
Synergistic Anti-Inflammatory Effects of Lipophilic Grape Seed Proanthocyanidin and Camellia Oil Combination in LPS-Stimulated RAW264.7 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of LGSP
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. NO Production Measurement
2.6. Synergistic Effect Analysis
2.7. Quantitative Real-Time PCR Analysis
2.8. Measurement of Reactive Oxygen Species (ROS) Production
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results and Discussion
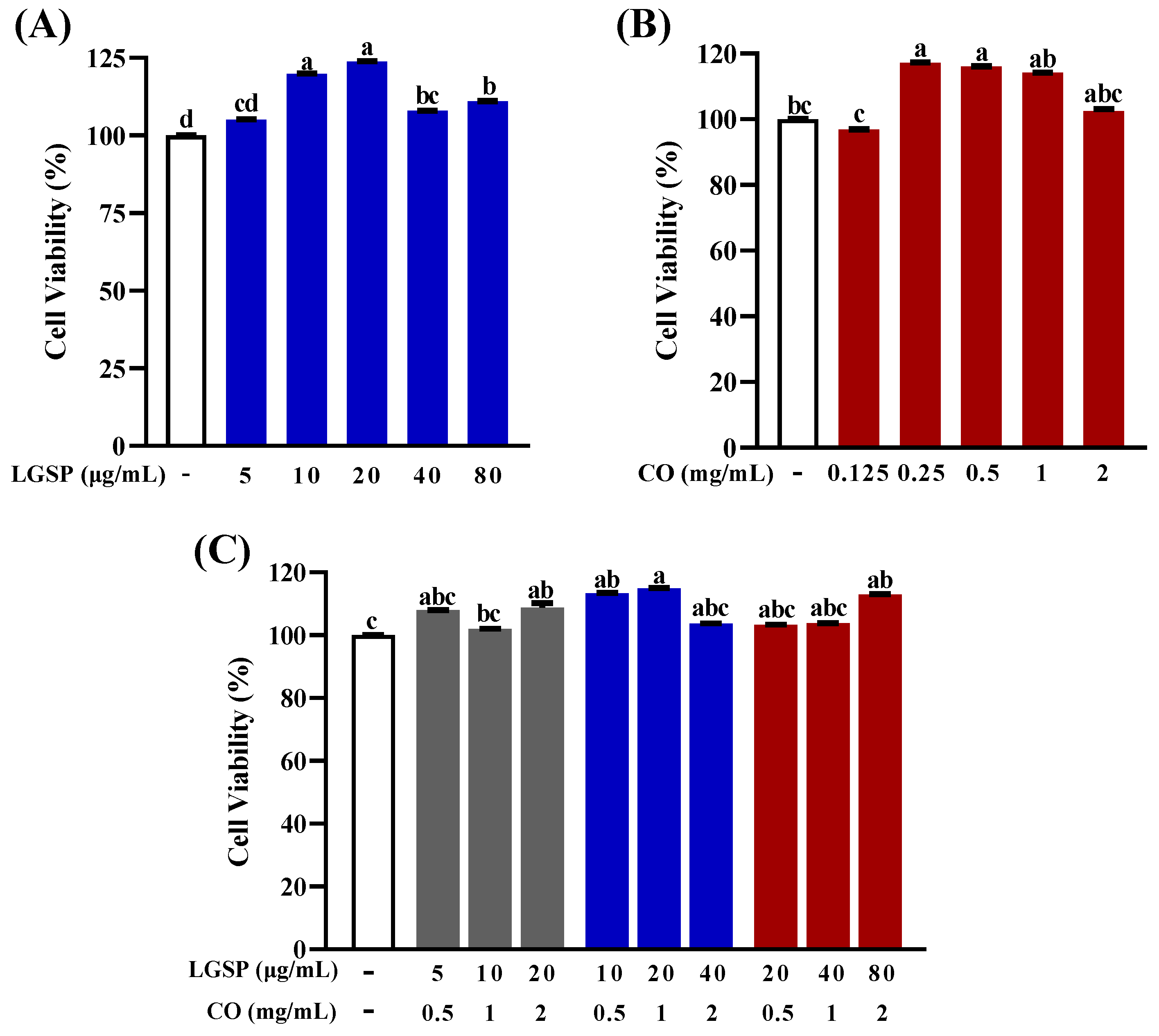
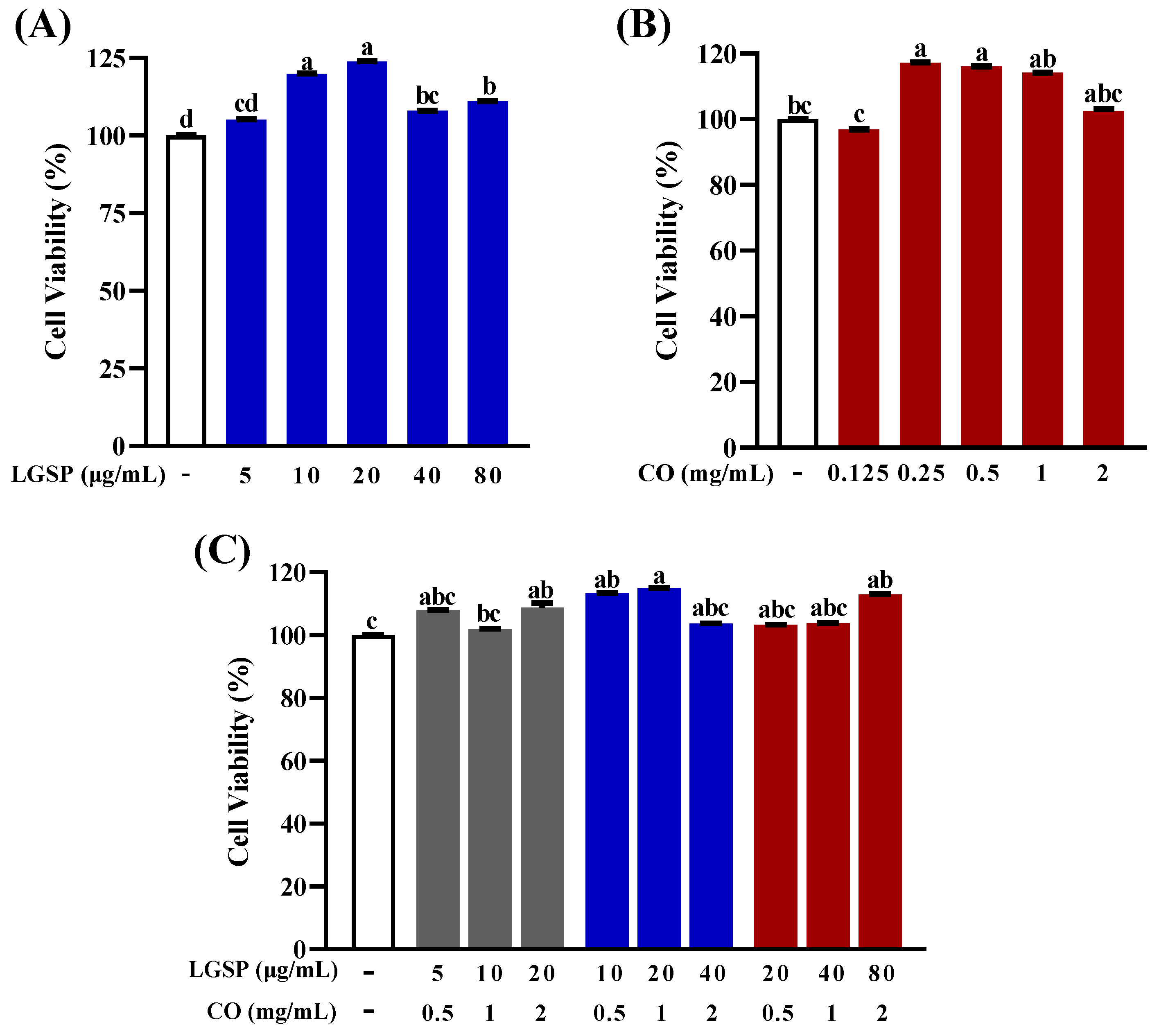
3.1. Effect of LGSP and CO on Cell Viability of RAW264.7 Cells
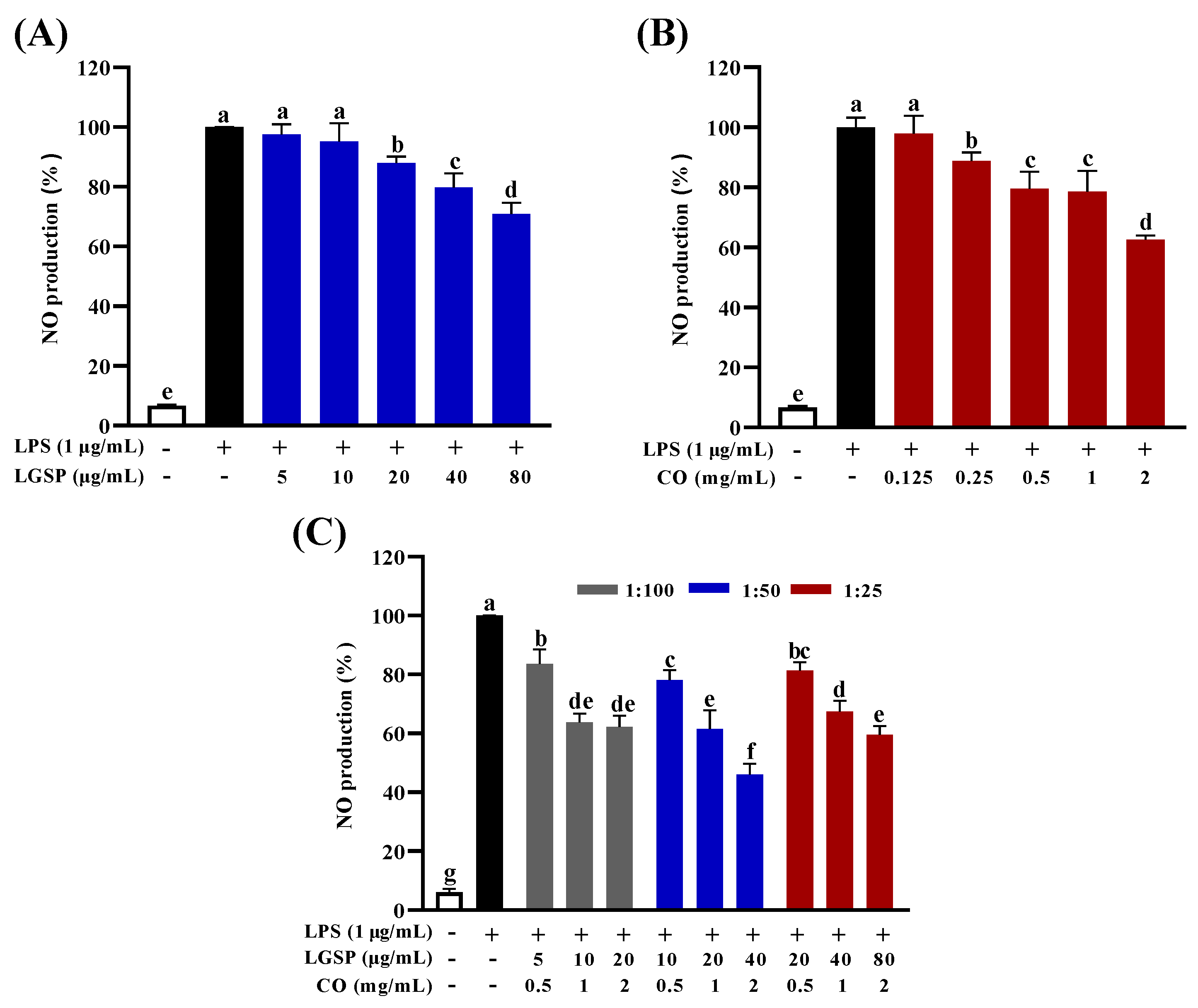
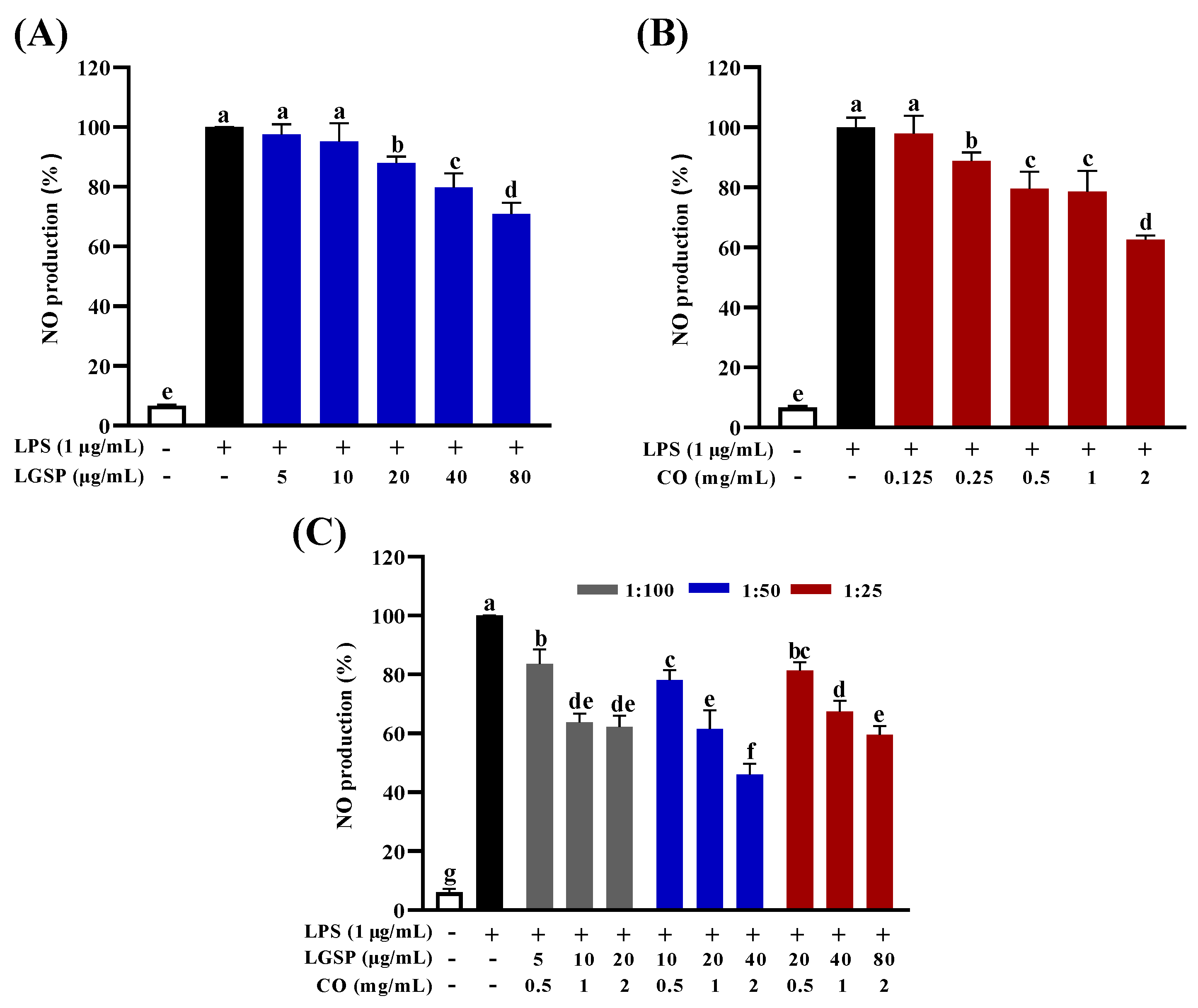
3.2. Effect of LGSP and CO on Inhibition of NO Production
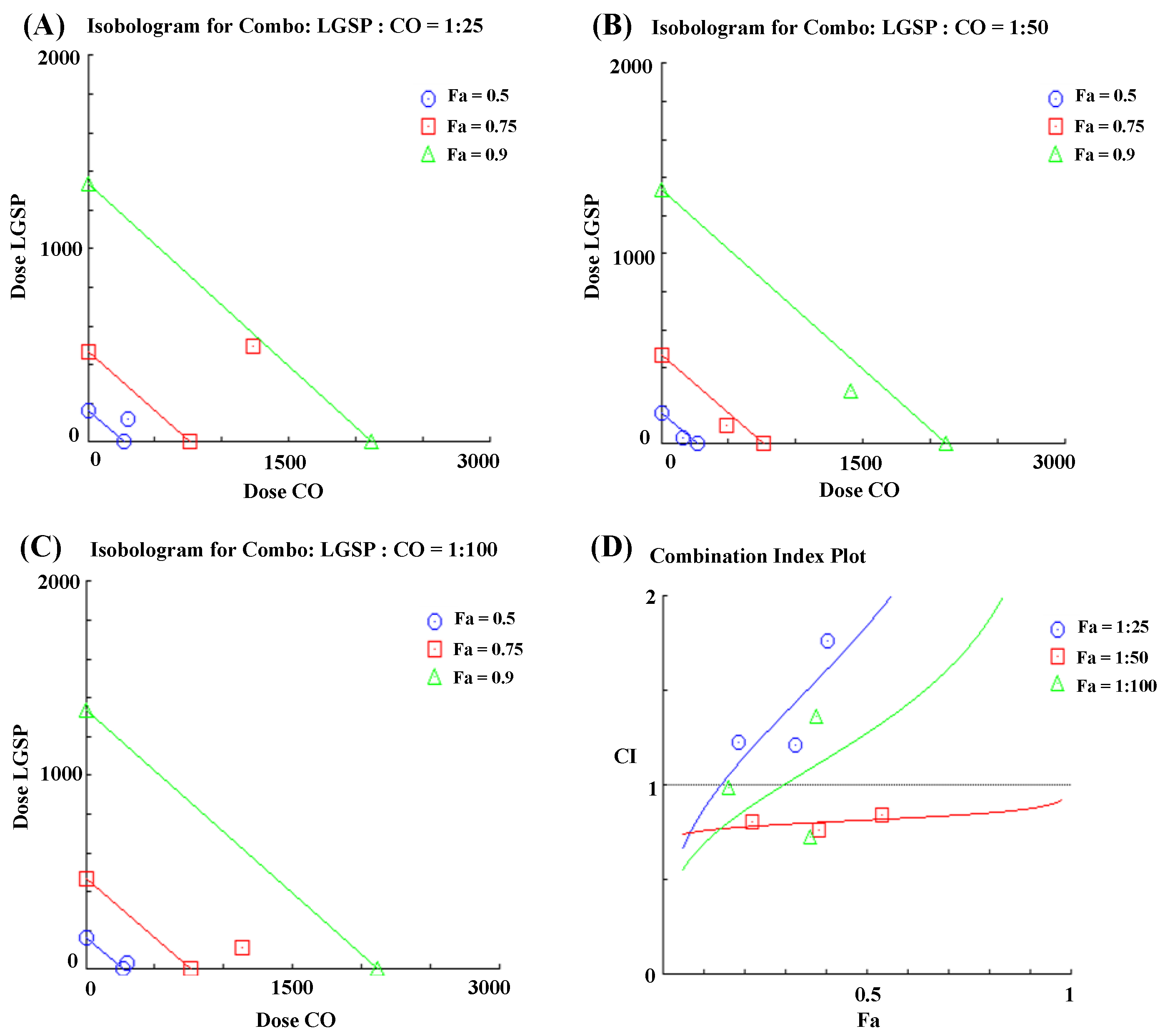
3.3. Synergistic Effect Analysis
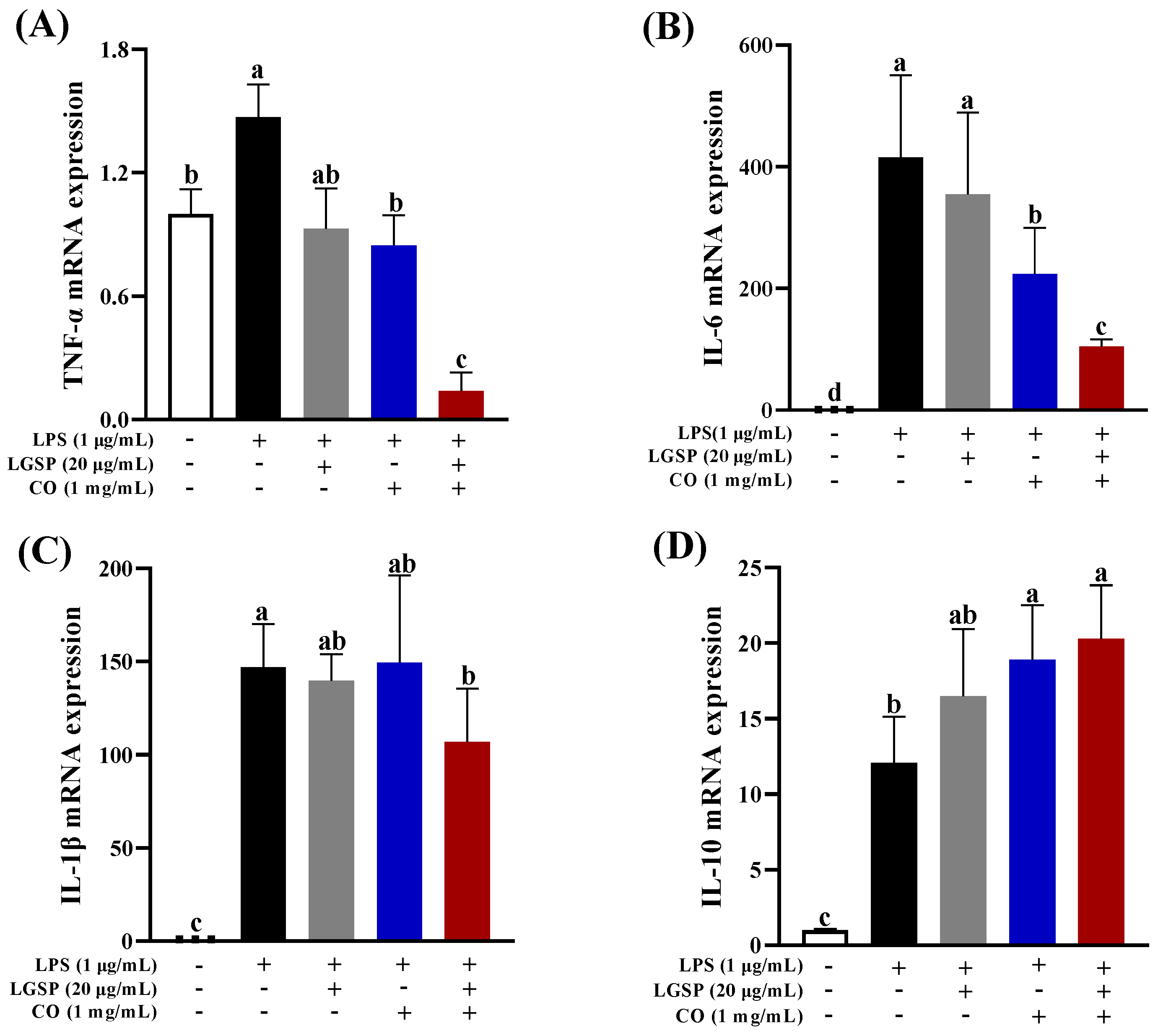
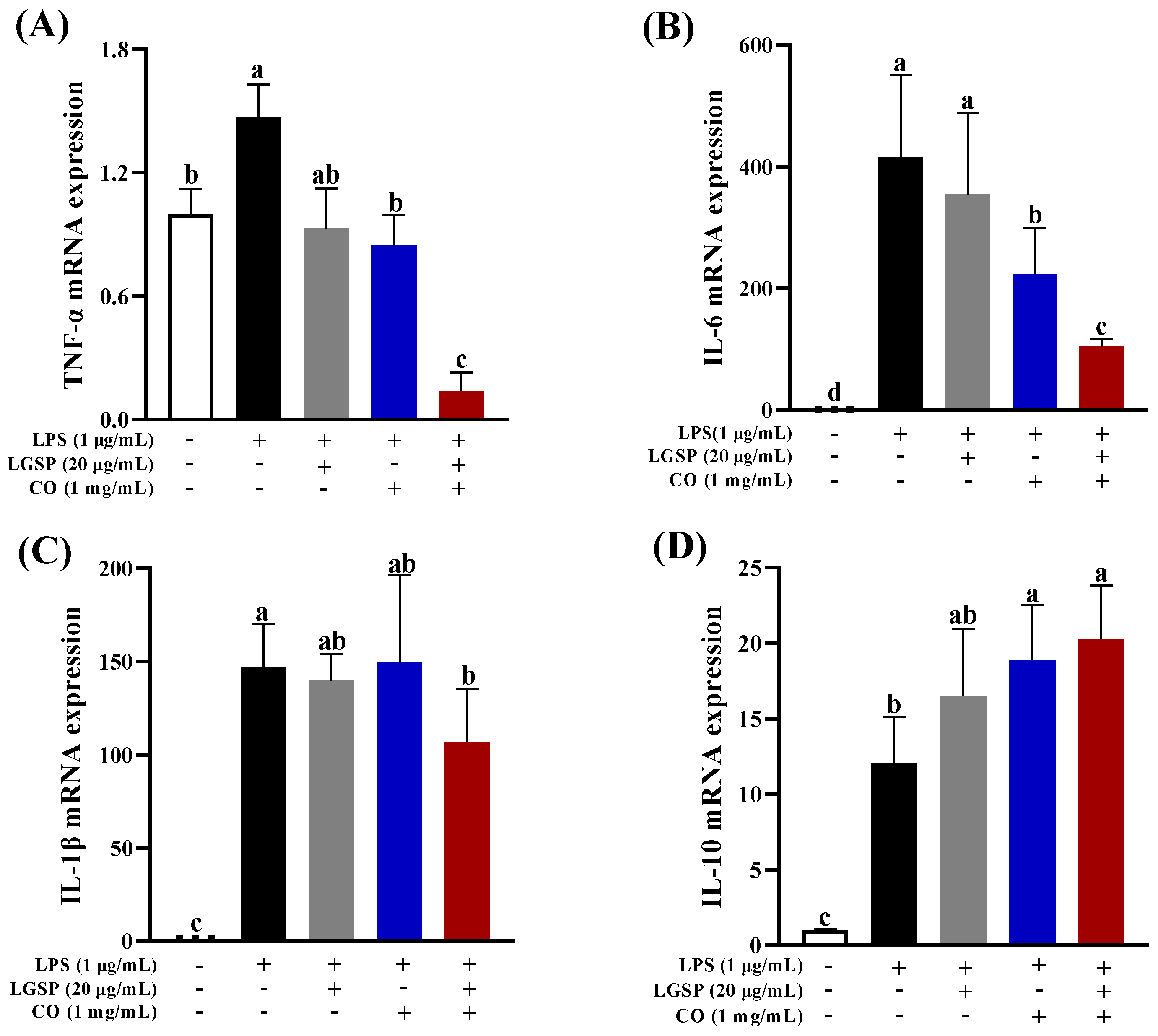
3.4. Effect of LGSP and CO on the mRNA Expression of Inflammatory Cytokines
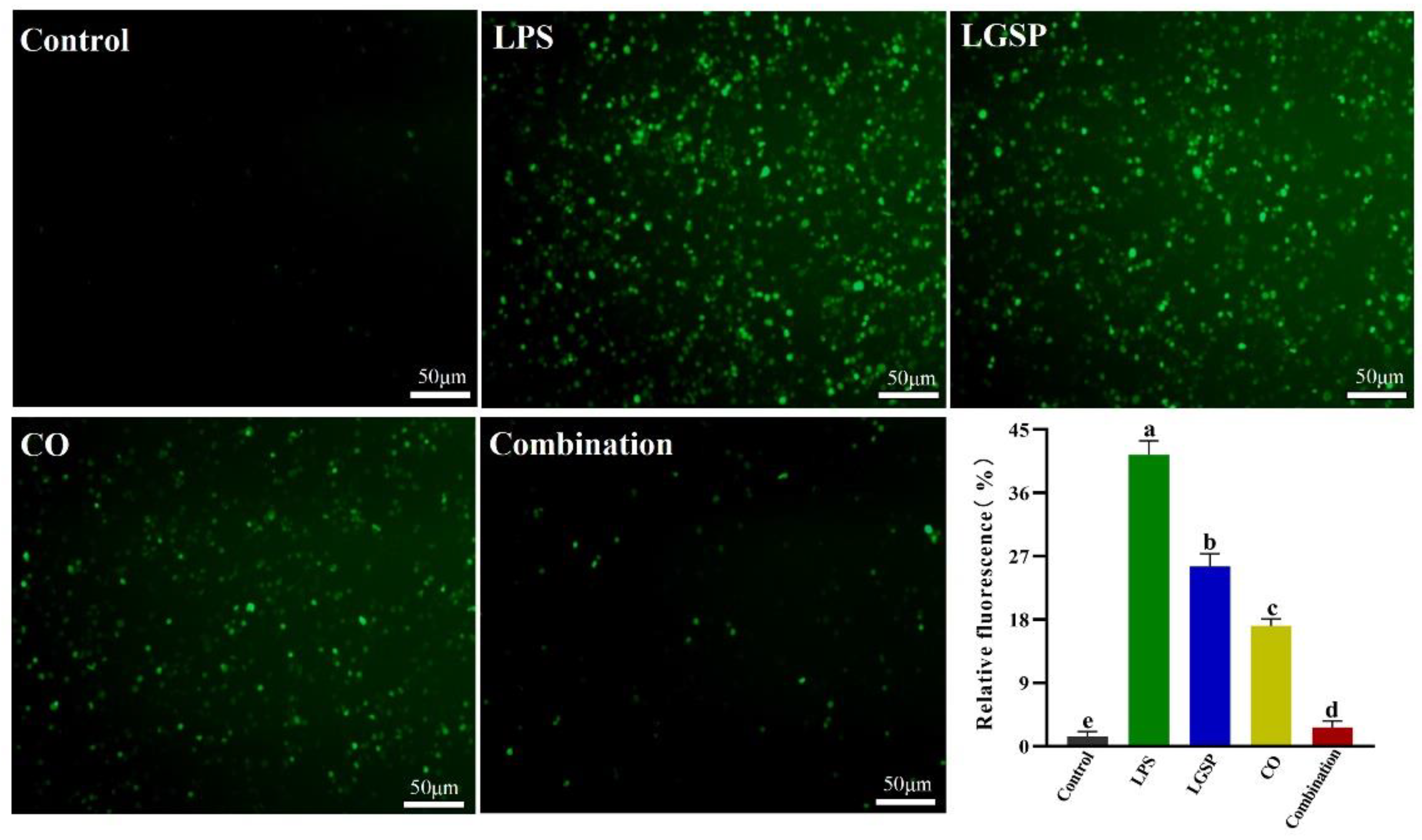
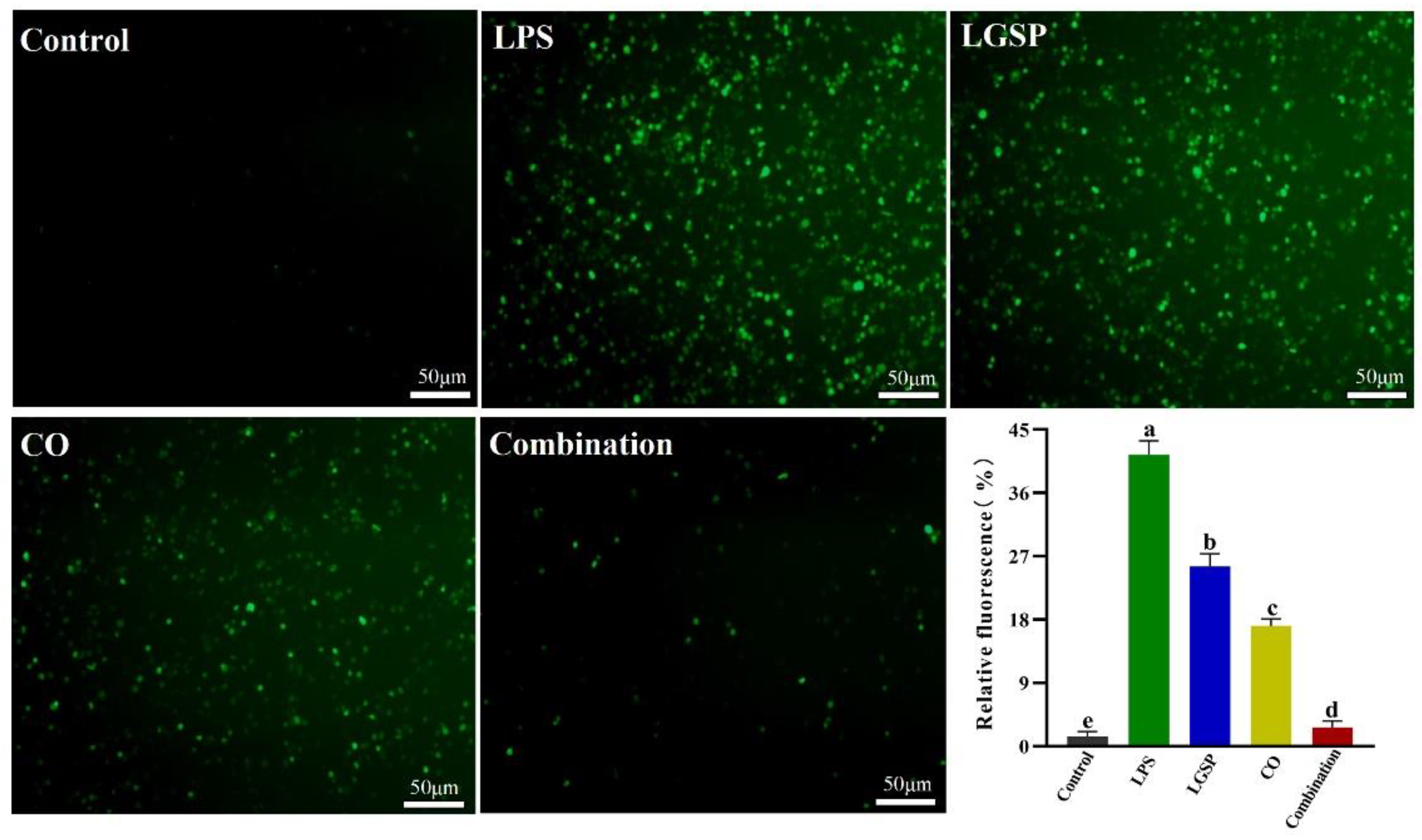
3.5. Effect of LGSP and CO on LPS-Induced ROS
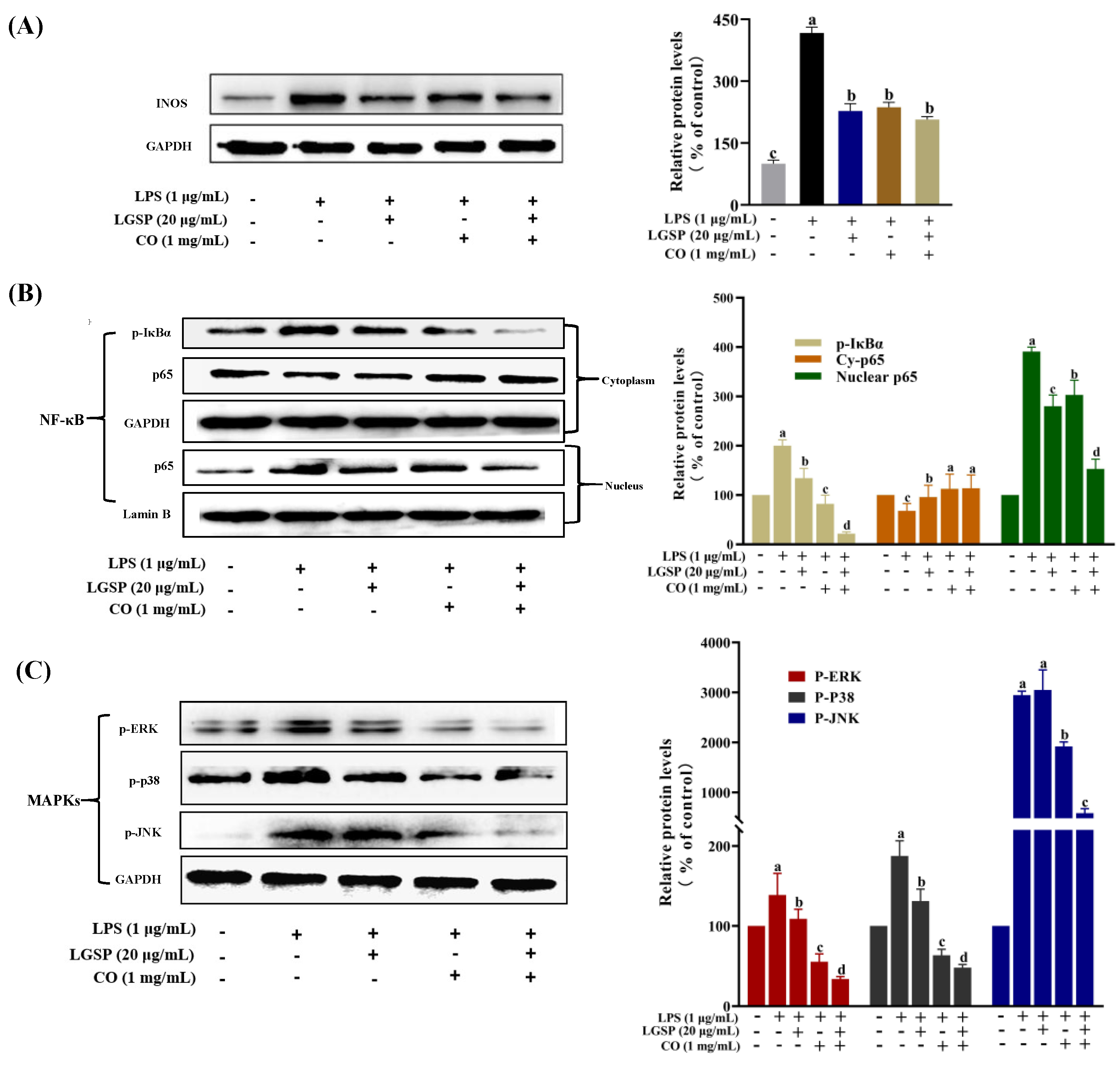
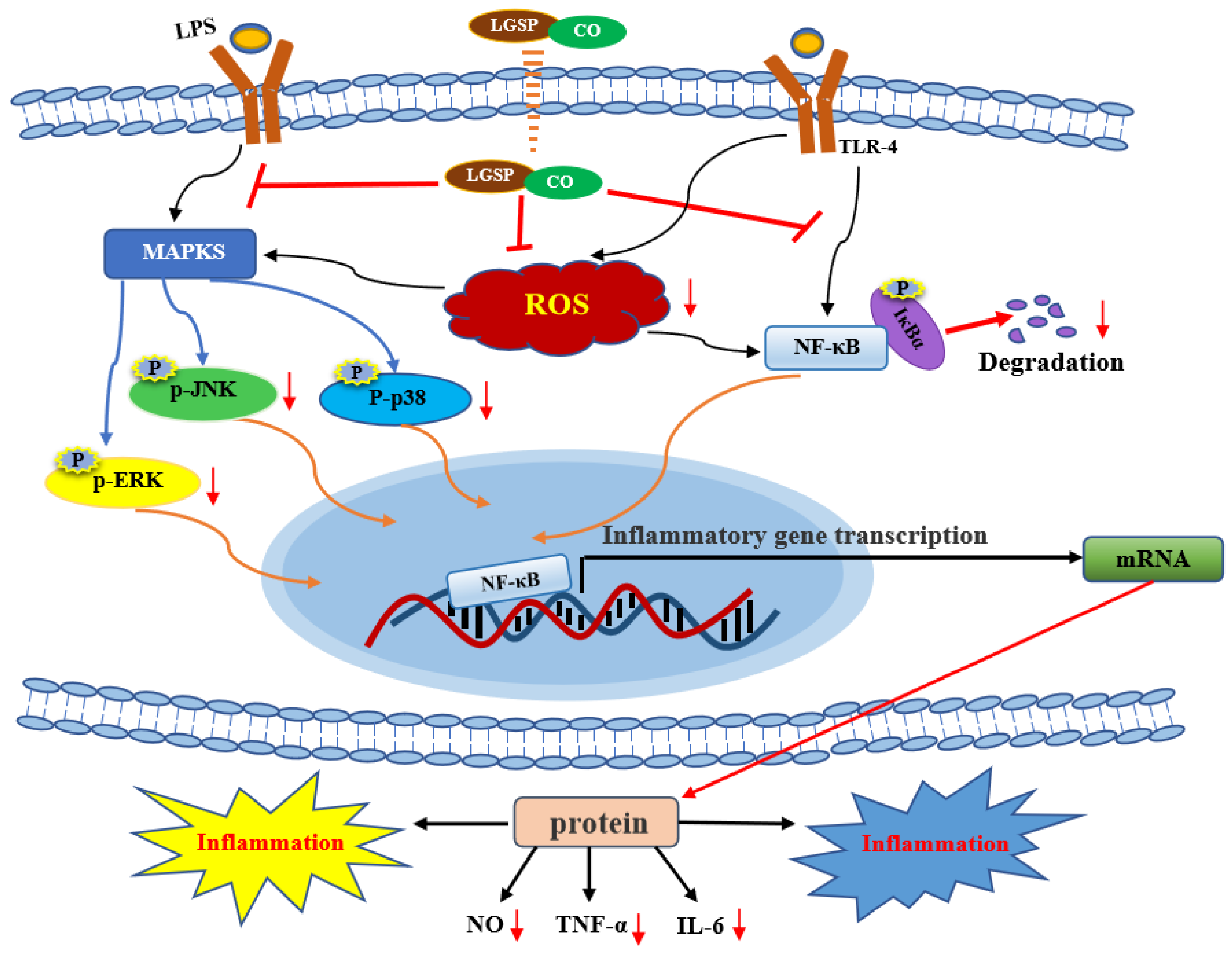
3.6. Effect of LGSP and CO on LPS-Induced iNOS, NF-κB and MAPK Pathway
