
A Chlorophyll-Derived Phylloxanthobilin Is a Potent Antioxidant That Modulates Immunometabolism in Human PBMC

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. General
2.2. Plant Material
2.3. Chromatography
2.4. Spectroscopy
2.5. Isolation and Preparation of Phyllobilins
2.6. Cell Culture and Isolation of PBMC
2.7. Stability of PxB in Medium
2.8. Oxygen Radical Absorbance Capacity (ORAC) Assay
2.9. Intracellular ROS Measurement
2.10. PBMC Treatments and Metabolite Measurements
2.11. Statistical Analysis
3. Results
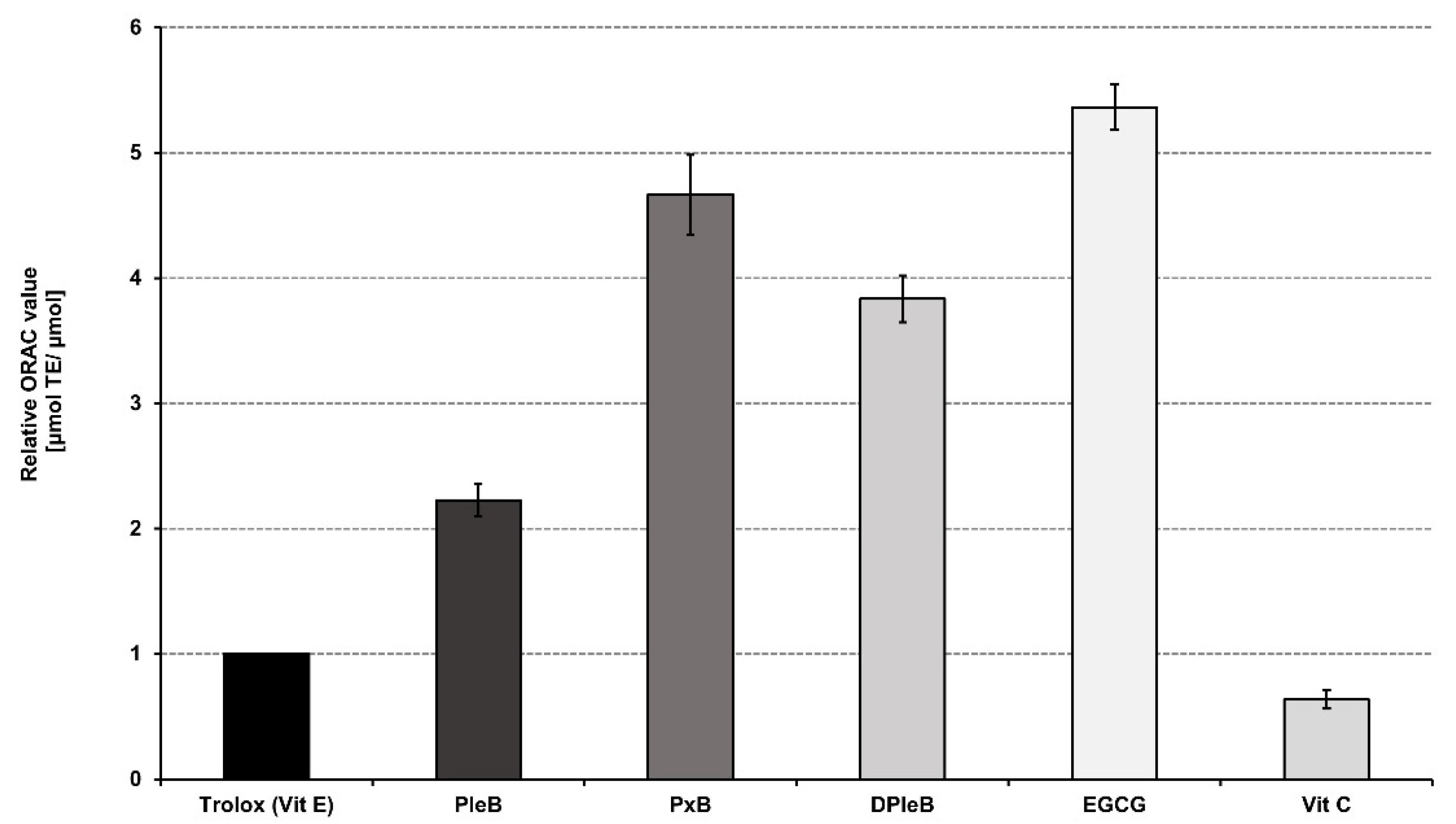
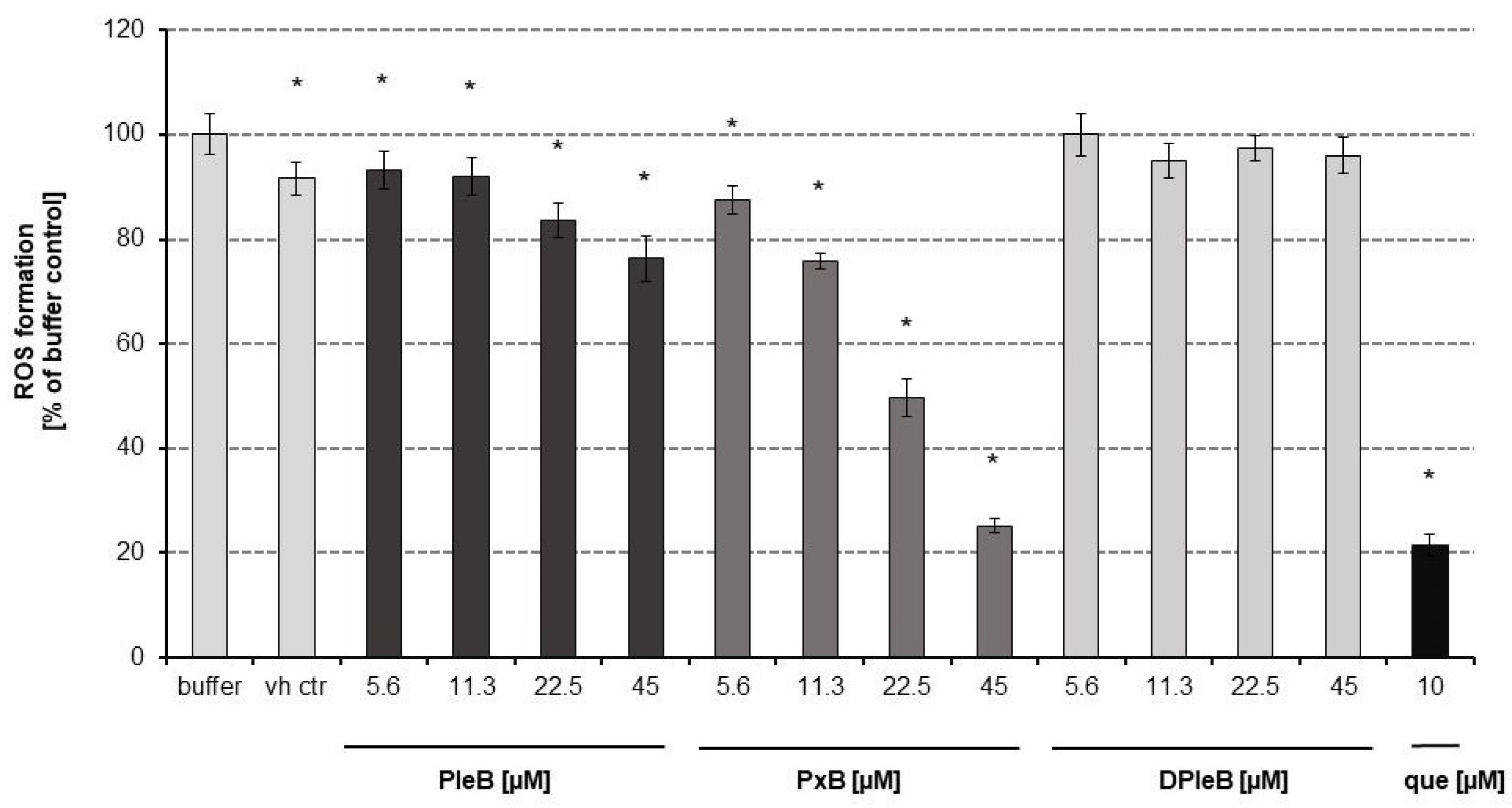
3.1. Antioxidative Properties of Phyllobilins
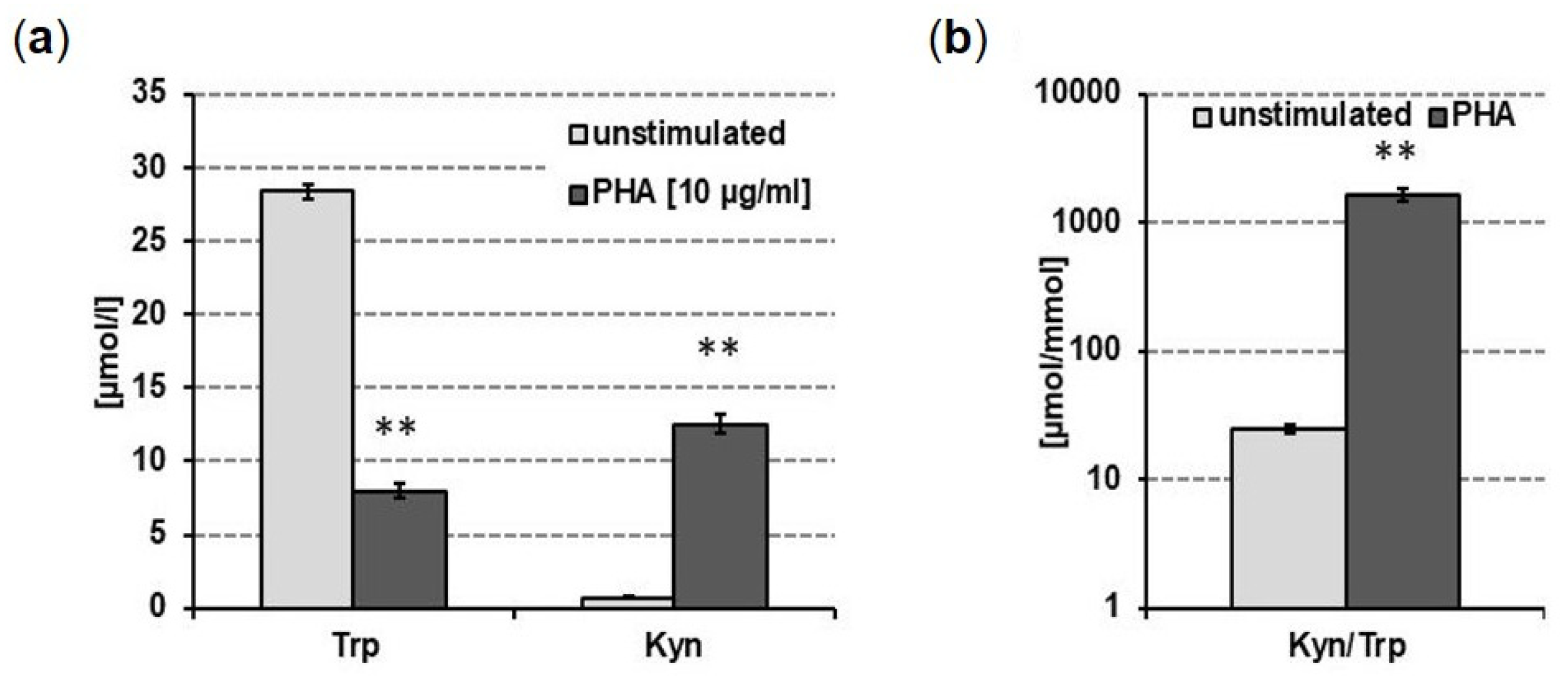
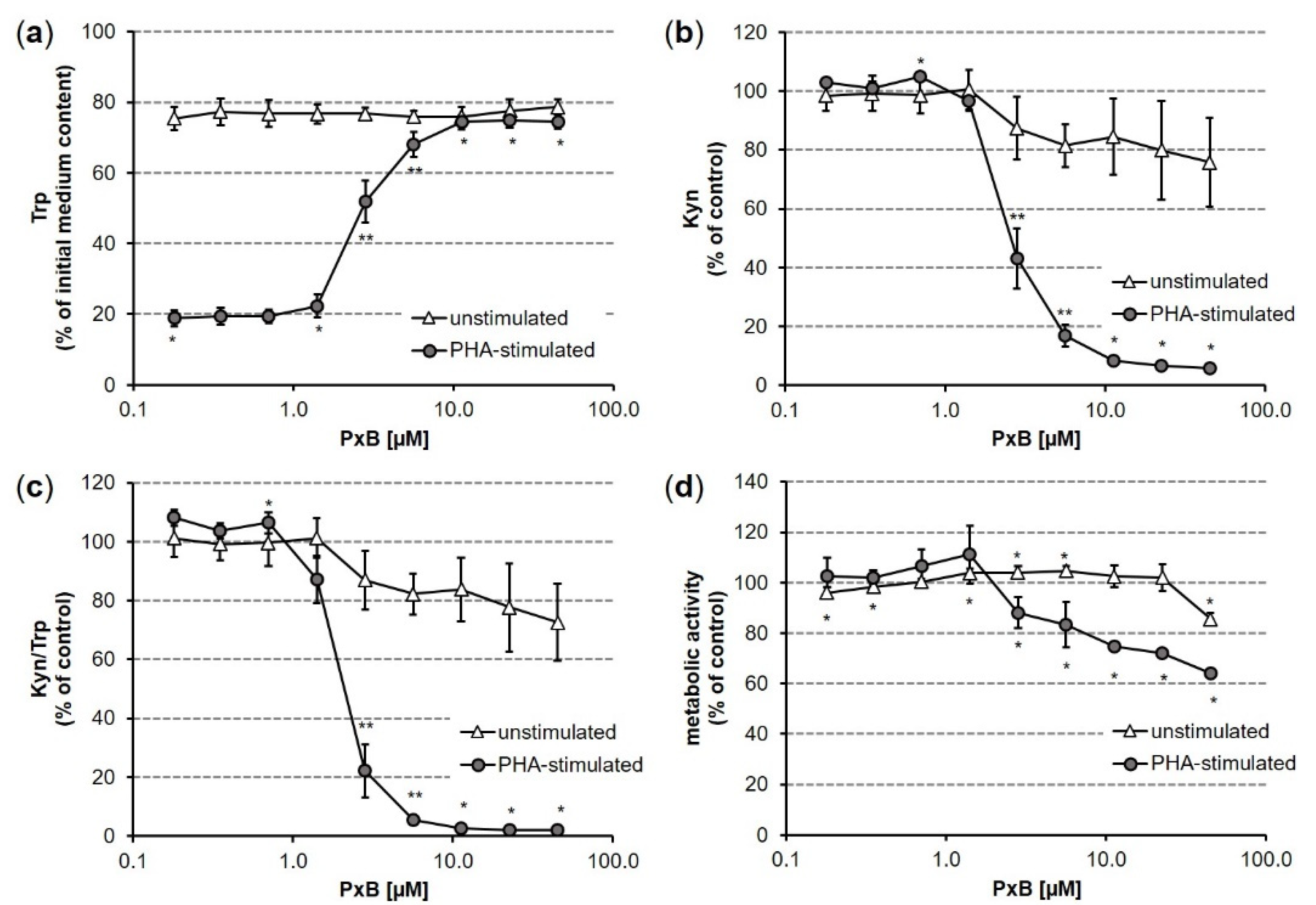
3.2. Impact of PxB on Tryptophan Breakdown in Human PBMC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmidt, B.M.; Ribnicky, D.M.; Lipsky, P.E.; Raskin, I. Revisiting the ancient concept of botanical therapeutics. Nat. Chem. Biol. 2007, 3, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Hendry, G.A.F.; Houghton, J.D.; Brown, S.B. The Degradation of Chlorophyll—A Biological Enigma. New Phytol. 1987, 107, 255–302. [Google Scholar] [CrossRef]
- Mur, L.A.J.; Ougham, H.; Thomas, H. Chlorophyll Metabolism and Its Role in Responses and Adaptations to Stress. In Annual Plant Reviews Online; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2021; pp. 713–732. [Google Scholar]
- Kräutler, B. Phyllobilins—The Abundant Tetrapyrrolic Catabolites of the Green Plant Pigment Chlorophyll. Chem. Soc. Rev. 2014, 43, 6227–6238. [Google Scholar] [CrossRef] [PubMed]
- Kräutler, B.; Jaun, B.; Bortlik, K.; Schellenberg, M.; Matile, P. On the Enigma of Chlorophyll Degradation—The Constitution of a Secoporphinoid Catabolite. Angew. Chem. Int. Ed. Engl. 1991, 30, 1315–1318. [Google Scholar] [CrossRef]
- Kräutler, B.; Matile, P. Solving the riddle of chlorophyll breakdown. Acc. Chem. Res. 1999, 32, 35–43. [Google Scholar] [CrossRef]
- Hörtensteiner, S.; Kräutler, B. Chlorophyll breakdown in higher plants. Biochim. Biophys. Acta Bioenerget. 2011, 1807, 977–988. [Google Scholar] [CrossRef]
