
Effect of Artemisinin on the Redox System of NADPH/FNR/Ferredoxin from Malaria Parasites

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site-Directed Mutagenesis of PfFd and PfFNR and Preparation of Recombinant Proteins
2.2. Enzymatic Analyses
2.3. In Silico Docking Analysis
3. Results and Discussion
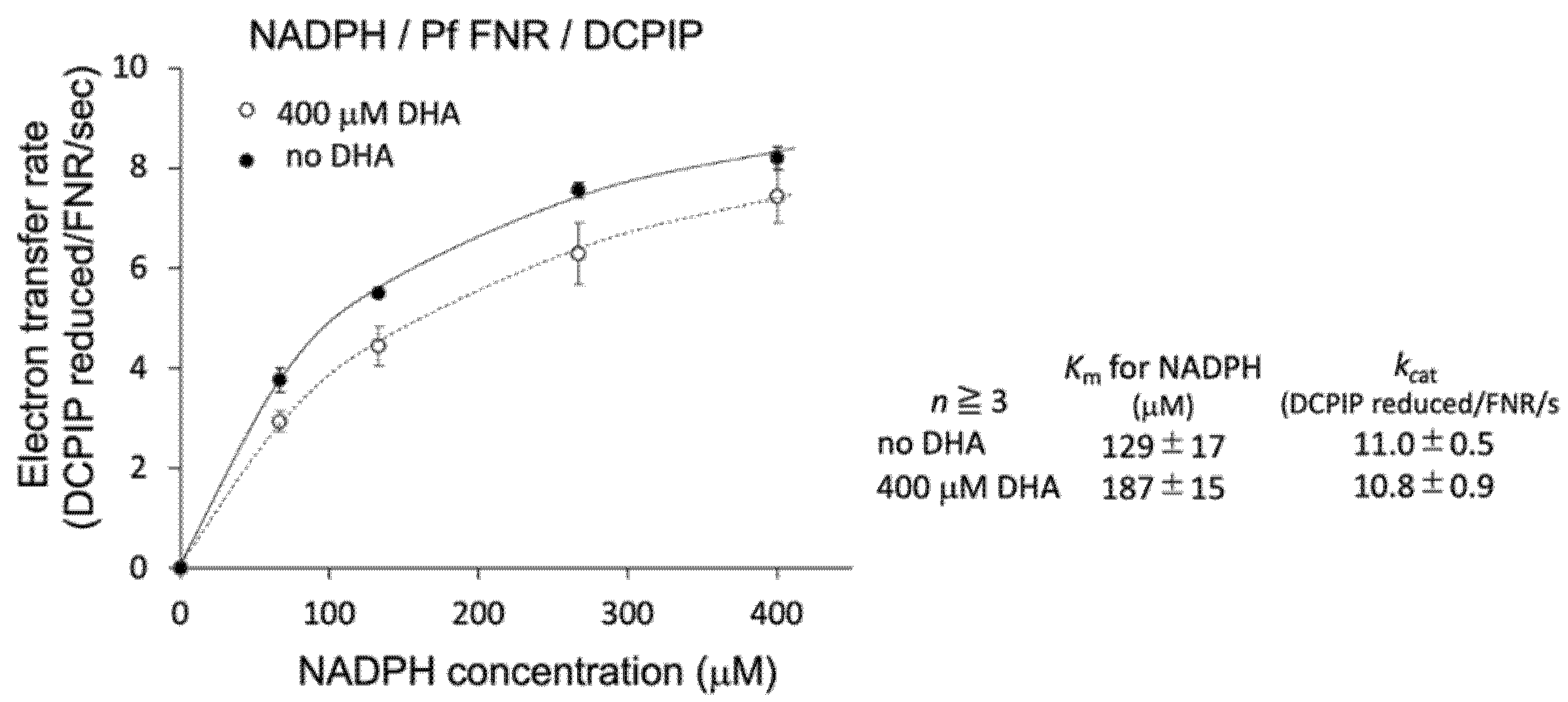
3.1. Effect of DHA on the Catalytic Activity of PfFNR
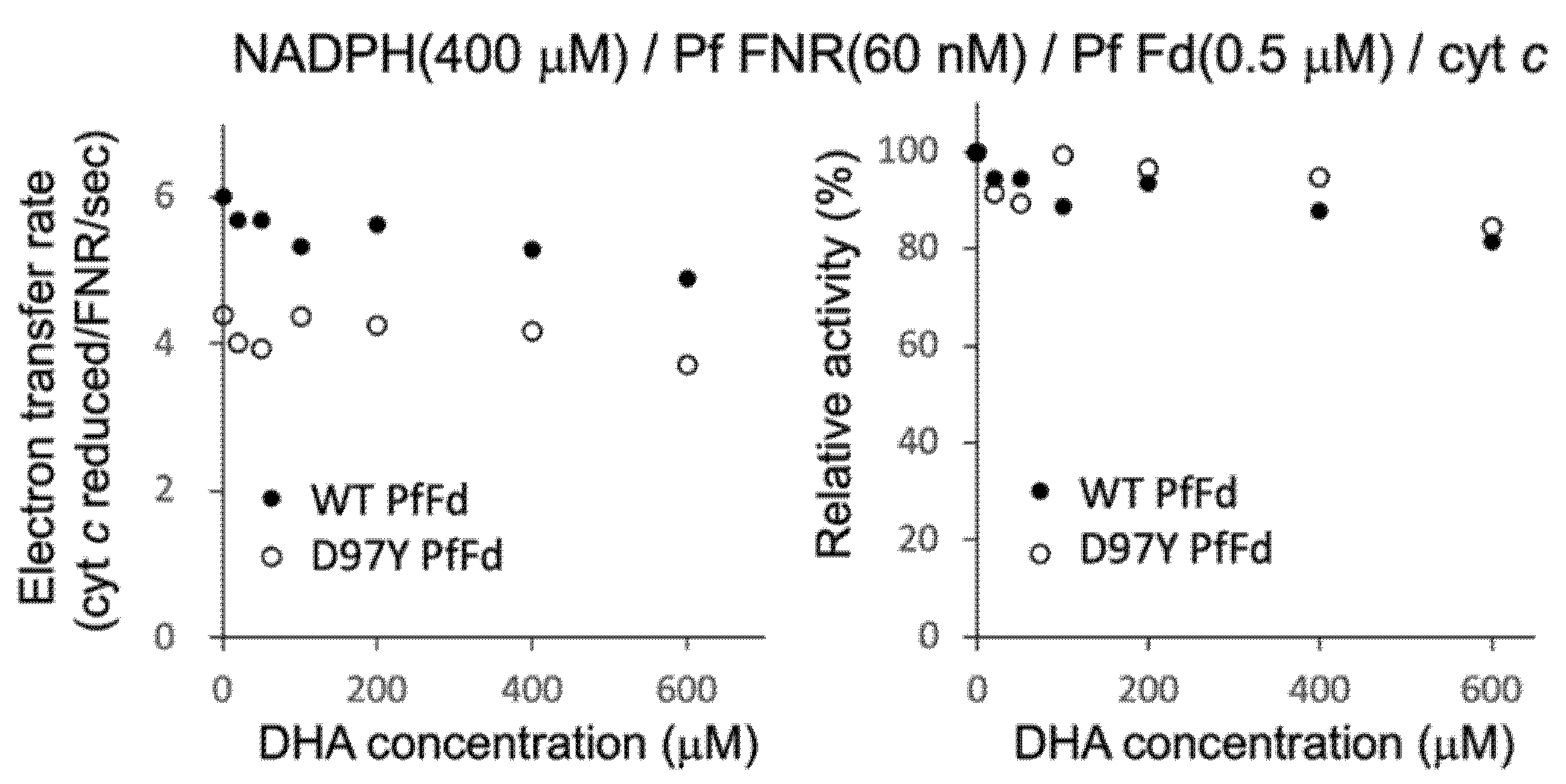
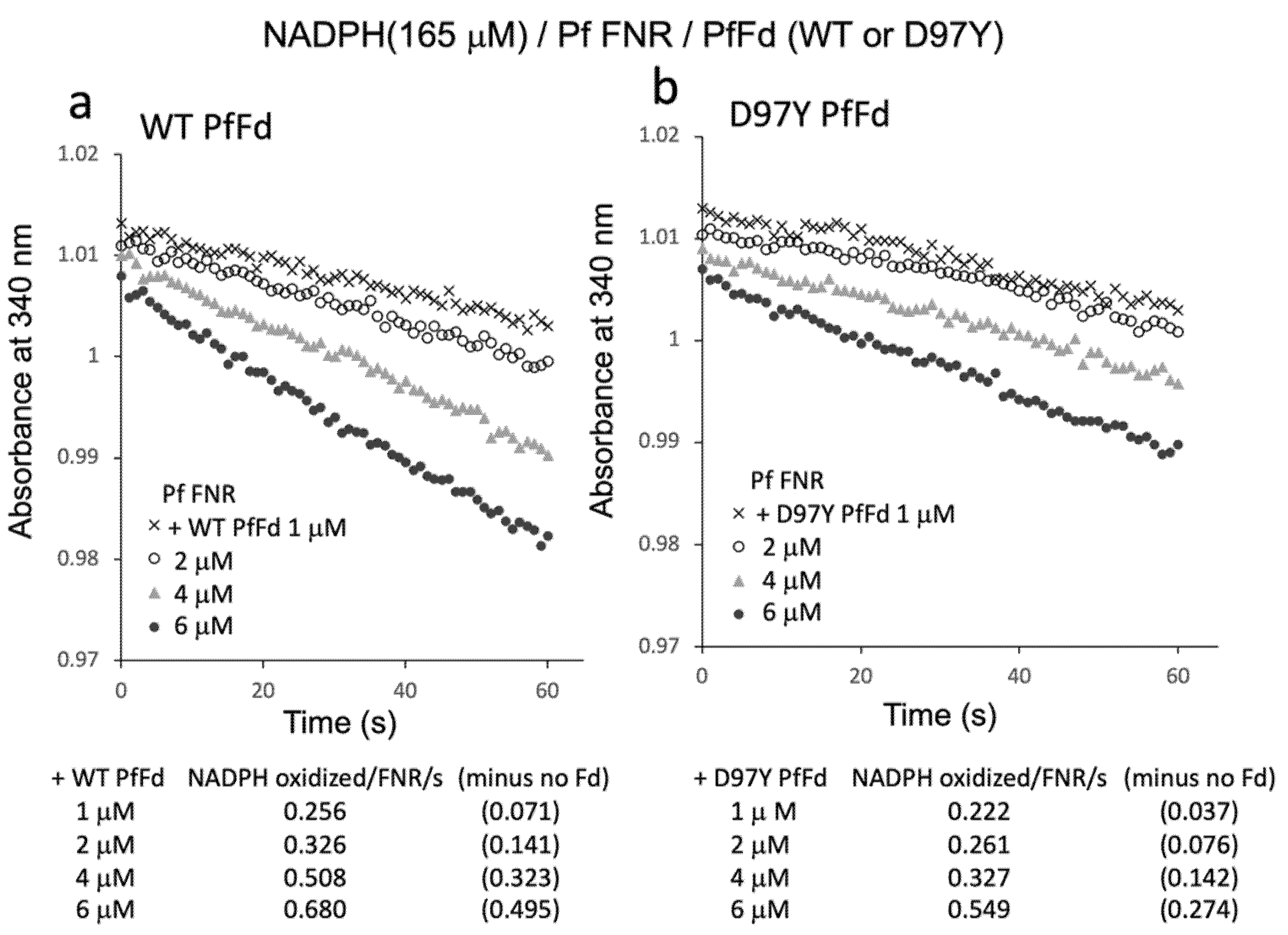
3.2. Effect of DHA on the Electron Transfer Activity from PfFNR to Wild-Type and Mutant PfFds
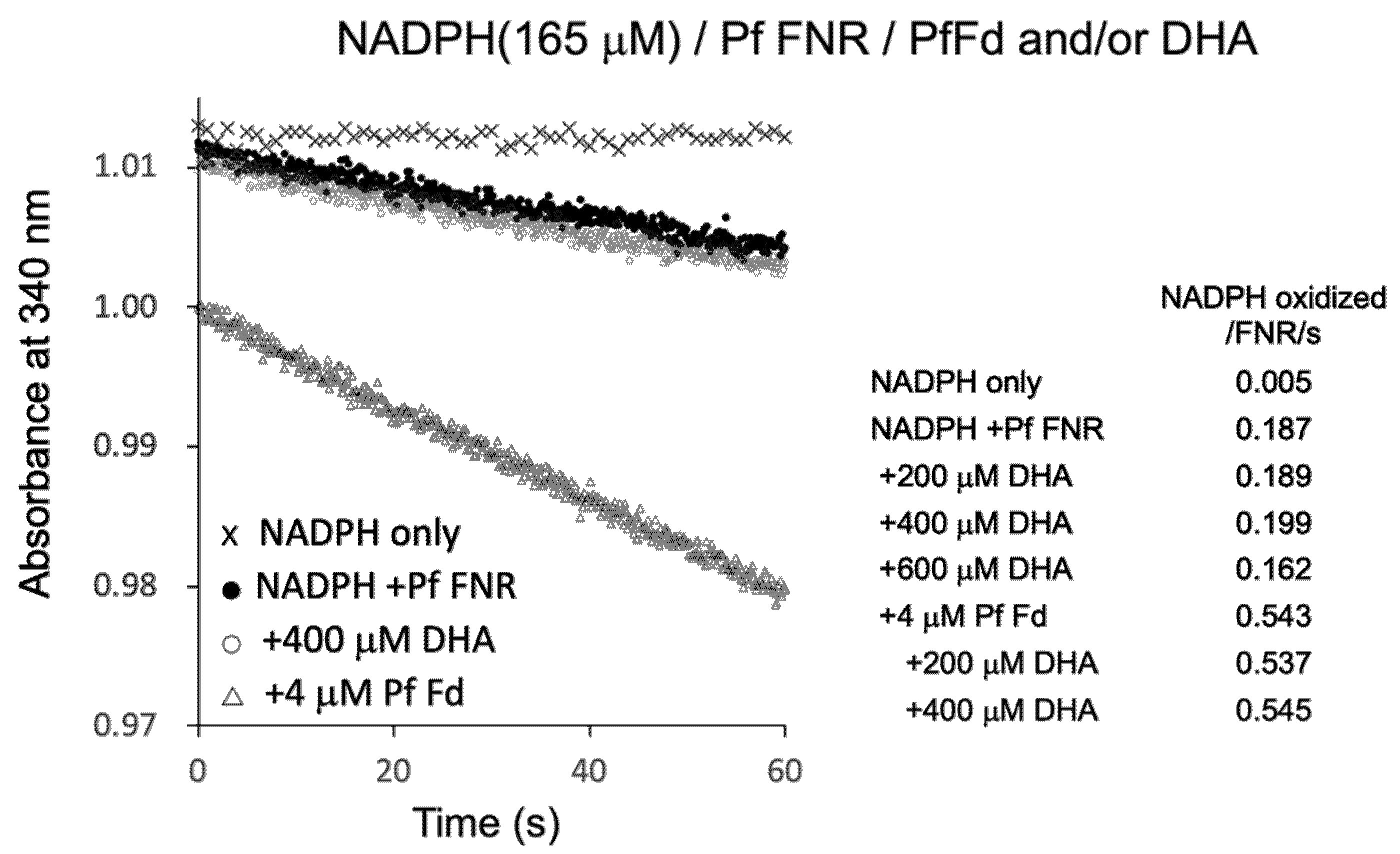
3.3. Reaction of PfFNR with DHA
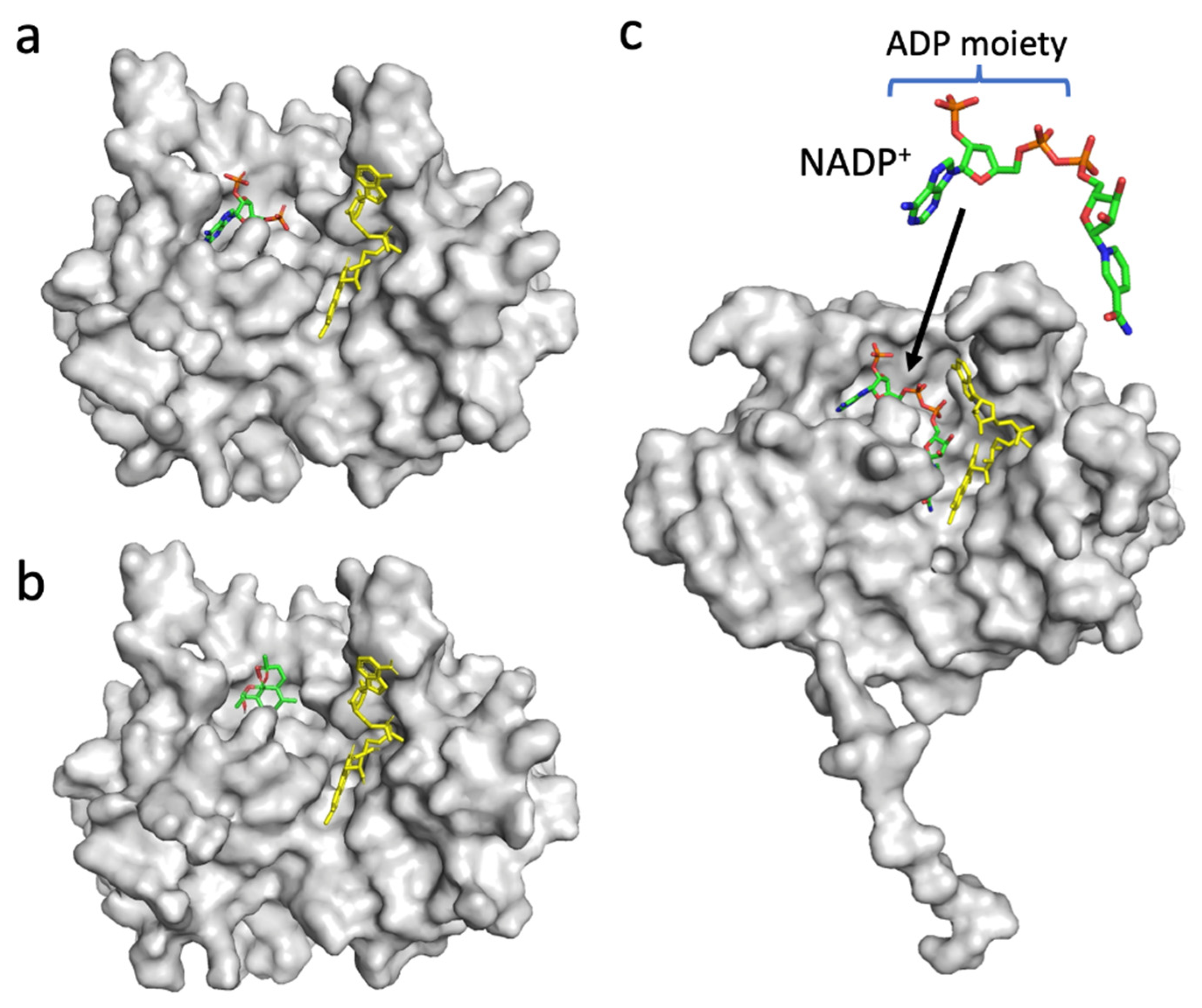
3.4. In Silico Docking Analysis of PfFNR and DHA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miotto, O.; Amato, R.; Ashley, E.A.; MacInnis, B.; Almagro-Garcia, J.; Amaratunga, C.; Lim, P.; Mead, D.; Oyola, S.O.; Dhorda, M.; et al. Genetic architecture of artemisinin-resistant Plasmodium falciparum. Nat. Genet. 2015, 47, 226–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miotto, O.; Sekihara, M.; Tachibana, S.-I.; Yamauchi, M.; Pearson, R.D.; Amato, R.; Gonçalves, S.; Mehra, S.; Noviyanti, R.; Marfurt, J.; et al. Emergence of artemisinin-resistant Plasmodium falciparum with kelch13 C580Y mutations on the island of New Guinea. PLoS Pathog. 2020, 16, e1009133. [Google Scholar] [CrossRef] [PubMed]
