
Lipophilic Grape Seed Proanthocyanidin Exerts Anti-Cervical Cancer Effects in HeLa Cells and a HeLa-Derived Xenograft Zebrafish Model



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of LGSP
2.3. Cell Culture
2.4. Cell Proliferation Analysis
2.5. Cell Apoptosis Analysis
2.6. Cell Cycle Distribution Analysis
2.7. Determination of ROS Production
2.8. Measurement of MMP
2.9. Western Blot Analysis
2.10. Zebrafish Maintenance
2.11. Embryo Acute Toxicity Test
2.12. HeLa-Derived Xenograft Zebrafish Model
2.13. Statistical Analysis
3. Results
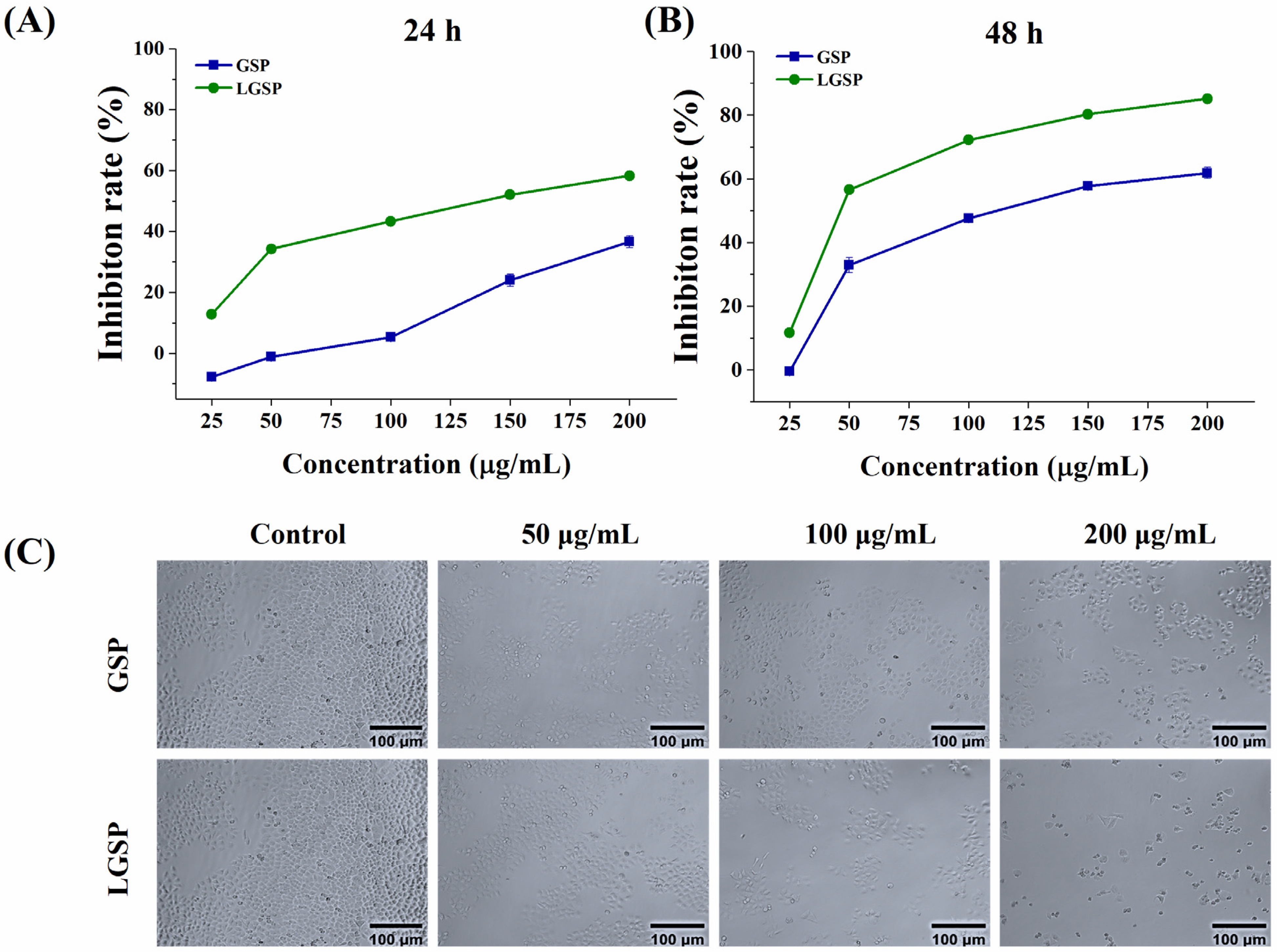
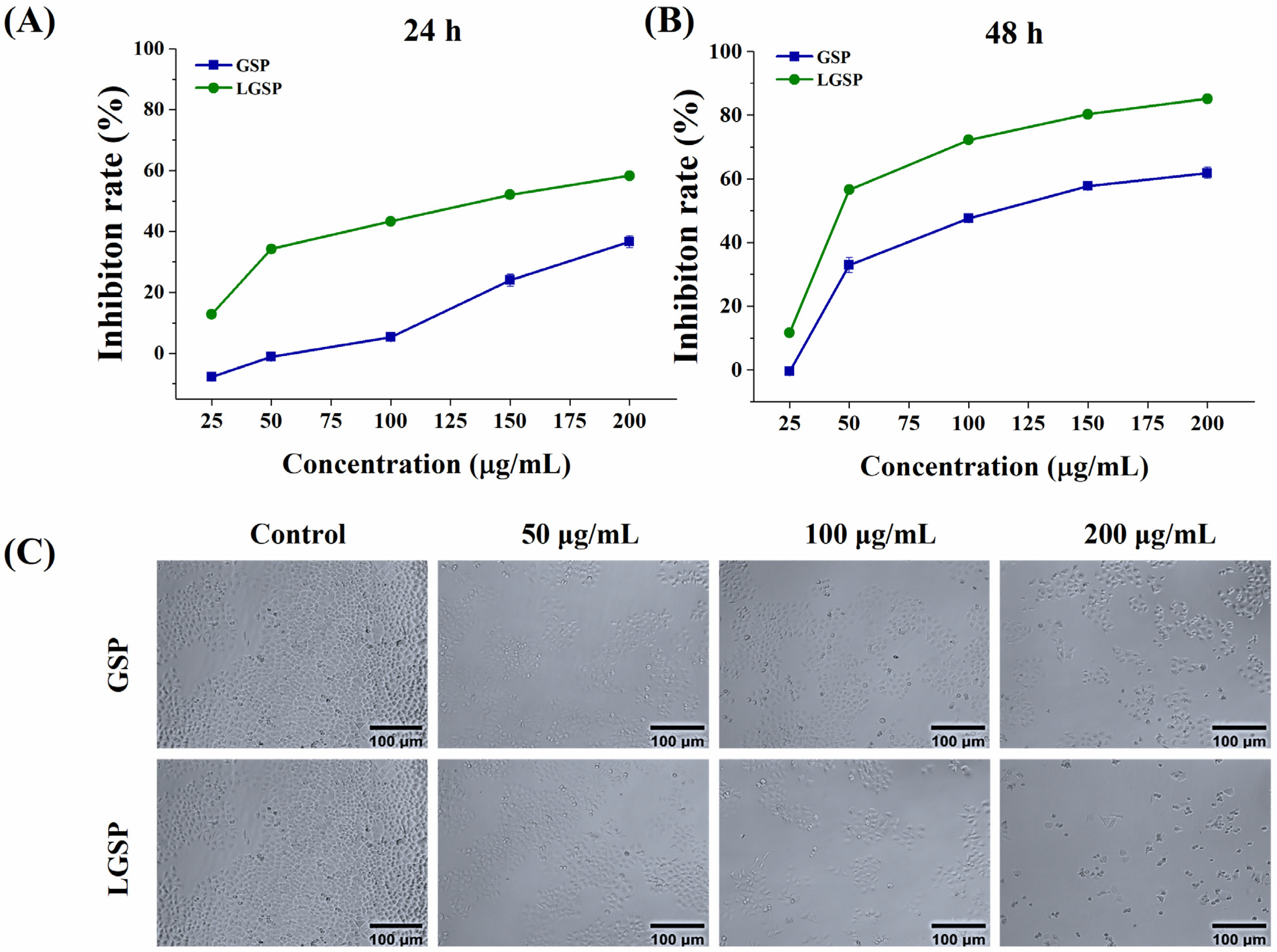
3.1. Effect of LGSP on the Proliferation of HeLa Cells
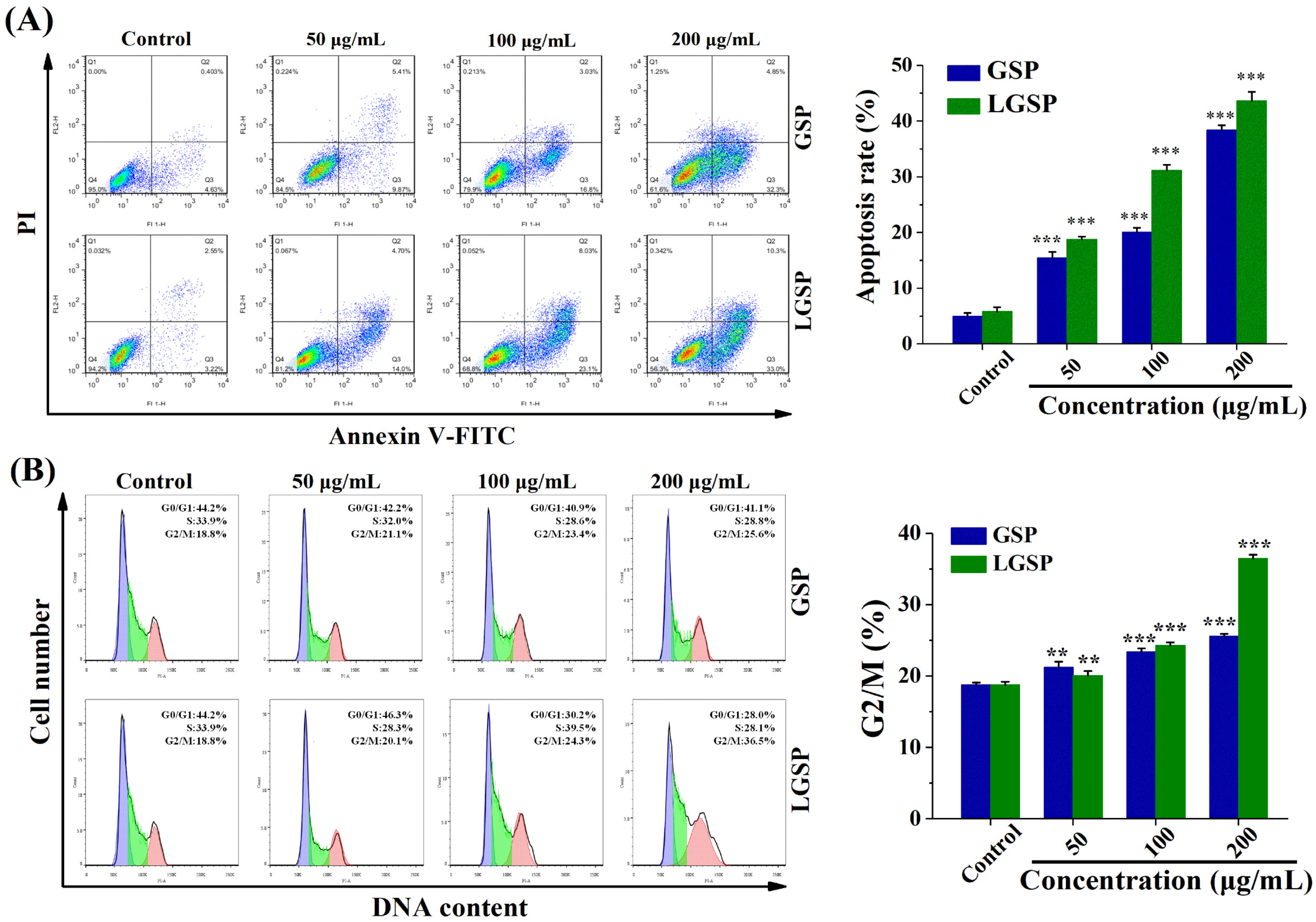
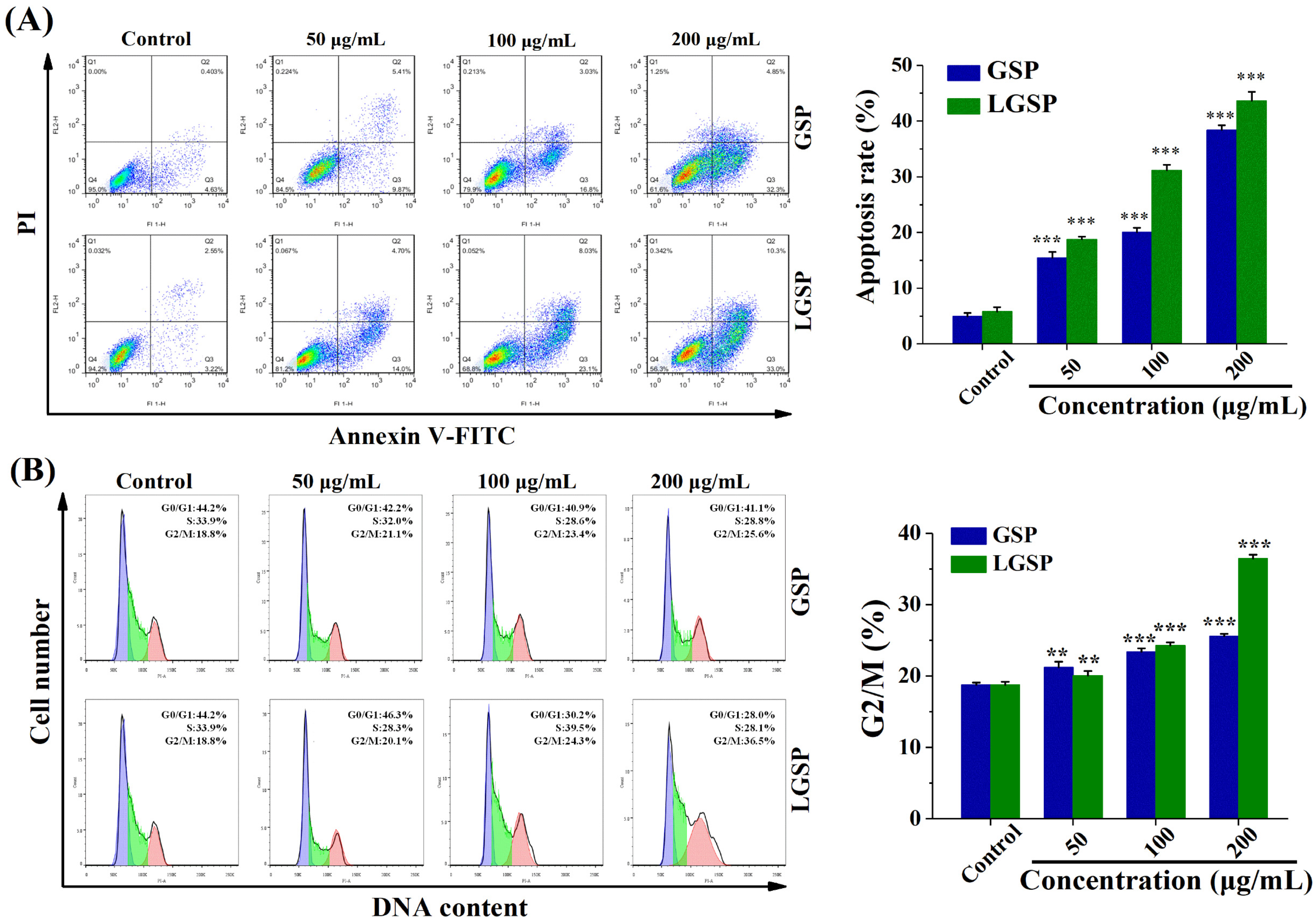
3.2. LGSP-Induced HeLa Cells Apoptosis
3.3. Effect of LGSP on the Cell Cycle of HeLa Cells
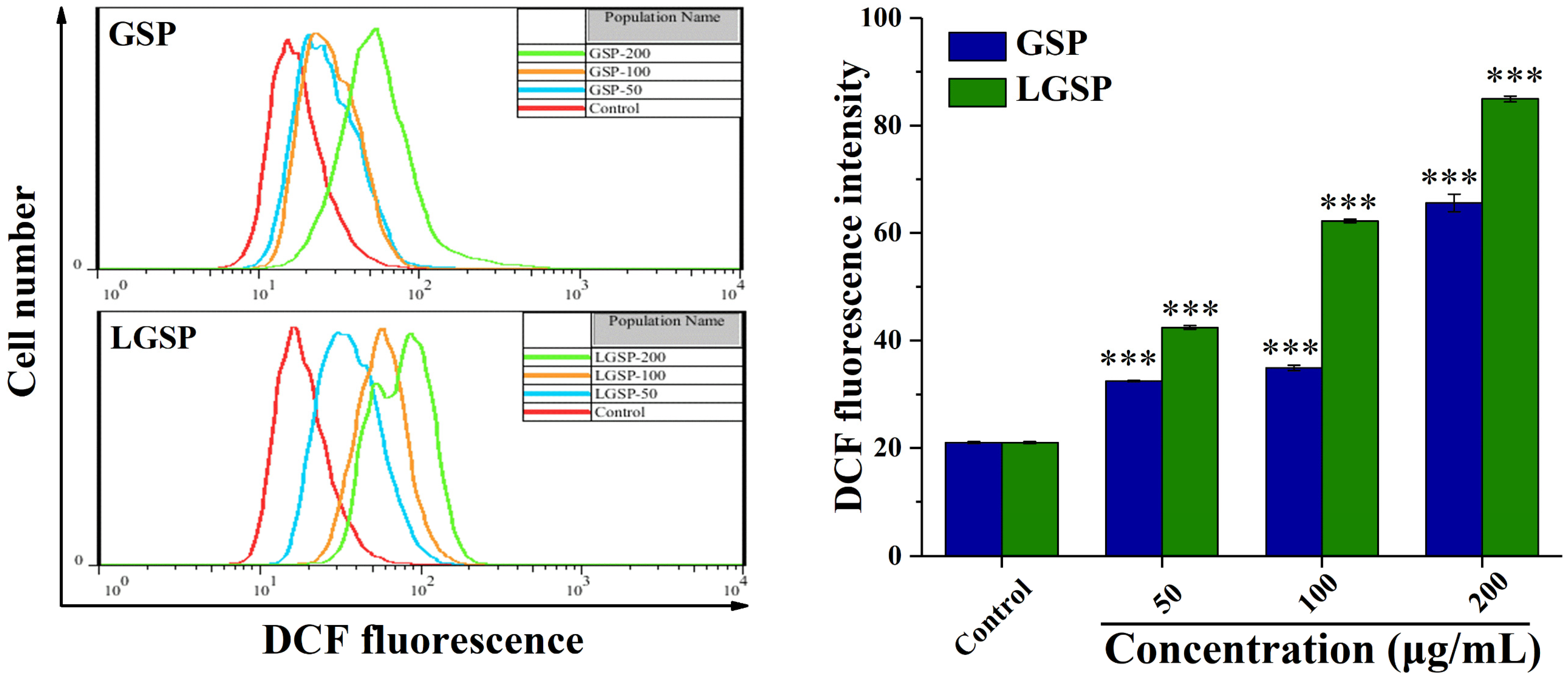
3.4. LGSP Enhanced the ROS Levels in HeLa Cells
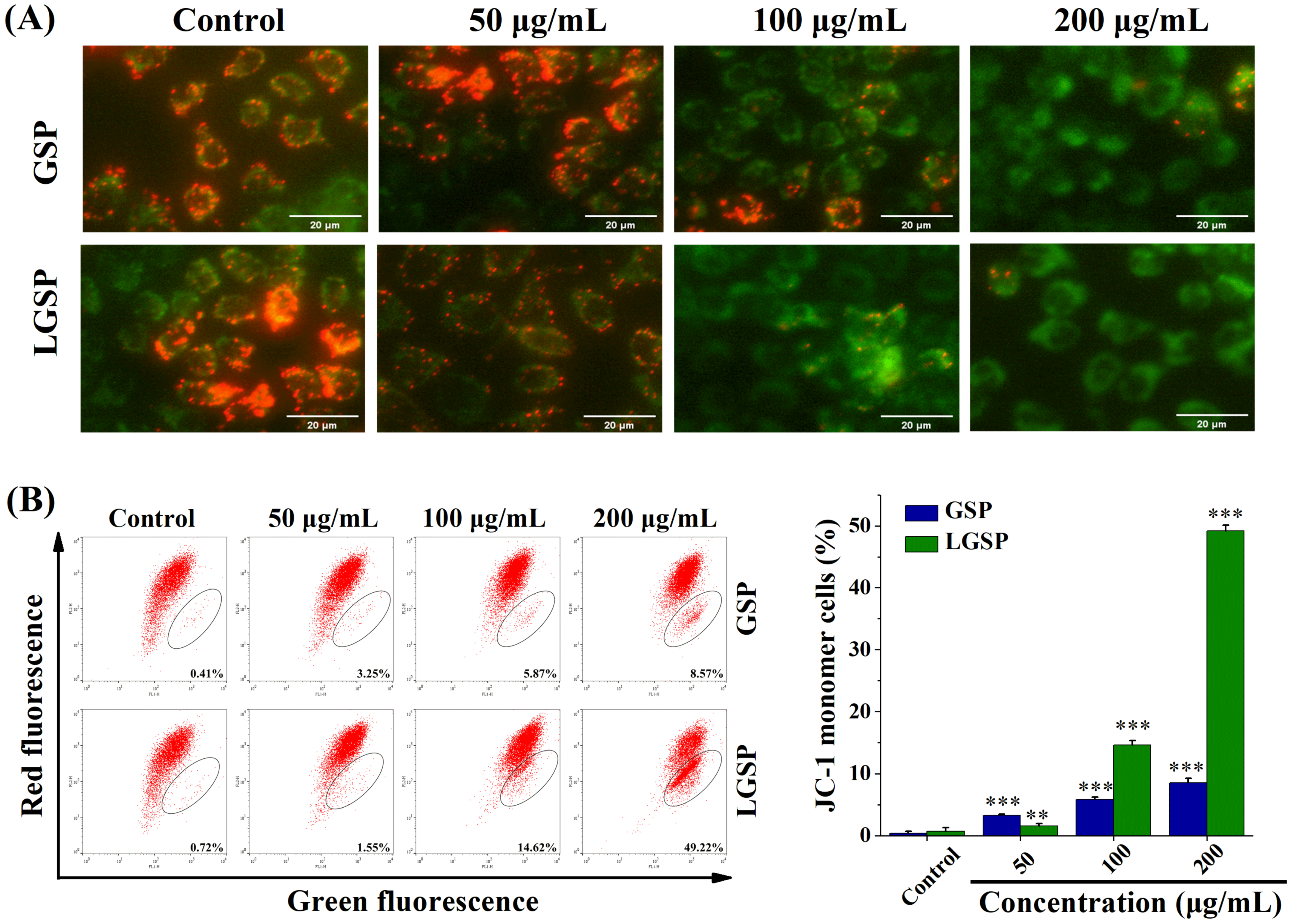
3.5. LGSP Induced Mitochondrial Dysfunction in HeLa Cells
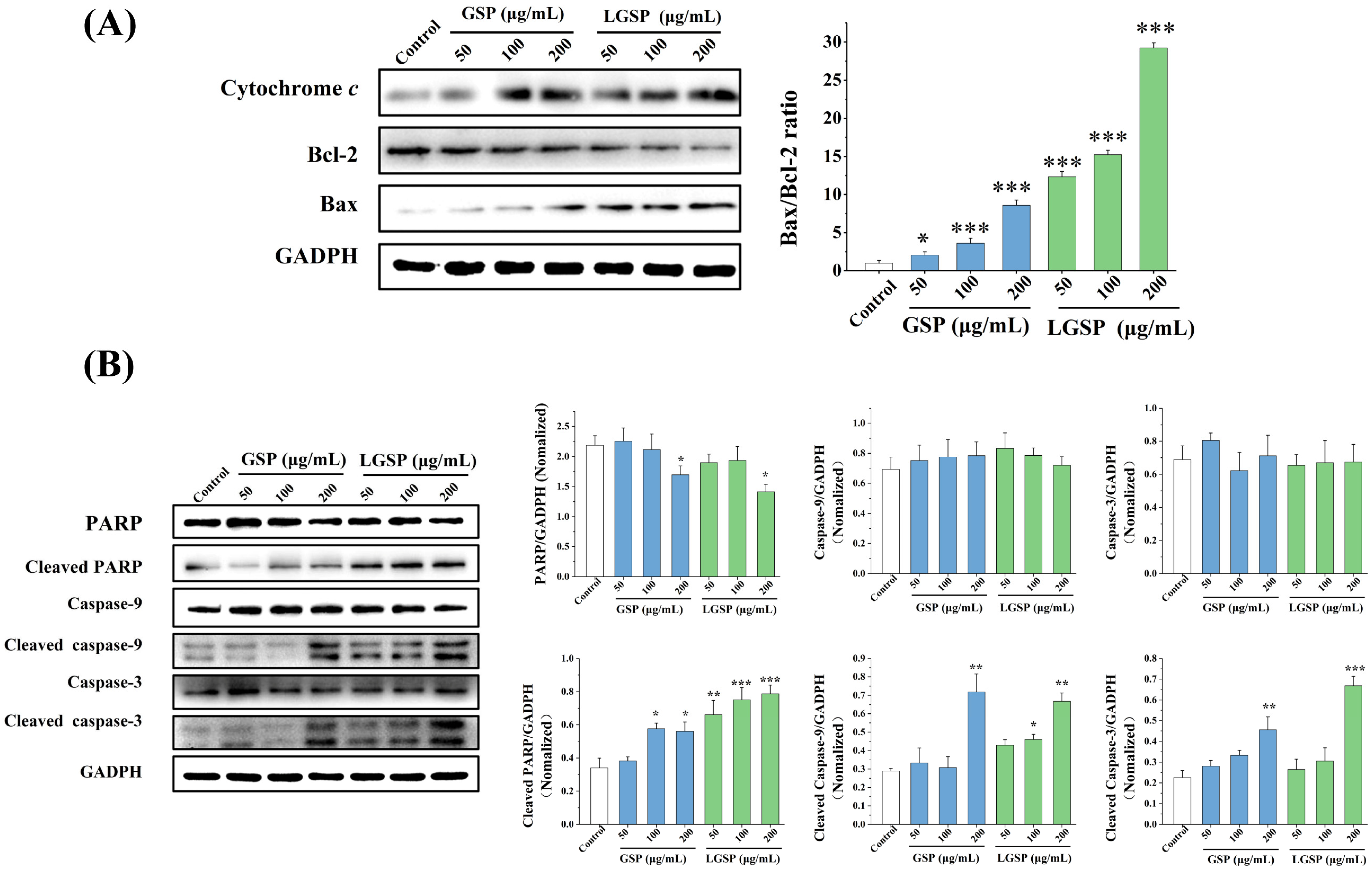
3.6. Effect of LGSP on Bax, Bcl-2 and Cytochrome c Expression in HeLa Cells
3.7. LGSP-Induced Activation of Caspases and the Cleavage of PARP Protein in HeLa Cells
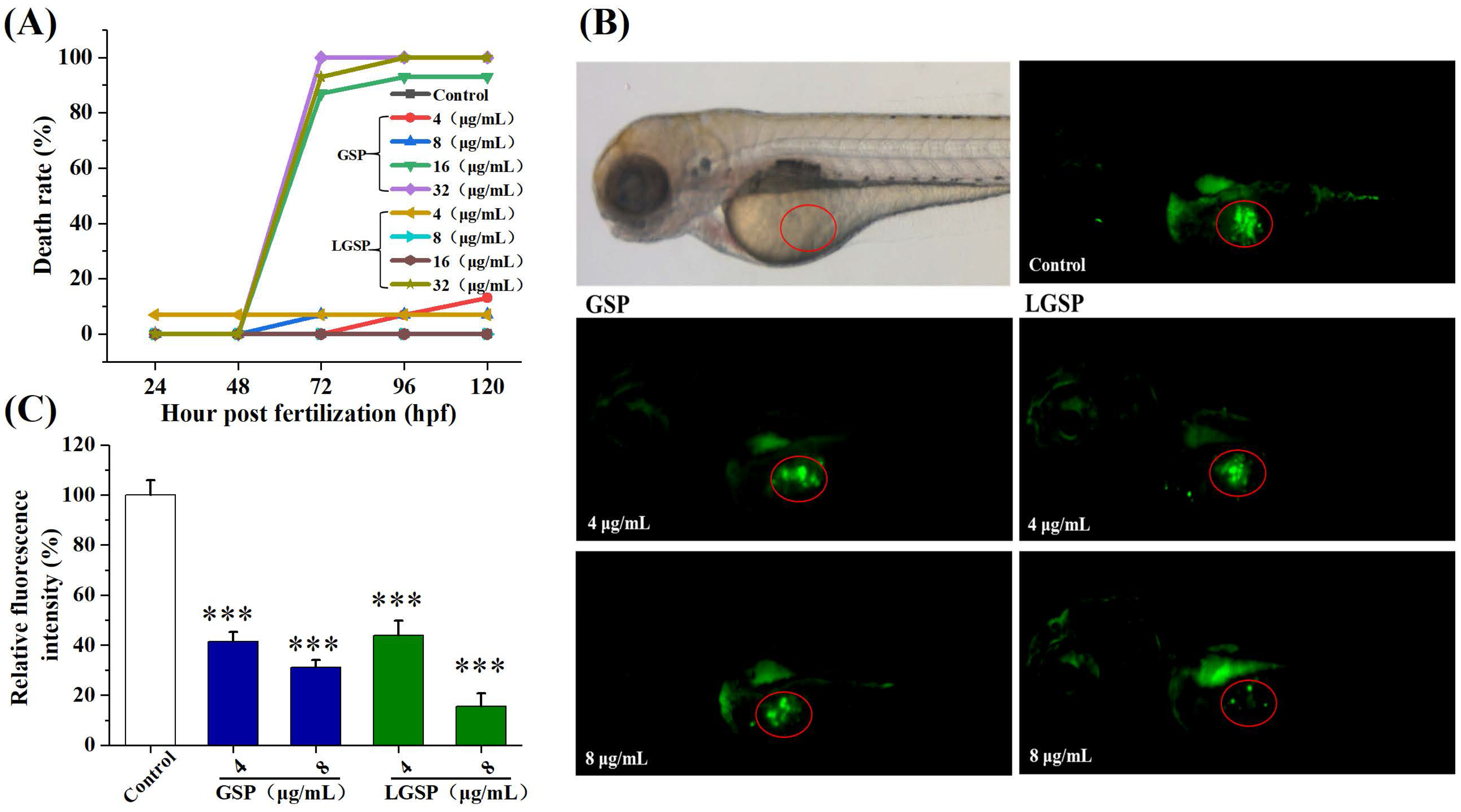