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ono, M. Molecular links between tumor angiogenesis and inflammation: Inflammatory stimuli of macrophages and cancer cells as targets for therapeutic strategy. Cancer Sci. 2008, 99, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Du, L. Diabetes is an inflammatory disease: Evidence from traditional chinese medicines. Diabetes Obes. Metab. 2011, 13, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Chae, U.; Kim, H.-S.; Kim, K.-M.; Lee, H.; Lee, H.-S.; Park, J.-W.; Lee, D.-S. IDH2 deficiency in microglia decreases the pro-inflammatory response via the ERK and NF-κB pathways. Inflammation 2018, 41, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Ip, W.K.E.; Hoshi, N.; Shouval, D.S.; Snapper, S.; Medzhitov, R. Anti-inflammatory effect of IL-10 mediated by metabolic reprogramming of macrophages. Science 2017, 356, 513–519. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K. Stress, food, and inflammation: Psychoneuroimmunology and nutrition at the cutting edge. Psychosom. Med. 2010, 72, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Lee, W.-H.; Choi, E.-J.; Choi, J.-P.; Heo, Y.-J.; Gho, Y.-S.; Jee, Y.-K.; Oh, Y.-M.; Kim, Y.-K. Extracellular vesicles derived from gram-negative bacteria, such as Escherichia coli, induce emphysema mainly via IL-17A-mediated neutrophilic inflammation. J. Immunol. 2015, 194, 3361–3368. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.Y.; Lin, J.; Lu, K.; Xu, H.G.; Geng, Z.Z.; Sun, P.H.; Chen, W.M. Anti-inflammatory effects of cajaninstilbene acid and its derivatives. J. Agr. Food Chem. 2016, 64, 2893–2900. [Google Scholar] [CrossRef]
