
Transit and Metabolic Pathways of Quercetin in Tubular Cells: Involvement of Its Antioxidant Properties in the Kidney



Abstract
:1. Introduction
2. Quercetin Pharmacokinetics
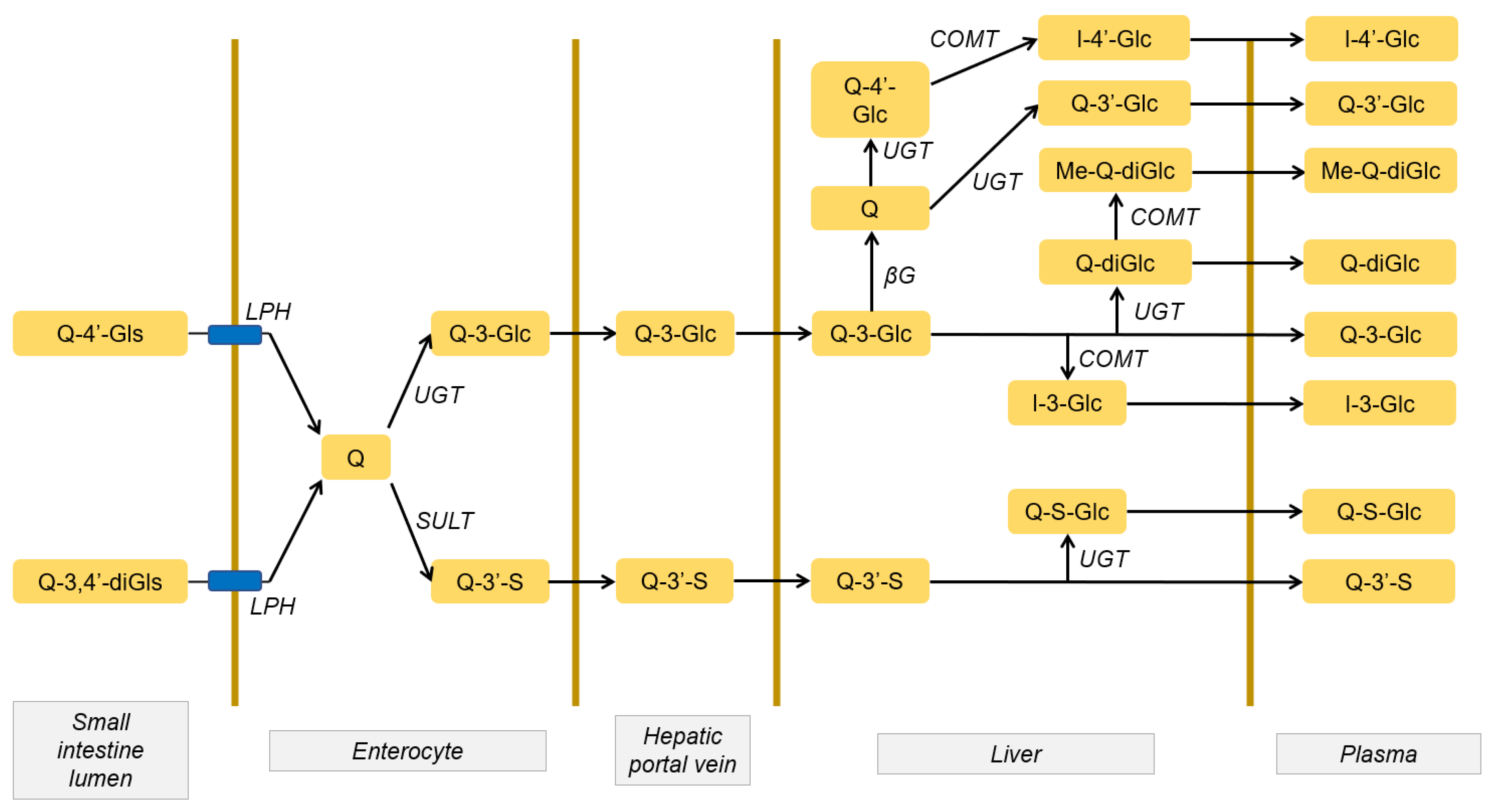
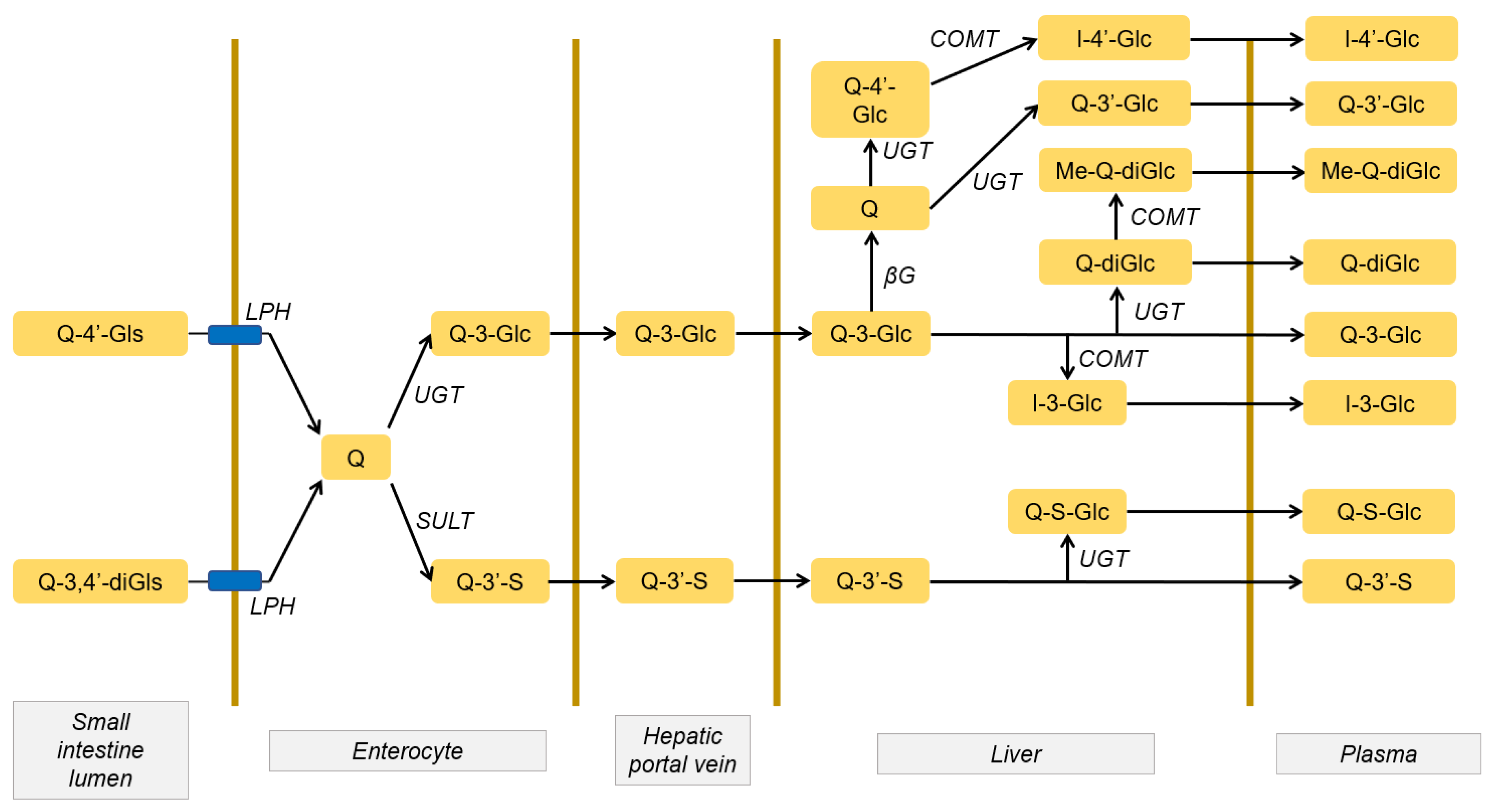
2.1. Quercetin Absorption and Metabolism
2.2. Distribution and Accumulation of Quercetin and its Metabolites
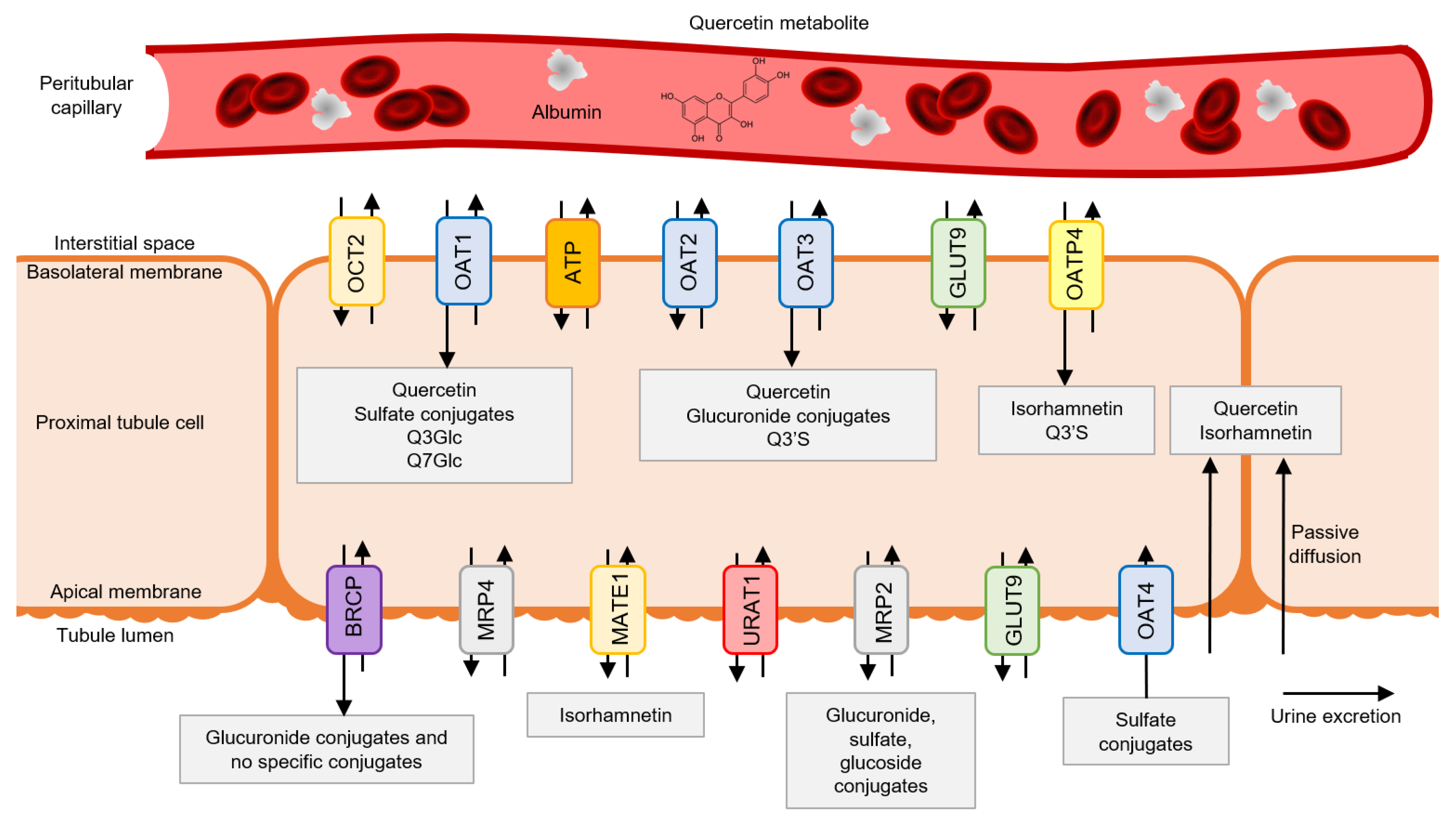
3. Transit of Quercetin through the Kidney
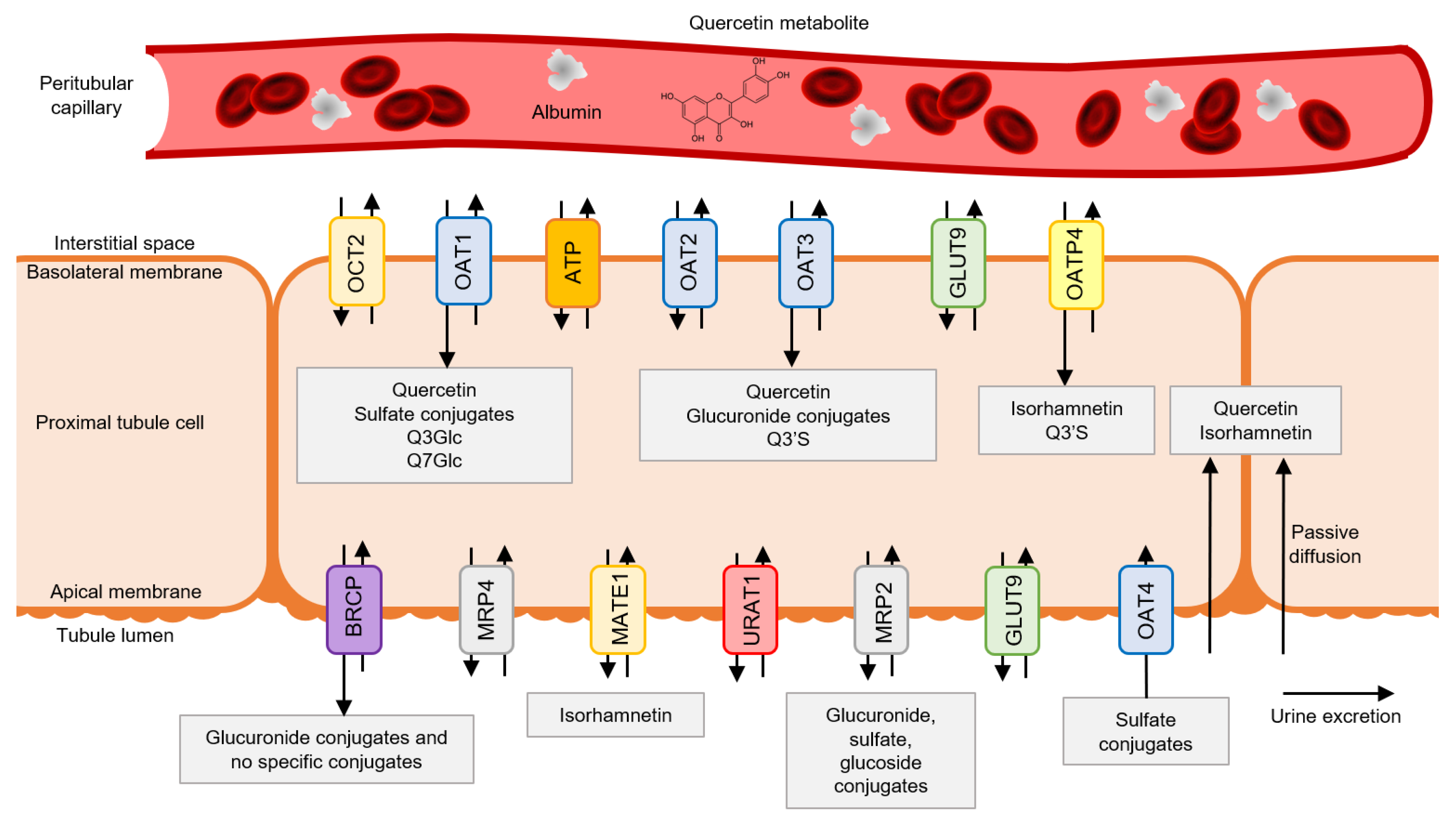
3.1. Quercetin Metabolite Uptake by Tubular Cells
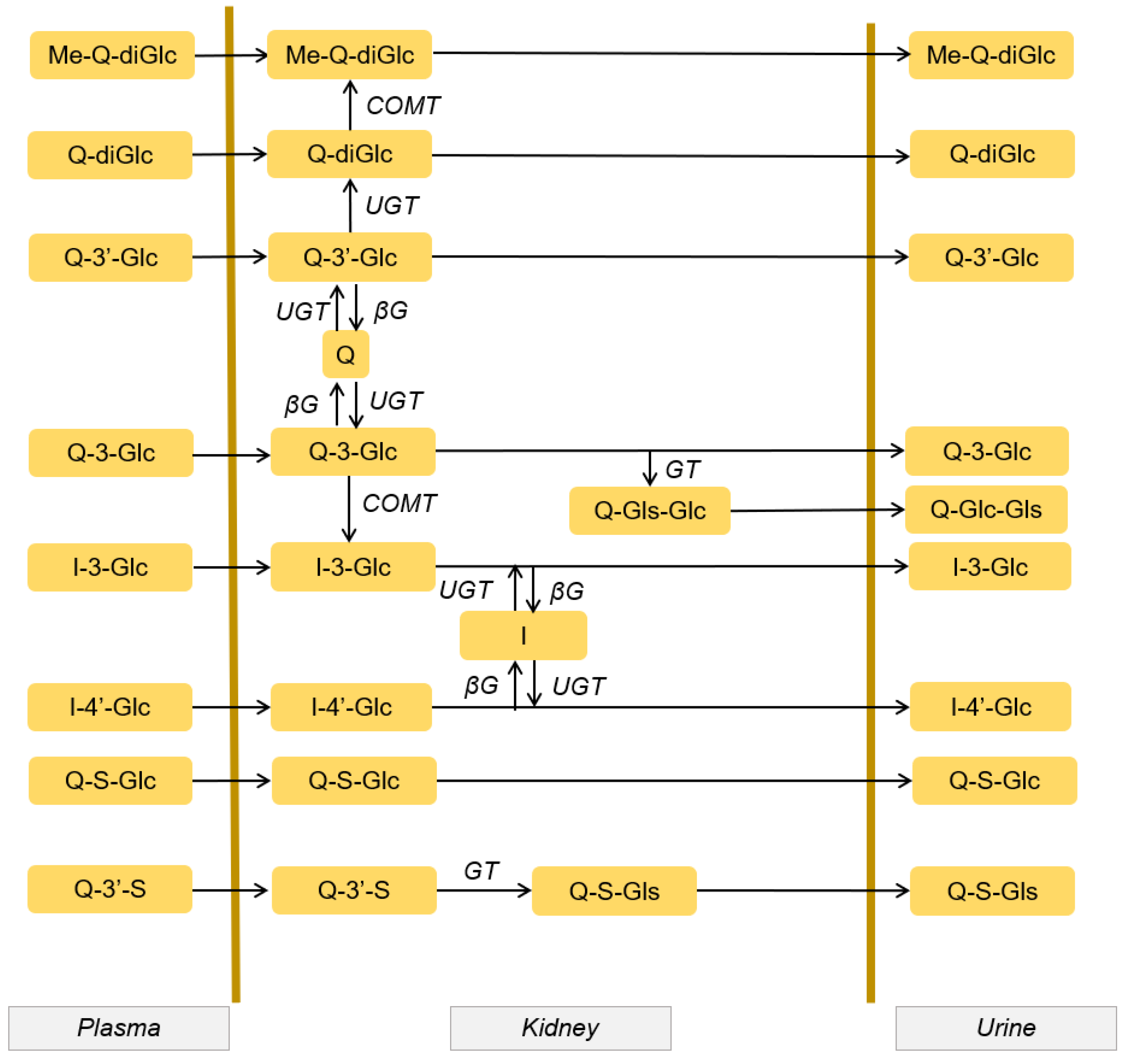
3.2. Quercetin Metabolism in Tubular Cells
3.3. Quercetin Metabolite Accumulation in Tubular Cells
3.4. Quercetin Metabolites’ Luminal Efflux by Tubular Cells
3.5. Quercetin Metabolites in Urine
4. Antioxidant and Renoprotective Effects of Quercetin and Its Metabolites
4.1. Quercetin Antioxidant Activity
4.1.1. Scavenger Activity
4.1.2. Glutathione Level Regulation
4.1.3. Antioxidant Enzyme Modulation
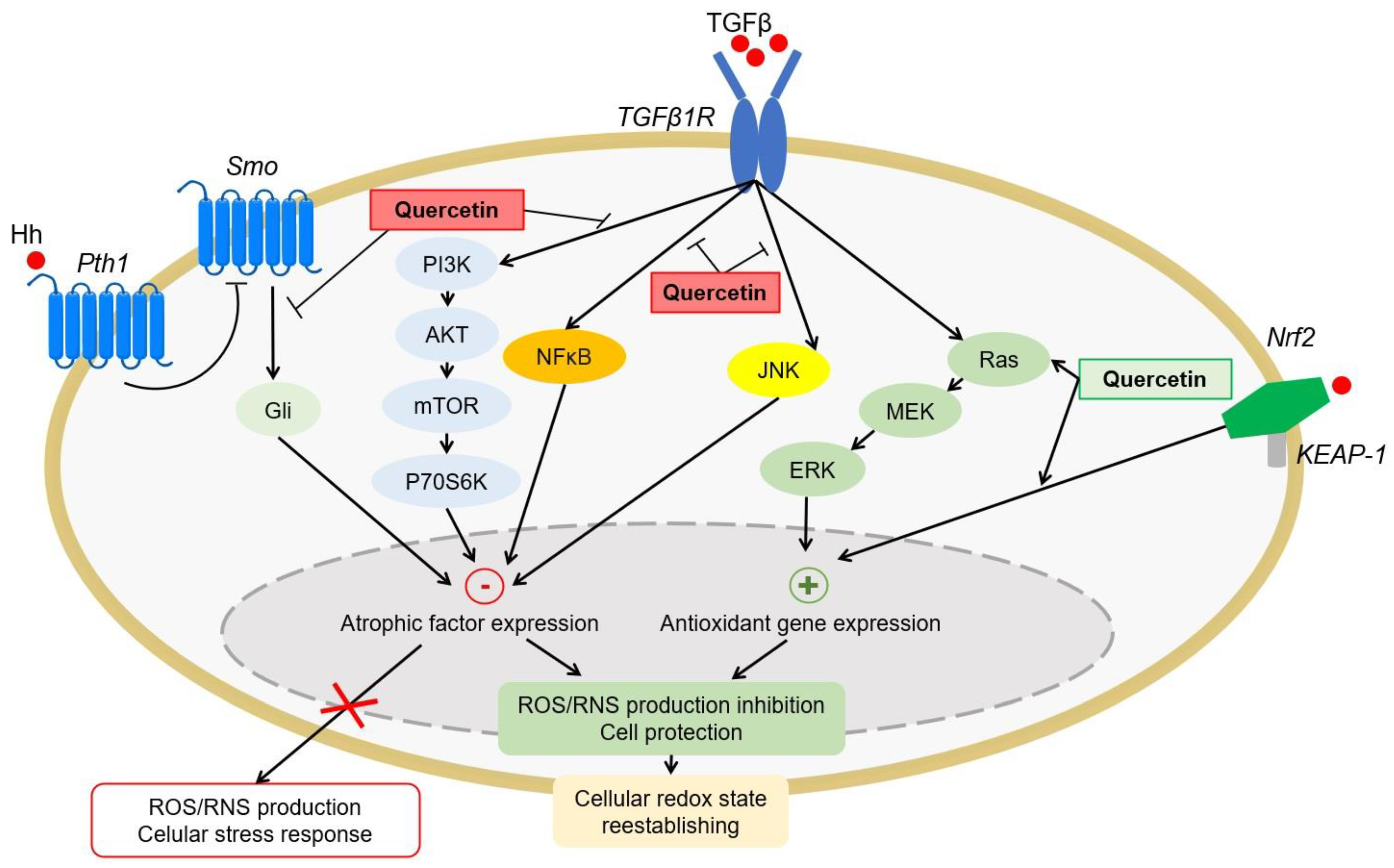
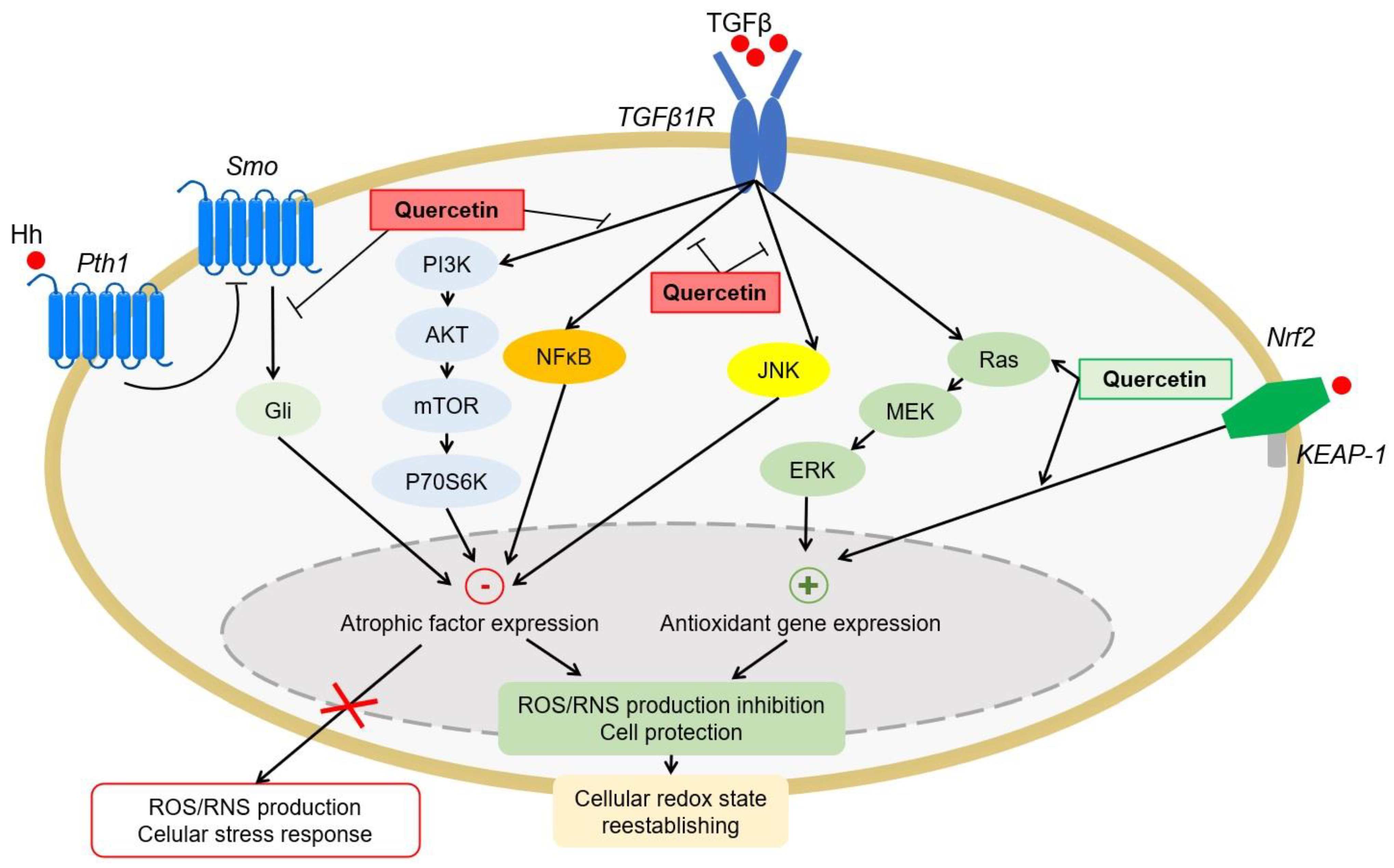
4.1.4. Antioxidant Signaling Pathways
4.2. Renoprotective Activity of Quercetin and Its Metabolites
Aspects to Be Elucidated and Limitations of Current Studies
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of quercetin in humans with a focus on interindividual variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Wang, D.; Sun-Waterhouse, D.; Li, F.; Xin, L.; Li, D. MicroRNAs as molecular targets of quercetin and its derivatives underlying their biological effects: A preclinical strategy. Crit. Rev. Food Sci. Nutr. 2019, 59, 2189–2201. [Google Scholar] [CrossRef]
- Darsandhari, S.; Bae, J.Y.; Shrestha, B.; Yamaguchi, T.; Jung, H.J.; Han, J.M.; Rha, C.-S.; Pandey, R.P.; Sohng, J.K. Enzymatic synthesis of novel quercetin sialyllactoside derivatives. Nat. Prod. Res. 2019, 33, 1944–1952. [Google Scholar] [CrossRef]