3.8. LGSP Blocked Cervical Cancer Growth in a HeLa-Derived Xenograft Zebrafish Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjose, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of incidence and mortality of cervical cancer in 2018: A worldwide analysis. Lancet Glob. Health 2020, 8, E191–E203. [Google Scholar] [CrossRef] [Green Version]
- Small, W., Jr.; Bacon, M.A.; Bajaj, A.; Chuang, L.T.; Fisher, B.J.; Harkenrider, M.M.; Jhingran, A.; Kitchener, H.C.; Mileshkin, L.R.; Viswanathan, A.N.; et al. Cervical Cancer: A Global Health Crisis. Cancer 2017, 123, 2404–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothwell, J.A.; Knaze, V.; Zamora-Ros, R. Polyphenols: Dietary assessment and role in the prevention of cancers. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Yeo, J. Bioactivities of Phenolics by Focusing on Suppression of Chronic Diseases: A Review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roleira, F.M.F.; Tavares-da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant derived and dietary phenolic antioxidants: Anticancer properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef]
- Unusan, N. Proanthocyanidins in grape seeds: An updated review of their health benefits and potential uses in the food industry. J. Funct. Foods 2020, 67, 103861. [Google Scholar] [CrossRef]
- Long, M.; Yang, S.H.; Han, J.X.; Li, P.; Zhang, Y.; Dong, S.; Chen, X.L.; Guo, J.Y.; Wang, J.; He, J.B. The Protective Effect of Grape-Seed Proanthocyanidin Extract on Oxidative Damage Induced by Zearalenone in Kunming Mice Liver. Int. J. Mol. Sci. 2016, 17, 808. