
Protective Effect of Beta-Carotene against Myeloperoxidase- Mediated Oxidative Stress and Inflammation in Rat Ischemic Brain Injury

 ,
,  , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. Induction of Focal Cerebral I/R Injury
2.4. Experimental Protocol
2.5. Evaluation of Neurological Deficit
2.6. Behavioural Evaluation
2.6.1. Evaluation of Motor Coordination and Balance
2.6.2. Evaluation of Spontaneous Motor Activity
2.7. Euthanasia and Brain Tissue Samplings
2.8. Biochemical Analyses
2.8.1. Measurement of Brain Glutathione (GSH) Content
2.8.2. Assessment of Lipid Peroxidation (LPO) in Brain Tissues
2.8.3. Assessment of Superoxide Dismutase (SOD) Activity in Brain Tissues
2.8.4. Determination of Glutathione Peroxidase (GPx), Catalase (CAT), Nuclear Factor Kappa B (NF-κB), and Myeloperoxidase (MPO) in Brain Tissues
2.9. Histopathological Evaluation
2.10. Neuronal Cell Count
2.11. Immunohistochemistry of Caspase-3
2.12. Gene Expression Analysis by Real Time PCR
2.13. Statistical Analysis
3. Results
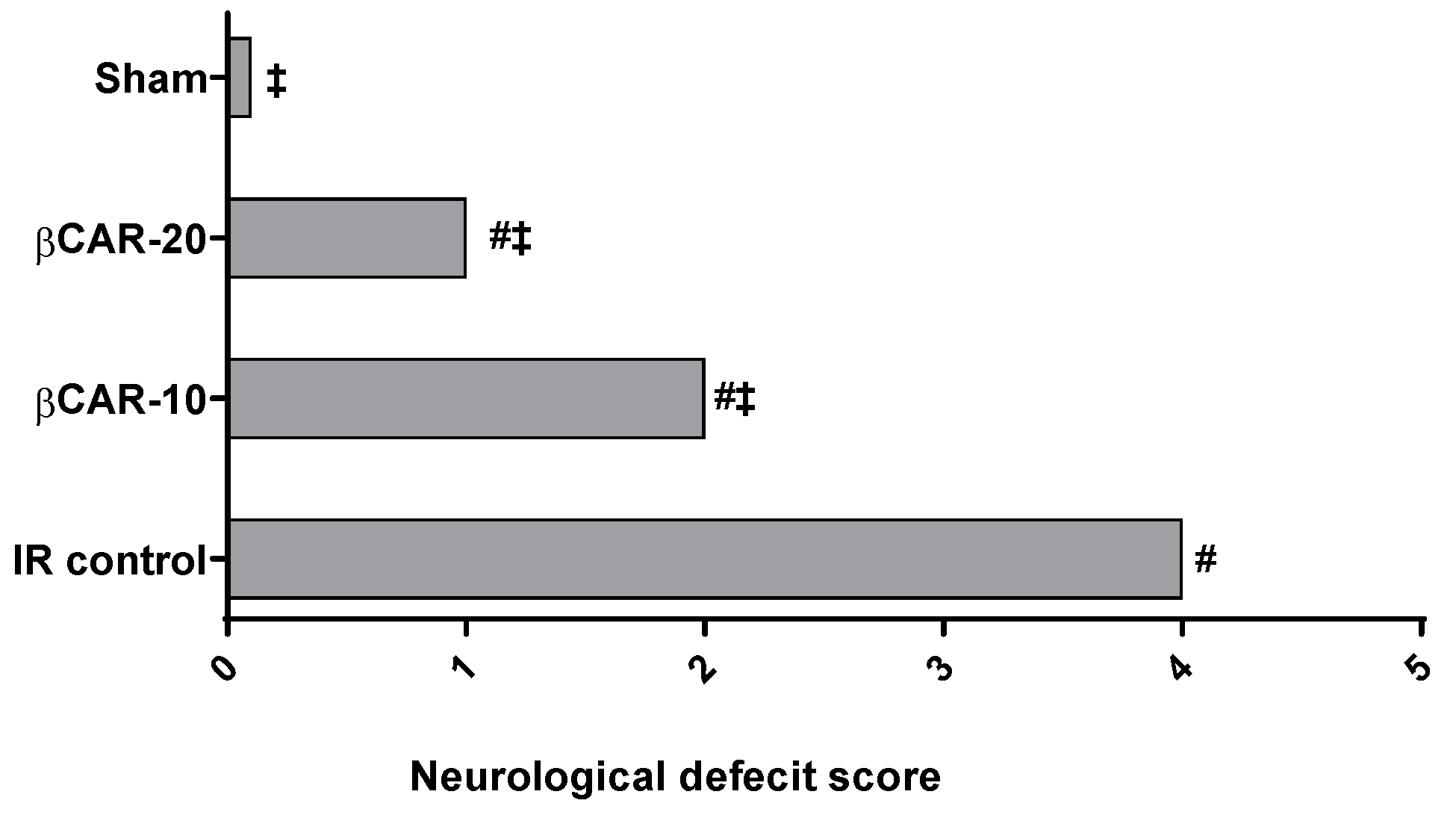
3.1. Evaluation of Neurological Deficit
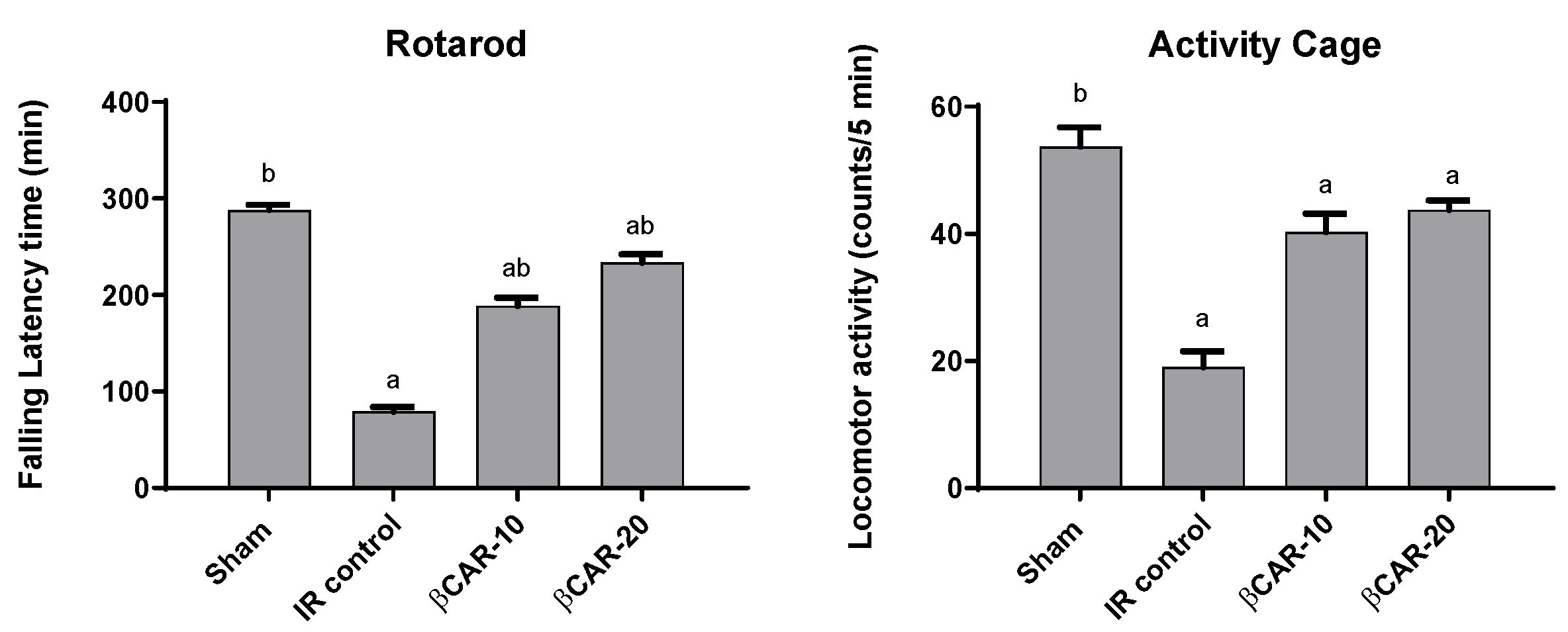
3.2. Behavioural Evaluation
3.2.1. Evaluation of Motor Coordination and Balance
3.2.2. Evaluation of Spontaneous Motor Activity
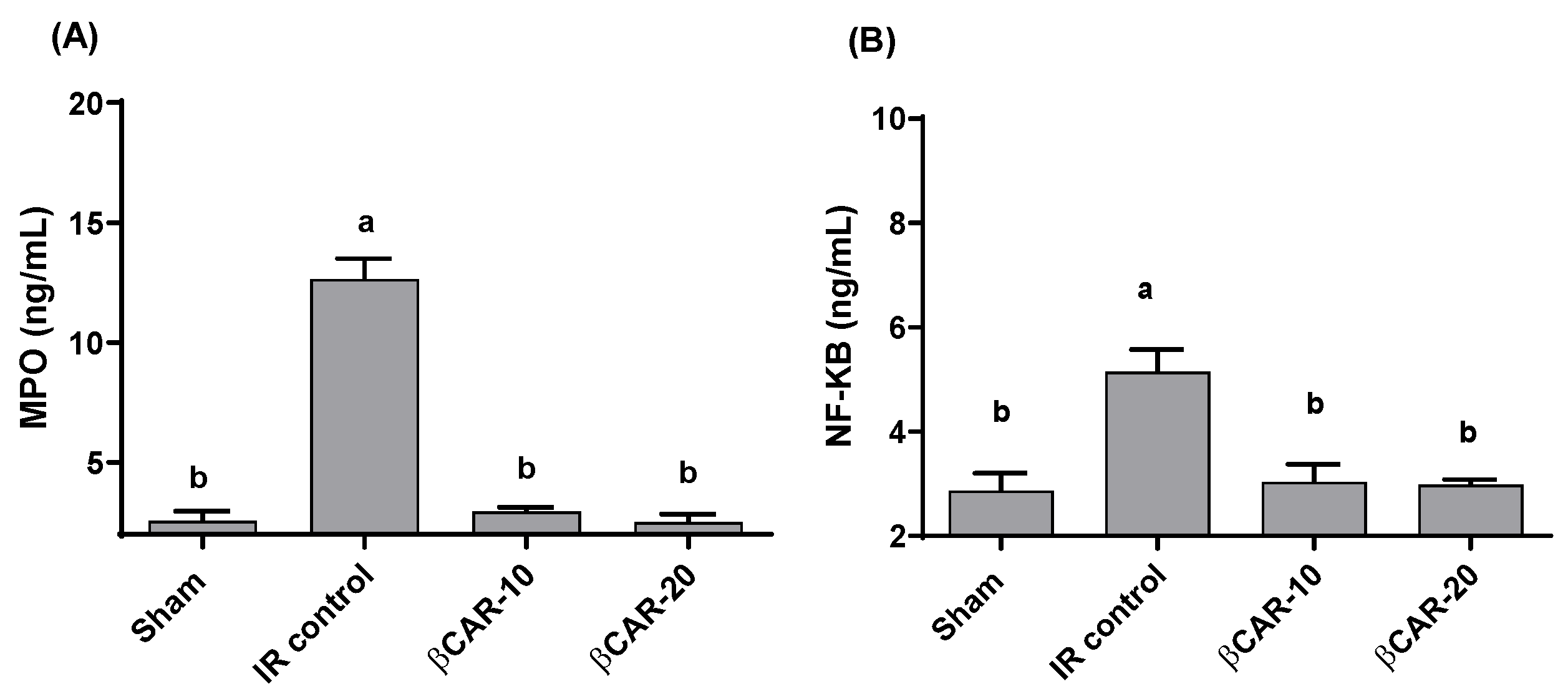
3.3. Effect of βCAR on Oxidative Stress Biomarkers
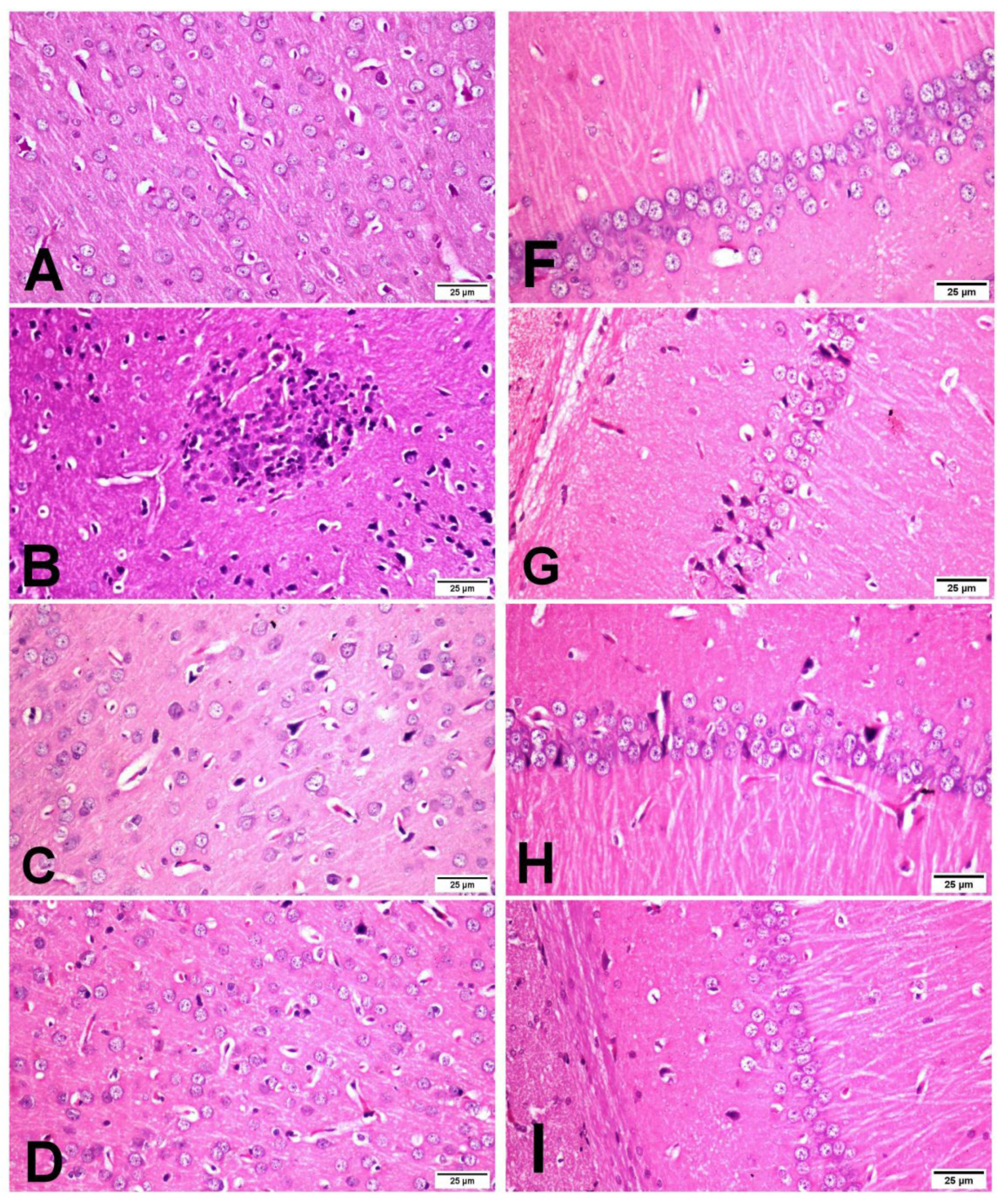
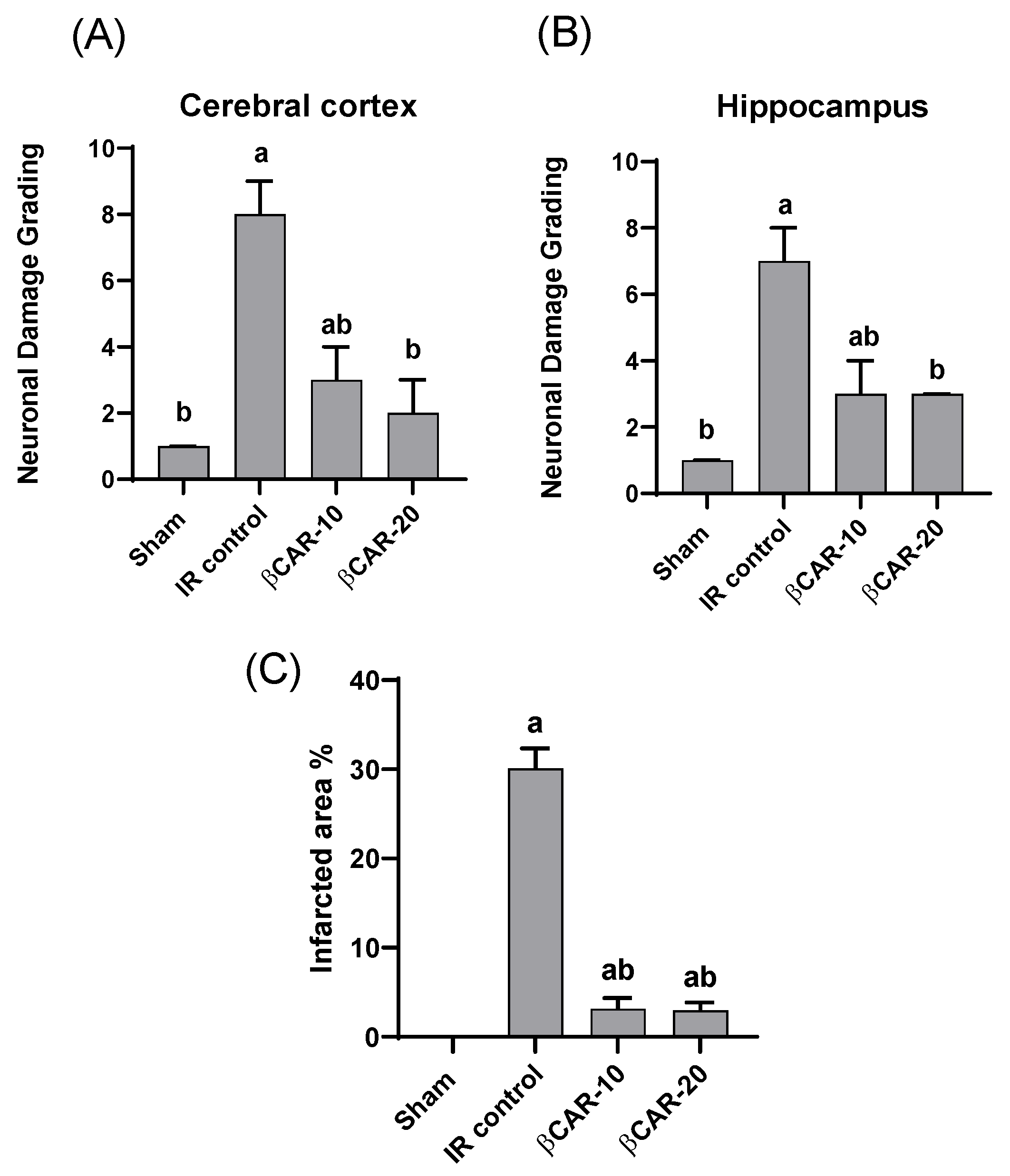
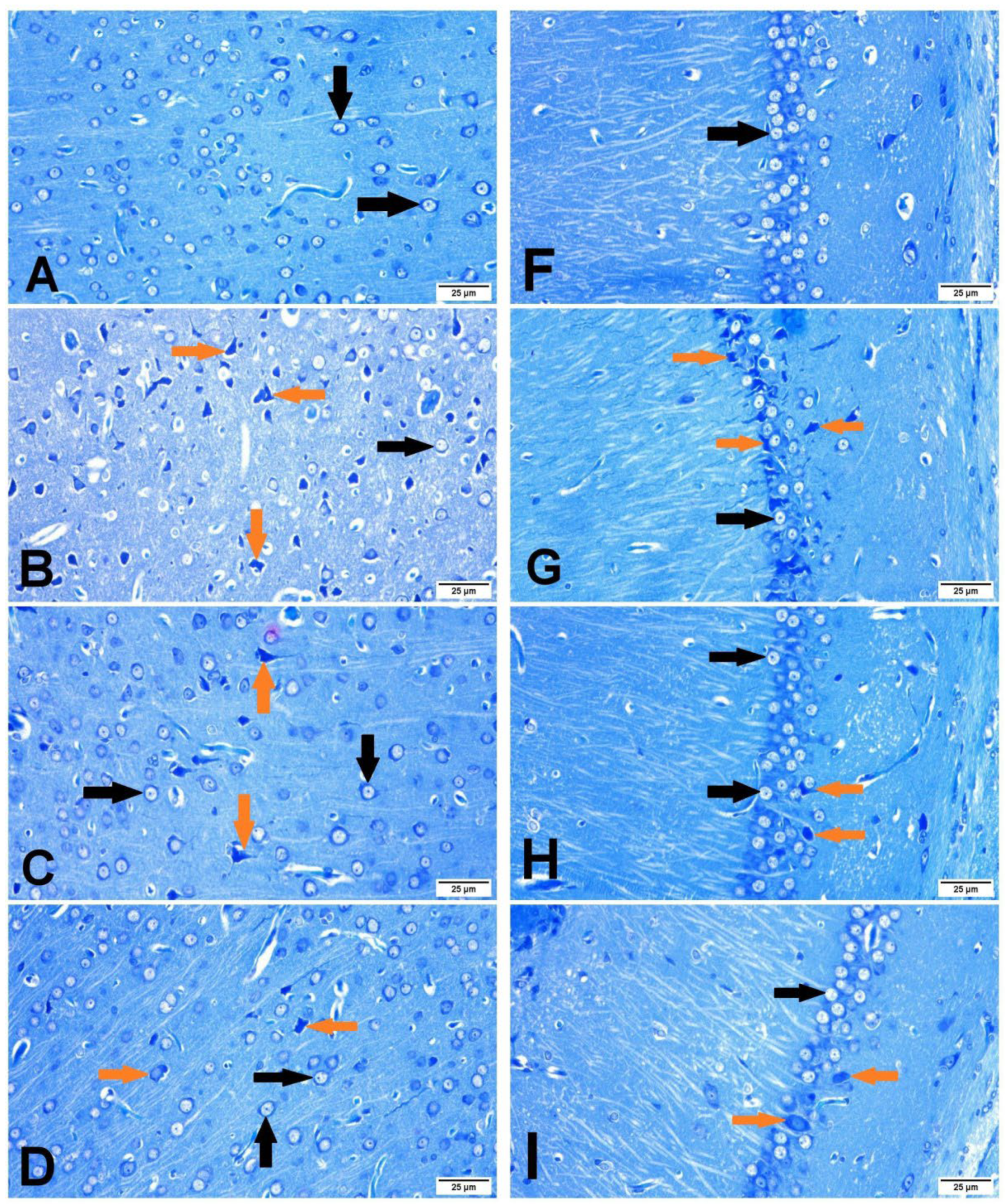
3.4. Histopathology of the Brain
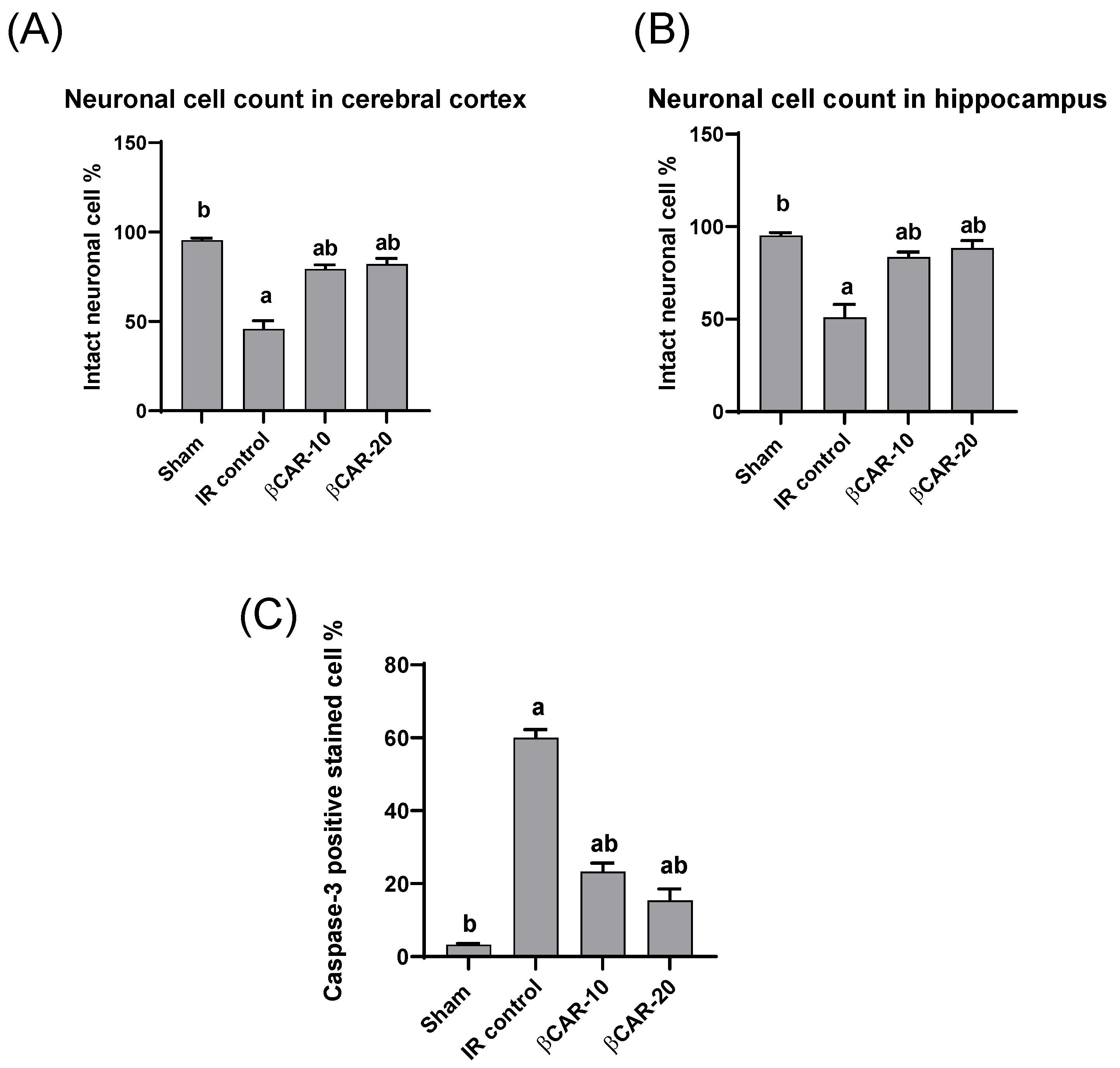
3.5. The Neuronal Cell Count
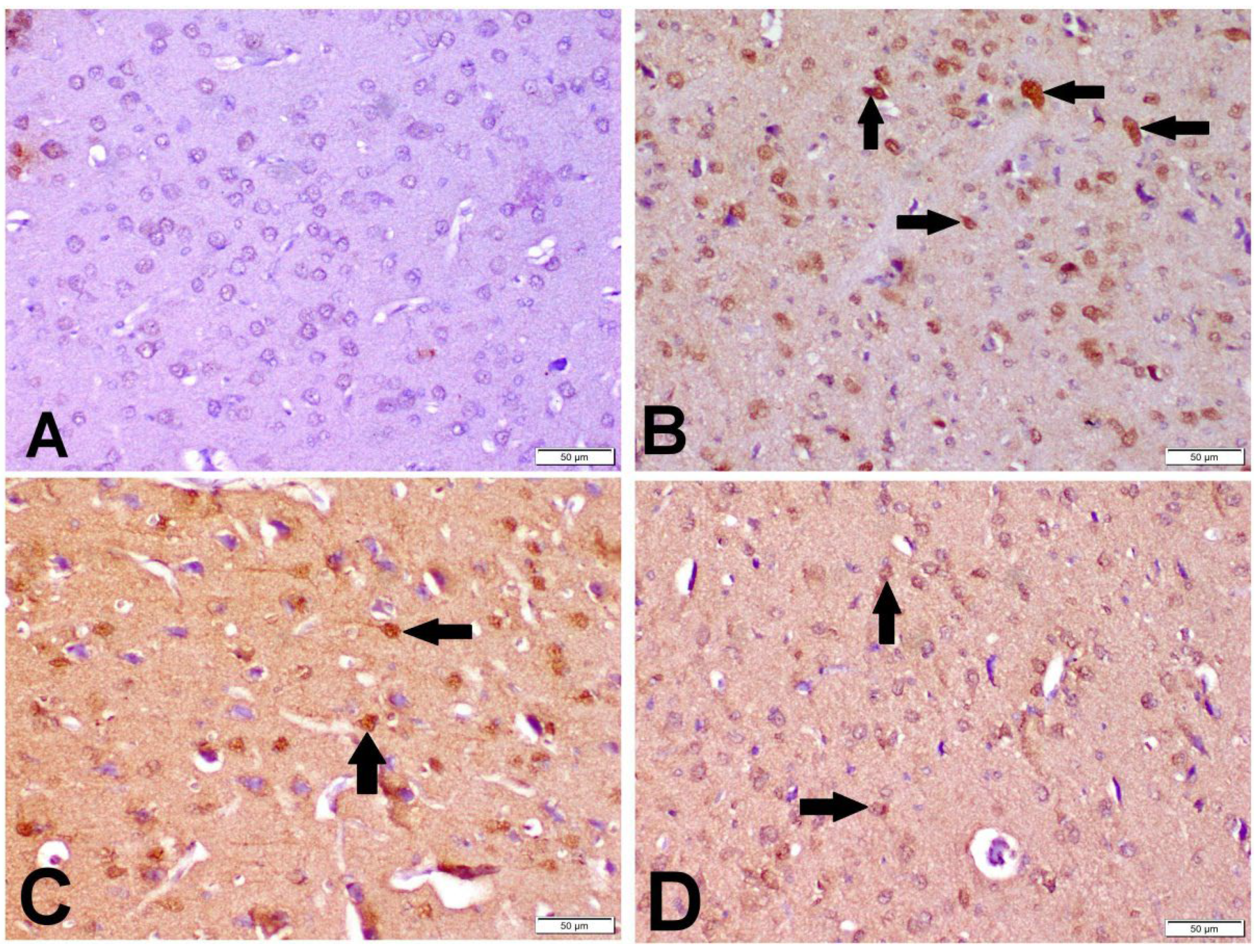
3.6. Immunohistochemistry of Caspase-3
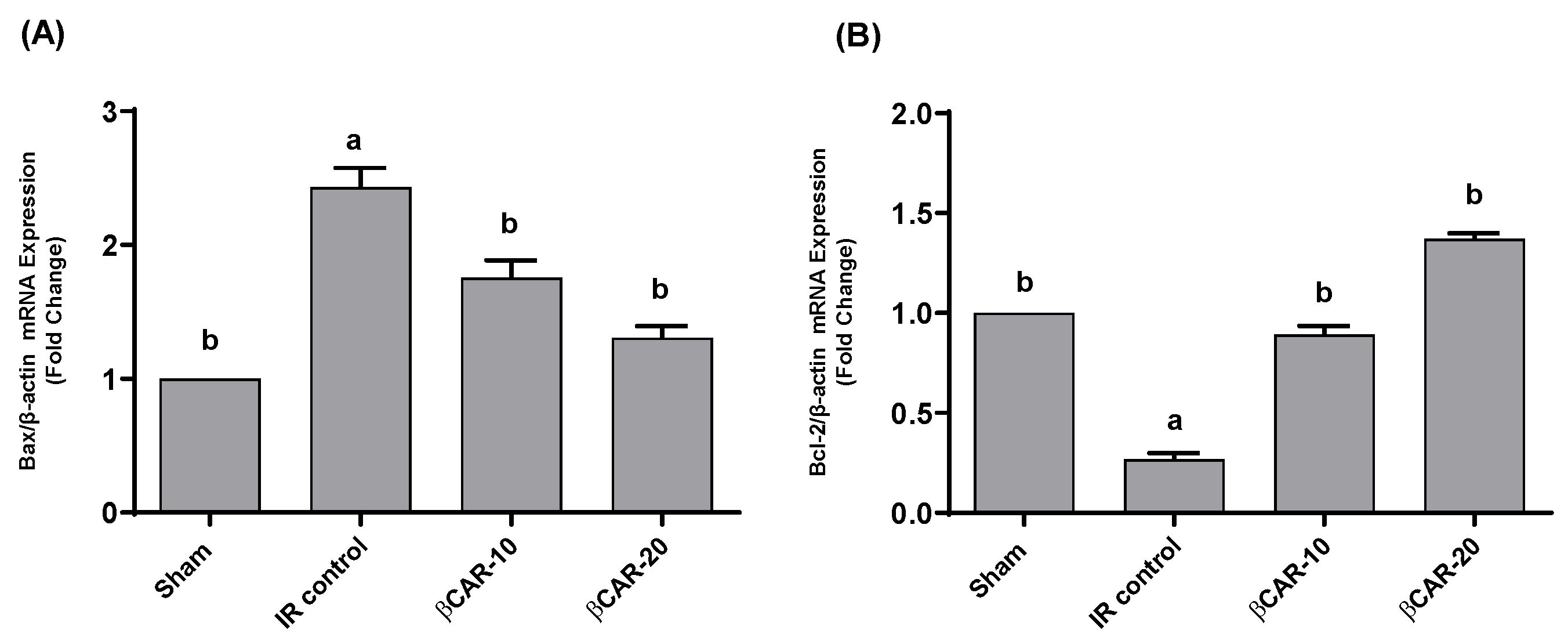
3.7. Expression of Bax and Bcl-2 mRNA in Brain Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, M.; Qu, Y.Z.; Zhao, Z.W.; Wu, S.X.; Liu, Y.Y.; Wei, X.Y.; Gao, L.; Gao, G.D. Astragaloside IV protects against focal cerebral ischemia/reperfusion injury correlating to suppression of neutrophils adhesion-related molecules. Neurochem. Int. 2012, 60, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Tian, T.; Cai, E.