Abstract
Oats (Avena sativa L.) are rich in protein, fiber, calcium, vitamins (B, C, E, and K), amino acids, and antioxidants (beta-carotene, polyphenols, chlorophyll, and flavonoids). β-glucan and avenanthramides improve the immune system, eliminate harmful substances from the body, reduce blood cholesterol, and help with dietary weight loss by enhancing the lipid profile and breaking down fat in the body. β-glucan regulates insulin secretion, preventing diabetes. Progladins also lower cholesterol levels, suppress the accumulation of triglycerides, reduce blood sugar levels, suppress inflammation, and improve skin health. Saponin-based avanacosidase and functional substances of flavone glycoside improve the immune function, control inflammation, and prevent infiltration in the skin. Moreover, lignin and phytoestrogen prevent hormone-related cancer and improve the quality of life of postmenopausal women. Sprouted oats are rich in saponarin in detoxifying the liver. The literatures have been reviewed and the recent concepts and prospects have been summarized with figures and tables. This review discusses recent trends in research on the functionality of oats rather than their nutritional value with individual immunity for self-medication. The oat and its acting components have been revisited for the future prospect and development of human healthy and functional sources.
1. Introduction
So-called “superfoods” are food products that boost the immunity of the body through their rich contents of nutrients and antioxidants. Representative examples of superfoods include tomatoes, spinach, broccoli, salmon, garlic, and blueberries [1]. Recently, interest in so-called “functional grains”, which are also rich in nutrients and antioxidants, has increased, particularly in the United States of America, Europe, and Japan [2]. These functional grains include oats, quinoa, lentils, chickpeas, amaranth, chia seeds, wild rice, and flaxseeds. Although there are differences in the level of nutrients between each grain, these grains are rich in protein, vitamins, minerals, and dietary fiber compared to the commonly consumed rice (white rice), barley, and wheat. Functional grains help the modern population suffering from “poverty in the midst of plenty” due to busy daily life, irregular meals, and frequent consumption of take-out restaurant meals to ingest abundant nutrients [3,4,5,6,7,8].
Oats (Avena sativa L.) are biennial herbaceous plants that belong to the Poaceae family. They are one of the representative crops that grow in cool and humid weather conditions [9]. Oats have a shape similar to barley and are available in different types, including black, red, yellow, and white oats. They are the sixth most produced grain worldwide, following wheat, corn, rice, barley, and sorghum [10,11]. Traditionally, oats have been considered healthy as they are abundant in protein, fiber, vitamins, and minerals and are mainly consumed as oat meals. In particular, oats contain high levels of proteins and lipids as well as balanced amounts of essential amino acids, such as lysine, and 2–6% of β-glucan and are, therefore, recognized as a high-value crop [12,13,14,15]. Compared to other crops that contain an inversely proportional amount of protein to that of lysine, oats consist of a constant amount of lysine regardless of the protein levels [16]. Globulin accounts for 70–80% of the protein content of oats, and they also contain a low amount of prolamin [16,17]. Oats have 5–12% fat, which is higher than that of other cereal crops [18] (Table 1). Approximately 95% of the fat content in oats are palmitic, oleic, and linoleic acids, and 75–80% are unsaturated fatty acids. These unsaturated fatty acids are associated with various beneficial physiological properties, such as the prevention of dementia and antioxidant activity [10,15,19]. Recently, unsaturated fatty acids were shown to lower blood cholesterol, leading to increased interest in oats as a functional food [20,21,22]. Oats also contain polyphenols including caffeic acids, coumaric acids, gallic acids, hydroxybenzoic acids, protocatechuic acids, syringic acids and vanillic acids as the bioactive compounds [2,9,10]. Alkaloids such as avenanthramides (Avns) are also found in oat [9,10]. In addition, oat bran-derived by-products such as proteins, β-glucan, saponin, albumin, prolamins, and glutelins are also valuable for the nutritional components. Therefore, human consumption of whole oat grain is implicated with health benefits due to the acting components in the whole oat grain. Metabolic diseases have been suggested for the benefits and the whole grains or healthy grains has recently been emphasized through systematic meta-analyses of the available information [23].

Table 1.
Comparison in the functional components in white rice, wheat, oat, and barley [18].
2. Nutritional Benefits of Oats
The nutritional benefits of oats as a food product have been studied in different ways. As shown in Table 1, oats are a good source of high-quality proteins, carbohydrates, dietary fibers such as β-glucan and soluble dietary fibers, fat, minerals, phenolic acids, flavonoids, and antioxidants [13,18,24]. Compared to other grains, proteins in oats are superior in quantity and quality, especially for humans and non-ruminants, based on the composition of essential amino acids. Seed storage proteins of grains are divided into globulins, which are soluble in salt, and prolamins, which are soluble in alcohol [16,25]. Among essential amino acids, lysine, which plays a crucial role in protein biosynthesis, is particularly important as this amino acid cannot be synthesized and must be obtained from the external environment in mammals [26]. Grains are a preferred source of these essential amino acids. In particular, proteins in oats are rich in lysine and have a higher ratio of globulin proteins than other grains that have a high prolamin content [27]. Oats generally contain 3–11% of fat, and several strains contain up to 18% of fat. In addition, most of the fats in oats are stored in the endosperm compared to other grains that store a high fat content in the germini and germinal disk [28]. Fatty acids in oats include oleic (18:1), linoleic (18:2), and linolenic acid (18:3), which are unsaturated fatty acids, and myristic (14:0), palmitic (16:0), and stearic acid (18:0), which are saturated fatty acids (Figure 1) [29,30]. Among these, oats contain high amounts of oleic, linoleic, and palmitic acid. Oleic and linoleic acid are nutritionally essential unsaturated fatty acids, and palmitic acid prevents peroxidation of fat, which causes toxicity and reduces the flavor of grains [31,32]. Thus, oats contain nutritionally excellent fat content and fatty acid composition [33]. However, depending on the use of the grain, it is necessary to evaluate the fat content in more detail. The energy value of fat (37 kJ/g) is much higher than that of proteins or carbohydrates (16–17 kJ/g) [34,35]. Thus, grains with high fat content are generally preferred for animal feeds and not for consumption by humans because they lack flavor and brown excessively during cooking [35,36].
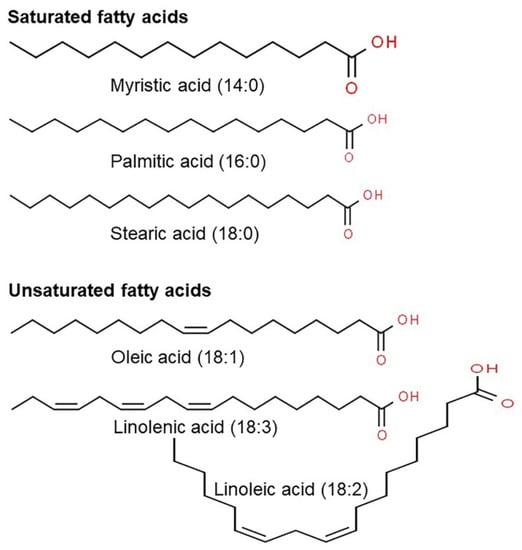
Figure 1.
Representative chemical structure of saturated and unsaturated fatty acids present in oats using ChemSpider.
3. Functionality of β-Glucan in Oats
In 1998, the Food and Agriculture Organization approved the labeling of health effects on food products containing oat extract, and this has attracted attention globally [37]. In addition, oats were selected as one of the world’s top 10 superfoods. Because of the growing interest of consumers in healthy foods, the consumption of oats as an excellent functional food has increased recently [38]. Oats are an important functional ingredient and contain high amounts of dietary fiber and β-glucan. The β-glucan content of oats varies depending on the type and part of the grain (Figure 2A) [39,40]. β-glucans in oats are present as insoluble and soluble dietary fibers. As soluble dietary fibers have superior physiological functions, interest in oats with high soluble β-glucan content has increased. β-glucans in oats mainly consist of (1→3), (1→4)-beta-d-glucan, which is a linear polysaccharide, and this glucan is often abbreviated as β-glucan (Figure 2B) [41,42]. The health benefits of β-glucan in wheat and barley are relatively well-known [43]. β-glucans in oats are known to differ from those in wheat and barley in terms of solubility, gelation, and relative molecular weight [44]. In oats, a large amount of β-glucan is contained in the endosperm and aleurone layer cell wall (Figure 2A) [40]. β-glucan regulates the gastrointestinal transit rate after meals and starch digestion to dilute blood sugar level through changes in the glycemic index, thereby reducing the insulinemic response in diabetes [41,45]. In addition, β-glucan has various physiological effects such as preventing cardiovascular diseases by controlling the blood pressure and anti-obesity and anti-cancer effects (e.g., prevention of colorectal cancer) [18,46,47,48,49,50,51,52,53,54,55]. It lowers low-density lipoprotein (LDL) cholesterol level in the blood and elevates high-density lipoprotein (HDL) cholesterol level to help maintain normal blood lipid concentration and body weight [56]. Furthermore, β-glucan activates leucocytes/macrophages and promotes immune function by increasing immunoglobulin levels, and NK and killer T cell numbers [57,58]. This leads to increased resistance against cancer and infections as well as diseases caused by parasites [57,59,60]. β-glucan water-soluble dietary fiber in oats increases intestinal viscosity and shortens the transit time of the intestinal contents and nutrients during peristalsis [61]. This helps to reduce the absorption rate of proteins, lipids, and glucose, leading to decreased body weight and dietary efficiency (Figure 3 and Figure 4) [40,43,62,63]. The cell wall of oats contains a high level of mixed-linked β-d-glucan that consists of β(1→3) and β(1→4) glucosides in a ratio of 3:7 [64]. This mixed-linked β-d-glucan is also known to lower blood cholesterol levels [56]. β-glucans noted above are a class of fiber identified in yeast, algae, bacteria, fungi, and some plants such as oats and barley [43]. The particulate or soluble form of oat-derived β-glucan appears safe when digested [39]. However, some moderate adverse effects have been reported. Symptoms including ulcerative colitis, diarrhea, back pain, joint pain, kidney disease, circadian disruption-induced metabolic syndrome, bile acid storage and vascular calcium storage are reported to be ameliorated in the specific conditions such as ingestion with a high-dose-limiting concentration of β-glucan [48,49,65,66,67,68]. It remains to be elucidated whether this intake is safe for women who are pregnant or breastfeeding. In addition, dietary intake of β-glucan may be unsafe for an individual with certain disease-related conditions such as acquired immune deficiency syndrome (AIDS) and AIDS-related complex [69]. However, no serious adverse effects related to β-glucan supplementations have been reported during laboratory and clinical trials except in some special cases as mentioned above [49,70,71].