- Köhler, S.; Delwiche, C.F.; Denny, P.W.; Tilney, L.G.; Webster, P.; Wilson, R.J.M.; Palmer, J.D.; Roos, D.S. A Plastid of Probable Green Algal Origin in Apicomplexan Parasites. Science 1997, 275, 1485–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fichera, M.E.; Roos, D. A plastid organelle as a drug target in apicomplexan parasites. Nature 1997, 390, 407–409. [Google Scholar] [CrossRef]
- Yeh, E.; DeRisi, J.L. Chemical Rescue of Malaria Parasites Lacking an Apicoplast Defines Organelle Function in Blood-Stage Plasmodium falciparum. PLoS Biol. 2011, 9, e1001138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimata-Ariga, Y.; Saitoh, T.; Ikegami, T.; Horii, T.; Hase, T. Molecular interaction of ferredoxin and ferredoxin-NADP+ re-ductase from human malaria parasite. J. Biochem. 2007, 142, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Kimata-Ariga, Y.; Sakamoto, A.; Kamatani, M.; Saitoh, T.; Hase, T. C-terminal aromatic residue of Plasmodium ferredoxin important for the interaction with ferredoxin: NADP (H) oxidoreductase: Possible involvement for artemisinin resistance of human malaria parasites. J. Biochem. 