
Palmitoylethanolamide/Baicalein Regulates the Androgen Receptor Signaling and NF-κB/Nrf2 Pathways in Benign Prostatic Hyperplasia

 ,
,  , ,
, ,  , ,
, ,  ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. BPH Induction and Drugs
2.3. Experimental Groups
2.4. Prostate Weight
2.5. Histology
2.6. Staining of Mast Cells
2.7. Western Blot Analysis of 5α-red 2, AR, PSA, TGF-β, Bax, Bcl2, IκBα, Nuclear Factor kappaB (NF-κB), Nrf-2, Heme Oxigenase-1 (HO-1) and Mn-SOD
2.8. Evaluation of Testosterone and DHT Levels in Serum and Prostate
2.9. Evaluation of Cytokine Levels
2.10. Statistical Evaluation
3. Results
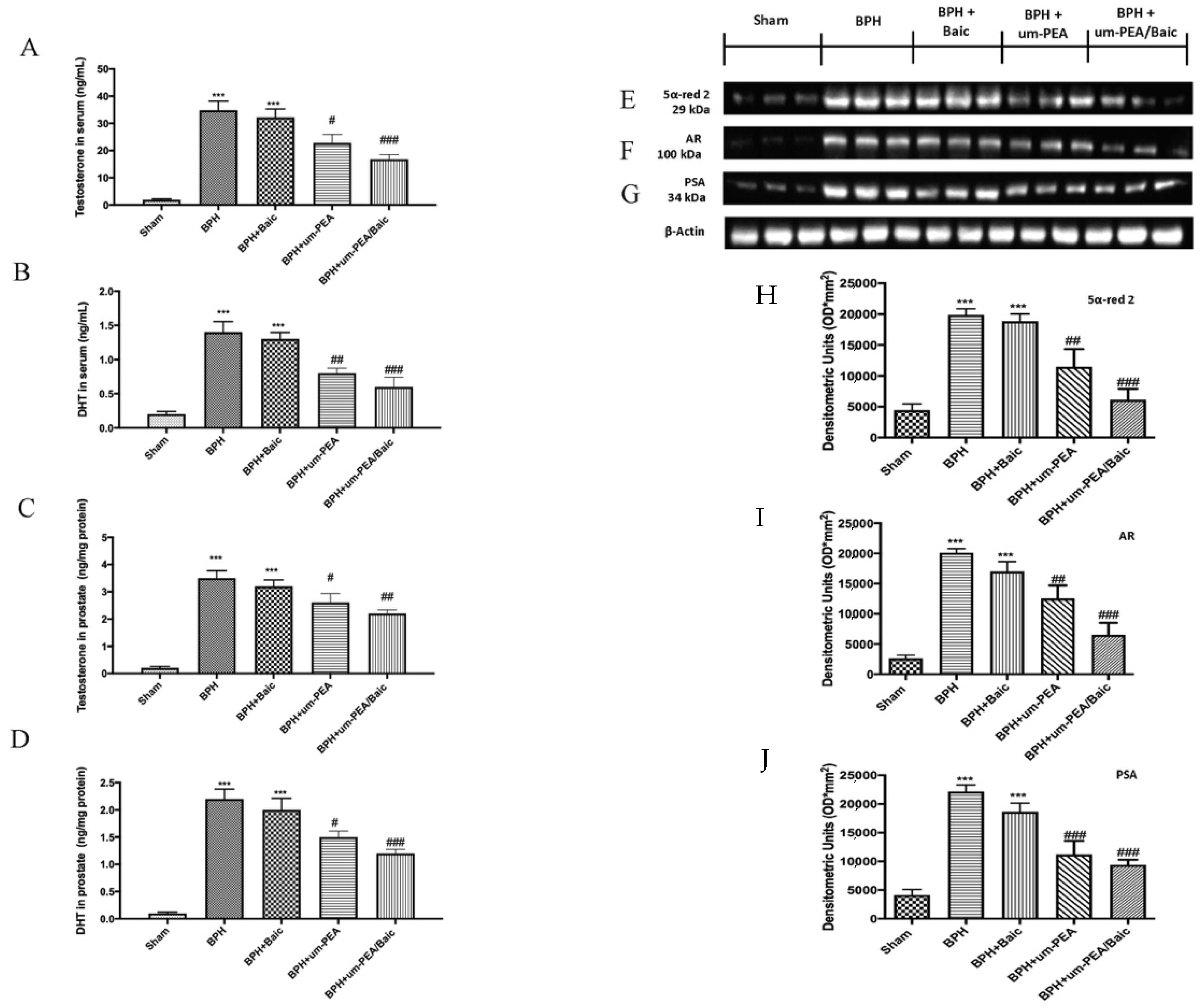
3.1. Effect of um-PEA/Baic on Testosterone and DHT levels and 5α-red 2, AR and PSA Expression
3.2. Effect of um-PEA/Baic on Cell Growth and Apoptotic Pathway
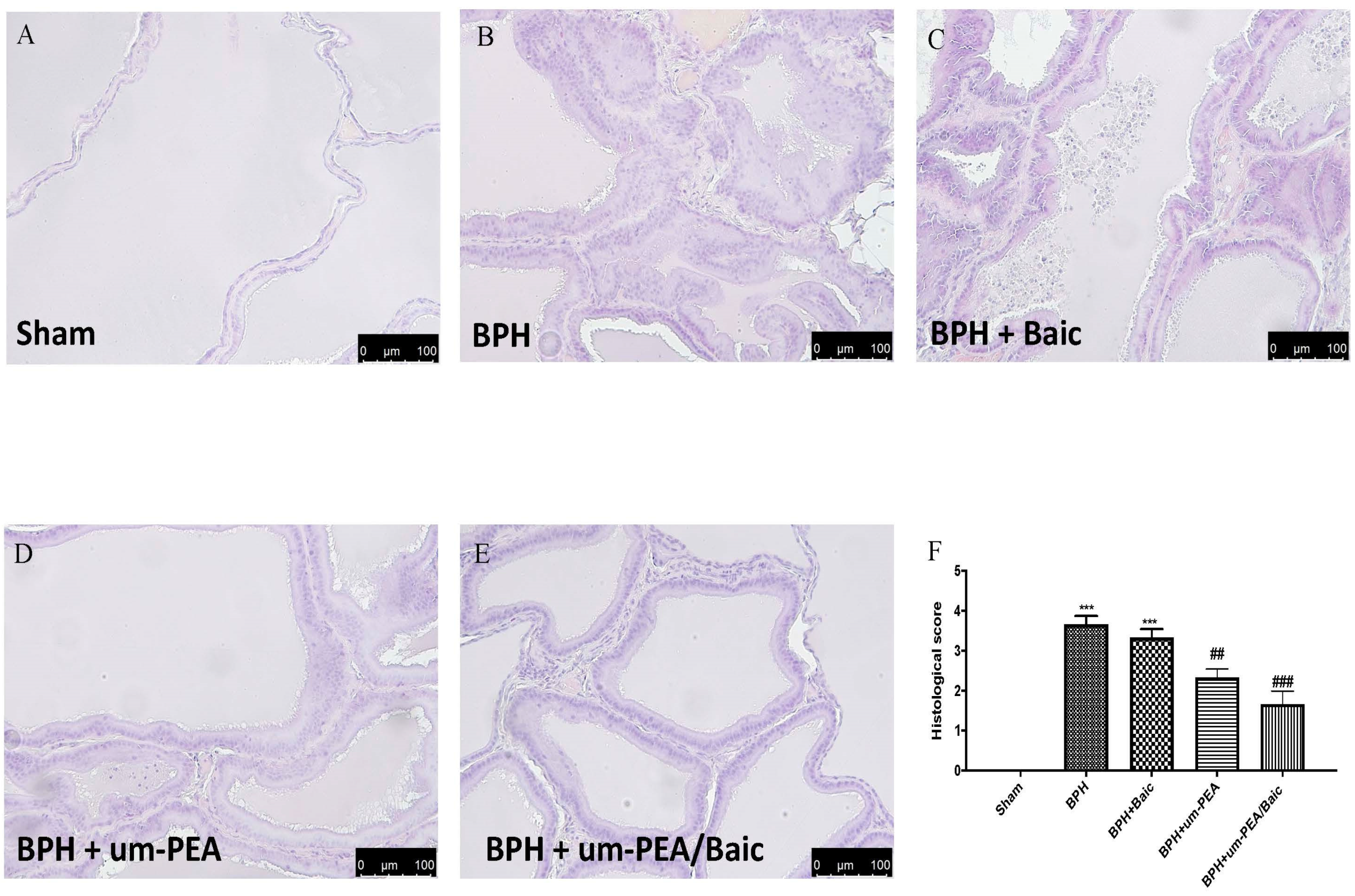
3.3. Effect of um-PEA/Baic on Prostate Morphology
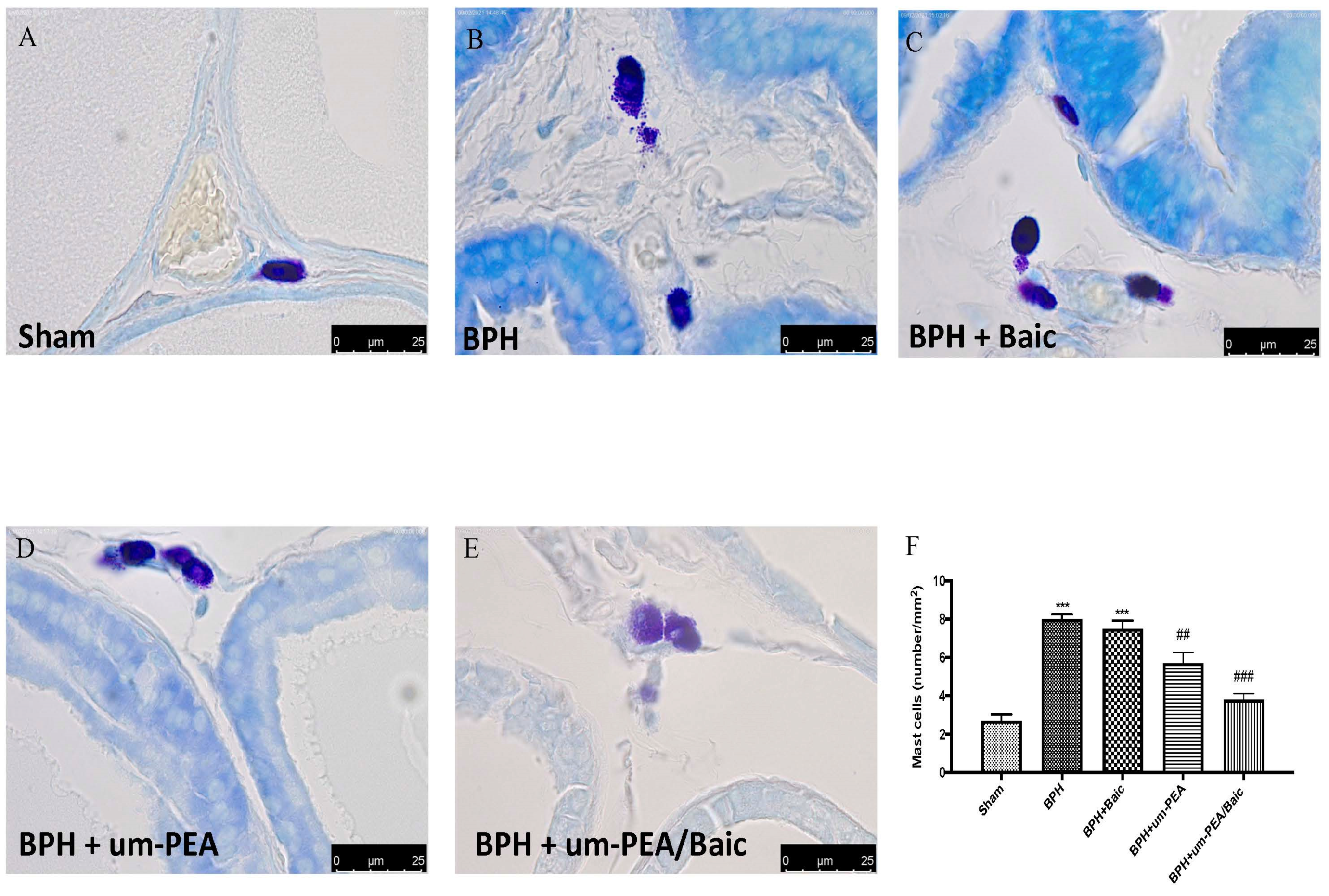
3.4. Effect of um-PEA/Baic on Mast Cell Density
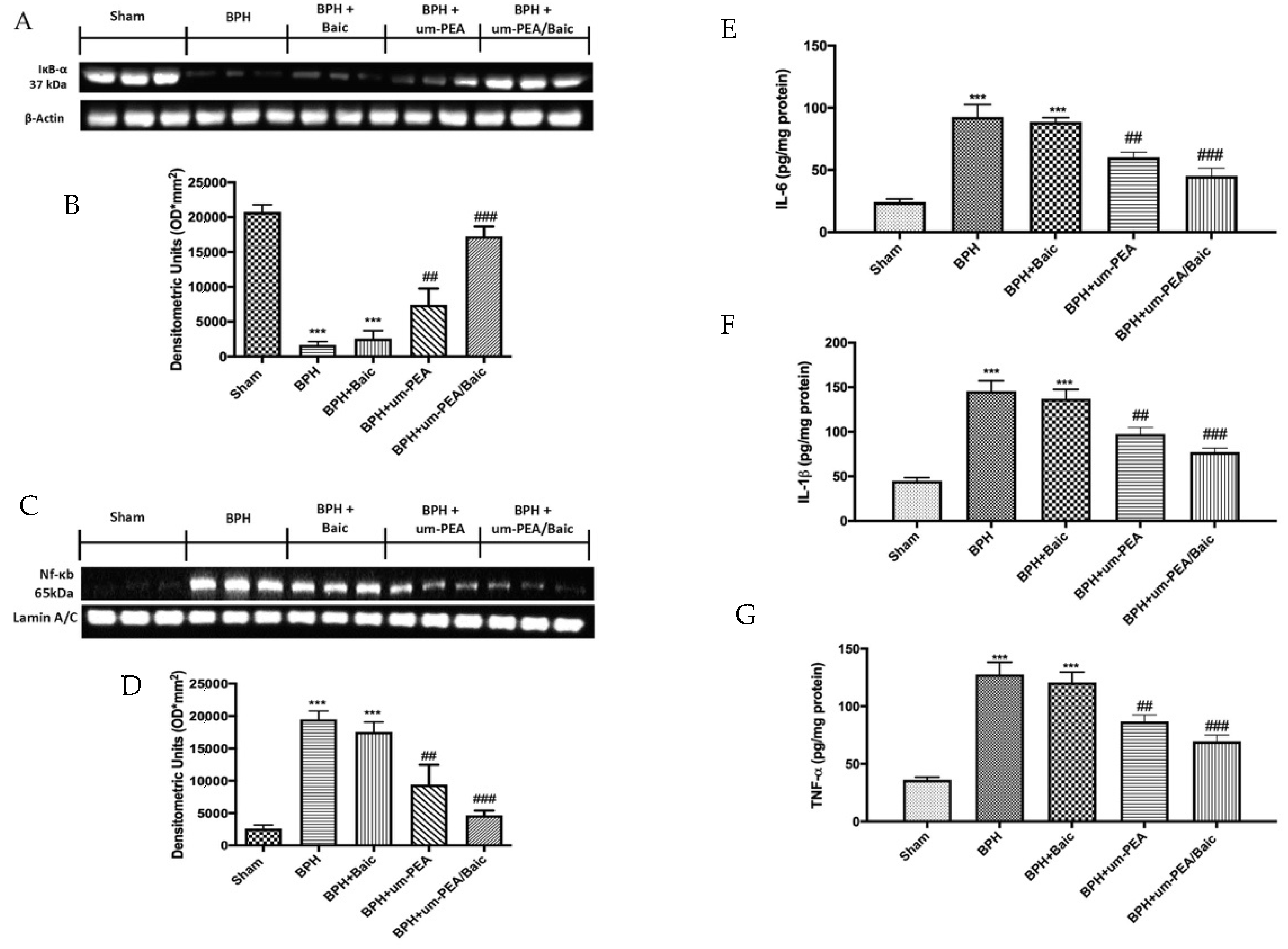
3.5. Effect of um-PEA/Baic on Inflammation Pathway
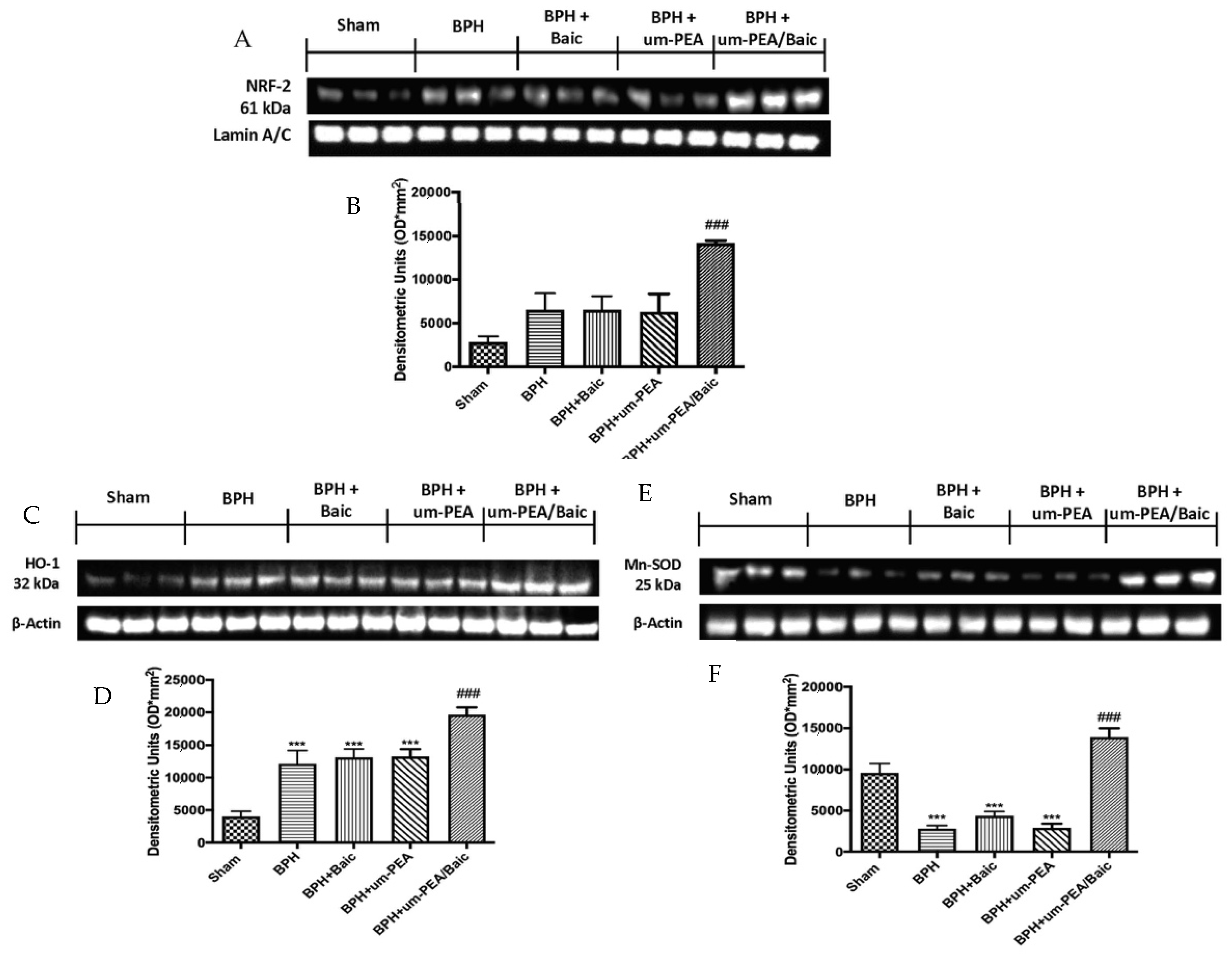
3.6. Effect of um-PEA/Baic on Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fang, T.; Xue, Z.S.; Li, J.X.; Liu, J.K.; Wu, D.; Li, M.Q.; Song, Y.T.; Yun, S.F.; Yan, J. Rauwolfia vomitoria extract suppresses benign prostatic hyperplasia by reducing expression of androgen receptor and 5alpha-reductase in a rat model. J. Integr. Med. 2020, 19, 258–264. [Google Scholar] [CrossRef]
- Minutoli, L.; Rinaldi, M.; Marini, H.; Irrera, N.; Crea, G.; Lorenzini, C.; Puzzolo, D.; Valenti, A.; Pisani, A.; Adamo, E.B.; et al. Apoptotic Pathways Linked to Endocrine System as Potential Therapeutic Targets for Benign Prostatic Hyperplasia. Int. J. Mol. Sci. 2016, 17, 1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peyronnet, B.; Brucker, B.M.; Michel, M.C. Lower Urinary Tract Symptoms: What’s New in Medical Treatment? Eur. Urol. Focus 2018, 4, 17–24. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; Li, W.; Zhang, K.; Yuan, Z.; Cai, Y.; Xu, K.; Zhou, J.; Du, Z. Curcuma oil ameliorates benign prostatic hyperplasia through suppression of the nuclear factor-kappa B signaling pathway in rats. J. Ethnopharmacol. 2020, 2020, 113703. [Google Scholar] [CrossRef]