- Kuai, B.; Chen, J.; Hörtensteiner, S. The biochemistry and molecular biology of chlorophyll breakdown. J. Exp. Bot. 2017, 69, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Aubry, S.; Christ, B.; Kräutler, B.; Martinoia, E.; Thomas, H.; Zipfel, C. An evergreen mind and a heart for the colors of fall. J. Exp. Bot. 2021, 72, 4625–4633. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Hörtensteiner, S. Mechanism and Significance of Chlorophyll Breakdown. J. Plant Growth Regul. 2014, 33, 4–20. [Google Scholar] [CrossRef]
- Kräutler, B.; Hörtensteiner, S. Chlorophyll Breakdown: Chemistry, Biochemistry, and Biology. In Handbook of Porphyrin Science; World Scientific Publishing Company: Singapore, 2012; Volume 28, pp. 117–185. [Google Scholar]
- Mühlecker, W.; Ongania, P.-D.D.K.-H.; Kräutler, B.; Matile, P.; Hörtensteiner, S. Tracking Down Chlorophyll Breakdown in Plants: Elucidation of the Constitution of a “Fluorescent” Chlorophyll Catabolite. Angew. Chem. Int. Ed. 1997, 36, 401–404. [Google Scholar] [CrossRef]
- Kräutler, B. Breakdown of Chlorophyll in Higher Plants—Phyllobilins as Abundant, Yet Hardly Visible Signs of Ripening, Senescence and Cell Death. Angew. Chem. Int. Ed. 2016, 55, 4882–4907. [Google Scholar] [CrossRef] [PubMed]
- Matile, P.; Ginsburg, S.; Schellenberg, M.; Thomas, H. Catabolites of chlorophyll in senescing barley leaves are localized in the vacuoles of mesophyll cells. Proc. Natl. Acad. Sci. USA 1988, 85, 9529–9532. [Google Scholar] [CrossRef] [PubMed]
- Oberhuber, M.; Berghold, J.; Breuker, K.; Hörtensteiner, S.; Kraütler, B. Breakdown of chlorophyll: A nonenzymatic reaction accounts for the formation of the colorless “nonfluorescent” chlorophyll catabolites. Proc. Natl. Acad. Sci. USA 2003, 100, 6910–6915. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Süssenbacher, I.; Moser, S.; Bichsel, N.; Egert, A.; Müller, T.; Kräutler, B.; Hörtensteiner, S. Cytochrome P450 CYP89A9 is involved in the formation of major chlorophyll catabolites during leaf senescence in Arabidopsis thaliana. Plant Cell 2013, 25, 1868–1880. [Google Scholar] [CrossRef]