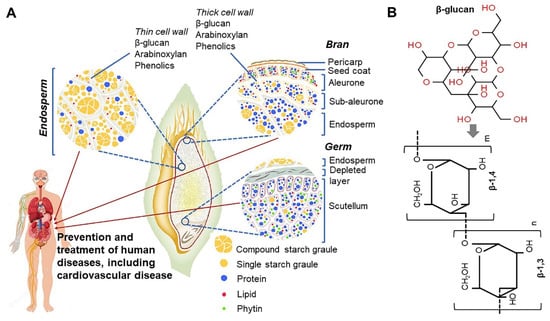
Figure 2.
Structural diagram of the seed-derived cellular components from different oat tissue, including bran, germ, and endosperm, and nutrient distribution and organization within these tissues (A) [18,40], and chemical structure of β-glucan linked to β-1,3 and β-1,4 bridge (B).
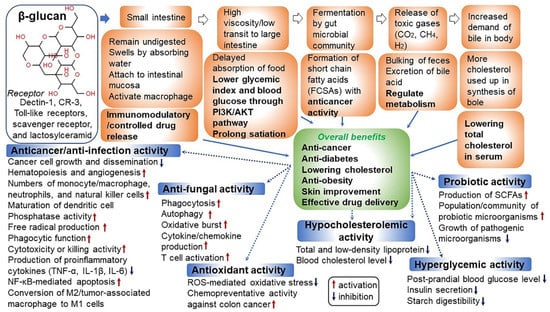
Figure 3.
Human health benefits of β-glucan associated with anti-cancer/anti-infection, anti-fungal, antioxidant, hypocholesterolemic, hyperglycemic, and probiotic activity. Binding receptors for β-glucan have been known to be dectin-1, complement receptor 3 (CR3), scavenger receptor, and lactosylceramid. Of these, dectin-1 is a major receptor [18,52,53,54,55].
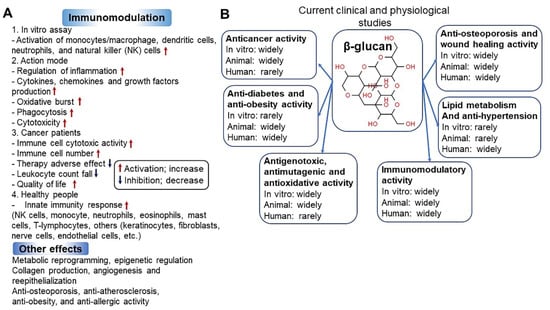
Figure 4.
Immunomodulatory activity (A) [57] and clinical and physiological perspectives (B) [58] of β-glucan.
4. Fermentation Enhancement of Functionality of Oat β-Glucan and Ingredients
Several previous studies have prepared lactic acid fermented beverages from various plant materials. Soymilk has been extensively studied for its property of improved storage and sensory characteristics after lactic acid fermentation [72]. Oligosaccharides such as raffinose and stachyose, which cause flatulence, are hydrolyzed by lactic acid bacteria containing α-galactosidase, thereby improving the nutritional properties [73]. Lactic acid fermentation of peanut oil reduces the content of n-hexanal, a substance that causes a fishy smell in soybean, and thus improves the sensory properties [74]. A study was conducted to assess the changes in the microbiological characteristics to evaluate the growth potential of lactic acid bacteria during fermentation with oat extract as microbial substrate.
Gut microbiome have recently received a light in human health promotion in views of dietary grain consumption and health enhancement. Polyphenolic compounds are rich bioactive components present in grains, indicating the crucial polyphenolic compounds as human health factors [75]. Polyphenolic compounds and fiber present in whole grain diet are helpful to reduce risks of human diseases. Fiber consumption is associated with specific microbial flora such as Bacteroides. Dietary fiber such as arabinoxylans, pectins or insulin promotes enrichment of health-related bacterial flora [76]. In gut, the consumed components are known to interact with the microbiome. Therefore, the oat components are suggested to interact with the gut microbiome to exert human health [76]. The dietary consumption of oat fiber and polyphenolic compounds is considered to help gut bacterial promotion associated with healthy intestinal benefit.
It was observed that fermentation using a combination of Lactobacillus delbrueckii subsp. bulgaricus (L. bulgaricus) and Streptococcus salivarius subsp. thermophilus (S. thermophilus) increased the number of microorganisms and acid production compared to fermentation using only L. bulgaricus or S. thermophilus. In addition, fermentation with S. thermophilus led to a lower pH than fermentation with L. bulgaricus. Fermentation using a mix of L. bulgaricus and S. thermophilus led to a faster decrease in the pH. These results suggest that there is a growth-promoting phenomenon between L. bulgaricus and S. thermophilus, similar to that in yogurt made from milk. The increased number of microorganisms and acid production and lower pH in fermentation using S. thermophilus may be attributed to the better adaptation to oat extract of S. thermophilus than of L. bulgaricus [75,76,77,78,79,80]. These studies have demonstrated that the increased content of the β-glucan in oat plantation is associated with maximum health benefits and hence, biosynthetic enhancement of the β-glucan would be an important goal of functional oat breeding.
5. Other Bioactive Ingredients and Functionalities
Physiologically active ingredients of oats include vitamin E, carotenoids, anthocyanins, lignans, phytic acid, phenolics, and phytosterol, and Avn, which is a phenol present only in oats [81,82]. These components are secondary metabolites produced as defense mechanisms during plant growth and act as antioxidants that control cell damage from oxidative stress by removing reactive oxygen species in the human body [82,83,84,85]. Furthermore, the addition of oat components during the processing of food products helps to suppress fatty acid plaque development because of its antioxidant action and improves storage properties [23,86,87].
Vitamin E consists of four tocopherol isomers (α-, β-, γ-, and δ-tocopherol) and four tocotrienol isoforms (α-, β-, γ-, and δ-tocotrienol) (Figure 5) [88]. Among these, α-tocotrienol has 40–60 times greater antioxidant capacity than β-tocotrienol, a key antioxidant [89]. It lowers blood cholesterol, has anti-inflammatory effects, and inhibits tumor cell proliferation in humans [90]. The main polyphenolic compound found in oats includes protocatechuic, syringic, vanillic, p-hydroxybenzoic, gallic, p-coumaric, o-coumaric, and caffeic acids (Figure 6) [91,92,93]. Among them, Avn biosynthesized from phenylalnine as an alkaloid (Figure 7) [9,94,95] is a polyphenol with various physiological properties, including antioxidant, anti-inflammatory, anti-cancer, anti-thrombotic, anti-proliferative, and anti-itch activities [9,95,96,97,98,99,100,101,102,103]. Avn has 30 times higher antioxidant activity than other phenolic compounds [98,99,102]. There are various types of Avn found in oats. Depending on the residue of N-cinnamoyl anthranilic acid, Avn A combined with p-coumaric acid, Avn B combined with ferulic acid, and Avn C combined with caffeic acid are mainly found in oats [101,102,104]. The structure of Avn is similar to tranilast, a commercially available anti-allergic drug, and many studies assessed the anti-inflammatory and anti-atherogenic effects of Avn [105,106,107,108]. In particular, Avn is known to inhibit the release of inflammatory substances by macrophages or adhesion of monocytes to vascular endothelial cells and exhibit anti-cancer effects through anti-proliferative and pro-apoptotic activities (Figure 8, Figure 9 and Figure 10) [78,99,106,107,108,109,110,111,112,113]. In a recent study, Avn C, among different types of Avn, was shown to be effective against dementia and hearing loss [98,114], as well as prevention of misfolded aggregation (Figure 11) [115]. Avn is also helpful in relieving itching of dry skin, and oat extract is widely used as a cosmetic material in Western Europe [65]. Avns (A, B and C) dose-dependently inhibit cellular tyrosinase and melanin synthesizing tyrosinase activities with the competitive inhibitory manner. They also inhibit the expression level of melanogenic proteins (TRP1 and 2). The tyrosinase-binding affinity of Avns, which obtained by molecular docking simulation and the Derek Nexus quantitative structure–activity relationship system, suggests that the binding affinities are ranged between −7.5 kcal/mol and −6.8 kcal/mol [116]. Moreover, creams containing oat extract showed improved facial skin and were safe to use [117]. Based on these findings, it is possible to explore the feasibility of oat extract as an active ingredient in functional cosmetics to alleviate redness and pigmentation. In addition, combining oats with other natural ingredient extracts for cosmetic products to cover wrinkles, whiten, alleviate redness, and have other functions can lead to several positive changes [65].
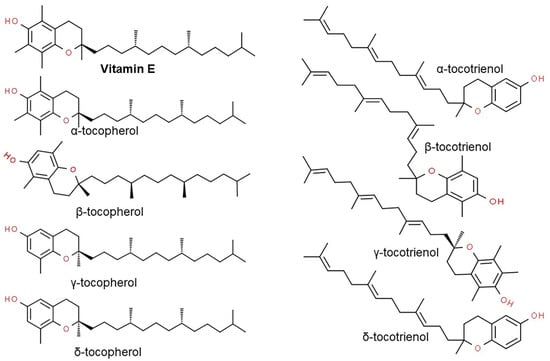
Figure 5.
Chemical structure of vitamin E-derived tocopherol and tocotrienol isomers.
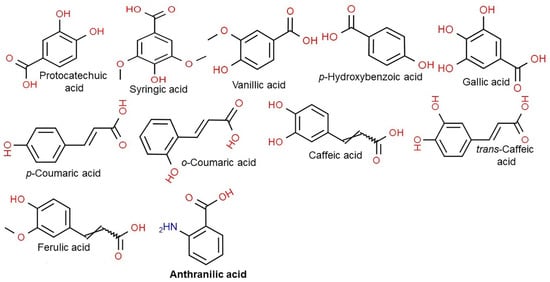
Figure 6.
Chemical structure of main polyphenolic compounds found in oats.
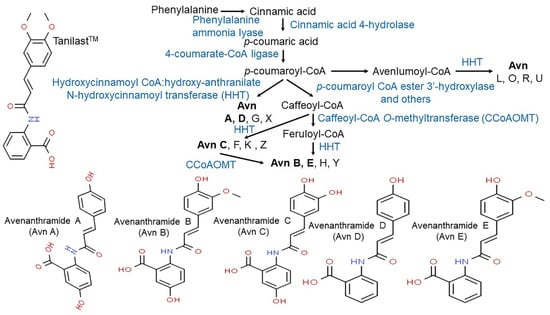
Figure 7.
Proposed biosynthetic pathway of major avenanthramides in oat (Avena sativa L.) [9,94,95].
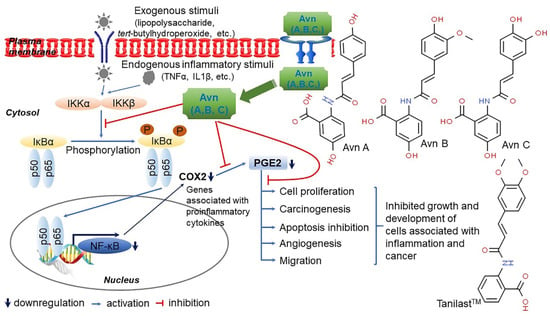
Figure 8.