- Samarinas, M.; Gacci, M.; de la Taille, A.; Gravas, S. Prostatic inflammation: A potential treatment target for male LUTS due to benign prostatic obstruction. Prostate Cancer Prostatic Dis. 2018, 21, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Udensi, U.K.; Tchounwou, P.B. Oxidative stress in prostate hyperplasia and carcinogenesis. J. Exp. Clin. Cancer Res. 2016, 35, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, P.P.; Banerjee, S.; Brown, T.R.; Zirkin, B.R. Androgen action in prostate function and disease. Am. J. Clin. Exp. Urol. 2018, 6, 62–77. [Google Scholar]
- Vickman, R.E.; Franco, O.E.; Moline, D.C.; Vander Griend, D.J.; Thumbikat, P.; Hayward, S.W. The role of the androgen receptor in prostate development and benign prostatic hyperplasia: A review. Asian J. Urol. 2020, 7, 191–202. [Google Scholar] [CrossRef]
- Kang, H.Y.; Lin, H.K.; Hu, Y.C.; Yeh, S.; Huang, K.E.; Chang, C. From transforming growth factor-beta signaling to androgen action: Identification of Smad3 as an androgen receptor coregulator in prostate cancer cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3018–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K.; Seok, H.; Park, H.J.; Jeon, H.S.; Kang, S.W.; Lee, B.C.; Yi, J.; Song, S.Y.; Lee, S.H.; Kim, Y.O.; et al. Inhibitory effect of curcumin on testosterone induced benign prostatic hyperplasia rat model. BMC Complement. Altern. Med. 2015, 15, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traish, A.M. Negative Impact of Testosterone Deficiency and 5alpha-Reductase Inhibitors Therapy on Metabolic and Sexual Function in Men. Adv. Exp. Med. Biol. 2017, 1043, 473–526. [Google Scholar] [CrossRef]
- Petrosino, S.; Di Marzo, V. The pharmacology of palmitoylethanolamide and first data on the therapeutic efficacy of some of its new formulations. Br. J. Pharmacol. 2017, 174, 1349–1365. [Google Scholar] [CrossRef]
- Siracusa, R.; Fusco, R.; Cordaro, M.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Crupi, R.; Genovese, T.; Evangelista, M.; Di Paola, R.; et al. The Protective Effects of Pre- and Post-Administration of Micronized Palmitoylethanolamide Formulation on Postoperative Pain in Rats. Int. J. Mol. Sci. 2020, 21, 7700. [Google Scholar] [CrossRef]