-L.; Yang, C.; Yang, X. miR-124 Alleviates Ischemic Stroke-Induced Neuronal Death by Targeting DAPK1 in Mice. Front. Neurosci. 2021, 15, 649982. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Zong, N.; Hu, Y.; Chen, Y.; Xu, Y. Neuronal Death Mechanisms and Therapeutic Strategy in Ischemic Stroke. Neurosci. Bull. 2022, 38, 1229–1247. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, M.; Jing, X.; Shi, H.; Ren, M.; Lou, H. (−)-Epigallocatechin gallate protects against cerebral ischemia-induced oxidative stress via Nrf2/ARE signaling. Neurochem. Res. 2014, 39, 1292–1299. [Google Scholar] [CrossRef]
- Del Zoppo, G.J.; Mabuchi, T. Cerebral microvessel responses to focal ischemia. J. Cereb. Blood Flow Metab. 2003, 23, 879–894. [Google Scholar] [CrossRef]
- Diaz-Ruiz, A.; Zavala, C.; Montes, S.; Ortiz-Plata, A.; Salgado-Ceballos, H.; Orozco-Suarez, S.; Nava-Ruiz, C.; Pérez-Neri, I.; Perez-Severiano, F.; Ríos, C. Antioxidant, antiinflammatory and antiapoptotic effects of dapsone in a model of brain ischemia/reperfusion in rats. J. Neurosci. Res. 2008, 86, 3410–3419. [Google Scholar] [CrossRef]
- Chen, H.; Yoshioka, H.; Kim, G.S.; Jung, J.E.; Okami, N.; Sakata, H.; Maier, C.M.; Narasimhan, P.; Goeders, C.E.; Chan, P.H. Oxidative stress in ischemic brain damage: Mechanisms of cell death and potential molecular targets for neuroprotection. Antioxid. Redox Signal. 2011, 14, 1505–1517. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Structures and analysis of carotenoid molecules. Carotenoids in Nature. In Subcellular Biochemistry; Springer: Cham, Switzerland, 2016; pp. 71–108. [Google Scholar]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Marine carotenoids against oxidative stress: Effects on human health. Mar. Drugs 2015, 13, 6226–6246. [Google Scholar] [CrossRef]
- Maoka, T. Recent progress in structural studies of carotenoids in animals and plants. Arch. Biochem. Biophys. 2009, 483, 191–195. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef]
- Shete, V.; Quadro, L. Mammalian metabolism of β-carotene: Gaps in knowledge. Nutrients 2013, 5, 4849–4868. [Google Scholar] [CrossRef]