A predicted mechanism of avenanthramides (Avns)-mediated anti-inflammatory in skeletal muscle C2C12 cells. Avns, the polyphenolic molecules identified solely in oats, exhibit anti-inflammatory activity mainly by inducing nuclear factor-kappaB (NF-κB) inactivation in C2C12 cells. Avns downregulated the expression of IκB kinase beta (IKKβ) as an inhibitor of NF-κB kinase subunit beta in cellular response to tert-butyl hydroperoxide (tBHP)-meditated oxidative stress and attenuated the expression tumor necrosis factor alpha (TNFα) and interleukin 1β (IL-1β) at the transcriptional level under the same condition. Furthermore, Avns reduced the expression of cyclooxygenase-2 (COX-2) protein, along with decreased prostaglandin E2 (PGE2) levels. The downregulated COX2/PGE2 pathway leads to the inhibition of cell proliferation, migration, apoptosis suppression, angiogenesis, and carcinogenesis in various cell lines. Thus, Avns can be potent inhibitors of NF-κB-mediated inflammatory response following the downregulation of IKKβ activity in C2C12 cells [109,110].
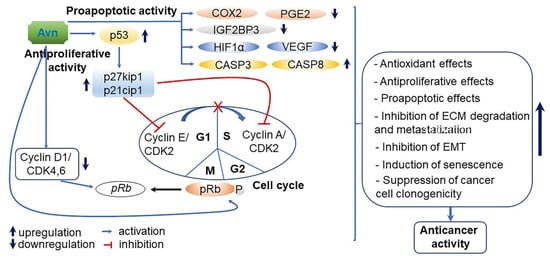
Figure 9.
A proposed action mode of avenanthramides (Avns)-derived antiproliferative and proapoptotic activity. p53, and p27kip1 and p21cip1 activated by Avns treatment suppress the expression of cyclin E/ cyclin-dependent kinase 2 (CDK2) and cyclin A/CDK2 associated with cell cycle, and lead to the cell cycle arrest (G1 to S phase). In addition, Avns also downregulate the expression of cyclin D1/CDK4,6 and enhances phosphorylation of Rb protein (pRb) as a tumor suppressor. As a result, Avns cause cell cycle arrest of M phase. Based on these results, Avns play a vital role in the positive control of the cell cycle and in tumor progression [110,111]. With regard to proapoptotic activity, Avns upregulate caspase 3 (CASP3) and caspase (CASP8), while they downregulate insulin-like growth factor 2 mRNA-binding protein 3 (IGF2BP3), hypoxia-inducible factor 1-alpha (HIF1α), vascular endothelial growth factor (VEGF), cyclooxygenase 2 (COX2), and prostaglandin E2 (PGE2) in tumor cell lines [112]. Therefore, Avns reinforce anticancer effects through increased antioxidative, antiproliferative and proapoptotic effects, as well as induction of senescence, and inhibition of extracellular matrix (ECM) degradation and metastatization and epithelial-mesenchymal transition (EMT).
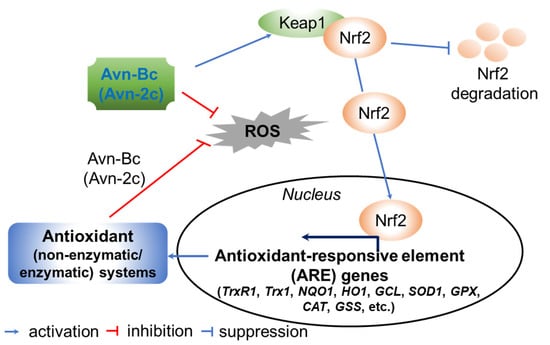
Figure 10.
A proposed action mode of avenanthramide (Avn)-derived antioxidant activity. Avn-Bc unties the complex of nuclear factor erythroid 2-related factor 2 (Nrf2) and Kelch-like ECH-associated protein 1 (Keap1). Detached Ntf2 moves to the nucleus, binds to the antioxidant-responsive element (ARE) domain, and activates a wide range of cytoprotective and antioxidative genes, including thioredoxin reductase (TrxR1), thioredoxin 1 (Trx1), NAD(P)H:quinone oxidoreductase (NQO1), heme oxygenase 1 (HO-1), glutamate-cysteine ligase (GCL), superoxide dismutase 1 (SOD1), glutathione peroxidase (GPX), catalase (CAT) and glutathione synthetase (GSS), at the transcriptional and translational levels under oxidative stress conditions. The activated antioxidant systems improve redox homeostasis by neutralizing reactive oxygen species (ROS). Thus, Avn-Bc enhances cell homeostasis in response to oxidative stress [99,112].
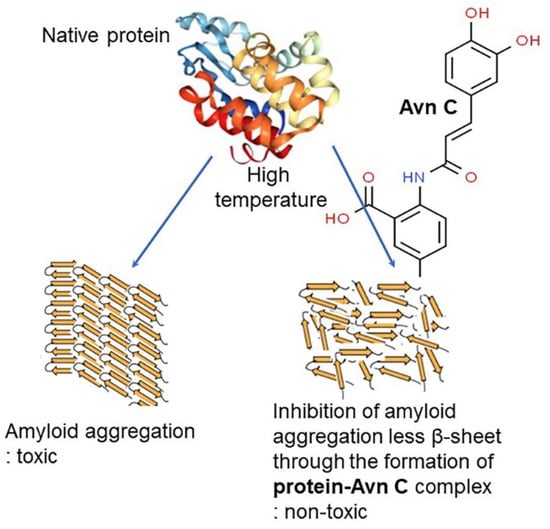
Figure 11.
Prevention of amyloid formation in the presence of avenanthramide C (Avn C). The assembly of misfolded protein into amyloid fibrils that have a high β-sheet-rich secondary structure is associated with many human diseases, including central nervous diseases (Parkinson’s, Alzheimer’s, and Huntington’s disease), amyotrophic lateral sclerosis, and type 2 diabetes, and diseases related to the accumulation of insoluble serum amyloid A protein in liver, spleen, and kidney. Although great efforts have been made to elucidate the pathogenesis of these diseases and development of effective therapy to date, there is still no evidence for the treatment and prevention associated with amyloid-related diseases. Polyphenols such as avenanthramides have been widely studied as a key factor of amyloid aggregation inhibitors. Their bioactive effects depend on the number and position of hydroxyl groups around the flavone backbone. Avn C can act as a potential biomolecule in inhibiting protein aggregation by decreasing the formation of β-sheet structure of protein aggregates [115].
In a study that compared the antioxidant effects and inhibition of cancer cell proliferation of oat extract according to extraction solvents, the antioxidant activity of the extract was measured by assessing the scavenging of 2,2´-azinobis 3-ethylbenzothiazoline-6-sulphonic acid (ABTS) and 1,1-diphenyl-1-picrylhydrazyl (DPPH) radicals and reducing power, whereas the inhibition of cancer cell proliferation was assessed using colorectal, lung, and breast cancer cell lines. It was observed that the total polyphenol content, scavenging of ABTS and DPPH radicals, and reducing power were the greatest in methanol extracts [118]. In addition, methanol extracts had the most significant inhibitory effects on the proliferation of colorectal cancer (HCT116), lung cancer (NCI-H460), and breast cancer (MCF7) cells [119]. Although there were differences in the antioxidant effects and inhibition of cancer cell proliferation of oats depending on the extraction solvent, these findings demonstrate that oats have antioxidant and anti-cancer effects [98,120]. Notably, to date no studies have shown any adverse effects associated with Avn supplementation. Altogether, physiological activities of oat-derived Avns were summarized into Figure 12 [65,110,121].
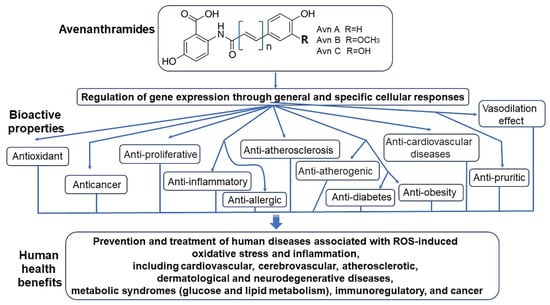
Figure 12.
Physiological bioactivities of oat-derived avenanthramides [65,110,120].
6. Functionality of Sprouted Oats
In light of the nutritional value of ordinary oats, interest in sprouted oats is also gradually increasing. Germination is a process in which the seed absorbs moisture and undergoes various metabolic processes to produce young roots and shoots. The germinal disk with determined genetic information germinates under suitable conditions, and biological processes are induced by activities of enzymes that decompose starch, resulting in the generation of a new plant [96,122]. During germination, various enzymes, nutrients, and genetic information in the germinal disk and endosperm of the seed are activated, and the maximum amount of nutrients is secured. In detail, the germinal disk germinates, and proteins undergo qualitative changes. Amino acids, carbohydrates, fatty acids, and vitamins B1, B2, and E are increased, and minerals and dietary fibers are changed (Figure 2). In addition, physiologically contents of active ingredients such as γ-aminobutyric acid, γ-oryzanol, and arabinoxylan are enhanced [123,124,125]. During germination, enzymes are activated, and softening of the grain can lead to improved texture. In addition, the digestibility of carbohydrates increases, leading to increased absorption of nutrients in the body. Moreover, germination extensively changes the chemical composition of grains [126,127,128]. For example, soaking barley in water and germinating them increases the biodegradability of proteins, vitamins, minerals, and other substances in barley, leading to improved physiological activities [129]. In rye, phytosterol, folate, lignan, and phenolic contents increase during germination [130]. Likewise, a similar phenomenon is observed in oats. The concentration of Avn in oats is significantly increased after germination compared to that before germination [9,131,132]. Furthermore, germination of grains can lead to saccharification effects, which can improve palatability such as by enhancing flavor [126,133]. Thus, various types of processed foods and functional products can be manufactured using these germination characteristics.
7. Functional Enhancement Using Oat By-Products
Recently, studies on the creation of new value-added products from food by-products have been actively conducted [134]. Thus, a considerable amount of by-products such as husks, seeds, grains, and bran are thrown away as wastes [135]. Although food by-products have been mainly used as animal feed until now, some of these by-products can be used in eco-friendly industries [135,136,137]. In addition, these by-products such as sugar cane, fruit beets, whey, bread, and wheat by-products as natural materials are also utilized for functional components [55,138,139,140]. However, the functional properties and nutritional value of these by-products have not been actively studied, although by-products can be utilized [141]. Food by-products are considered high-value food additives for their antioxidant, anti-bacterial, colorant, and flavoring functions [142,143,144,145]. More than 20 million tons of oats are annually produced, mainly as food or animal feed [146]. Oat bran is a key by-product and accounts for approximately 50% of the weight of dry oat grains [147]. Oat bran also contains high-quality proteins such as β-glucan, saponin, albumin, prolamins, and glutelins that lower cholesterol levels (Figure 13) [148,149,150,151,152]. To date, studies on oats mainly assessed the nutritional value and processing characteristics of oat grains or powder [13,42,55]. There is a lack of studies on its by-products, such as the bran, hull, and leaves, which are mostly thrown away as waste during the processing [153]. However, as recent studies reported the content of numerous functional components in these by-products, the utility of oats is gradually increasing [42,55].