- Cordaro, M.; Scuto, M.; Siracusa, R.; D’Amico, R.; Filippo Peritore, A.; Gugliandolo, E.; Fusco, R.; Crupi, R.; Impellizzeri, D.; Pozzebon, M.; et al. Effect of N-palmitoylethanolamine-oxazoline on comorbid neuropsychiatric disturbance associated with inflammatory bowel disease. FASEB J. 2020, 34, 4085–4106. [Google Scholar] [CrossRef]
- Di Paola, R.; Cordaro, M.; Crupi, R.; Siracusa, R.; Campolo, M.; Bruschetta, G.; Fusco, R.; Pugliatti, P.; Esposito, E.; Cuzzocrea, S. Protective Effects of Ultramicronized Palmitoylethanolamide (PEA-um) in Myocardial Ischaemia and Reperfusion Injury in vivo. Shock 2016, 46, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Peritore, A.F.; Cordaro, M.; Gugliandolo, E.; Siracusa, R.; Crupi, R.; D’Amico, R.; Fusco, R.; Evangelista, M.; Cuzzocrea, S.; et al. The neuroprotective effects of micronized PEA (PEA-m) formulation on diabetic peripheral neuropathy in mice. FASEB J. 2019, 33, 11364–11380. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain. Int. J. Mol. Sci. 2020, 21, 5330. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Impellizzeri, D.; Fusco, R.; Cordaro, M.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. Ultramicronized palmitoylethanolamide (PEA-um®) in the treatment of idiopathic pulmonary fibrosis. Pharmacol. Res. 2016, 111, 405–412. [Google Scholar] [CrossRef]
- Tripathi, S.S.; Kumar, R.; Bissoyi, A.; Rizvi, S.I. Baicalein maintains redox balance in experimental hyperlipidemic rats. Arch. Physiol. Biochem. 2020, 2020, 1760890. [Google Scholar] [CrossRef]
- Park, C.; Choi, E.O.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Yoo, Y.H.; Park, H.T.; Choi, Y.H. Protective Effect of Baicalein on Oxidative Stress-induced DNA Damage and Apoptosis in RT4-D6P2T Schwann Cells. Int. J. Med. Sci. 2019, 16, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Qiao, D.; Jin, J.; Xing, J.; Zhang, Y.; Jia, N.; Ren, X.; Lin, Z.; Jin, N.; Chen, L.; Piao, Y. Baicalein Inhibits Gastric Cancer Cell Proliferation and Migration through a FAK Interaction via AKT/mTOR Signaling. Am. J. Chin. Med. 2021, 49, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Chen, Q.; Liu, Y.; Wen, X. Baicalein suppresses the androgen receptor (AR)-mediated prostate cancer progression via inhibiting the AR N-C dimerization and AR-coactivators interaction. Oncotarget 2017, 8, 105561–105573. [Google Scholar] [CrossRef] [Green Version]