- Cardinal, S.; Azelmat, J.; Grenier, D.; Voyer, N. Anti-inflammatory properties of quebecol and its derivatives. Bioorg. Med. Chem. Lett. 2016, 26, 440–444. [Google Scholar] [CrossRef]
- Zhang, L.; Virgous, C.; Si, H. Synergistic anti-inflammatory effects and mechanisms of combined phytochemicals. J. Nutr. Biochem. 2019, 69, 19–30. [Google Scholar] [CrossRef]
- de Cassia da Silveira e Sa, R.; Andrade, L.N.; de Sousa, D.P. A review on anti-inflammatory activity of monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef]
- Kim, S.H.; Bang, J.; Son, C.N.; Baek, W.K.; Kim, J.M. Grape seed proanthocyanidin extract ameliorates murine autoimmune arthritis through regulation of TLR4/MyD88/NF-κB signaling pathway. Korean J. Intern. Med. 2018, 33, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yu, S. Lipophilized grape seed proanthocyanidin derivatives as novel antioxidants. J. Agr. Food Chem. 2017, 65, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yu, S. Lipophilic grape seed proanthocyanidin exerts anti-proliferative and pro-apoptotic effects on PC3 human prostate cancer cells and suppresses PC3 xenograft tumor growth in vivo. J. Agr. Food Chem. 2019, 67, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.T.; Howell, K.; Chan, M.; Zhang, P.; Ng, K. Modulation of the human gut microbiota by phenolics and phenolic fiber-rich foods. Compr. Rev. Food Sci. F. 2020, 19, 1268–1298. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Qiu, F.; Lan, N.; Zhang, T.; Guo, X.; Jin, Q.; Liu, R.; Wang, X. Analysis of phytochemical composition of Camellia oleifera oil and evaluation of its anti-inflammatory effect in lipopolysaccharide-stimulated RAW 264.7 macrophages. Lipids 2020, 55, 353–363. [Google Scholar] [CrossRef]
- Xiao, X.; He, L.; Chen, Y.; Wu, L.; Wang, L.; Liu, Z. Anti-inflammatory and antioxidative effects of Camellia oleifera abel components. Future Med. Chem. 2017, fmc-2017-0109. [Google Scholar] [CrossRef]
- Pallares, V.; Calay, D.; Cedo, L.; Castell-Auvi, A.; Raes, M.; Pinent, M.; Ardevol, A.; Arola, L.; Blay, M. Additive, antagonistic, and synergistic effects of procyanidins and polyunsaturated fatty acids over inflammation in RAW 264.7 macrophages activated by lipopolysaccharide. Nutrition 2012, 28, 447–457. [Google Scholar] [CrossRef]
- Tao, J.-Y.; Zheng, G.-H.; Zhao, L.; Wu, J.-G.; Zhang, X.-Y.; Zhang, S.-L.; Huang, Z.-J.; Xiong, F.-L.; Li, C.-M. Anti-inflammatory effects of ethyl acetate fraction from melilotus suaveolens ledeb on LPS-stimulated RAW 264.7 cells. J. Ethnopharmacol. 2009, 123, 97–105. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Kim, J.-H.; So, Y.; Kang, S.-Y.; Jeong, H.-G.; Jin, C.-H. Anti-inflammatory effect of lupinalbin a isolated from Apios americna on lipopolysaccharide-treated RAW264.7 cells. Molecules 2018, 23, 583. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Fu, J.-N.; Chou, T.-C. Synergistic combination of microtubule targeting anticancer fludelone with cytoprotective panaxytriol derived from panax ginseng against MX-1 cells in vitro: Experimental design and data analysis using the combination index method. Am. J. Cancer Res. 2016, 6, 97–104. [Google Scholar]
- Huang, H.; Chen, X.; Li, D.; He, Y.; Li, Y.; Du, Z.; Zhang, K.; DiPaola, R.; Goodin, S.; Zheng, X. Combination of α-tomatine and curcumin inhibits growth and induces apoptosis in human prostate cancer cells. PLoS ONE 2015, 10, e0144293. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Tollefsbol, T.O. Combinational proanthocyanidins and resveratrol synergistically inhibit human breast cancer cells and impact epigenetic–mediating machinery. Int. J. Mol. Sci. 2018, 19, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.S. Antioxidant and antibacterial activities of chitosan-phloroglucinol conjugate. Fish. Aquat. Sci. 2013, 16, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Lechner, M.; Lirk, P.; Rieder, J. Inducible nitric oxide synthase (iNOS) in tumor biology: The two sides of the same coin. Semin. Cancer Biol. 2005, 15, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Hu, L.; Jin, X.-L.; Ma, Q.-X.; Marcucci, M.C.; Netto, A.A.L.; Sawaya, A.C.H.F.; Huang, S.; Ren, W.-K.; Conlon, M.A.; et al. Polyphenol-rich propolis extracts from china and brazil exert anti-inflammatory effects by modulating ubiquitination of TRAF6 during the activation of NF-κB. J. Funct. Foods 2015, 19, 464–478. [Google Scholar] [CrossRef]