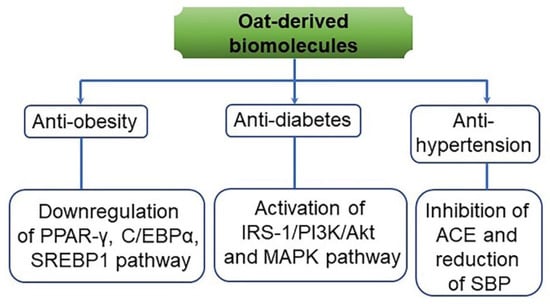
Figure 13.
A predicted action mode of anti-obesity, anti-diabetes, and anti-hypertension effects from oat-derived biomolecules and by-products. Anti-obesity is associated with the downregulation of peroxisome proliferator-activated receptor gamma (PPAR-γ), CCAAT-enhancer-binding protein isoform alpha (C/EBPα), and sterol regulatory element-binding protein 1 (SREBP1). Anti-diabetes is involved in the activation of insulin receptor substrate 1 (IRS-1)/phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) and AMP-activated protein kinase (AMPK) signaling pathway. Anti-hypertension is related to the inhibition of angiotensin-converting enzyme (ACE) and the reduction of systolic blood pressure (SBP) [147].
8. Importance of Oat Breeding
Compared to other grains, oats grow well in harsh environmental conditions with insufficient nutrients, can be cultivated in cold and humid climates, have low chemical fertilizer requirements, have fewer pests, and require less chemical control, making them eco-friendly [154]. Approximately 70% of the produced oat grains are consumed for animal food, and the remaining 30% are consumed as food products for humans [155]. However, farmers prefer crops such as wheat, rice, and barley, which provide higher yields and profits compared to oats, and thus, oat production has gradually decreased over the past few decades globally. As a result, unlike wheat, barley, and rice, agricultural research on oats is limited [156]. Previous studies by oat breeders mainly focused on phenotype selection and disease resistance related to the yield, and there are fewer studies on genetics and other necessary traits of oats than those of other grains [157,158,159]. However, following the recognition of their excellent nutritional effects and health benefits, studies on oats and their consumption have steadily increased, including in Korea [24]. However, studies have been mainly focused on the cultivation or quality evaluation as fodder crops and development of bulky feed varieties rather than edible varieties [160,161,162]. Moreover, previous studies often assessed the extraction and physiological activity of its functional component, β-glucan [163]. Thus, the development of new varieties for food or processed products has not been actively conducted. In particular, studies on genetic resources as breeding materials and breeding for improved nutritional function are limited [164]. However, the public awareness regarding the health functionalities of oats has rapidly enhanced their consumption, and such improvement in nutritional function may have positive effects in promoting their utilization in the future [165,166]. Therefore, future studies should not only actively seek to increase the yield but also improve the nutritional value by cultivating varieties with high contents of Avn and β-glucan (Figure 14) [23,166,167,168,169].
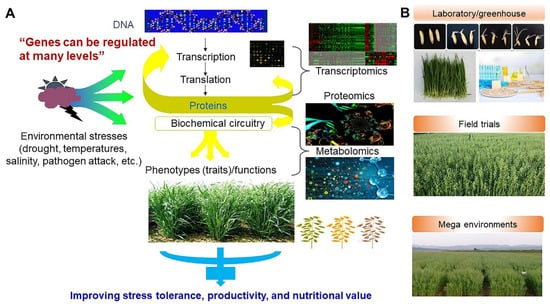
Figure 14.
Breeding strategy of oat crop plants to improve stress tolerance, productivity, and high nutritional value based on transcriptomics, proteomics, and metabolomics (A), and evaluation of agronomic traits on laboratory and greenhouse conditions, field trials, and mega environments (B).
9. Conclusions and Perspectives
Oats, which belong to the Poaceae family, grow well in cool and humid weather and are annual crops. Oats are aware of the surrounding environment with superior senses compared to other plants, effectively compete with limited resources available in the soil and atmosphere, and accurately judge the surroundings. Oats also have the ability to determine and implement strategies to respond to environmental stimuli. In other words, oats detect changes and produce various useful substances while being sensitive to altered environmental conditions. The physiological activities of oats, such as lowering blood cholesterol levels, have been demonstrated in studies, and thus, interest in oats as a functional food has increased. Oats contain 7–14% of dietary fiber and 3–5% of β-glucan, which is one of the important functional components of oats. β-glucan is a water-soluble dietary fiber found in oats and exhibits viscosity. It can reduce the risk of heart disease by inhibiting the absorption of cholesterol in the intestine. β-glucan exhibits anti-obesity effects by inhibiting the accumulation of adipocytes (thereby reducing the formation of body fat), preventing the accumulation of cholesterol in the liver, and improving lipid metabolism. Furthermore, Avn, which is present only in oats, has various physiological activities along with β-glucan in increasing the nutritional and functional values of oats. The oat β-glucans are potentially suggested as a celiac disease-targeting diet resource, especially for gluten-free diets [165]. However, the scientific studies to apply the β-glucans for the autoimmune diseases are not performed yet. Oats also contain high levels of tocotrienol (another type of vitamin E), which possesses antioxidant and anti-cancer effects and improves hyperlipidemia. Therefore, oats embody a much more resistant and modern crop model than other plants.
Oats are a prime example of a combination of solidity and flexibility. The modular composition of oats is the essence of modernization, and oats can actively respond to repeated changes in the environment without losing their functions through their distributed cooperative structure without a control center for quick adaptation. Their unique evolution has led to solutions different from those of other plants, suggesting that oats are much more modern than other plants. Thus, the unique characteristics of oats may offer several opportunities for the development of novel functional foods in the coming years.
Oats are important to Koreans, who mainly consume rice. Many Koreans reduce the proportion of rice in meals to decrease sugar intake; therefore, oats, which have a good nutritional balance, are an excellent alternative to rice or can be mixed with rice. Asians, who consume grain-oriented meals, often lack essential amino acids that must be additionally obtained from protein foods. Thus, the intake of oats can help to improve the consumption of more high-quality proteins such as soybeans. In addition, individuals often suffer from low immunity due to stress, overwork, excessive drinking, chronic fatigue, lack of sleep, and incorrect dietary habits. Although they are aware of their problems, proper health management in a busy life is not easy, leading to the intake of expensive nutritional supplements or health foods. However, it is necessary to balance the nutrients in daily meals naturally. The intake of essential nutrients leads to improved immunity to fight against various stresses. Oats, containing various nutrients in small amounts, would help prevent and treat different adult diseases by enhancing the immune system through nutritional balance relatively easily. Therefore, future studies on oats must not only actively seek to increase the yield but also improve the nutritional value by cultivating varieties with high contents of Avn and β-glucan. In addition, studies should simultaneously assess the physiological activity and related mechanisms of functional ingredients.
Author Contributions
Conceptualization, I.-S.K., C.-W.H., W.-S.Y. and C.-H.K.; writing—original draft preparation, I.-S.K.; writing—review and editing, C.-W.H.; W.-S.Y. and C.-H.K.; visualization, I.-S.K. and W.-S.Y.; supervision, C.-W.H., W.-S.Y. and C.-H.K.; project administration, C.-H.K., C.-W.H. and W.-S.Y.; funding acquisition and revision, C.-H.K., C.-W.H. and W.-S.Y. All authors have read and agreed to the published version of the manuscript.