- Susmitha, G.D.; Miyazato, K.; Ogura, K.; Yokoyama, S.; Hayakawa, Y. Anti-metastatic Effects of Baicalein by Targeting STAT3 Activity in Breast Cancer Cells. Biol. Pharm. Bull. 2020, 43, 1899–1905. [Google Scholar] [CrossRef]
- Cordaro, M.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Fusco, R.; Inferrera, A.; Esposito, E.; Di Paola, R.; Cuzzocrea, S. Effects of a co-micronized composite containing palmitoylethanolamide and polydatin in an experimental model of benign prostatic hyperplasia. Toxicol. Appl. Pharmacol. 2017, 329, 231–240. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, R.; Fusco, R.; Gugliandolo, E.; Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Peritore, A.F.; Crupi, R.; Cuzzocrea, S.; Di Paola, R. Effects of a new compound containing Palmitoylethanolamide and Baicalein in myocardial ischaemia/reperfusion injury in vivo. Phytomedicine 2019, 54, 27–42. [Google Scholar] [CrossRef]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; Evangelista, M.; Granese, R.; Cuzzocrea, S. Co-micronized Palmitoylethanolamide/Polydatin Treatment Causes Endometriotic Lesion Regression in a Rodent Model of Surgically Induced Endometriosis. Front. Pharmacol. 2016, 7, 382. [Google Scholar] [CrossRef] [Green Version]
- Peritore, A.F.; D’Amico, R.; Cordaro, M.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Di Paola, R.; Cuzzocrea, S.; et al. PEA/Polydatin: Anti-Inflammatory and Antioxidant Approach to Counteract DNBS-Induced Colitis. Antioxidants 2021, 10, 464. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Impellizzeri, D.; D’Amico, R.; Peritore, A.F.; Crupi, R.; Gugliandolo, E.; Fusco, R.; Di Paola, R.; Schievano, C.; et al. Safety and efficacy of a new micronized formulation of the ALIAmide palmitoylglucosamine in preclinical models of inflammation and osteoarthritis pain. Arthritis Res. Ther. 2019, 21, 254. [Google Scholar] [CrossRef] [Green Version]