2020, 168, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Kimata-Ariga, Y.; Morihisa, R. Functional analyses of plasmodium ferredoxin Asp97Tyr mutant related to artemisinin resistance of human malaria parasites. J. Biochem. 2021, 170, 521–529. [Google Scholar] [CrossRef]
- Posner, G.H.; O’neill, P.M. Knowledge of the Proposed Chemical Mechanismof Action and Cytochrome P450 Metabolismof Antimalarial Trioxanes Like Artemisinin Allows Rational Design of New Antimalarial Peroxides. Acc. Chem. Res. 2004, 37, 397–404. [Google Scholar] [CrossRef]
- Egwu, C.O.; Tsamesidis, I.; Pério, P.; Augereau, J.-M.; Benoit-Vical, F.; Reybier, K. Superoxide: A major role in the mechanism of action of essential antimalarial drugs. Free Radic. Biol. Med. 2021, 167, 271–275. [Google Scholar] [CrossRef]
- Müller, S. Redox and antioxidant systems of the malaria parasite Plasmodium falciparum. Mol. Microbiol. 2004, 53, 1291–1305. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A.R.; Rodriguez, R.E.; Poli, H.O.; Paladini, D.H.; Palatnik, J.F.; Carrillo, N. The flavoenzyme ferredoxin (flavodox-in)-NADP(H) reductase modulates NADP(H) homeostasis during the soxRS response of Escherichia coli. J. Bacteriol. 