- Sy, C.; Gleize, B.; Dangles, O.; Landrier, J.F.; Veyrat, C.C.; Borel, P. Effects of physicochemical properties of carotenoids on their bioaccessibility, intestinal cell uptake, and blood and tissue concentrations. Mol. Nutr. Food Res. 2012, 56, 1385–1397. [Google Scholar] [CrossRef]
- Haider, C.; Ferk, F.; Bojaxhi, E.; Martano, G.; Stutz, H.; Bresgen, N.; Knasmüller, S.; Alija, A.; Eckl, P.M. Effects of β-carotene and its cleavage products in primary pneumocyte type II cells. Antioxidants 2017, 6, 37. [Google Scholar] [CrossRef]
- De Oliveira, B.F.; Costa, D.C.; Nogueira-Machado, J.A.; Chaves, M.M. β-Carotene, α-tocopherol and ascorbic acid: Differential profile of antioxidant, inflammatory status and regulation of gene expression in human mononuclear cells of diabetic donors. Diabetes Metab. Res. Rev. 2013, 29, 636–645. [Google Scholar] [CrossRef]
- Chen, Q.-H.; Wu, B.-K.; Pan, D.; Sang, L.-X.; Chang, B. Beta-carotene and its protective effect on gastric cancer. World J. Clin. Cases 2021, 9, 6591. [Google Scholar] [CrossRef]
- Abdel-Rahman, R.F.; Alqasoumi, S.I.; Ogaly, H.A.; Abd-Elsalam, R.M.; El-Banna, H.A.; Soliman, G.A. Propolis ameliorates cerebral injury in focal cerebral ischemia/reperfusion (I/R) rat model via upregulation of TGF-β1. Saudi Pharm. J. 2020, 28, 116–126. [Google Scholar] [CrossRef]
- Cui, B.; Liu, S.; Wang, Q.; Lin, X. Effect of β-carotene on immunity function and tumour growth in hepatocellular carcinoma rats. Molecules 2012, 17, 8595–8603. [Google Scholar] [CrossRef]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar] [CrossRef]
- Vijitruth, R.; Liu, M.; Choi, D.-Y.; Nguyen, X.V.; Hunter, R.L.; Bing, G. Cyclooxygenase-2 mediates microglial activation and secondary dopaminergic cell death in the mouse MPTP model of Parkinson’s disease. J. Neuroinflammation 2006, 3, 6. [Google Scholar] [CrossRef]