- Sarbishegi, M.; Khajavi, O.; Arab, M.R. Withania coagulans Extract Induces Cell Apoptosis and Inhibits COX-2 Expression in a Rat Model of Benign Prostatic Hyperplasia. Nephrourol. Mon. 2016, 8, e39284. [Google Scholar] [CrossRef] [Green Version]
- Gugliandolo, E.; Fusco, R.; D’Amico, R.; Militi, A.; Oteri, G.; Wallace, J.L.; Di Paola, R.; Cuzzocrea, S. Anti-inflammatory effect of ATB-352, a H2S -releasing ketoprofen derivative, on lipopolysaccharide-induced periodontitis in rats. Pharmacol. Res. 2018, 132, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Fusco, R.; Ginestra, G.; D’Amico, R.; Bisignano, C.; Mandalari, G.; Cuzzocrea, S.; Di Paola, R. Involvement of TLR4 and PPAR-alpha Receptors in Host Response and NLRP3 Inflammasome Activation, Against Pulmonary Infection with Pseudomonas Aeruginosa. Shock 2019, 51, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Peritore, A.F.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Crupi, R.; Genovese, T.; Impellizzeri, D.; Cuzzocrea, S.; et al. Ultramicronized Palmitoylethanolamide and Paracetamol, a New Association to Relieve Hyperalgesia and Pain in a Sciatic Nerve Injury Model in Rat. Int. J. Mol. Sci. 2020, 21, 3509. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Fusco, R.; Peritore, A.F.; Cordaro, M.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. The Antioxidant and Anti-Inflammatory Properties of Anacardium occidentale L. Cashew Nuts in a Mouse Model of Colitis. Nutrients 2020, 12, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusco, R.; Siracusa, R.; D’Amico, R.; Peritore, A.F.; Cordaro, M.; Gugliandolo, E.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Melatonin Plus Folic Acid Treatment Ameliorates Reserpine-Induced Fibromyalgia: An Evaluation of Pain, Oxidative Stress, and Inflammation. Antioxidants 2019, 8, 628. [Google Scholar] [CrossRef] [Green Version]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Biochemical Evaluation of the Antioxidant Effects of Hydroxytyrosol on Pancreatitis-Associated Gut Injury. Antioxidants 2020, 9, 781. [Google Scholar] [CrossRef]