2002, 184, 1474–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozuleva, M.; Goss, T.; Twachtmann, M.; Rudi, K.; Trapka, J.; Selinski, J.; Ivanov, B.; Garapati, P.; Steinhoff, H.-J.; Hase, T.; et al. Ferredoxin:NADP(H) Oxidoreductase Abundance and Location Influences Redox Poise and Stress Tolerance. Plant Physiol. 2016, 172, 1480–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, M.; Balconi, E.; Aliverti, A.; Mastrangelo, E.; Seeber, F.; Bolognesi, M.; Zanetti, G. Ferredoxin-NADP+ Reductase from Plasmodium falciparum Undergoes NADP+-dependent Dimerization and Inactivation: Functional and Crystallographic Analysis. J. Mol. Biol. 2007, 367, 501–513. [Google Scholar] [CrossRef]
- Onda, Y.; Matsumura, T.; Kimata-Ariga, Y.; Sakakibara, H.; Sugiyama, T.; Hase, T. Differential Interaction of Maize Root Ferredoxin:NADP+ Oxidoreductase with Photosynthetic and Non-Photosynthetic Ferredoxin Isoproteins. Plant Physiol. 2000, 123, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Kimata-Ariga, Y.; Yuasa, S.; Saitoh, T.; Fukuyama, H.; Hase, T. Plasmodium-specific basic amino acid residues important for the interaction with ferredoxin on the surface of ferredoxin-NADP+ reductase. J. Biochem. 2018, 164, 231–237. [Google Scholar] [CrossRef]