- Abdel-Rahman, R.F.; El Awdan, S.A.; Hegazy, R.R.; Mansour, D.F.; Ogaly, H.A.; Abdelbaset, M. Neuroprotective effect of Crocus sativus against cerebral ischemia in rats. Metab. Brain Dis. 2020, 35, 427–439. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Bulaj, G.; Kortemme, T.; Goldenberg, D.P. Ionization− reactivity relationships for cysteine thiols in polypeptides. Biochemistry 1998, 37, 8965–8972. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Salam, O.M.E.; Abdel-Rahman, R.F.; Gaafar, A.E.-D.M. Behavioral and biochemical effects of Cannabis Sativa and their modulation by antidepressant drugs. Rev. Latinoam. De Química 2013, 41, 21–37. [Google Scholar]
- Ruiz-Larrea, M.B.; Leal, A.M.; Liza, M.; Lacort, M.; de Groot, H. Antioxidant effects of estradiol and 2-hydroxyestradiol on iron-induced lipid peroxidation of rat liver microsomes. Steroids 1994, 59, 383–388. [Google Scholar] [PubMed]
- Mansour, D.F.; Nada, S.A.; El-Denshary, E.S.; Omara, E.A.; Asaad, G.F.; Abdel-Rahman, R.F. Milk whey proteins modulate endotoxemia-induced hepatotoxicity in rats. Int. J. Pharm. Pharm. Sci. 2015, 7, 65–71. [Google Scholar]
- Dereski, M.O.; Chopp, M.; Knight, R.A.; Rodolosi, L.C.; Garcia, J.H. The heterogeneous temporal evolution of focal ischemic neuronal damage in the rat. Acta Neuropathol. 1993, 85, 327–333. [Google Scholar] [CrossRef]
- Kirino, T.; Tamura, A.; Sano, K. Delayed neuronal death in the rat hippocampus following transient forebrain ischemia. Acta Neuropathol. 1984, 64, 139–147. [Google Scholar] [CrossRef]
- Ogaly, H.A.; Alsherbiny, M.A.; El Badawy, S.A.; Abd-Elsalam, R.M.; Li, C.G.; Azouz, A.A. Gastroprotective effects and metabolomic profiling of Chasteberry fruits against indomethacin-induced gastric injury in rats. J. Funct. Foods 2021, 86, 104732. [Google Scholar] [CrossRef]
- Soliman, G.A.; Abdel-Rahman, R.F.; Ogaly, H.A.; Althurwi, H.N.; Abd-Elsalam, R.M.; Albaqami, F.F.; Abdel-Kader, M.S. Momordica charantia extract protects against diabetes-related spermatogenic dysfunction in male rats: Molecular and biochemical study. Molecules 2020, 25, 5255. [Google Scholar] [CrossRef]
- Sun, X.; Guo, S.; Wang, Y.; Fu, B.; Liu, J.; Zhang, Y.; Yang, R.; Li, C.; Gao, J.; Gao, L. Neuroprotective effect of Longshengzhi capsule following permanent middle cerebral artery occlusion in rats. J. Tradit. Chin. Med. Sci. 2020, 7, 12–19. [Google Scholar] [CrossRef]