- Süssenbacher, I.; Hörtensteiner, S.; Kräutler, B. A Dioxobilin-Type Fluorescent Chlorophyll Catabolite as a Transient Early Intermediate of the Dioxobilin-Branch of Chlorophyll Breakdown in Arabidopsis thaliana. Angew. Chem. Int. Ed. 2015, 54, 13777–13781. [Google Scholar] [CrossRef]
- Christ, B.; Hauenstein, M.; Hörtensteiner, S. A liquid chromatography-mass spectrometry platform for the analysis of phyllobilins, the major degradation products of chlorophyll in Arabidopsis thaliana. Plant J. 2016, 88, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Süssenbacher, I.; Christ, B.; Hörtensteiner, S.; Kräutler, B. Hydroxymethylated phyllobilins: A puzzling new feature of the dioxobilin branch of chlorophyll breakdown. Chem. Eur. J. 2014, 20, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Erhart, T.; Mittelberger, C.; Liu, X.; Podewitz, M.; Li, C.; Scherzer, G.; Stoll, G.; Valls, J.; Robatscher, P.; Liedl, K.R.; et al. Novel Types of Hypermodified Fluorescent Phyllobilins from Breakdown of Chlorophyll in Senescent Leaves of Grapevine (Vitis vinifera). Chem. Eur. J. 2018, 24, 17268–17279. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Rafelsberger, M.; Vergeiner, C.; Kräutler, B. A Dioxobilane as Product of a Divergent Path of Chlorophyll Breakdown in Norway Maple. Angew. Chem. Int. Ed. 2011, 50, 10724–10727. [Google Scholar] [CrossRef]
- Moser, S.; Ulrich, M.; Müller, T.; Kräutler, B. A yellow chlorophyll catabolite is a pigment of the fall colours. Photochem. Photobiol. Sci. 2008, 7, 1577–1581. [Google Scholar] [CrossRef]
- Scherl, M.; Müller, T.; Kräutler, B. Chlorophyll Catabolites in Senescent Leaves of the Lime Tree (Tilia cordata). Chem. Biodivers. 2012, 9, 2605–2617. [Google Scholar] [CrossRef] [PubMed]
- Wakana, D.; Kato, H.; Momose, T.; Sasaki, N.; Ozeki, Y.; Goda, Y. NMR-based characterization of a novel yellow chlorophyll catabolite, Ed-YCC, isolated from Egeria densa. Tetrahedron Lett. 2014, 55, 2982–2985. [Google Scholar] [CrossRef]
- Scherl, M.; Müller, T.; Kreutz, C.; Huber, R.G.; Zass, E.; Liedl, K.R.; Kräutler, B. Chlorophyll Catabolites in Fall Leaves of the Wych Elm Tree Present a Novel Glycosylation Motif. Chem. Eur. J. 2016, 22, 9498–9503. [Google Scholar] [CrossRef] [PubMed]