- Massey, V. The microestimation of succinate and the extinction coefficient of cytochrome c. Biochim. Biophys. Acta 1959, 34, 255–256. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Grellier, P.; Marozienė, A.; Nivinskas, H.; Šarlauskas, J.; Aliverti, A.; Čėnas, N. Antiplasmodial activity of quinones: Roles of aziridinyl substituents and the inhibition of Plasmodium falciparum glutathione reductase. Arch. Biochem. Biophys. 2010, 494, 32–39. [Google Scholar] [CrossRef]
- Marozienė, A.; Lesanavičius, M.; Davioud-Charvet, E.; Aliverti, A.; Grellier, P.; Šarlauskas, J.; Čėnas, N. Antiplasmodial ac-tivity of nitroaromatic compounds: Correlation with their reduction potential and inhibitory action on Plasmodium falciparum glutathione reductase. Molecules 2019, 24, 4509. [Google Scholar] [CrossRef] [Green Version]
- Lesanavičius, M.; Aliverti, A.; Šarlauskas, J.; Čėnas, N. Reactions of Plasmodium falciparum Ferredoxin:NADP+ Oxidoreductase with Redox Cycling Xenobiotics: A Mechanistic Study. Int. J. Mol. Sci. 2020, 21, 3234. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.-J.; Ni Chia, W.; Loh, C.C.Y.; Li, Z.; Lee, Y.M.; He, Y.; Yuan, L.-X.; Lim, T.K.; Liu, M.; et al. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Aliverti, A.; Zanetti, G.; Arakaki, A.K.; Ottado, J.; Orellano, E.G.; Calcaterra, N.B.; Ceccarelli, E.A.; Carrillo, N.; Karplus, P.A. A productive NADP+ binding mode of ferredoxin-NADP + reductase revealed by protein engineering and crystallographic studies. Nat. Genet. 1999, 6, 847–853. [Google Scholar] [CrossRef]
- Siddiqui, F.A.; Liang, X.; Cui, L. Plasmodium falciparum resistance to ACTs: Emergence, mechanisms, and outlook. Int. J. Parasitol. Drugs Drug Resist. 2021, 16, 102–118. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Dietz, K.J.; Horling, F.; König, J.; Baier, M. The function of the chloroplast 2-cysteine peroxiredoxin in peroxide detoxification and its regulation. J. Exp. Bot. 2002, 53, 1321–1329. [Google Scholar]
- Zhang, M.; Wang, C.; Oberstaller, J.; Thomas, P.; Otto, T.D.; Casandra, D.; Boyapalle, S.; Adapa, S.R.; Xu, S.; Button-Simons, K.; et al. The apicoplast link to fever-survival and artemisinin-resistance in the malaria parasite. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Röhrich, R.C.; Englert, N.; Troschke, K.; Reichenberg, A.; Hintz, M.; Seeber, F.; Balconi, E.; Aliverti, A.; Zanetti, G.; Köhler, U.; et al. Reconstitution of an apicoplast-localised electron transfer pathway involved in the isoprenoid biosynthesis ofPlasmodium falciparum. FEBS Lett. 2005, 579, 6433–6438. [Google Scholar] [CrossRef] [Green Version]