Funding
The study has supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET 119010033SB010 derived from the IPET No.119010033CG000) during 2019–2021.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yuanqing, H.; Pengyao, Y.; Yangyang, D.; Min, C.; Rui, G.; Yuqing, D.; Haihui, Z.; Haile, M. The preparation, antioxidant activity evaluation, and iron-deficient anemic improvement of oat (Avena sativa L.) peptides–ferrous chelate. Front. Nutr. 2021, 8, 687133. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.W.; Nash, P.; Buttar, H.S.; Griffiths, K.; Singh, R.; De Meester, F.; Horiuchi, R.; Takahashi, T. The role of food antioxidants, benefits of functional foods, and influence of feeding habits on the health of the older person: An overview. Antioxidants 2017, 6, 81. [Google Scholar] [CrossRef]
- Damiran, D.; Yu, P. Structural makeup, biopolymer conformation, and biodegradation characteristics of a newly developed super genotype of oats (CDC SO-I versus conventional varieties): A novel approach. J. Agric. Food Chem. 2010, 58, 2377–2387. [Google Scholar] [CrossRef] [PubMed]
- Acharjee, S.; Sarmah, B.K. Biotechnologically generating ‘super chickpea’ for food and nutritional security. Plant Sci. 2013, 207, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Yu, H.; Wang, B.; Li, J. Retrospective and perspective of rice breeding in China. J. Genet. Genom. 2018, 45, 603–612. [Google Scholar] [CrossRef]
- Li, G.; Zhu, F. Quinoa starch: Structure, properties, and applications. Carbohydr. Polym. 2018, 181, 851–861. [Google Scholar] [CrossRef]
- Wing, R.A.; Purugganan, M.D.; Zhang, Q. The rice genome revolution: From an ancient grain to Green Super Rice. Nat. Rev. Genet. 2018, 19, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Ahmad, M.; Ahmed, M.; Iftikhar Hussain, M. Rising atmospheric temperature impact on wheat and thermotolerance strategies. Plants 2020, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, T.; Jeong, Y.; Park, S.; Park, S.; Lee, J.; Yang, K.; Jeong, J.; Kim, C. Synergistic effect of methyl jasmonate and abscisic acid co-treatment on avenanthramide production in germinating oats. Int. J. Mol. Sci. 2021, 22, 4779. [Google Scholar] [CrossRef]
- Martín-Diana, A.B.; García-Casas, M.J.; Martínez-Villaluenga, C.; Frías, J.; Peñas, E.; Rico, D. Wheat and oat brans as sources of polyphenol compounds for development of antioxidant nutraceutical ingredients. Foods 2021, 10, 115. [Google Scholar] [CrossRef]
- Mert, I.D. The applications of microfluidization in cereals and cereal-based products: An overview. Crit. Rev. Food Sci. Nutr. 2020, 60, 1007–1024. [Google Scholar] [CrossRef]
- McKevith, B. Nutritional aspects of cereals. Nutr. Bull. 2004, 29, 111–142. [Google Scholar] [CrossRef]
- Rasane, P.; Jha, A.; Sabikhi, L.; Kumar, A.; Unnikrishnan, V.S. Nutritional advantages of oats and opportunities for its pro-cessing as value added foods—A review. J. Food Sci. Technol. 2015, 52, 662–675. [Google Scholar] [CrossRef]
- Saccomanno, B.; Chambers, A.H.; Hayes, A.; Mackay, I.; McWilliam, S.C.; Trafford, K. Starch granule morphology in oat endosperm. J. Cereal Sci. 2017, 73, 46–54. [Google Scholar] [CrossRef]
- Nogala-Kalucka, M.; Kawka, A.; Dwiecki, K.; Siger, A. Evaluation of bioactive compounds in cereals. Study of wheat, barley, oat and selected grain products. Acta Sci. Pol. Technol. Aliment 2020, 19, 405–423. [Google Scholar]
- Klose, C.; Arendt, E.K. Proteins in oats; their synthesis and changes during germination: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Gell, G.; Kovacs, K.; Veres, G.; Korponay-Szabo, I.R.; Juhasz, A. Characterization of globulin storage proteins of a low prola-min cereal species in relation to celiac disease. Sci. Rep. 2017, 7, 39876. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, G.; Sowrirajan, S.; Joseph, B. β-Glucan, a dietary fiber in effective prevention of lifestyle diseases—An insight. Bioact. Carbohydr. Dietary Fibre 2019, 19, 100187. [Google Scholar] [CrossRef]
- Capurso, C. Whole-grain intake in the mediterranean diet and a low protein to carbohydrates ratio can help to reduce mor-tality from cardiovascular disease, slow down the progression of aging, and to improve lifespan: A review. Nutrients 2021, 13, 2540. [Google Scholar] [CrossRef]
- Varma, P.; Bhankharia, H.; Bhatia, S. Oats: A multi-functional grain. J. Clin. Prev. Cardiol. 2016, 5, 9–17. [Google Scholar] [CrossRef]
- Díaz, L.D.; Fernández-Ruiz, V.; Cámara, M. An international regulatory review of food health-related claims in functional food products labeling. J. Funct. Foods 2020, 68, 103896. [Google Scholar] [CrossRef]
- Shvachko, N.; Loskutov, I.; Semilet, T.; Popov, V.; Kovaleva, O.; Konarev, A. Bioactive components in oat and barley grain as a promising breeding trend for functional food production. Molecules 2021, 26, 2260. [Google Scholar] [CrossRef] [PubMed]
- Tieri, M.; Ghelfi, F.; Vitale, M.; Vetrani, C.; Marventano, S.; Lafranconi, A.; Godos, J.; Titta, L.; Gambera, A.; Alonzo, E.; et al. Whole grain consumption and human health: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2020, 71, 668–677. [Google Scholar] [CrossRef]
- Chen, O.; Mah, E.; Dioum, E.; Marwaha, A.; Shanmugam, S.; Malleshi, N.; Sudha, V.; Gayathri, R.; Unnikrishnan, R.; Anjana, R.; et al. The role of oat nutrients in the immune system: A narrative review. Nutrients 2021, 13, 1048. [Google Scholar] [CrossRef]
- Daly, M.; Bromilow, S.N.; Nitride, C.; Shewry, P.R.; Gethings, L.A.; Mills, E.N.C. Mapping coeliac toxic motifs in the prolamin seed storage proteins of barley, rye, and oats using a curated sequence database. Front. Nutr. 2020, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhao, D.; Liu, Q. Connections between amino acid metabolisms in plants: Lysine as an example. Front. Plant Sci. 2020, 11, 928. [Google Scholar] [CrossRef]
- Jaeger, A.; Zannini, E.; Sahin, A.W.; Arendt, E.K. Barley protein properties, extraction and applications, with a focus on brewers’ spent grain protein. Foods 2021, 10, 1389. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Gao, Z.; Greenway, F.L.; Burton, J.; Johnson, W.; Keenan, M.J.; Enright, F.M.; Martin, R.J.; Chu, Y.; Zheng, J. Oat consumption reduced intestinal fat deposition and improved health span in Caenorhabditis elegans model. Nutr. Res. 2015, 35, 834–843. [Google Scholar] [CrossRef]
- Ahmet, B.U.G.; Musa, O.M.; Ziya, D.; Nurhans, U. Oil contents and fatty acid composition of oat (Avena sativa L.) seed and oils. J. Agroaliment. Proc. Technol. 2019, 25, 182–186. [Google Scholar]
- Culetu, A.; Ionescu, V.; Todasca, C.; Duta, D.E. Evaluation of the storage-associated changes in the fatty acid profile of oat-based gluten-free cookies prepared with different fats. Food Sci. Biotechnol. 2020, 29, 759–767. [Google Scholar] [CrossRef]
- Sterna, V.; Zute, S.; Brunava, L. Oat grain composition and its nutrition benefice. Agric. Agric. Sci. Procedia 2016, 8, 252–256. [Google Scholar] [CrossRef]
- Yau, Y.F.; El-Nezami, H.; Galano, J.M.; Kundi, Z.M.; Durand, T.; Lee, J.C. Lactobacillus rhamnosus GG and oat beta-glucan regulated fatty acid profiles along the gut-liver-brain axis of mice fed with high fat diet and demonstrated antioxidant and anti-inflammatory potentials. Mol. Nutr. Food Res. 2020, 64, e2000566. [Google Scholar] [CrossRef] [PubMed]
- Banaś, K.; Harasym, J. Current knowledge of content and composition of oat oil—Future perspectives of oat as oil source. Food Bioproc. Technol. 2020, 14, 232–247. [Google Scholar] [CrossRef]
- Livesey, G. A perspective on food energy standards for nutrition labelling. Br. J. Nutr. 2001, 85, 271–287. [Google Scholar] [CrossRef]
- Kowalska, H.; Kowalska, J.; Ignaczak, A.; Masiarz, E.; Domian, E.; Galus, S.; Ciurzyńska, A.; Salamon, A.; Zając, A.; Marzec, A. Development of a high-fibre multigrain bar technology with the addition of curly kale. Molecules 2021, 26, 3939. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, H.; Landberg, R.; Sundberg, B.; Lundin, E.; Hallmans, G.; Zhang, J.X.; Tidehag, P.; Knudsen, K.E.B.; Moazzami, A.A.; Aman, P. High-fiber rye diet increases ileal excretion of energy and macronutrients compared with low-fiber wheat diet independent of meal frequency in ileostomy subjects. Food Nutr. Res. 2013, 57, 18519. [Google Scholar] [CrossRef][Green Version]
- Lunn, J.; Buttriss, J.L. Carbohydrates and dietary fibre. Nutr. Bull. 2007, 32, 21–64. [Google Scholar] [CrossRef]
- Proestos, C. Superfoods: Recent data on their role in the prevention of diseases. Curr. Res. Nutr. Food Sci. 2018, 6, 576–593. [Google Scholar] [CrossRef]
- Henrion, M.; Francey, C.; Le, K.A.; Lamothe, L. Cereal β-glucans: The impact of processing and how it affects physiological responses. Nutrients 2019, 11, 1729. [Google Scholar] [CrossRef] [PubMed]
- Grundy, M.M.; Fardet, A.; Tosh, S.M.; Rich, G.T.; Wilde, P.J. Processing of oat: The impact on oat’s cholesterol lowering effect. Food Funct. 2018, 9, 1328–1343. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Cao, J.; Zhong, S.; Gao, Y.; Cui, X. Seeking aggregation-induced emission materials in food: Oat beta-glucan and its diverse applications. J. Agric. Food Chem. 2021, 69, 7680–7686. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Dong, R.; Hu, X.; Ren, C.; Li, Y. Oat-based foods: Chemical constituents, glycemic index, and the effect of processing. Foods 2021, 10, 1304. [Google Scholar] [CrossRef]
- El Khoury, D.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta glucan: Health benefits in obesity and metabolic syndrome. J. Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A concise review on the molecular structure and function relationship of beta-glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar] [CrossRef] [PubMed]