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.X.; Bao, L.; Ren, J.W.; Li, Y.; Zhang, Z.F. Grape seed procyanidin B2 protects podocytes from high glucose-induced mitochondrial dysfunction and apoptosis via the AMPK-SIRT1-PGC-1 alpha axis in vitro. Food Funct. 2016, 7, 805–815. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, S.Q.; Wang, J.Q.; Shi, J.; Sun, Y.K.; Wang, W.Q.; Ning, G.; Hong, J.; Liu, R.X. Grape seed proanthocyanidin extract ameliorates inflammation and adiposity by modulating gut microbiota in high-fat diet mice. Mol. Nutr. Food Res. 2017, 61, 1601082. [Google Scholar] [CrossRef]
- Huang, L.L.; Pan, C.; Wang, L.; Ding, L.; Guo, K.; Wang, H.Z.; Xu, A.m.; Gao, S. Protective effects of grape seed proanthocyanidins on cardiovascular remodeling in DOCA-salt hypertension rats. J. Nutr. Biochem. 2015, 26, 841–849. [Google Scholar] [CrossRef]
- Zhu, S.; Li, Y.; Ma, C.Y.; Lou, Z.X.; Chen, S.W.; Dai, J.; Wang, H.X. Optimization of lipase-catalyzed synthesis of acetylated EGCG by response surface methodology. J. Mol. Catal. B Enzym. 2013, 97, 87–94. [Google Scholar] [CrossRef]
- Mellou, F.; Loutrari, H.; Stamatis, H.; Roussos, C.; Kolisis, F.N. Enzymatic esterification of flavonoids with unsaturated fatty acids: Effect of the novel esters on vascular endothelial growth factor release from K562 cells. Process Biochem. 2006, 41, 2029–2034. [Google Scholar] [CrossRef]
