M.; Cherbut, C. Prolonged intake of fructo-oligosaccharides induces a short-term elevation of lactic acid-producing bacteria and a persistent increase in cecal butyrate in rats. J. Nutr. 1999, 129, 2231–2235. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Jädert, C.; Holm, L.; Nyman, M.E. Propionic and butyric acids, formed in the caecum of rats fed highly fermentable dietary fibre, are reflected in portal and aortic serum. Br. J. Nutr. 2013, 110, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Adnan, S.; Nelson, J.W.; Ajami, N.J.; Venna, V.R.; Petrosino, J.F.; Bryan, R.M., Jr.; Durgan, D.J. Alterations in the gut microbiota can elicit hypertension in rats. Physiol. Genom. 2017, 49, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.W.; Romano, N.; Ebrahimi, M.; Natrah, I. The effects of dietary fructooligosaccharide on growth, intestinal short chain fatty acids level and hepatopancreatic condition of the giant freshwater prawn (macrobrachium rosenbergii) post-larvae. Aquaculture 2017, 469, 95–101. [Google Scholar] [CrossRef]
- Toral, M.; Gómez-Guzmán, M.; Jiménez, R.; Romero, M.; Sánchez, M.; Utrilla, M.P.; Garrido-Mesa, N.; Rodríguez-Cabezas, M.E.; Olivares, M.; Gálvez, J.; et al. The probiotic Lactobacillus coryniformis CECT5711 reduces the vascular pro-oxidant and pro-inflammatory status in obese mice. Clin. Sci. 2014, 127, 33–45. [Google Scholar] [CrossRef]
- Malik, M.; Suboc, T.M.; Tyagi, S.; Salzman, N.; Wang, J.; Ying, R.; Tanner, M.J.; Kakarla, M.; Baker, J.E.; Widlansky, M.E. Lactobacillus plantarum 299v Supplementation Improves Vascular Endothelial Function and Reduces Inflammatory Biomarkers in Men with Stable Coronary Artery Disease. Circ. Res. 2018, 123, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Toral, M.; Duarte, J. Microbiota and Hypertension: Role of the Sympathetic Nervous System and the Immune System. Am. J. Hypertens. 2020, 33, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Santisteban, M.M.; Qi, Y.; Zubcevic, J.; Kim, S.; Yang, T.; Shenoy, V.; Cole-Jeffrey, C.T.; Lobaton, G.O.; Stewart, D.C.; Rubiano, A.; et al. Hypertension-Linked Pathophysiological Alterations in the Gut. Circ. Res. 2017, 120, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Zubcevic, J.; Richards, E.M.; Yang, T.; Kim, S.; Sumners, C.; Pepine, C.J.; Raizada, M.K. Impaired Autonomic Nervous System-Microbiome Circuit in Hypertension. Circ. Res. 2019, 125, 104–116. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Sánchez, M.; Gómez-Guzmán, M.; Muñoz, R.; Algieri, F.; Vezza, T.; Jiménez, R.; Gálvez, J.; et al. Changes to the gut microbiota induced by losartan contributes to its antihypertensive effects. Br. J. Pharmacol. 2020, 177, 2006–2023. [Google Scholar] [CrossRef]
- Toral, M.; Robles-Vera, I.; de la Visitación, N.; Romero, M.; Yang, T.; Sánchez, M.; Gómez-Guzmán, M.; Jiménez, R.; Raizada, M.K.; Duarte, J. Critical Role of the Interaction Gut Microbiota–Sympathetic Nervous System in the Regulation of Blood Pressure. Front. Physiol. 2019, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; Tan, F.; Liao, W.; Wang, Q.; Mu, J.; Zhou, X.; Yang, Z.; Zhao, X. Isolation and Identification of Lactobacillus plantarum HFY05 from Natural Fermented Yak Yogurt and Its Effect on Alcoholic Liver Injury in Mice. Microorganisms 2019, 7, 530. [Google Scholar] [CrossRef]