- Vrba, J.; Kren, V.; Vacek, J.; Papouskova, B.; Ulrichova, J. Quercetin, quercetin glycosides and taxifolin differ in their ability to induce AhR activation and CYP1A1 expression in HepG2 cells. Phytother. Res. 2012, 26, 1746–1752. [Google Scholar] [CrossRef]
- Wang, W.; Sun, C.; Mao, L.; Ma, P.; Liu, F.; Yang, J.; Gao, Y. The biological activities, chemical stability, metabolism and delivery systems of quercetin: A review. Trends Food Sci. Technol. 2016, 56, 21–38. [Google Scholar] [CrossRef]
- Ferenczyova, K.; Kalocayova, B.; Bartekova, M. Potential implications of quercetin and its derivatives in cardioprotection. Int. J. Mol. Sci. 2020, 21, 1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Materska, M. Quercetin and its derivatives: Chemical structure and bioactivity—A review. Pol. J. Food Nutr. Sci. 2008, 58, 407–413. [Google Scholar]
- Khan, F.; Niaz, K.; Maqbool, F.; Hassan, F.I.; Abdollahi, M.; Venkata, K.N.; Nabavi, S.; Bishayee, A. Molecular targets underlying the anticancer effects of quercetin: An update. Nutrients 2016, 8, 529. [Google Scholar] [CrossRef] [PubMed]
- Kariu, T.; Nakao, R.; Ikeda, T.; Nakashima, K.; Potempa, J.; Imamura, T. Inhibition of gingipains and Porphyromonas gingivalis growth and biofilm formation by prenyl flavonoids. J. Periodontal Res. 2017, 52, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Hisanaga, A.; Mukai, R.; Sakao, K.; Terao, J.; Hou, D.-X. Anti-inflammatory effects and molecular mechanisms of 8-prenyl quercetin. Mol. Nutr. Food Res. 2016, 60, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Jakaria, M.; Azam, S.; Jo, S.-H.; Kim, I.-S.; Dash, R.; Choi, D.