- Karg, C.A.; Wang, P.; Vollmar, A.M.; Moser, S. Re-opening the stage for Echinacea research—Characterization of phylloxanthobilins as a novel anti-oxidative compound class in Echinacea purpurea. Phytomedicine 2019, 60, 152969. [Google Scholar] [CrossRef] [PubMed]
- Berghold, J.; Breuker, K.; Oberhuber, M.; Hörtensteiner, S.; Kräutler, B. Chlorophyll breakdown in spinach: On the structure of five nonfluorescent chlorophyll catabolites. Photosynth. Res. 2002, 74, 109–119. [Google Scholar] [CrossRef]
- Kräutler, B.; Banala, S.; Moser, S.; Vergeiner, C.; Müller, T.; Lütz, C.; Holzinger, A. A novel blue fluorescent chlorophyll catabolite accumulates in senescent leaves of the peace lily and indicates a split path of chlorophyll breakdown. FEBS Lett. 2010, 584, 4215–4221. [Google Scholar] [CrossRef]
- Karg, C.A.; Wang, P.; Kluibenschedl, F.; Müller, T.; Allmendinger, L.; Vollmar, A.M.; Moser, S. Phylloxanthobilins are Abundant Linear Tetrapyrroles from Chlorophyll Breakdown with Activities Against Cancer Cells. Eur. J. Org. Chem. 2020, 2020, 4499–4509. [Google Scholar] [CrossRef]
- Schumacher, I.; Menghini, D.; Ovinnikov, S.; Hauenstein, M.; Fankhauser, N.; Zipfel, C.; Hörtensteiner, S.; Aubry, S. Evolution of chlorophyll degradation is associated with plant transition to land. Plant J. Cell Mol. Biol. 2022, 109, 1473–1488. [Google Scholar] [CrossRef] [PubMed]
- Hauenstein, M.; Hörtensteiner, S.; Aubry, S. Side-chain modifications of phyllobilins may not be essential for chlorophyll degradation in Arabidopsis. Plant Direct 2022, 6, e441. [Google Scholar] [CrossRef] [PubMed]
- Gostner, J.M.; Becker, K.; Fuchs, D.; Sucher, R. Redox regulation of the immune response. Redox Rep. 2013, 18, 88–94. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Talalay, P. Direct and indirect antioxidant properties of inducers of cytoprotective proteins. Mol. Nutr. Food Res. 2008, 52, S128–S138. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Schwaiger, S.; Waltenberger, B.; Fuchs, D.; Pezzei, C.K.; Schennach, H.; Stuppner, H.; Gostner, J.M. Immunomodulatory Effects of Diterpene Quinone Derivatives from the Roots of Horminum pyrenaicum in Human PBMC. Oxidative Med. Cell. Longev. 2018, 2018, 2980295. [Google Scholar] [CrossRef]