- Barati, Z.; Iravani, M.; Karandish, M.; Haghighizadeh, M.H.; Masihi, S. The effect of oat bran consumption on gestational diabetes: A randomized controlled clinical trial. BMC Endocr. Disord. 2021, 21, 67. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Chen, Y.; Zhao, J.; Jing, R.; Guo, K.; Wang, L.; Li, X.; Li, C.; Hu, Z.; Xu, N. Oat beta-glucan ameliorates epidermal barrier disruption by upregulating the expression of CaSR through dectin-1-mediated ERK and p38 signaling pathways. Int. J. Biol. Macromol. 2021, 185, 876–889. [Google Scholar] [CrossRef]
- Climova, A.; Ibrahim, M.N.G.; Salamahina, A.; Savin, A.M.; Dukhinova, M.S.; Barakova, N.V.; Krivoshapkina, E.F. Application of extracted beta-glucan from oat for beta-carotene encapsulation. J. Food Sci. Technol. 2021, 58, 2641–2650. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Lam, K.L.; Li, X.; Kong, A.P.; Cheung, P.C. Circadian disruption-induced metabolic syndrome in mice is ameliorated by oat beta-glucan mediated by gut microbiota. Carbohydr. Polym. 2021, 267, 118216. [Google Scholar] [CrossRef]
- Wolever, T.M.S.; Rahn, M.; Dioum, E.H.; Jenkins, A.L.; Ezatagha, A.; Campbell, J.E.; Chu, Y. Effect of oat beta-glucan on affective and physical feeling states in healthy adults: Evidence for reduced headache, fatigue, anxiety and limb/joint pains. Nutrients 2021, 13, 1534. [Google Scholar] [CrossRef]
- Zyla, E.; Dziendzikowska, K.; Kamola, D.; Wilczak, J.; Sapierzynski, R.; Harasym, J.; Gromadzka-Ostrowska, J. Anti-inflammatory activity of oat beta-glucans in a Crohn’s disease model: Time- and molar mass-dependent effects. Int. J. Mol. Sci. 2021, 22, 4485. [Google Scholar] [CrossRef]
- Ames, N.; Malunga, L.N.; Mollard, R.; Johnson, J.; Chu, Y.; Thandapilly, S.J. Effect of processing on oat β-glucan viscosity, postprandial glycemic response and subjective measures of appetite. Food Funct. 2021, 12, 3672–3679. [Google Scholar] [CrossRef]
- Schmidt, M. Cereal beta-glucans: An underutilized health endorsing food ingredient. Crit. Rev. Food Sci. Nutr. 2020, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J.; Jesenák, M. β-Glucans: Multi-functional modulator of wound healing. Molecules 2018, 23, 806. [Google Scholar] [CrossRef] [PubMed]
- Ciecierska, A.; Drywień, M.; Hamulka, J.; Sadkowski, T. Nutraceutical functions of beta-glucans in human nutrition. Rocz. Panstw. Zakl. Hig. 2019, 70, 315–324. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, Y.; Li, Y.; Qi, X.; Yang, J.; Qi, H.; Li, Q.; Ma, Y.; Zhang, Y.; Zhang, X.; et al. A bifunctional zwitterion—modified porphyrin for photodynamic nondestructive tooth whitening and biofilm eradication. Adv. Funct. Mater. 2021, 2104799. [Google Scholar] [CrossRef]
- Ms Wolever, T.; Rahn, M.; Dioum, E.; Spruill, S.E.; Ezatagha, A.; Campbell, J.E.; Jenkins, A.L.; Chu, Y. An oat beta-glucan beverage reduces LDL cholesterol and cardiovascular disease risk in men and women with borderline high cholesterol: A double-blind, randomized, controlled clinical trial. J. Nutr. 2021. [Google Scholar] [CrossRef] [PubMed]
- Del Cornò, M.; Gessani, S.; Conti, L. Shaping the innate immune response by dietary glucans: Any role in the control of cancer? Cancers 2020, 12, 155. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.M.; Choi, J.S. Clinical and physiological perspectives of β-glucans: The past, present, and future. Int. J. Mol. Sci. 2017, 18, 1906. [Google Scholar] [CrossRef] [PubMed]
- Blaszczyk, K.; Gajewska, M.; Wilczak, J.; Kamola, D.; Majewska, A.; Harasym, J.; Gromadzka-Ostrowska, J. Oral administra-tion of oat beta-glucan preparations of different molecular weight results in regulation of genes connected with immune re-sponse in peripheral blood of rats with LPS-induced enteritis. Eur. J. Nutr. 2019, 58, 2859–2873. [Google Scholar] [CrossRef] [PubMed]
- Moerings, B.G.J.; de Graaff, P.; Furber, M.; Witkamp, R.F.; Debets, R.; Mes, J.J.; van Bergenhenegouwen, J.; Govers, C. Con-tinuous exposure to non-soluble beta-glucans induces trained immunity in M-CSF-differentiated macrophages. Front. Immu-nol. 2021, 12, 672796. [Google Scholar] [CrossRef]
- Schop, M.; Jansman, A.J.M.; de Vries, S.; Gerrits, W.J.J. Increased diet viscosity by oat beta-glucans decreases the passage rate of liquids in the stomach and affects digesta physicochemical properties in growing pigs. Animal 2020, 14, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Joyce, S.A.; Kamil, A.; Fleige, L.; Gahan, C.G.M. The cholesterol-lowering effect of oats and at beta glucan: Modes of action and potential role of bile acids and the microbiome. Front. Nutr. 2019, 6, 171. [Google Scholar] [CrossRef] [PubMed]
- Pino, J.L.; Mujica, V.; Arredondo, M. Effect of dietary supplementation with oat β-glucan for 3 months in subjects with type 2 diabetes: A randomized, double-blind, controlled clinical trial. J. Funct. Foods 2021, 77, 104311. [Google Scholar] [CrossRef]
- Yoo, H.-U.; Ko, M.-J.; Chung, M.-S. Hydrolysis of beta-glucan in oat flour during subcritical-water extraction. Food Chem. 2019, 308, 125670. [Google Scholar] [CrossRef]
- Liu, B.; Lin, Q.; Yang, T.; Zeng, L.; Shi, L.; Chen, Y.; Luo, F. Oat β-glucan ameliorates dextran sulfate sodium (DSS)—Induced ulcerative colitis in mice. Food Funct. 2015, 6, 3454–3463. [Google Scholar] [CrossRef]
- Greenway, F.; Wang, S.; Heiman, M. A novel cobiotic containing a prebiotic and an antioxidant augments the glucose control and gastrointestinal tolerability of metformin: A case report. Benef. Microbes 2014, 5, 29–32. [Google Scholar] [CrossRef]
- Mäkelä, N.; Rosa-Sibakov, N.; Wang, Y.-J.; Mattila, O.; Nordlund, E.; Sontag-Strohm, T. Role of β-glucan content, molecular weight and phytate in the bile acid binding of oat β-glucan. Food Chem. 2021, 358, 129917. [Google Scholar] [CrossRef]
- Fehlbaum, S.; Prudence, K.; Kieboom, J.; Heerikhuisen, M.; Broek, T.V.D.; Schuren, F.H.J.; Steinert, R.E.; Raederstorff, D. In vitro fermentation of selected prebiotics and their effects on the composition and activity of the adult gut microbiota. Int. J. Mol. Sci. 2018, 19, 3097. [Google Scholar] [CrossRef]
- Rahar, S.; Swami, G.; Nagpal, N.; Nagpal, M.A.; Singh, G.S. Preparation, characterization, and biological properties of beta-glucans. J. Adv. Pharm. Technol. Res. 2011, 2, 94–103. [Google Scholar] [CrossRef]
- Hill, E.; Sapa, H.; Negrea, L.; Bame, K.; Hostetter, T.; Barkoukis, H.; Dusso, A.; Dobre, M. Effect of oat beta-glucan supple-mentation on chronic kidney disease: A feasibility study. J. Ren. Nutr. 2020, 30, 208–215. [Google Scholar] [CrossRef]
- Biorklund, M.; van Rees, A.; Mensink, R.P.; Onning, G. Changes in serum lipids and postprandial glucose and insulin con-centrations after consumption of beverages with beta-glucans from oats or barley: A randomised dose-controlled trial. Eur. J. Clin. Nutr. 2005, 59, 1272–1281. [Google Scholar] [CrossRef]
- Mishra, B.K.; Hati, S.; Das, S.; Prajapati, J.B. Biofunctional attributes and storage study of soy milk fermented by Lactobacillus rhamnosus and Lactobacillus helveticus. Food Technol. Biotechnol. 2019, 57, 399–407. [Google Scholar] [CrossRef]
- Hu, Y.; Tian, G.; Geng, X.; Zhang, W.; Zhao, L.; Wang, H.; Ng, T.B. A protease-resistant α-galactosidase from Pleurotus citrinopileatus with broad substrate specificity and good hydrolytic activity on raffinose family oligosaccharides. Process Biochem. 2016, 51, 491–499. [Google Scholar] [CrossRef]
- Xiang, H.; Sun-Waterhouse, D.; Waterhouse, G.I.N.; Cui, C.; Ruan, Z. Fermentation-enabled wellness foods: A fresh perspective. Food Sci. Hum. Well. 2019, 8, 203–243. [Google Scholar] [CrossRef]
- Mena, P.; Bresciani, L. Dietary fibre modifies gut microbiota: What’s the role of (poly)phenols? Int. J. Food Sci. Nutr. 2020, 71, 783–784. [Google Scholar] [CrossRef] [PubMed]
- van der Merwe, M. Gut microbiome changes induced by a diet rich in fruits and vegetables. Int. J. Food Sci. Nutr. 2020, 72, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Bruckner-Guhmann, M.; Vasil’eva, E.; Culetu, A.; Duta, D.; Sozer, N.; Drusch, S. Oat protein concentrate as alternative ingredient for non-dairy yoghurt-type product. J. Sci. Food Agric. 2019, 99, 5852–5857. [Google Scholar] [CrossRef]
- Wu, H.; Rui, X.; Li, W.; Xiao, Y.; Zhou, J.; Dong, M. Whole-grain oats (Avena sativa L.) as a carrier of lactic acid bacteria and a supplement rich in angiotensin I-converting enzyme inhibitory peptides through solid-state fermentation. Food Funct. 2018, 9, 2270–2281. [Google Scholar] [CrossRef] [PubMed]
- Kedia, G.; Vázquez, J.A.; Pandiella, S. Evaluation of the fermentability of oat fractions obtained by debranning using lactic acid bacteria. J. Appl. Microbiol. 2008, 105, 1227–1237. [Google Scholar] [CrossRef][Green Version]
- Patel, H.M.; Wang, R.; Chandrashekar, O.; Pandiella, S.S.; Webb, C. Proliferation of Lactobacillus plantarum in solid-state fermentation of oats. Biotechnol. Prog. 2004, 20, 110–116. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Raguindin, P.F.; Itodo, O.A.; Stoyanov, J.; Dejanovic, G.M.; Gamba, M.; Asllanaj, E.; Minder, B.; Bussler, W.; Metzger, B.; Muka, T.; et al. A systematic review of phytochemicals in oat and buckwheat. Food Chem. 2020, 338, 127982. [Google Scholar] [CrossRef]
- Guan, X.; Jin, S.; Li, S.; Huang, K.; Liu, J. Process optimization, characterization and antioxidant capacity of oat (Avena Sativa L.) bran oil extracted by subcritical butane extraction. Molecules 2018, 23, 1546. [Google Scholar] [CrossRef] [PubMed]
- Ratnasari, N.; Walters, M.; Tsopmo, A. Antioxidant and lipoxygenase activities of polyphenol extracts from oat brans treated with polysaccharide degrading enzymes. Heliyon 2017, 3, e00351. [Google Scholar] [CrossRef] [PubMed]
- Starowicz, M.; Arpaci, S.; Topolska, J.; Wronkowska, M. Phytochemicals and antioxidant activity in oat-buckwheat dough and cookies with added spices or herbs. Molecules 2021, 26, 2267. [Google Scholar] [CrossRef]
- Orme, A.; Louveau, T.; Stephenson, M.J.; Appelhagen, I.; Melton, R.; Cheema, J.; Li, Y.; Zhao, Q.; Zhang, L.; Fan, D.; et al. A noncanonical vacuolar sugar transferase required for biosynthesis of antimicrobial defense compounds in oat. Proc. Natl. Acad. Sci. USA 2019, 116, 27105–27114. [Google Scholar] [CrossRef] [PubMed]