-K. Potential therapeutic targets of quercetin and its derivatives: Its role in the therapy of cognitive impairment. J. Clin. Med. 2019, 8, 1789. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.-L.; Xiao, N.; Li, X.-W.; Fan, Y.; Alolga, R.N.; Sun, X.-Y.; Wang, S.-L.; Li, P.; Qi, L.-W. Pharmacokinetic comparison between quercetin and quercetin 3-O-β-glucuronide in rats by UHPLC-MS/MS. Sci. Rep. 2016, 6, 35460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Russo, M.; Spagnuolo, C. The pleiotropic flavonoid quercetin: From its metabolism to the inhibition of protein kinases in chronic lymphocytic leukemia. Food Funct. 2014, 5, 2393–2401. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H. Membrane interactions of phytochemicals as their molecular mechanism applicable to the discovery of drug leads from plants. Molecules 2015, 20, 18923–18966. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Ma, J.; Han, J.; Li, M.; Shang, J. Pharmacokinetic comparison of quercetin, isoquercitrin, and quercetin-3-O-β-D-glucuronide in rats by HPLC-MS. PeerJ 2019, 7, e6665. [Google Scholar] [CrossRef]
- Graf, B.A.; Ameho, C.; Dolnikowski, G.G.; Milbury, P.E.; Chen, C.-Y.; Blumberg, J.B. Rat gastrointestinal tissues metabolize quercetin. J. Nutr. 2006, 136, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.-L.; Lin, Y.-C.; Lin, Y.-L.; Li, C.-C.; Chuang, C.-H. Comparing the metabolism of quercetin in rats, mice and gerbils. Eur. J. Nutr. 2016, 55, 413–422. [Google Scholar] [CrossRef]
- Murota, K.; Hotta, A.; Ido, H.; Kawai, Y.; Moon, J.-H.; Sekido, K.; Hayashi, H.; Inakuma, T.; Terao, J. Antioxidant capacity of albumin-bound quercetin metabolites after onion consumption in humans. J. Med. Investig. 2007, 54, 370–374. [Google Scholar] [CrossRef] [Green Version]