- Saggu, G.S.; Garg, S.; Pala, Z.R.; Yadav, S.K.; Kochar, S.K.; Kochar, D.K.; Saxena, V. Characterization of 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase (IspG) from Plasmodium vivax and it’s potential as an antimalarial drug target. Int. J. Biol. Macromol. 2017, 96, 466–473. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimata-Ariga, Y.; Morihisa, R. Effect of Artemisinin on the Redox System of NADPH/FNR/Ferredoxin from Malaria Parasites. Antioxidants 2022, 11, 273. https://doi.org/10.3390/antiox11020273
Kimata-Ariga Y, Morihisa R. Effect of Artemisinin on the Redox System of NADPH/FNR/Ferredoxin from Malaria Parasites. Antioxidants. 2022; 11(2):273. https://doi.org/10.3390/antiox11020273
Chicago/Turabian StyleKimata-Ariga, Yoko, and Rena Morihisa. 2022. "Effect of Artemisinin on the Redox System of NADPH/FNR/Ferredoxin from Malaria Parasites" Antioxidants 11, no. 2: 273. https://doi.org/10.3390/antiox11020273
APA StyleKimata-Ariga, Y., & Morihisa, R. (2022). Effect of Artemisinin on the Redox System of NADPH/FNR/Ferredoxin from Malaria Parasites. Antioxidants, 11(2), 273. https://doi.org/10.3390/antiox11020273