- Fukuda, A.M.; Badaut, J. Aquaporin 4: A player in cerebral edema and neuroinflammation. J. Neuroinflammation 2012, 9, 279. [Google Scholar] [CrossRef]
- Sommer, C.J. Ischemic stroke: Experimental models and reality. Acta Neuropathol. 2017, 133, 245–261. [Google Scholar] [CrossRef]
- Grober, E.; Hall, C.B.; Lipton, R.B.; Zonderman, A.B.; Resnick, S.M.; Kawas, C. Memory impairment, executive dysfunction, and intellectual decline in preclinical Alzheimer’s disease. J. Int. Neuropsychol. Soc. 2008, 14, 266–278. [Google Scholar] [CrossRef]
- El-Marasy, S.A.; Abdel-Rahman, R.F.; Abd-Elsalam, R.M. Neuroprotective effect of vildagliptin against cerebral ischemia in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 1133–1145. [Google Scholar] [CrossRef]
- Chen, P.; Li, L.; Gao, Y.; Xie, Z.; Zhang, Y.; Pan, Z.; Tu, Y.; Wang, H.; Han, Q.; Hu, X. β-carotene provides neuroprotection after experimental traumatic brain injury via the Nrf2-ARE pathway. J. Integr. Neurosci. 2019, 18, 153–161. [Google Scholar]
- Hattori, K.; Lee, H.; Hurn, P.D.; Crain, B.J.; Traystman, R.J.; DeVries, A.C. Cognitive deficits after focal cerebral ischemia in mice. Stroke 2000, 31, 1939–1944. [Google Scholar] [CrossRef]
- Zhao, Q.; Cheng, X.; Wang, X.; Wang, J.; Zhu, Y.; Ma, X. Neuroprotective effect and mechanism of Mu-Xiang-You-Fang on cerebral ischemia-reperfusion injury in rats. J. Ethnopharmacol. 2016, 192, 140–147. [Google Scholar] [CrossRef]
- Hira, S.; Saleem, U.; Anwar, F.; Sohail, M.F.; Raza, Z.; Ahmad, B. β-Carotene: A Natural Compound Improves Cognitive Impairment and Oxidative Stress in a Mouse Model of Streptozotocin-Induced Alzheimer’s Disease. Biomolecules 2019, 9, 441. [Google Scholar] [CrossRef]
- Granger, D.N.; Kvietys, P.R. Reperfusion injury and reactive oxygen species: The evolution of a concept. Redox Biol. 2015, 6, 524–551. [Google Scholar] [CrossRef]
- Bahonar, A.; Saadatnia, M.; Khorvash, F.; Maracy, M.; Khosravi, A. Carotenoids as Potential Antioxidant Agents in Stroke Prevention: A Systematic Review. Int. J. Prev. Med. 2017, 8, 70. [Google Scholar]