- Aboulgheit, A.; Karbasiafshar, C.; Zhang, Z.; Sabra, M.; Shi, G.; Tucker, A.; Sodha, N.; Abid, M.R.; Sellke, F.W. Lactobacillus plantarum probiotic induces Nrf2-mediated antioxidant signaling and eNOS expression resulting in improvement of myocardial diastolic function. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H839–H849. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.; Zhang, L.; Chen, Q.; Tan, F.; Zhao, X. Effect of Lactobacillus fermentum HFY03 on the Antifatigue and Antioxidation Ability of Running Exhausted Mice. Oxid. Med. Cell. Longev. 2021, 2021, 8013681. [Google Scholar] [CrossRef] [PubMed]
- Iring, A.; Jin, Y.J.; Albarrán-Juárez, J.; Siragusa, M.; Wang, S.; Dancs, P.T.; Nakayama, A.; Tonack, S.; Chen, M.; Künne, C.; et al. Shear stress-induced endothelial adrenomedullin signaling regulates vascular tone and blood pressure. J. Clin. Investig. 2019, 129, 2775–2791. [Google Scholar] [CrossRef]
- Segawa, Y.; Hashimoto, H.; Maruyama, S.; Shintani, M.; Ohno, H.; Nakai, Y.; Osera, T.; Kurihara, N. Dietary capsaicin-mediated attenuation of hypertension in a rat model of renovascular hypertension. Clin. Exp. Hypertens. 2020, 42, 352–359. [Google Scholar] [CrossRef]
- Lin, H.J.; Mahendran, R.; Huang, H.Y.; Chiu, P.L.; Chang, Y.M.; Day, C.H.; Chen, R.J.; Padma, V.V.; Liang-Yo, Y.; Kuo, W.W.; et al. Aqueous extract of Solanum nigrum attenuates Angiotensin-II induced cardiac hypertrophy and improves cardiac function by repressing protein kinase C-ζ to restore HSF2 deSUMOlyation and Mel-18-IGF-IIR signaling suppression. J. Ethnopharmacol. 2022, 284, 114728. [Google Scholar] [CrossRef]
- Heuvelin, E.; Lebreton, C.; Bichara, M.; Cerf-Bensussan, N.; Heyman, M. A Bifidobacterium probiotic strain and its soluble factors alleviate chloride secretion by human intestinal epithelial cells. J. Nutr. 2010, 140, 7–11. [Google Scholar] [CrossRef]
- Zhou, Y.K.; Qin, H.L.; Zhang, M.; Shen, T.Y.; Chen, H.Q.; Ma, Y.L.; Chu, Z.X.; Zhang, P.; Liu, Z.H. Effects of Lactobacillus plantarum on gut barrier function in experimental obstructive jaundice. World J. Gastroenterol. 2012, 18, 3977–3991. [Google Scholar] [CrossRef]
- Ting, W.J.; Kuo, W.W.; Kuo, C.H.; Yeh, Y.L.; Shen, C.Y.; Chen, Y.H.; Ho, T.J.; Viswanadha, V.P.; Chen, Y.H.; Huang, C.Y. Supplementary heat-killed Lactobacillus reuteri GMNL-263 ameliorates hyperlipidaemic and cardiac apoptosis in high-fat diet-fed hamsters to maintain cardiovascular function. Br. J. Nutr. 2015, 114, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Paveljšek, D.; Juvan, P.; Košir, R.; Rozman, D.; Hacin, B.; Ivičak-Kocjan, K.; Rogelj, I. Lactobacillus fermentum L930BB and Bifidobacterium animalis subsp. animalis IM386 initiate signalling pathways involved in intestinal epithelial barrier protection. Benef. Microbes 2018, 9, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Luo, Y.; Liu, Z.; Yang, P.; Gui, Y. Probiotics SOD inhibited food allergy via downregulation of STAT6-TIM4 signaling on DCs. Mol. Immunol. 2018, 103, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Jia, L.; Su, Y.; Du, J.; Guo, L.; Luo, Z.; Liu, Y. Lactobacillus reuteri extracts promoted wound healing via PI3K/AKT/β-catenin/TGFβ1 pathway. Stem Cell Res. Ther. 2019, 10, 243. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Sharma, A.; Gupta, M.; Padwad, Y.; Sharma, R. Cell-Free Culture Supernatant of Probiotic Lactobacillus fermentum Protects Against H2O2-Induced Premature Senescence by Suppressing ROS-Akt-mTOR Axis in Murine Preadipocytes. Probiotics Antimicrob. Proteins 2020, 12, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Llévenes, P.; Balfagón, G.; Blanco-Rivero, J. Thyroid hormones affect nitrergic innervation function in rat mesenteric artery: Role of the PI3K/AKT pathway. Vascul. Pharmacol. 2018, 108, 36–45. [Google Scholar] [CrossRef]