- Chen, M.; Yu, S. Lipophilic grape seed proanthocyanidin exerts anti-proliferative and pro-apoptotic effects on PC3 Human Prostate cancer cells and suppresses PC3 xenograft tumor growth in vivo. J. Agric. Food Chem. 2019, 67, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.Z.; Hu, X.F.; Pan, L.H.; Zheng, Z.; Zhao, Y.Y.; Cao, L.L.; Pang, M.; Hou, Z.G.; Jiang, S.T. Preparation of camellia oil-based W/O emulsions stabilized by tea polyphenol palmitate: Structuring camellia oil as a potential solid fat replacer. Food Chem. 2019, 276, 209–217. [Google Scholar] [CrossRef]
- Chen, M.S.; Yu, S.J. Lipophilized grape seed proanthocyanidin derivatives as novel antioxidants. J. Agric. Food Chem. 2017, 65, 1598–1605. [Google Scholar] [CrossRef]
- Zhang, L.L.; Chen, J.; Liao, H.J.; Li, C.H.; Chen, M.S. Anti-inflammatory effect of lipophilic grape seed proanthocyanidin in RAW 264.7 cells and a zebrafish model. J. Funct. Foods 2020, 75, 104217. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.T.; Yang, L.; Jiang, J.G. Effects of thonningianin A in natural foods on apoptosis and cell cycle arrest of HepG-2 human hepatocellular carcinoma cells. Food Funct. 2015, 6, 2588–2597. [Google Scholar] [CrossRef]
- Wang, J.; Liao, A.M.; Thakur, K.; Zhang, J.G.; Huang, J.H.; Wei, Z.J. Licochalcone B extracted from glycyrrhiza uralensis fisch induces apoptotic effects in human hepatoma cell HepG2. J. Agric. Food Chem. 2019, 67, 3341–3353. [Google Scholar] [CrossRef]