- Fiorani, M.; Guidarelli, A.; Blasa, M.; Azzolini, C.; Candiracci, M.; Piatti, E.; Cantoni, O. Mitochondria accumulate large amounts of quercetin: Prevention of mitochondrial damage and release upon oxidation of the extramitochondrial fraction of the flavonoid. J. Nutr. Biochem. 2010, 21, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-González, P.D.; López-Hernández, F.J.; Dueñas, M.; Prieto, M.; Sánchez-López, E.; Thomale, J.; Ruiz-Ortega, M.; López-Novoa, J.M.; Morales, A.I. Differential effect of quercetin on cisplatin-induced toxicity in kidney and tumor tissues. Food Chem. Toxicol. 2017, 107, 226–236. [Google Scholar] [CrossRef]
- Ren, J.; Li, J.; Liu, X.; Feng, Y.; Gui, Y.; Yang, J.; He, W.; Dai, C. Quercetin inhibits fibroblast activation and kidney fibrosis involving the suppression of mammalian target of rapamycin and β-catenin signaling. Sci. Rep. 2016, 6, 23968. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.C.; Botting, N.P.; Orfila, C.; Al-Maharik, N.; Williamson, G. Flavonoid conjugates interact with organic anion transporters (OATs) and attenuate cytotoxicity of adefovir mediated by organic anion transporter 1 (OAT1/SLC22A6). Biochem. Pharmacol. 2011, 81, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Mandíková, J.; Volková, M.; Pávek, P.; Česnek, M.; Janeba, Z.; Kubíček, V.; Trejtnar, F. Interactions with selected drug renal transporters and transporter-mediated cytotoxicity in antiviral agents from the group of acyclic nucleoside phosphonates. Toxicology 2013, 311, 135–146. [Google Scholar] [CrossRef]