- Cojocaru, I.M.; Cojocaru, M.; Iliescu, I.; Botnaru, L.; Gurban, C.V.; Sfrijan, F.; Tănăsescu, R. Plasma myeloperoxidase levels in patients with acute ischemic stroke. Rom. J. Intern. Med. 2010, 48, 101–104. [Google Scholar] [PubMed]
- Kong, L.L.; Wang, Z.Y.; Han, N.; Zhuang, X.M.; Wang, Z.Z.; Li, H.; Chen, N.H. Neutralization of chemokine-like factor 1, a novel C-C chemokine, protects against focal cerebral ischemia by inhibiting neutrophil infiltration via MAPK pathways in rats. J. Neuroinflammation 2014, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, H.; Du, Q.; Shen, J. Targeting Myeloperoxidase (MPO) Mediated Oxidative Stress and Inflammation for Reducing Brain Ischemia Injury: Potential Application of Natural Compounds. Front. Physiol. 2020, 11, 433. [Google Scholar] [CrossRef]
- Jover-Mengual, T.; Hwang, J.-Y.; Byun, H.-R.; Court-Vazquez, B.L.; Centeno, J.M.; Burguete, M.C.; Zukin, R.S. The Role of NF-κB Triggered Inflammation in Cerebral Ischemia. Front. Cell. Neurosci. 2021, 15, 633610. [Google Scholar] [CrossRef]
- Snow, W.M.; Albensi, B.C. Neuronal Gene Targets of NF-κB and Their Dysregulation in Alzheimer’s Disease. Front. Mol. Neurosci. 2016, 9, 118. [Google Scholar] [CrossRef]
- Shaik, S.M.; Mp, E.; Al Omar, S.Y.; Mohammad, A.; Kodidhela, L.D. Combined cardio-protective ability of syringic acid and resveratrol against isoproterenol induced cardio-toxicity in rats via attenuating NF-kB and TNF-α pathways. Sci. Rep. 2020, 10, 3426. [Google Scholar]
- Dresselhaus, E.C.; Meffert, M.K. Cellular specificity of NF-κB function in the nervous system. Front. Immunol. 2019, 10, 1043. [Google Scholar] [CrossRef]
- Harari, O.A.; Liao, J.K. NF-κB and innate immunity in ischemic stroke. Ann. N. Y. Acad. Sci. 2010, 1207, 32–40. [Google Scholar] [CrossRef]
- Xu, D.; Xia, N.; Hou, K.; Li, F.; Chen, S.; Hu, Y.; Fang, W.; Li, Y. Clematichinenoside Facilitates Recovery of Neurological and Motor Function in Rats after Cerebral Ischemic Injury through Inhibiting Notch/NF-κB Pathway. J. Stroke Cerebrovasc. Dis. 2019, 28, 104288. [Google Scholar] [CrossRef]