- Lee, G.; Shin, J.; Choi, H.; Jo, A.; Pan, S.; Bae, D.; Lee, Y.; Choi, C. Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5alpha-Reductase and Androgen Receptor Activities in a Rat Model. Nutrients 2017, 9, 1070. [Google Scholar] [CrossRef] [Green Version]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Campolo, M.; Latteri, S.; Carughi, A.; Mandalari, G.; Cuzzocrea, S. The Antioxidant Activity of Pistachios Reduces Cardiac Tissue Injury of Acute Ischemia/Reperfusion (I/R) in Diabetic Streptozotocin (STZ)-Induced Hyperglycaemic Rats. Front. Pharmacol. 2018, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Cuzzocrea, S.; Mazzon, E.; Esposito, E.; Muia, C.; Abdelrahman, M.; Di Paola, R.; Crisafulli, C.; Bramanti, P.; Thiemermann, C. Glycogen synthase kinase-3beta inhibition attenuates the development of ischaemia/reperfusion injury of the gut. Intensive Care Med. 2007, 33, 880–893. [Google Scholar] [CrossRef]
- Cordaro, M.; Fusco, R.; D’Amico, R.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Mandalari, G.; Cuzzocrea, S.; et al. Cashew (Anacardium occidentale L.) Nuts Modulate the Nrf2 and NLRP3 Pathways in Pancreas and Lung after Induction of Acute Pancreatitis by Cerulein. Antioxidants 2020, 9, 992. [Google Scholar] [CrossRef]
- Choi, Y.J.; Fan, M.; Tang, Y.; Yang, H.P.; Hwang, J.Y.; Kim, E.K. In Vivo Effects of Polymerized Anthocyanin from Grape Skin on Benign Prostatic Hyperplasia. Nutrients 2019, 11, 2444. [Google Scholar] [CrossRef] [Green Version]
- Gacci, M.; Eardley, I.; Giuliano, F.; Hatzichristou, D.; Kaplan, S.A.; Maggi, M.; McVary, K.T.; Mirone, V.; Porst, H.; Roehrborn, C.G. Critical analysis of the relationship between sexual dysfunctions and lower urinary tract symptoms due to benign prostatic hyperplasia. Eur. Urol. 2011, 60, 809–825. [Google Scholar] [CrossRef]
- Heinlein, C.A.; Chang, C. The roles of androgen receptors and androgen-binding proteins in nongenomic androgen actions. Mol. Endocrinol. 2002, 16, 2181–2187. [Google Scholar] [CrossRef]
- Carson, C., 3rd; Rittmaster, R. The role of dihydrotestosterone in benign prostatic hyperplasia. Urology 2003, 61, 2–7. [Google Scholar] [CrossRef]