- Wang, M.; Qi, H.; Li, J.; Xu, Y.; Zhang, H. Transmembrane transport of steviol glucuronide and its potential interaction with selected drugs and natural compounds. Food Chem. Toxicol. 2015, 86, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Akiyama, Y.; Abe, T.; Lippiat, J.D.; Orfila, C.; Williamson, G. Carrier-mediated transport of quercetin conjugates: Involvement of organic anion transporters and organic anion transporting polypeptides. Biochem. Pharmacol. 2012, 84, 564–570. [Google Scholar] [CrossRef]
- Duan, J.; Xie, Y.; Luo, H.; Li, G.; Wu, T.; Zhang, T. Transport characteristics of isorhamnetin across intestinal Caco-2 cell monolayers and the effects of transporters on it. Food Chem. Toxicol. 2014, 66, 313–320. [Google Scholar] [CrossRef] [PubMed]
- George, B.; You, D.; Joy, M.S.; Aleksunes, L.M. Xenobiotic transporters and kidney injury. Adv. Drug Deliv. Rev. 2017, 116, 73–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hai, Y.; Zhang, Y.; Liang, Y.; Ma, X.; Qi, X.; Xiao, J.; Xue, W.; Luo, Y.; Yue, T. Advance on the absorption, metabolism, and efficacy exertion of quercetin and its important derivatives. Food Front. 2020, 1, 420–434. [Google Scholar] [CrossRef]
- Graf, B.A.; Mullen, W.; Caldwell, S.T.; Hartley, R.C.; Duthie, G.G.; Lean, M.E.J.; Crozier, A.; Edwards, C.A. Disposition and metabolism of [2-14 C]Quercetin-4′-glucoside in rats. Drug Metab. Dispos. 2005, 33, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- De Boer, V.C.; Dihal, A.A.; van der Woude, H.; Arts, I.C.; Wolffram, S.; Alink, G.M.; Rietjens, I.M.; Keijer, J.; Hollman, P.C. Tissue distribution of quercetin in rats and pigs. J. Nutr. 2005, 135, 1718–1725. [Google Scholar] [CrossRef] [Green Version]
- Mukai, R.; Fujikura, Y.; Murota, K.; Uehara, M.; Minekawa, S.; Matsui, N.; Kawamura, T.; Nemoto, H.; Terao, J. Prenylation enhances quercetin uptake and reduces efflux in caco-2 cells and enhances tissue accumulation in mice fed long-term. J. Nutr. 2013, 143, 1558–1564. [Google Scholar] [CrossRef] [Green Version]