- Yuan, Y.; Men, W.; Shan, X.; Zhai, H.; Qiao, X.; Geng, L.; Li, C. Baicalein exerts neuroprotective effect against ischaemic/reperfusion injury via alteration of NF-kB and LOX and AMPK/Nrf2 pathway. Inflammopharmacology 2020, 28, 1327–1341. [Google Scholar] [CrossRef]
- Yao, Y.; Chen, L.; Xiao, J.; Wang, C.; Jiang, W.; Zhang, R.; Hao, J. Chrysin protects against focal cerebral ischemia/reperfusion injury in mice through attenuation of oxidative stress and inflammation. Int. J. Mol. Sci. 2014, 15, 20913–20926. [Google Scholar] [CrossRef]
- Cho, S.O.; Kim, M.-H.; Kim, H. β-Carotene Inhibits Activation of NF-κB, Activator Protein-1, and STAT3 and Regulates Abnormal Expression of Some Adipokines in 3T3-L1 Adipocytes. J. Cancer Prev. 2018, 23, 37–43. [Google Scholar] [CrossRef]
- Le, D.A.; Wu, Y.; Huang, Z.; Matsushita, K.; Plesnila, N.; Augustinack, J.C.; Hyman, B.T.; Yuan, J.; Kuida, K.; Flavell, R.A.; et al. Caspase activation and neuroprotection in caspase-3-deficient mice after in vivo cerebral ischemia and in vitro oxygen glucose deprivation. Proc. Natl. Acad. Sci. USA 2002, 99, 15188–15193. [Google Scholar] [CrossRef]
- Shen, H.; Kuo, C.-C.; Chou, J.; Delvolve, A.; Jackson, S.N.; Post, J.; Woods, A.S.; Hoffer, B.J.; Wang, Y.; Harvey, B.K. Astaxanthin reduces ischemic brain injury in adult rats. FASEB J. 2009, 23, 1958–1968. [Google Scholar] [CrossRef]
- Aboutaleb, N.; Shamsaei, N.; Rajabi, H.; Khaksari, M.; Erfani, S.; Nikbakht, F.; Motamedi, P.; Shahbazi, A. Protection of Hippocampal CA1 Neurons Against Ischemia/Reperfusion Injury by Exercise Preconditioning via Modulation of Bax/Bcl-2 Ratio and Prevention of Caspase-3 Activation. Basic Clin. Neurosci. 2016, 7, 21–29. [Google Scholar]
- Wagner, D.-C.; Riegelsberger, U.M.; Michalk, S.; Härtig, W.; Kranz, A.; Boltze, J. Cleaved caspase-3 expression after experimental stroke exhibits different phenotypes and is predominantly non-apoptotic. Brain Res. 2011, 1381, 237–242. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, X.; Chen, A.; Cheng, X.; Zhang, G.; Sun, J.; Zhao, Y.; Huang, Y.; Zhu, Y. Rhein protects against cerebral ischemic-/reperfusion-induced oxidative stress and apoptosis in rats. Int. J. Mol. Med. 2018, 41, 2802–2812. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GSH (μmol/g) | SOD (U/g) | MDA (nmol/g) | GPx (U/g) | CAT (U/g) | |