- Wu, Q.; Deng, J.J.; Fan, D.D.; Duan, Z.G.; Zhu, C.H.; Fu, R.Z.; Wang, S.S. Ginsenoside Rh4 induces apoptosis and autophagic cell death through activation of the ROS/JNK/p53 pathway in colorectal cancer cells. Biochem. Pharmacol. 2018, 148, 64–74. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, L.L.; Li, C.H.; Chen, R.C.; Liu, C.M.; Chen, M.S. Lipophilized epigallocatechin gallate derivative exerts anti-proliferation efficacy through induction of cell cycle arrest and apoptosis on DU145 human prostate cancer cells. Nutrients 2020, 12, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, G.H.; Zou, L.F.; Deng, Y.Y.; Meng, Y.L.; Liao, X.J.; Lu, H.Q. Clethodim exposure induces developmental immunotoxicity and neurobehavioral dysfunction in zebrafish embryos. Fish Shellfish Immunol. 2019, 86, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Song, G.; Lee, J.-Y.; Hong, T.; Chang, M.-J.; Lim, W. Laminarin-Derived from brown algae suppresses the growth of Ovarian cancer cells via mitochondrial dysfunction and ER stress. Mar. Drugs 2020, 18, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, A.; Sethi, G. Role of reactive oxygen species in cancer progression: Molecular mechanisms and recent advancements. Biomolecules 2019, 9, 753. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.Y.; Zhang, Y.; Liu, Y.T.; Xu, W.P.; Yang, Y.; Hao, Y.W.; Tao, L.M. A natural product enhances apoptosis via mitochondria/caspase-mediated pathway in HeLa cells. J. Cell. Biochem. 2019, 120, 16811–16823. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, X.F.; Zheng, P.S. Grape seed proanthocyanidins (GSPs) inhibit the growth of cervical cancer by inducing apoptosis mediated by the mitochondrial pathway. PLoS ONE. 2014, 9, e107045. [Google Scholar]
- Sudan, S.; Rupasinghe, H.P.V. Antiproliferative activity of long chain acylated esters of quercetin-3-O-glucoside in hepatocellular carcinoma HepG2 cells. Exp. Biol. Med. 2015, 240, 1452–1464. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Liao, W.; Wang, J.; Wang, P.; Gao, H.; Wang, M.; Xu, C.; Zhong, Y.; Ding, Y. CSTMP induces apoptosis and mitochondrial dysfunction in human myeloma RPMI8226 cells via CHOP-dependent endoplasmic reticulum stress. Biomed. Pharmacother. 2016, 83, 776–784. [Google Scholar] [CrossRef]
- Ye, X.C.; Zuo, D.D.; Yu, L.; Zhang, L.; Tang, J.; Cui, C.C.; Bao, L.; Zan, K.; Zhang, Z.H.; Yang, X.X.; et al. ROS/TXNIP pathway contributes to thrombin induced NLRP3 inflammasome activation and cell apoptosis in microglia. Biochem. Biophys. Res. Commun. 2017, 485, 499–505. [Google Scholar] [CrossRef]