- Bieger, J.; Cermak, R.; Blank, R.; de Boer, V.C.J.; Hollman, P.C.H.; Kamphues, J.; Wolffram, S. Tissue distribution of quercetin in pigs after long-term dietary supplementation. J. Nutr. 2008, 138, 1417–1420. [Google Scholar] [CrossRef] [PubMed]
- Riches, Z.; Stanley, E.L.; Bloomer, J.C.; Coughtrie, M.W.H. Quantitative evaluation of the expression and activity of five major sulfotransferases (SULTs) in human tissues: The SULT “Pie”. Drug Metab. Dispos. 2009, 37, 2255–2261. [Google Scholar] [CrossRef] [Green Version]
- Margaillan, G.; Rouleau, M.; Fallon, J.K.; Caron, P.; Villeneuve, L.; Turcotte, V.; Smith, P.C.; Joy, M.S.; Guillemette, C. Quantitative profiling of human renal UDP-glucuronosyltransferases and glucuronidation activity: A comparison of normal and tumoral kidney tissues. Drug Metab. Dispos. 2015, 43, 611–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strassburg, C.P.; Nguyen, N.; Manns, M.P.; Tukey, R.H. UDP-glucuronosyltransferase activity in human liver and colon. Gastroenterology 1999, 116, 149–160. [Google Scholar] [CrossRef]
- Webb, L.J.; Miles, K.K.; Auyeung, D.J.; Kessler, F.K.; Ritter, J.K. Analysis of substrate specificities and tissue expression of rat UDP-glucuronosyltransferases UGT1A7 and UGT1A8. Drug Metab. Dispos. 2005, 33, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Semenova, S.; Rozov, S.; Panula, P. Distribution, properties, and inhibitor sensitivity of zebrafish catechol-O-methyl transferases (COMT). Biochem. Pharmacol. 2017, 145, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, K.; Piskula, M.K. Food content, processing, absorption and metabolism of onion flavonoids. Crit. Rev. Food Sci. Nutr. 2007, 47, 397–409. [Google Scholar] [CrossRef]