- Hillebrand, A.C.; Pizzolato, L.S.; Neto, B.S.; Branchini, G.; Brum, I.S. Androgen receptor isoforms expression in benign prostatic hyperplasia and primary prostate cancer. PLoS ONE 2018, 13, e0200613. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Wang, X.; Jiang, C.; Ruan, Y.; Xia, S.; Wang, X. The androgen receptor plays different roles in macrophage-induced proliferation in prostate stromal cells between transitional and peripheral zones of benign prostatic hypertrophy. EXCLI J. 2017, 16, 939–948. [Google Scholar] [CrossRef]
- Kaplan, S.A. Re: Impact of 5alpha-Reductase Inhibitor and alpha-Blocker Therapy for Benign Prostatic Hyperplasia on Prostate Cancer Incidence and Mortality. J. Urol. 2020, 203, 856–857. [Google Scholar] [CrossRef]
- Rho, J.; Seo, C.S.; Park, H.S.; Jeong, H.Y.; Moon, O.S.; Seo, Y.W.; Son, H.Y.; Won, Y.S.; Kwun, H.J. Asteris Radix et Rhizoma suppresses testosterone-induced benign prostatic hyperplasia in rats by regulating apoptosis and inflammation. J. Ethnopharmacol. 2020, 255, 112779. [Google Scholar] [CrossRef]
- Kim, H.J.; Jin, B.R.; An, H.J. Psoralea corylifolia L. extract ameliorates benign prostatic hyperplasia by regulating prostate cell proliferation and apoptosis. J. Ethnopharmacol. 2021, 273, 113844. [Google Scholar] [CrossRef]
- Yeewa, R.; Sakuludomkan, W.; Kiriya, C.; Khanaree, C.; Chewonarin, T. Attenuation of benign prostatic hyperplasia by hydrophilic active compounds from pigmented rice in a testosterone implanted rat model. Food Funct. 2020, 11, 1585–1598. [Google Scholar] [CrossRef]
- Kim, E.H.; Brockman, J.A.; Andriole, G.L. The use of 5-alpha reductase inhibitors in the treatment of benign prostatic hyperplasia. Asian J. Urol. 2018, 5, 28–32. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, J.; Xue, Z.; Sun, J.; Huang, Z.; Jing, Y.; Han, B.; Shen, B.; Yan, J.; Huang, R. Pao Pereira extract suppresses benign prostatic hyperplasia by inhibiting inflammation-associated NFkappaB signaling. BMC Complement. Med. Ther. 2020, 20, 150. [Google Scholar] [CrossRef]
- D’Amico, R.; Siracusa, R.; Fusco, R.; Cordaro, M.; Genovese, T.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Protective effects of Colomast®, A New Formulation of Adelmidrol and Sodium Hyaluronate, in A Mouse Model of Acute Restraint Stress. Int. J. Mol. Sci. 2020, 21, 8136. [Google Scholar] [CrossRef]
- Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; D’Amico, R.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. Consumption of Anacardium Occidentale L. (Cashew Nuts) Inhibits Oxidative Stress through Modulation of the Nrf2/HO-1 and NF-kB Pathways. Molecules 2020, 25, 4426. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Scuto, M.; Crupi, R.; Mandalari, G.; et al. Cashew (Anacardium occidentale L.) Nuts Counteract Oxidative Stress and Inflammation in an Acute Experimental Model of Carrageenan-Induced Paw Edema. Antioxidants 2020, 9, 660. [Google Scholar] [CrossRef]
- Eid, B.G.; Abdel-Naim, A.B. Piceatannol Attenuates Testosterone-Induced Benign Prostatic Hyperplasia in Rats by Modulation of Nrf2/HO-1/NFkappaB Axis. Front. Pharmacol. 2020, 11, 614897. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.; Genovese, T.; Cordaro, M.; Siracusa, R.; Gugliandolo, E.; Peritore, A.F.; Interdonato, L.; Crupi, R.; Cuzzocrea, S.; Di Paola, R.; et al. Palmitoylethanolamide/Baicalein Regulates the Androgen Receptor Signaling and NF-κB/Nrf2 Pathways in Benign Prostatic Hyperplasia. Antioxidants 2021, 10, 1014. https://doi.org/10.3390/antiox10071014
D’Amico R, Genovese T, Cordaro M, Siracusa R, Gugliandolo E, Peritore AF, Interdonato L, Crupi R, Cuzzocrea S, Di Paola R, et al. Palmitoylethanolamide/Baicalein Regulates the Androgen Receptor Signaling and NF-κB/Nrf2 Pathways in Benign Prostatic Hyperplasia. Antioxidants. 2021; 10(7):1014. https://doi.org/10.3390/antiox10071014
Chicago/Turabian StyleD’Amico, Ramona, Tiziana Genovese, Marika Cordaro, Rosalba Siracusa, Enrico Gugliandolo, Alessio Filippo Peritore, Livia Interdonato, Rosalia Crupi, Salvatore Cuzzocrea, Rosanna Di Paola, and et al. 2021. "Palmitoylethanolamide/Baicalein Regulates the Androgen Receptor Signaling and NF-κB/Nrf2 Pathways in Benign Prostatic Hyperplasia" Antioxidants 10, no. 7: 1014. https://doi.org/10.3390/antiox10071014
APA StyleD’Amico, R., Genovese, T., Cordaro, M., Siracusa, R., Gugliandolo, E., Peritore, A. F., Interdonato, L., Crupi, R., Cuzzocrea, S., Di Paola, R., Fusco, R., & Impellizzeri, D. (2021). Palmitoylethanolamide/Baicalein Regulates the Androgen Receptor Signaling and NF-κB/Nrf2 Pathways in Benign Prostatic Hyperplasia. Antioxidants, 10(7), 1014. https://doi.org/10.3390/antiox10071014