- Moglia, A.; Goitre, L.; Gianoglio, S.; Baldini, E.; Trapani, E.; Genre, A.; Scattina, A.; Dondo, G.; Trabalzini, L.; Beekwilder, J.; et al. Evaluation of the bioactive properties of avenanthramide analogs produced in recombinant yeast. Biofactors 2015, 41, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Gonzalez, J.J.; Garvin, D.F. Subgenome-specific assembly of vitamin E biosynthesis genes and expression patterns during seed development provide insight into the evolution of oat genome. Plant Biotechnol. J. 2016, 14, 2147–2157. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sundaram, C.; Prasad, S.; Kannappan, R. Tocotrienols, the vitamin E of the 21st century: Its potential against cancer and other chronic diseases. Biochem. Pharmacol. 2010, 80, 1613–1631. [Google Scholar] [CrossRef]
- Szewczyk, K.; Chojnacka, A.; Gornicka, M. Tocopherols and tocotrienols-bioactive dietary compounds; what is certain, what is doubt? Int. J. Mol. Sci. 2021, 22, 6222. [Google Scholar] [CrossRef]
- Soycan, G.; Schar, M.Y.; Kristek, A.; Boberska, J.; Alsharif, S.N.S.; Corona, G.; Shewry, P.R.; Spencer, J.P.E. Composition and content of phenolic acids and avenanthramides in commercial oat products: Are oats an important polyphenol source for consumers? Food Chem. X 2019, 3, 100047. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Gangopadhyay, N.; Hossain, M.B.; Rai, D.K.; Brunton, N.P. A review of extraction and analysis of bioactives in oat and bar-ley and scope for use of novel food processing technologies. Molecules 2015, 20, 10884–10909. [Google Scholar] [CrossRef]
- Gutierrez-Gonzalez, J.J.; Tu, Z.J.; Garvin, D.F. Analysis and annotation of the hexaploid oat seed transcriptome. BMC Genom. 2013, 14, 471. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; Meesapyodsuk, D.; Qiu, X. The biosynthetic pathway of major avenanthramides in oat. Metabolites 2019, 9, 163. [Google Scholar] [CrossRef]
- Xochitl, A.; Rosalía, R.; Minerva, R.; Mendoza-Sánchez, M.; Mora, O.; Pérez-Ramírez, I.F. Polyphenols and avenanthramides extracted from oat (Avena sativa L.) grains and sprouts modulate genes involved in glucose and lipid metabolisms in 3T3 L1 adipocytes. J. Food Biochem. 2021, 45, e13738. [Google Scholar] [CrossRef]
- Campbell, S.C. Faecalibacterium prausnitzii abundance in mouse and human gut can predict metabolism of oat avenanthramides. J. Nutr. 2021, 151, 1369–1370. [Google Scholar] [CrossRef]
- Park, J.; Choi, H.; Abekura, F.; Lim, H.S.; Im, J.H.; Yang, W.S.; Hwang, C.W.; Chang, Y.C.; Lee, Y.C.; Park, N.G.; et al. Avenanthramide C suppresses matrix metalloproteinase-9 expression and migration through the MAPK/NF-kappaB signaling pathway in TNF-alpha-activated HASMC cells. Front. Pharmacol. 2021, 12, 621854. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Peng, S.; Song, Z.; Bai, F.; Li, X.; Fang, J. Oat polyphenol avenanthramide-2c confers protection from oxidative stress by regulating the Nrf2-ARE signaling pathway in PC12 cells. Arch. Biochem. Biophys. 2021, 706, 108857. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, S.; Yerke, A.; Ohland, C.L.; Gharaibeh, R.Z.; Fouladi, F.; Fodor, A.A.; Jobin, C.; Sang, S. Avenanthramide metabotype from whole-grain oat intake is influenced by Faecalibacterium prausnitzii in healthy adults. J. Nutr. 2021, 151, 1426–1435. [Google Scholar] [CrossRef] [PubMed]
- Sumayya, P.C.; Babu, G.M.; Muraleedharan, K. Quantum chemical investigation of the antiradical property of avenanthramides, oat phenolics. Heliyon 2021, 7, e06125. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Hernandez, O.; Pereira-Caro, G.; Borges, G.; Crozier, A.; Olsson, O. Characterization and antioxidant activity of avenanthramides from selected oat lines developed by mutagenesis technique. Food Chem. 2020, 343, 128408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, T.; Zhang, Y.; Liu, T.; Gagnon, G.; Ebrahim, J.; Johnson, J.; Chu, Y.F.; Ji, L.L. Avenanthramide supplementa-tion reduces eccentric exercise-induced inflammation in young men and women. J. Int. Soc. Sports Nutr. 2020, 17, 41. [Google Scholar] [CrossRef]
- Jagr, M.; Dvoracek, V.; Cepkova, P.H.; Dolezalova, J. Comprehensive analysis of oat avenanthramides using hybrid quad-rupole-Orbitrap mass spectrometry: Possible detection of new compounds. Rapid Commun. Mass Spectrom. 2020, 34, e8718. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Tang, Y.; Zhao, Y.; Sang, S. Quantitative analysis and anti-inflammatory activity evaluation of the A-type avenanthramides in commercial sprouted oat products. J. Agric. Food Chem. 2020, 68, 13068–13075. [Google Scholar] [CrossRef]
- Finetti, F.; Moglia, A.; Schiavo, I.; Donnini, S.; Berta, G.N.; Di Scipio, F.; Perrelli, A.; Fornelli, C.; Trabalzini, L.; Retta, S.F. Yeast-derived recombinant avenanthramides inhibit proliferation, migration and epithelial mesenchymal transition of colon cancer cells. Nutrients 2018, 10, 1159. [Google Scholar] [CrossRef]
- Wu, W.; Tang, Y.; Yang, J.; Idehen, E.; Sang, S. Avenanthramide aglycones and glucosides in oat bran: Chemical profile, levels in commercial oat products, and cytotoxicity to human colon cancer cells. J. Agric. Food Chem. 2018, 66, 8005–8014. [Google Scholar] [CrossRef]
- Yang, J.; Ou, B.; Wise, M.L.; Chu, Y. In vitro total antioxidant capacity and anti-inflammatory activity of three common oat-derived avenanthramides. Food Chem. 2014, 160, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Shin, W.S.; Yeo, D.; Lim, W.; Zhang, T.; Ji, L.L. Anti-inflammatory effect of avenanthramides via NF-kappaB pathways in C2C12 skeletal muscle cells. Free Radic. Biol. Med. 2018, 117, 30–36. [Google Scholar] [CrossRef]
- Tripathi, V.; Singh, A.; Ashraf, M.T. Avenanthramides of oats: Medicinal importance and future perspectives. Pharmacogn. Rev. 2018, 12, 66–71. [Google Scholar] [CrossRef]
- Giacinti, C.; Giordano, A. RB and cell cycle progression. Oncogene 2006, 25, 5220–5227. [Google Scholar] [CrossRef]
- Turrini, E.; Maffei, F.; Milelli, A.; Calcabrini, C.; Fimognari, C. Overview of the anticancer profile of avenanthramides from oat. Int. J. Mol. Sci. 2019, 20, 4536. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zhu, Y.; Yerke, A.; Wise, M.L.; Johnson, J.; Chu, Y.; Sang, S. Oat avenanthramides induce heme oxygenase-1 expression via Nrf2-mediated signaling in HK-2 cells. Mol. Nutr. Food Res. 2015, 59, 2471–2479. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.S.; Lee, D.Y.; Kim, H.S.; Park, S.C.; Park, J.T.; Kim, H.S.; Oh, W.K.; Cho, K.A. Identification of a novel senomorphic agent, avenanthramide C, via the suppression of the senescence-associated secretory phenotype. Mech. Ageing Dev. 2020, 192, 111355. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.G.Q.; Condado, D.M.; Hernandez, B.G.; Gonzalez-Horta, A. Avenanthramide-C prevents amyloid formation of bovine serum albumin. Biophys. Chem. 2020, 263, 106391. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-Y.; Choi, H.-J.; Park, T.; Lee, M.-J.; Lim, H.-S.; Yang, W.-S.; Hwang, C.-W.; Park, D.; Kim, C.-H. Inhibitory effect of avenanthramides (Avn) on tyrosinase activity and melanogenesis in α-MSH-activated SK-MEL-2 cells: In vitro and in silico analysis. Int. J. Mol. Sci. 2021, 22, 7814. [Google Scholar] [CrossRef] [PubMed]
- Becker, L.C.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G.; Shank, R.C., Jr.; Slaga, T.J.; Snyder, P.W.; et al. Safety assessment of Avena sativa (oat)-derived ingredients as used in cosmetics. Int. J. Toxicol. 2019, 38, 23S–47S. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, E.S.; Mari, M.; Antonini, E.; Palma, F.; Ninfali, P. Natural and synthetic avenanthramides activate caspases 2, 8, 3 and downregulate hTERT, MDR1 and COX-2 genes in CaCo-2 and Hep3B cancer cells. Food Funct. 2018, 9, 2913–2921. [Google Scholar] [CrossRef]
- Ham, H.; Woo, K.S.; Park, J.-Y.; Lee, B.; Choi, Y.-H.; Lee, C.; Kim, W.H.; Lee, J.; Lee, Y.-Y. Antioxidant and anti-proliferative activities of oats under different solvent extraction conditions. J. Korean Soc. Food Sci. Nutr. 2016, 45, 918–922. [Google Scholar] [CrossRef][Green Version]
- Chimento, A.; Santarsiero, A.; Iacopetta, D.; Ceramella, J.; De Luca, A.; Infantino, V.; Parisi, O.; Avena, P.; Bonomo, M.; Saturnino, C.; et al. A phenylacetamide resveratrol derivative exerts inhibitory effects on breast cancer cell growth. Int. J. Mol. Sci. 2021, 22, 5255. [Google Scholar] [CrossRef]
- Sang, S.; Chu, Y. Whole grain oats, more than just a fiber: Role of unique phytochemicals. Mol. Nutr. Food Res. 2017, 61, 1600715. [Google Scholar] [CrossRef]
- Aparicio-Garcia, N.; Martinez-Villaluenga, C.; Frias, J.; Penas, E. Sprouted oat as a potential gluten-free ingredient with en-hanced nutritional and bioactive properties. Food Chem. 2021, 338, 127972. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Sang, S. Triterpenoid saponins in oat bran and their levels in commercial oat products. J. Agric. Food Chem. 2020, 68, 6381–6389. [Google Scholar] [CrossRef]
- Damazo-Lima, M.; Rosas-Perez, G.; Reynoso-Camacho, R.; Perez-Ramirez, I.F.; Rocha-Guzman, N.E.; de Los Rios, E.A.; Ra-mos-Gomez, M. Chemopreventive effect of the germinated oat and its phenolic-AVA extract in azoxymethane/dextran sulfate sodium (AOM/DSS) model of colon carcinogenesis in mice. Foods 2020, 9, 169. [Google Scholar] [CrossRef]
- Aborus, N.E.; Šaponjac, V.T.; Canadanovic-Brunet, J.; Cetkovic, G.; Hidalgo, A.; Vulic, J.; Seregelj, V. Sprouted and freeze-dried wheat and oat seeds—Phytochemical profile and in vitro biological activities. Chem. Biodivers. 2018, 15, e1800119. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.-B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef]