- Jenny, M.; Klieber, M.; Zaknun, D.; Schroecksnadel, S.; Kurz, K.; Ledochowski, M.; Schennach, H.; Fuchs, D. In vitro testing for anti-inflammatory properties of compounds employing peripheral blood mononuclear cells freshly isolated from healthy donors. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2011, 60, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.C. Tryptophan nutrition and metabolism: An overview. Adv. Exp. Med. Biol. 1991, 294, 345–358. [Google Scholar] [CrossRef]
- Schroecksnadel, K.; Wirleitner, B.; Winkler, C.; Fuchs, D. Monitoring tryptophan metabolism in chronic immune activation. Clin. Chim. Acta Int. J. Clin. Chem. 2006, 364, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Bass, D.A.; Parce, J.W.; Dechatelet, L.R.; Szejda, P.; Seeds, M.C.; Thomas, M. Flow Cytometric Studies of Oxidative Product Formation by Neutrophils: A Graded Response to Membrane Stimulation. J. Immunol. 2016, 197, 683–690. [Google Scholar] [PubMed]
- Vergeiner, C.; Ulrich, M.; Li, C.; Liu, X.; Müller, T.; Kräutler, B. Stereo- and Regioselective Phyllobilane Oxidation in Leaf Homogenates of the Peace Lily (Spathiphyllum wallisii)—Hypothetical Endogenous Path to Yellow Chlorophyll Catabolites. Chem. Eur. J. 2015, 21, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Wurst, K.; Jockusch, S.; Gruber, K.; Podewitz, M.; Liedl, K.R.; Kräutler, B. Chlorophyll-Derived Yellow Phyllobilins of Higher Plants as Medium-Responsive Chiral Photoswitches. Angew. Chem. Int. Ed. 2016, 55, 15760–15765. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Gostner, J.M.; Schroecksnadel, S.; Jenny, M.; Klein, A.; Ueberall, F.; Schennach, H.; Fuchs, D. Coffee extracts suppress tryptophan breakdown in mitogen-stimulated peripheral blood mononuclear cells. J. Am. Coll. Nutr. 2015, 34, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Hofer, S.; Stonig, M.; Wally, V.; Hartmann, A.; Fuchs, D.; Hermann, M.; Paparella, M.; Ganzera, M.; Gostner, J.M. Contradictory effects of chemical filters in UV/ROS-stressed human keratinocyte and fibroblast cells. ALTEX 2019, 36, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Widner, B.; Werner, E.R.; Schennach, H.; Wachter, H.; Fuchs, D. Simultaneous measurement of serum tryptophan and kynurenine by HPLC. Clin. Chem. 1997, 43, 2424–2426. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, D.; Möller, A.A.; Reibnegger, G.; Stöckle, E.; Werner, E.R.; Wachter, H. Decreased serum tryptophan in patients with HIV-1 infection correlates with increased serum neopterin and with neurologic/psychiatric symptoms. J. Acquir. Immune Defic. Syndr. 1990, 3, 873–876. [Google Scholar] [PubMed]
- Katial, R.K.; Sachanandani, D.; Pinney, C.; Lieberman, M.M. Cytokine production in cell culture by peripheral blood mononuclear cells from immunocompetent hosts. Clin. Diagn. Lab. Immunol. 1998, 5, 78–81. [Google Scholar] [CrossRef]
- Nathan, C.F.; Murray, H.W.; Wiebe, M.E.; Rubin, B.Y. Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J. Exp. Med. 1983, 158, 670–689. [Google Scholar] [CrossRef]
- Schroecksnadel, K.; Winkler, C.; Werner, E.R.; Sarcletti, M.; Romani, N.; Ebner, S.; Schennach, H.; Fuchs, D.; Zangerle, R. Interferon-γ-mediated pathways and in vitro PBMC proliferation in HIV-infected patients. Biol. Chem. 2009, 390, 115–123. [Google Scholar] [CrossRef]
- Hörtensteiner, S.; Hauenstein, M.; Kräutler, B. Chlorophyll breakdown—Regulation, biochemistry and phyllobilins as its products. In Advances in Botanical Research; Grimm, B., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 90, pp. 213–271. [Google Scholar]
- Pruzinská, A.; Anders, I.; Aubry, S.; Schenk, N.; Tapernoux-Lüthi, E.; Müller, T.; Kräutler, B.; Hörtensteiner, S. In vivo participation of red chlorophyll catabolite reductase in chlorophyll breakdown. Plant Cell 2007, 19, 369–387. [Google Scholar] [CrossRef]
- Moser, S.; Erhart, T.; Neuhauser, S.; Kräutler, B. Phyllobilins from Senescence-Associated Chlorophyll Breakdown in the Leaves of Basil (Ocimum basilicum) Show Increased Abundance upon Herbivore Attack. J. Agric. Food Chem. 2020, 68, 7132–7142. [Google Scholar] [CrossRef]