- Xu, S.; Touyz, R.M. Reactive oxygen species and vascular remodelling in hypertension: Still alive. Can. J. Cardiol. 2006, 22, 947–951. [Google Scholar] [CrossRef]
- Wei, Y.; Whaley-Connell, A.T.; Chen, K.; Habibi, J.; Uptergrove, G.M.; Clark, S.E.; Stump, C.S.; Ferrario, C.M.; Sowers, J.R. NADPH oxidase contributes to vascular inflammation, insulin resistance, and remodeling in the transgenic (mRen2) rat. Hypertension 2007, 50, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Paravicini, T.M.; Touyz, R.M. NADPH oxidases, reactive oxygen species, and hypertension: Clinical implications and therapeutic possibilities. Diabetes Care 2008, 31 (Suppl. S2), S170–S180. [Google Scholar] [CrossRef] [PubMed]
- Dinh, Q.N.; Drummond, G.R.; Sobey, C.G.; Chrissobolis, S. Roles of inflammation, oxidative stress, and vascular dysfunction in hypertension. BioMed Res. Int. 2014, 2014, 406960. [Google Scholar] [CrossRef] [PubMed]
- Sastre, E.; Caracuel, L.; Balfagón, G.; Blanco-Rivero, J. Aerobic exercise training avoids endothelial and neuronal dysfunction produced by a high fat diet in rat mesenteric artery. Trauma Fund MAPFRE 2014, 25, 164–173. [Google Scholar]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Sánchez, M.; Romero, M.; Olivares, M.; Jiménez, R.; Duarte, J. The Probiotic Lactobacillus fermentum Prevents Dysbiosis and Vascular Oxidative Stress in Rats with Hypertension Induced by Chronic Nitric Oxide Blockade. Mol. Nutr. Food Res. 2018, 62, e1800298. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Bishnoi, M.; Kondepudi, K.K.; Shukla, G. Synbiotic (Lactiplantibacillus pentosus GSSK2 and isomalto-oligosaccharides) supplementation modulates pathophysiology and gut dysbiosis in experimental metabolic syndrome. Sci. Rep. 2021, 11, 21397. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.U.; Kim, H.W.; Kim, W. Effects of Probiotics, Prebiotics, and Synbiotics on Uremic Toxins, Inflammation, and Oxidative Stress in Hemodialysis Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Clin. Med. 2021, 10, 4456. [Google Scholar] [CrossRef]
- Cruz, B.; de Sousa Moraes, L.F.; De Nadai Marcon, L.; Dias, K.A.; Murad, L.B.; Sarandy, M.M.; Conceição, L.; Gonçalves, R.V.; Ferreira, C.; Peluzio, M. Evaluation of the efficacy of probiotic VSL#3 and synbiotic VSL#3 and yacon-based product in reducing oxidative stress and intestinal permeability in mice induced to colorectal carcinogenesis. J. Food Sci. 2021, 86, 1448–1462. [Google Scholar] [CrossRef] [PubMed]
- Simões, R.P.; Fardin, P.B.A.; Simões, M.R.; Vassallo, D.V.; Padilha, A.S. Long-term Mercury Exposure Accelerates the Development of Hypertension in Prehypertensive Spontaneously Hypertensive Rats Inducing Endothelial Dysfunction: The Role of Oxidative Stress and Cyclooxygenase-2. Biol. Trace Elem. Res. 2020, 196, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Hosseinifard, E.S.; Bavafa-Valenlia, K.; Saghafi-Asl, M.; Morshedi, M. Cardioprotective effects of the proline-rich oligopeptide Bj-PRO-7a in spontaneously hypertensive rats. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1693–1703. [Google Scholar] [CrossRef]