- Singh, A.K.; Rehal, J.; Kaur, A.; Jyot, G. Enhancement of attributes of cereals by germination and fermentation: A review. Crit. Rev. Food Sci. Nutr. 2013, 55, 1575–1589. [Google Scholar] [CrossRef]
- Kamau, E.H.; Nkhata, S.G.; Ayua, E.O. Extrusion and nixtamalization conditions influence the magnitude of change in the nutrients and bioactive components of cereals and legumes. Food Sci. Nutr. 2020, 8, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Rico, D.; Peñas, E.; García, M.D.C.; Martínez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martín-Diana, A.B. Sprouted barley flour as a nutritious and functional ingredient. Foods 2020, 9, 296. [Google Scholar] [CrossRef] [PubMed]
- Katina, K.; Liukkonen, K.-H.; Kaukovirta-Norja, A.; Adlercreutz, H.; Heinonen, S.-M.; Lampi, A.-M.; Pihlava, J.-M.; Poutanen, K. Fermentation-induced changes in the nutritional value of native or germinated rye. J. Cereal Sci. 2007, 46, 348–355. [Google Scholar] [CrossRef]
- Ding, J.; Johnson, J.; Chu, Y.F.; Feng, H. Enhancement of γ-aminobutyric acid, avenanthramides, and other health-promoting metabolites in germinating oats (Avena sativa L.) treated with and without power ultrasound. Food Chem. 2019, 283, 239–247. [Google Scholar] [CrossRef] [PubMed]
- de Bruijn, W.J.C.; van Dinteren, S.; Gruppen, H.; Vincken, J.P. Mass spectrometric characterisation of avenanthramides and enhancing their production by germination of oat (Avena sativa). Food Chem. 2019, 277, 682–690. [Google Scholar] [CrossRef]
- Kordialik-Bogacka, E.; Bogdan, P.; Diowksz, A. Malted and unmalted oats in brewing. J. Inst. Brew. 2014, 120, 390–398. [Google Scholar] [CrossRef]
- Lin, C.S.K.; Pfaltzgraff, L.A.; Herrero-Davila, L.; Mubofu, E.B.; Abderrahim, S.; Clark, J.H.; Koutinas, A.A.; Kopsahelis, N.; Stamatelatou, K.; Dickson, F.; et al. Food waste as a valuable resource for the production of chemicals, materials and fuels. Current situation and global perspective. Energy Environ. Sci. 2012, 6, 426–464. [Google Scholar] [CrossRef]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef]
- Torres-León, C.; Ramírez-Guzman, N.; Londoño-Hernandez, L.; Martinez-Medina, G.A.; Díaz-Herrera, R.; Navarro-Macias, V.; Alvarez-Pérez, O.B.; Picazo, B.; Villarreal-Vázquez, M.; Ascacio-Valdes, J.; et al. Food waste and byproducts: An opportunity to minimize malnutrition and hunger in developing countries. Front. Sustain. Food Syst. 2018, 2, 52. [Google Scholar] [CrossRef]
- Rao, M.; Bast, A.; de Boer, A. Valorized food processing by-products in the EU: Finding the balance between safety, nutrition, and sustainability. Sustainability 2021, 13, 4428. [Google Scholar] [CrossRef]
- Reddy, M.; Ponnamma, D.; Choudhary, R.; Sadasivuni, K. A comparative review of natural and synthetic biopolymer composite scaffolds. Polymers 2021, 13, 1105. [Google Scholar] [CrossRef]
- Saric, L.; Filipcev, B.; Simurina, O.; Plavsic, D.; Saric, B.; Lazarevic, J.; Milovanovic, I. Sugar beet molasses: Properties and applications in osmotic dehydration of fruits and vegetables. Food Feed Res. 2016, 43, 135–144. [Google Scholar] [CrossRef]
- Filipcev, B.; Levic, L.; Bodroza-Solarov, M.; Mišljenović, N.; Koprivica, G. Quality characteristics and antioxidant properties of breads supplemented with sugar beet molasses-based ingredients. Int. J. Food Prop. 2010, 13, 1035–1053. [Google Scholar] [CrossRef]
- Luithui, Y.; Nisha, R.B.; Meera, M.S. Cereal by-products as an important functional ingredient: Effect of processing. J. Food Sci. Technol. 2018, 56, 1–11. [Google Scholar] [CrossRef]
- Faustino, A.M.; Veiga, M.; Sousa, P.; Costa, E.M.; Silva, S.; Pintado, M. Agro-food byproducts as a new source of natural food additives. Molecules 2019, 24, 1056. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Lopez, C.; Fraga-Corral, M.; Carpena, M.; García-Oliveira, P.; Echave, J.; Pereira, A.G.; Lourenço-Lopes, C.; Prieto, M.A.; Simal-Gandara, J. Agriculture waste valorisation as a source of antioxidant phenolic compounds within a circular and sustainable bioeconomy. Food Funct. 2020, 11, 4853–4877. [Google Scholar] [CrossRef] [PubMed]
- Iriondo-DeHond, M.; Miguel, E.; Del Castillo, M.D. Food byproducts as sustainable ingredients for innovative and healthy dairy foods. Nutrients 2018, 10, 1358. [Google Scholar] [CrossRef] [PubMed]
- Guillard, V.; Gaucel, S.; Fornaciari, C.; Angellier-Coussy, H.; Buche, P.; Gontard, N. The next generation of sustainable food packaging to preserve our environment in a circular economy context. Front. Nutr. 2018, 5, 121. [Google Scholar] [CrossRef]
- Marshall, A.H.; Cowan, A.A.; Edwards, S.; Griffiths, I.M.; Howarth, C.; Langdon, T.; White, E. Crops that feed the world 9. Oats- a cereal crop for human and livestock feed with industrial applications. Food Secur. 2013, 5, 13–33. [Google Scholar] [CrossRef]
- Pasha, I.; Ahmad, F.; Usman, M. Elucidation of morphological characteristics, crystallinity, and molecular structures of native and enzyme modified cereal brans. J. Food Biochem. 2021, 45, e13768. [Google Scholar] [CrossRef]
- Qiao, Q.; Chen, L.; Li, X.; Lu, X.; Xu, Q. Roles of dietary bioactive peptides in redox balance and metabolic disorders. Oxid. Med. Cell. Longev. 2021, 2021, 5582245. [Google Scholar] [CrossRef]
- Esfandi, R.; Seidu, I.; Willmore, W.; Tsopmo, A. Antioxidant, pancreatic lipase, and alpha-amylase inhibitory properties of oat bran hydrolyzed proteins and peptides. J. Food Biochem. 2021, e13762. [Google Scholar] [CrossRef]
- Xue, Y.; Cui, L.; Qi, J.; Ojo, O.; Du, X.; Liu, Y.; Wang, X. The effect of dietary fiber (oat bran) supplement on blood pressure in patients with essential hypertension: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2458–2470. [Google Scholar] [CrossRef]
- Gillespie, R.; Ahlborn, G.J. Mechanical, sensory, and consumer evaluation of ketogenic, gluten-free breads. Food Sci. Nutr. 2021, 9, 3327–3335. [Google Scholar] [CrossRef] [PubMed]
- Bergh, C.; Landberg, R.; Andersson, K.; Heyman-Linden, L.; Rascon, A.; Magnuson, A.; Khalili, P.; Karegren, A.; Nilsson, J.; Pirazzi, C.; et al. Effects of Bilberry and Oat Intake on lipids, inflammation and exercise capacity after Acute Myocardial Infarction (BIOAMI): Study protocol for a randomized, double-blind, placebo-controlled trial. Trials 2021, 22, 338. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, O.; Ma, X.; Pasanen, E.; Zhou, P.; Yang, B.; Linderborg, K.M. Sensory characteristics contributing to pleasantness of oat product concepts by Finnish and Chinese consumers. Foods 2020, 9, 1234. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, E.; Karlsson, E.N.; Adlercreutz, P. Warming weather changes the chemical composition of oat hulls. Plant Biol. 2020, 22, 1086–1091. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Lizarazo, C.I.; Tuulos, A.; Jokela, V.; Mäkelä, P.S.A. Sustainable mixed cropping systems for the boreal-nemoral region. Front. Sustain. Food Syst. 2020, 4, 103. [Google Scholar] [CrossRef]
- Leišová-Svobodová, L.; Michel, S.; Tamm, I.; Chourová, M.; Janovska, D.; Grausgruber, H. Diversity and pre-breeding prospects for local adaptation in oat genetic resources. Sustainability 2019, 11, 6950. [Google Scholar] [CrossRef]
- Jan, S.F.; Khan, M.R.; Iqbal, A.; Khan, F.U.; Ali, S. Genetic diversity in exotic oat germplasm & resistance against barley yellow dwarf virus. Saudi J. Biol. Sci. 2020, 27, 2622–2631. [Google Scholar] [PubMed]
- Rispail, N.; Montilla-Bascon, G.; Sanchez-Martin, J.; Flores, F.; Howarth, C.; Langdon, T.; Rubiales, D.; Prats, E. Multi-environmental trials reveal genetic plasticity of oat agronomic traits associated with climate variable changes. Front. Plant Sci. 2018, 9, 1358. [Google Scholar] [CrossRef] [PubMed]
- Capstaff, N.; Miller, A.J. Improving the yield and nutritional quality of forage crops. Front. Plant Sci. 2018, 9, 535. [Google Scholar] [CrossRef]
- Kulkarni, K.P.; Tayade, R.; Asekova, S.; Song, J.T.; Shannon, J.G.; Lee, J.-D. Harnessing the potential of forage legumes, alfalfa, soybean, and cowpea for sustainable agriculture and global food security. Front. Plant Sci. 2018, 9, 1314. [Google Scholar] [CrossRef]
- Rodriguez, J.P.; Rahman, H.; Thushar, S.; Singh, R.K. Healthy and resilient cereals and pseudo-cereals for marginal agriculture: Molecular advances for improving nutrient bioavailability. Front. Genet. 2020, 11, 49. [Google Scholar] [CrossRef]
- Shunmugam, A.S.K.; Kannan, U.; Jiang, Y.; Daba, K.A.; Gorim, L.Y. Physiology based approaches for breeding of next-generation food legumes. Plants 2018, 7, 72. [Google Scholar] [CrossRef] [PubMed]
- McNish, I.G.; Zimmer, C.M.; Susko, A.Q.; Heuschele, D.; Tiede, T.; Case, A.J.; Smith, K.P. Mapping crown rust resistance at multiple time points in elite oat germplasm. Plant Genome 2020, 13, e20007. [Google Scholar] [CrossRef] [PubMed]
- Smulders, M.J.; van de Wiel, C.; Broeck, H.C.V.D.; van der Meer, I.M.; Israel-Hoevelaken, T.; Timmer, R.D.; van Dinter, B.-J.; Braun, S.; Gilissen, L.J. Oats in healthy gluten-free and regular diets: A perspective. Food Res. Int. 2018, 110, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.E.; Zhang, Y.; Omidvar, V.; Sperschneider, J.; Schwessinger, B.; Raley, C.; Palmer, J.M.; Garnica, D.; Upadhyaya, N.; Rathjen, J.; et al. De vovo assembly and phasing of dikaryotic genomes from two isolates of Puccinia coronata f. sp. avenae, the causal agent of oat crown rust. mBio 2018, 9, e01650-17. [Google Scholar] [CrossRef]
- Loskutov, I.G.; Khlestkina, E.K. Wheat, barley, and oat breeding for health benefit components in grain. Plants 2021, 10, 86. [Google Scholar] [CrossRef]
- Stewart, D.; McDougall, G. Oat agriculture, cultivation and breeding targets: Implications for human nutrition and health. Br. J. Nutr. 2014, 112, S50–S57. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Cowling, W.; Pareek, A.; Gupta, K.J.; Singla-Pareek, S.L.; Foyer, C.H. Gaining acceptance of novel plant breeding technologies. Trends Plant Sci. 2021, 26, 575–587. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).