- Guo, J.; Du, L.; Shang, E.; Li, T.; Liu, Y.; Qian, D.; Tang, Y.; Duan, J. Conjugated metabolites represent the major circulating forms of Abelmoschus manihot in vivo and show an altered pharmacokinetic profile in renal pathology. Pharm. Biol. 2016, 54, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Graf, B.A.; Caldwell, S.T.; Hartley, R.C.; Duthie, G.G.; Edwards, C.A.; Lean, M.E.J.; Crozier, A. Determination of flavonol metabolites in plasma and tissues of rats by HPLC-radiocounting and tandem mass spectrometry following oral ingestion of [2-14C]quercetin-4‘-glucoside. J. Agric. Food Chem. 2002, 50, 6902–6909. [Google Scholar] [CrossRef] [PubMed]
- Paulke, A.; Eckert, G.P.; Schubert-Zsilavecz, M.; Wurglics, M. Isoquercitrin provides better bioavailability than quercetin: Comparison of quercetin metabolites in body tissue and brain sections after six days administration of isoquercitrin and quercetin. Pharmazie 2012, 67, 991–996. [Google Scholar] [CrossRef]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P.C. Multidrug resistance proteins: Role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Mohos, V.; Fliszár-Nyúl, E.; Ungvári, O.; Kuffa, K.; Needs, P.W.; Kroon, P.A.; Telbisz, Á.; Özvegy-Laczka, C.; Poór, M. Inhibitory effects of quercetin and its main methyl, sulfate, and glucuronic acid conjugates on cytochrome P450 enzymes, and on OATP, BCRP and MRP2 transporters. Nutrients 2020, 12, 2306. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Ito, H.; Omote, H. Components of foods inhibit a drug exporter, human multidrug and toxin extrusion transporter. Biol. Pharm. Bull. 2014, 37, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Walgren, R.A.; Karnaky, K.J.; Lindenmayer, G.E.; Walle, T. Efflux of dietary flavonoid quercetin 4’-beta-glucoside across human intestinal Caco-2 cell monolayers by apical multidrug resistance-associated protein-2. J. Pharmacol. Exp. Ther. 2000, 294, 830–836. [Google Scholar]