- Cheng, Y.-T.; Wu, S.-L.; Ho, C.-Y.; Huang, S.-M.; Cheng, C.-L.; Yen, G.-C. Beneficial effects of camellia oil (Camellia oleifera abel.) on ketoprofen-induced gastrointestinal mucosal damage through upregulation of HO-1 and VEGF. J. Agr. Food Chem. 2014, 62, 642–650. [Google Scholar] [CrossRef]
- Vasarri, M.; Leri, M.; Barletta, E.; Ramazzotti, M.; Marzocchini, R.; Degl’Innocenti, D. Anti-inflammatory properties of the marine plant Posidonia oceanica (L.) delile. J. Ethnopharmacol. 2020, 247, 112252. [Google Scholar] [CrossRef]
- Miller, M.W.; Lin, A.P.; Wolf, E.J.; Miller, D.R. Oxidative stress, inflammation, and neuroprogression in chronic PTSD. Harvard Rev. Psychiat. 2018, 26, 57–69. [Google Scholar] [CrossRef]
- Bumrungpert, A.; Pavadhgul, P.; Kalpravidh, R.W. Camellia oil-enriched diet attenuates oxidative stress and inflammatory markers in hypercholesterolemic subjects. J. Med. Food 2016, 19, 895–898. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: An updated review of their molecular mechanisms and experimental models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Baltimore, D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences (reprinted from cell, vol 46, pg 705–716, 1986). J. Immunol. 2006, 177, 7485–7496. [Google Scholar] [PubMed]