- Mittelberger, C.; Yalcinkaya, H.; Pichler, C.; Gasser, J.; Scherzer, G.; Erhart, T.; Schumacher, S.; Holzner, B.; Janik, K.; Robatscher, P.; et al. Pathogen-Induced Leaf Chlorosis: Products of Chlorophyll Breakdown Found in Degreened Leaves of Phytoplasma-Infected Apple (Malus x domestica Borkh.) and Apricot (Prunus armeniaca L.) Trees Relate to the Pheophorbide a Oxygenase/Phyllobilin Pathway. J. Agric. Food Chem. 2017, 65, 2651–2660. [Google Scholar] [CrossRef]
- McArdle, P.F.; Whitcomb, B.W.; Tanner, K.; Mitchell, B.D.; Shuldiner, A.R.; Parsa, A. Association between bilirubin and cardiovascular disease risk factors: Using Mendelian randomization to assess causal inference. BMC Cardiovasc. Disord. 2012, 12, 16. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; Zheng, J.; Kobayashi, K.; Takayanagi, R. Bilirubin and biliverdin protect rodents against diabetic nephropathy by downregulating NAD(P)H oxidase. Kidney Int. 2010, 78, 905–919. [Google Scholar] [CrossRef]
- Zheng, J.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; McCarty, M.F.; Fujii, M.; Ikeda, N.; Kobayashi, K.; Sonoda, N.; Takayanagi, R. Phycocyanin and phycocyanobilin from Spirulina platensis protect against diabetic nephropathy by inhibiting oxidative stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 304, R110–R120. [Google Scholar] [CrossRef]
- Drummond, G.R.; Selemidis, S.; Griendling, K.K.; Sobey, C.G. Combating oxidative stress in vascular disease: NADPH oxidases as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 453–471. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.W.; Wang, J.; Zhang, Q.; Wang, R.; Dhandapani, K.M.; Vadlamudi, R.K.; Brann, D.W. NADPH oxidase in brain injury and neurodegenerative disorders. Mol. Neurodegener. 2017, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Paik, Y.H.; Kim, J.; Aoyama, T.; De Minicis, S.; Bataller, R.; Brenner, D.A. Role of NADPH oxidases in liver fibrosis. Antioxid. Redox Signal. 2014, 20, 2854–2872. [Google Scholar] [CrossRef]
- Müller, T.; Ulrich, M.; Ongania, K.-H.; Kräutler, B. Colourless tetrapyrrolic chlorophyll catabolites found in ripening fruit are effective antioxidants. Angew. Chem. Int. Ed. 2007, 46, 8699–8702. [Google Scholar] [CrossRef]
- Moser, S.; Kräutler, B. In Search of Bioactivity—Phyllobilins, an Unexplored Class of Abundant Heterocyclic Plant Metabolites from Breakdown of Chlorophyll. Isr. J. Chem. 2019, 59, 420–431. [Google Scholar] [CrossRef]