- Wu, G.S.; Lu, J.J.; Guo, J.J.; Li, Y.B.; Tan, W.; Dang, Y.Y.; Zhong, Z.F.; Xu, Z.T.; Chen, X.P.; Wang, Y.T. Ganoderic acid DM, a natural triterpenoid, induces DNA damage, G1 cell cycle arrest and apoptosis in human breast cancer cells. Fitoterapia 2012, 83, 408–414. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Shi, L.F.; Song, M.Y.; Meng, Y. Antitumor activity of paederosidic acid in human non-small cell lung cancer cells via inducing mitochondria-mediated apoptosis. Chem. Biol. Interact. 2017, 269, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Ben Othmene, Y.; Monceaux, K.; Karoui, A.; Ben Salem, I.; Belhadef, A.; Abid-Essefi, S.; Lemaire, C. Tebuconazole induces ROS-dependent cardiac cell toxicity by activating DNA damage and mitochondrial apoptotic pathway. Ecotoxicol. Environ. Saf. 2020, 204, 111040. [Google Scholar] [CrossRef]
- McArthur, K.; Whitehead, L.W.; Heddleston, J.M.; Li, L.; Padman, B.S.; Oorschot, V.; Geoghegan, N.D.; Chappaz, S.; Davidson, S.; Chin, H.S.; et al. BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science. 2018, 359, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Bi, Y.L.; Min, M.; Shen, W.; Liu, Y. Genistein induced anticancer effects on pancreatic cancer cell lines involves mitochondrial apoptosis, G(0)/G(1) cell cycle arrest and regulation of STAT3 signaling pathway. Phytomedicine 2018, 39, 10–16. [Google Scholar] [CrossRef]
- Zhang, B.B.; Xuan, C.; Ji, Y.X.; Zhang, W.M.; Wang, D.G. Zebrafish xenotransplantation as a tool for in vivo cancer study. Fam. Cancer 2015, 14, 487–493. [Google Scholar] [CrossRef]