- Hamadou, M.H.; Kerkatou, M.; Zucal, C.; Bisio, A.; Provenzani, A.; Inga, A.; Menad, A.; Benayache, S.; Benayache, F.; Ameddah, S. Limonium duriusculum (de Girard) kuntze exhibits anti-inflammatory effect via NF-κB pathway modulation. Braz. Arch. Biol. Techn. 2021, 64, e21200179. [Google Scholar] [CrossRef]
- Sanchez-Fidalgo, S.; Villegas, I.; Rosillo, M.A.; Aparicio-Soto, M.; de la Lastra, C.A. Dietary squalene supplementation improves dss-induced acute colitis by downregulating p38 MAPK and NFκB signaling pathways. Mol. Nutr. Food Res. 2015, 59, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Lagha, A.B.; Grenier, D. Tea polyphenols inhibit the activation of NF-κB and the secretion of cytokines and matrix metalloproteinases by macrophages stimulated with Fusobacterium nucleatum. Sci. Rep. 2016, 6, 34520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Shu, W.; Shen, Y.; Sun, Q.; Bai, F.; Wang, J.; Li, D.; Li, Y.; Jin, W.; Yuan, L. Sturgeon protein-derived peptides exert anti-inflammatory effects in lps-stimulated RAW264.7 macrophages via the MAPK pathway. J. Funct. Foods 2020, 72, 104044. [Google Scholar] [CrossRef]
- Liu, J.; Huang, H.; Huang, Z.; Ma, Y.; Zhang, L.; He, Y.; Li, D.; Liu, W.; Goodin, S.; Zhang, K.; et al. Eriocitrin in combination with resveratrol ameliorates LPS-induced inflammation in RAW264.7 cells and relieves TPA-induced mouse ear edema. J. Funct. Foods 2019, 56, 321–332. [Google Scholar] [CrossRef]