- Karg, C.A.; Wang, S.; Al Danaf, N.; Pemberton, R.P.; Bernard, D.; Kretschmer, M.; Schneider, S.; Zisis, T.; Vollmar, A.M.; Lamb, D.C.; et al. Tetrapyrrolic Pigments from Heme- and Chlorophyll Breakdown are Actin-Targeting Compounds. Angew. Chem. Int. Ed. 2021, 60, 22578–22584. [Google Scholar] [CrossRef]
- Karg, C.A.; Schilling, C.M.; Allmendinger, L.; Moser, S. Isolation, characterization, and antioxidative activity of a dioxobilin-type phylloxanthobilin from savoy cabbage. J. Porphyr. Phthalocyanines 2019, 23, 881–888. [Google Scholar] [CrossRef]
- Schmidt, B.; Ribnicky, D.M.; Poulev, A.; Logendra, S.; Cefalu, W.T.; Raskin, I. A natural history of botanical therapeutics. Metab. Clin. Exp. 2008, 57, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Kräutler, B. Transition Metal Complexes of Phyllobilins—A New Realm of Bioinorganic Chemistry. Dalton Trans. 2015, 44, 10116–10127. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karg, C.A.; Parráková, L.; Fuchs, D.; Schennach, H.; Kräutler, B.; Moser, S.; Gostner, J.M. A Chlorophyll-Derived Phylloxanthobilin Is a Potent Antioxidant That Modulates Immunometabolism in Human PBMC. Antioxidants 2022, 11, 2056. https://doi.org/10.3390/antiox11102056
Karg CA, Parráková L, Fuchs D, Schennach H, Kräutler B, Moser S, Gostner JM. A Chlorophyll-Derived Phylloxanthobilin Is a Potent Antioxidant That Modulates Immunometabolism in Human PBMC. Antioxidants. 2022; 11(10):2056. https://doi.org/10.3390/antiox11102056
Chicago/Turabian StyleKarg, Cornelia A., Lucia Parráková, Dietmar Fuchs, Harald Schennach, Bernhard Kräutler, Simone Moser, and Johanna M. Gostner. 2022. "A Chlorophyll-Derived Phylloxanthobilin Is a Potent Antioxidant That Modulates Immunometabolism in Human PBMC" Antioxidants 11, no. 10: 2056. https://doi.org/10.3390/antiox11102056
APA StyleKarg, C. A., Parráková, L., Fuchs, D., Schennach, H., Kräutler, B., Moser, S., & Gostner, J. M. (2022). A Chlorophyll-Derived Phylloxanthobilin Is a Potent Antioxidant That Modulates Immunometabolism in Human PBMC. Antioxidants, 11(10), 2056. https://doi.org/10.3390/antiox11102056