- Hosseinifard, E.S.; Bavafa-Valenlia, K.; Saghafi-Asl, M.; Morshedi, M. Antioxidative and Metabolic Effects of Lactobacillus plantarum, Inulin, and Their Synbiotic on the Hypothalamus and Serum of Healthy Rats. Nutr. Metab. Insights 2020, 13, 1178638820925092. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhang, B.; Ru, Q.J.; Chen, X.M.; Lv, B.D. Tocopheryl quinone improves non-alcoholic steatohepatitis (NASH) associated dysmetabolism of glucose and lipids by upregulating the expression of glucagon-like peptide 1 (GLP-1) via restoring the balance of intestinal flora in rats. Pharm. Biol. 2021, 59, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.P.; Nurrahma, B.A.; Kumar, R.; Wu, C.H.; Yeh, T.H.; Chiu, C.C.; Lee, Y.P.; Liao, Y.C.; Huang, C.H.; Yeh, Y.T.; et al. Probiotic Enhancement of Antioxidant Capacity and Alterations of Gut Microbiota Composition in 6-Hydroxydopamin-Induced Parkinson’s Disease Rats. Antioxidants 2021, 10, 1823. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides produced by Lactobacillus rhamnosus GG alleviate hydrogen peroxide-induced intestinal oxidative damage and apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. Food Funct. 2021, 12, 9632–9641. [Google Scholar] [CrossRef]
- Wu, Y.; Hu, J.; Long, X.; Pan, Y.; Mu, J.; Park, K.Y.; Zhao, X. Lactobacillus plantarum ZS62 Alleviates Alcohol-Induced Gastric Injury in Mice via an Anti-Oxidative Mechanism. Drug Des. Dev. Ther. 2021, 15, 1667–1676. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Albiñana, P.; Martínez-González, Á.; Camacho-Rodríguez, L.; Ferreira-Lazarte, Á.; Villamiel, M.; Rodrigues-Díez, R.; Balfagón, G.; García-Redondo, A.B.; Prieto-Nieto, M.I.; Blanco-Rivero, J. Supplementation with the Symbiotic Formulation Prodefen® Increases Neuronal Nitric Oxide Synthase and Decreases Oxidative Stress in Superior Mesenteric Artery from Spontaneously Hypertensive Rats. Antioxidants 2022, 11, 680. https://doi.org/10.3390/antiox11040680
Méndez-Albiñana P, Martínez-González Á, Camacho-Rodríguez L, Ferreira-Lazarte Á, Villamiel M, Rodrigues-Díez R, Balfagón G, García-Redondo AB, Prieto-Nieto MI, Blanco-Rivero J. Supplementation with the Symbiotic Formulation Prodefen® Increases Neuronal Nitric Oxide Synthase and Decreases Oxidative Stress in Superior Mesenteric Artery from Spontaneously Hypertensive Rats. Antioxidants. 2022; 11(4):680. https://doi.org/10.3390/antiox11040680
Chicago/Turabian StyleMéndez-Albiñana, Pablo, Ángel Martínez-González, Laura Camacho-Rodríguez, Álvaro Ferreira-Lazarte, Mar Villamiel, Raquel Rodrigues-Díez, Gloria Balfagón, Ana B. García-Redondo, Mª Isabel Prieto-Nieto, and Javier Blanco-Rivero. 2022. "Supplementation with the Symbiotic Formulation Prodefen® Increases Neuronal Nitric Oxide Synthase and Decreases Oxidative Stress in Superior Mesenteric Artery from Spontaneously Hypertensive Rats" Antioxidants 11, no. 4: 680. https://doi.org/10.3390/antiox11040680
APA StyleMéndez-Albiñana, P., Martínez-González, Á., Camacho-Rodríguez, L., Ferreira-Lazarte, Á., Villamiel, M., Rodrigues-Díez, R., Balfagón, G., García-Redondo, A. B., Prieto-Nieto, M. I., & Blanco-Rivero, J. (2022). Supplementation with the Symbiotic Formulation Prodefen® Increases Neuronal Nitric Oxide Synthase and Decreases Oxidative Stress in Superior Mesenteric Artery from Spontaneously Hypertensive Rats. Antioxidants, 11(4), 680. https://doi.org/10.3390/antiox11040680