- Ekstrand-Hammarstrom, B.; Osterlund, C.; Lilliehook, B.; Bucht, A. Vitamin e down-modulates mitogen-activated protein kinases, nuclear factor-κB and inflammatory responses in lung epithelial cells. Clin. Exp. Immunol. 2007, 147, 359–369. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Chen, J.; Liang, R.; Liu, C.; Chen, M.; Chen, J. Synergistic Anti-Inflammatory Effects of Lipophilic Grape Seed Proanthocyanidin and Camellia Oil Combination in LPS-Stimulated RAW264.7 Cells. Antioxidants 2022, 11, 289. https://doi.org/10.3390/antiox11020289
Zhang L, Chen J, Liang R, Liu C, Chen M, Chen J. Synergistic Anti-Inflammatory Effects of Lipophilic Grape Seed Proanthocyanidin and Camellia Oil Combination in LPS-Stimulated RAW264.7 Cells. Antioxidants. 2022; 11(2):289. https://doi.org/10.3390/antiox11020289
Chicago/Turabian StyleZhang, Linli, Juan Chen, Ruihong Liang, Chengmei Liu, Mingshun Chen, and Jun Chen. 2022. "Synergistic Anti-Inflammatory Effects of Lipophilic Grape Seed Proanthocyanidin and Camellia Oil Combination in LPS-Stimulated RAW264.7 Cells" Antioxidants 11, no. 2: 289. https://doi.org/10.3390/antiox11020289
APA StyleZhang, L., Chen, J., Liang, R., Liu, C., Chen, M., & Chen, J. (2022). Synergistic Anti-Inflammatory Effects of Lipophilic Grape Seed Proanthocyanidin and Camellia Oil Combination in LPS-Stimulated RAW264.7 Cells. Antioxidants, 11(2), 289. https://doi.org/10.3390/antiox11020289