|---|---|---|---|---|---|
| Sham | 3.3 ± 0.11 b | 87.4 ± 0.3 b | 14.0 ± 0.89 b | 302.0 ± 19.73 b | 2.79 ± 0.061 b |
| I/R control | 1.7 ± 0.07 a | 52.5 ± 2.9 a | 30.6 ± 2.12 a | 150.3 ± 8.43 a | 0.58 ± 0.026 a |
| βCAR-10 | 2.7 ± 0.09 ab | 65.8 ± 4.06 ab | 19.8 ± 1.42 ab | 230.4 ± 7.61 ab | 1.91 ± 0.023 ab |
| βCAR-20 | 3.1 ± 0.13 b | 69.2 ± 2.12 ab | 16.0 ± 1.07 b | 245.9 ± 11.8 ab | 2.31 ± 0.068 ab |
| Bax/Bcl2 Ratio | |
|---|---|
| Sham | 1 |
| I/R control | 9.07 a |
| βCAR-10 | 1.97 b |
| βCAR-20 | 0.95 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Althurwi, H.N.; Abdel-Rahman, R.F.; Soliman, G.A.; Ogaly, H.A.; Alkholifi, F.K.; Abd-Elsalam, R.M.; Alqasoumi, S.I.; Abdel-Kader, M.S. Protective Effect of Beta-Carotene against Myeloperoxidase- Mediated Oxidative Stress and Inflammation in Rat Ischemic Brain Injury. Antioxidants 2022, 11, 2344. https://doi.org/10.3390/antiox11122344
Althurwi HN, Abdel-Rahman RF, Soliman GA, Ogaly HA, Alkholifi FK, Abd-Elsalam RM, Alqasoumi SI, Abdel-Kader MS. Protective Effect of Beta-Carotene against Myeloperoxidase- Mediated Oxidative Stress and Inflammation in Rat Ischemic Brain Injury. Antioxidants. 2022; 11(12):2344. https://doi.org/10.3390/antiox11122344
Chicago/Turabian StyleAlthurwi, Hassan N., Rehab F. Abdel-Rahman, Gamal A. Soliman, Hanan A. Ogaly, Faisal K. Alkholifi, Reham M. Abd-Elsalam, Saleh I. Alqasoumi, and Maged S. Abdel-Kader. 2022. "Protective Effect of Beta-Carotene against Myeloperoxidase- Mediated Oxidative Stress and Inflammation in Rat Ischemic Brain Injury" Antioxidants 11, no. 12: 2344. https://doi.org/10.3390/antiox11122344
APA StyleAlthurwi, H. N., Abdel-Rahman, R. F., Soliman, G. A., Ogaly, H. A., Alkholifi, F. K., Abd-Elsalam, R. M., Alqasoumi, S. I., & Abdel-Kader, M. S. (2022). Protective Effect of Beta-Carotene against Myeloperoxidase- Mediated Oxidative Stress and Inflammation in Rat Ischemic Brain Injury. Antioxidants, 11(12), 2344. https://doi.org/10.3390/antiox11122344