- Jin, W.D.; Zhou, L.; Yan, B.; Yan, L.; Liu, F.C.; Tong, P.J.; Yu, W.H.; Dong, X.Q.; Xie, L.; Zhang, J.; et al. Theabrownin triggers DNA damage to suppress human osteosarcoma U2OS cells by activating p53 signaling pathway. J. Cell. Mol. Med. 2018, 22, 4423–4436. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Zhang, L.; Liu, C.; He, X.; Chen, M.; Chen, J. Lipophilic Grape Seed Proanthocyanidin Exerts Anti-Cervical Cancer Effects in HeLa Cells and a HeLa-Derived Xenograft Zebrafish Model. Antioxidants 2022, 11, 422. https://doi.org/10.3390/antiox11020422
Li C, Zhang L, Liu C, He X, Chen M, Chen J. Lipophilic Grape Seed Proanthocyanidin Exerts Anti-Cervical Cancer Effects in HeLa Cells and a HeLa-Derived Xenograft Zebrafish Model. Antioxidants. 2022; 11(2):422. https://doi.org/10.3390/antiox11020422
Chicago/Turabian StyleLi, Changhong, Linli Zhang, Chengmei Liu, Xuemei He, Mingshun Chen, and Jun Chen. 2022. "Lipophilic Grape Seed Proanthocyanidin Exerts Anti-Cervical Cancer Effects in HeLa Cells and a HeLa-Derived Xenograft Zebrafish Model" Antioxidants 11, no. 2: 422. https://doi.org/10.3390/antiox11020422
APA StyleLi, C., Zhang, L., Liu, C., He, X., Chen, M., & Chen, J. (2022). Lipophilic Grape Seed Proanthocyanidin Exerts Anti-Cervical Cancer Effects in HeLa Cells and a HeLa-Derived Xenograft Zebrafish Model. Antioxidants, 11(2), 422. https://doi.org/10.3390/antiox11020422