- Biancatelli, R.M.L.C.; Berrill, M.; Catravas, J.D.; Marik, P.E. Quercetin and vitamin C: An experimental, synergistic therapy for the prevention and treatment of SARS-CoV-2 related disease (COVID-19). Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Coppolino, G.; Leonardi, G.; Andreucci, M.; Bolignano, D. Oxidative stress and kidney function: A brief update. Curr. Pharm. Des. 2019, 24, 4794–4799. [Google Scholar] [CrossRef]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant mechanisms in renal injury and disease. Antioxid. Redox Signal. 2016, 25, 119–146. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Batiha, G.E.-S.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The pharmacological activity, biochemical properties, and pharmacokinetics of the major natural polyphenolic flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef] [Green Version]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Terao, J.; Murota, K.; Kawai, Y. Conjugated quercetin glucuronides as bioactive metabolites and precursors of aglycone in vivo. Food Funct. 2011, 2, 11–17. [Google Scholar] [CrossRef]
- Johinke, D.; de Graaf, S.P.; Bathgate, R. Quercetin reduces the in vitro production of H2O2 during chilled storage of rabbit spermatozoa. Anim. Reprod. Sci. 2014, 151, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Seo, K.; Ki, S.H.; Shin, S.M. Isorhamnetin inhibits reactive oxygen species-dependent hypoxia inducible factor (HIF)-1α accumulation. Biol. Pharm. Bull. 2016, 39, 1830–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchionatti, A.M.; Pacciaroni, A.; de Talamoni, N.G.T. Effects of quercetin and menadione on intestinal calcium absorption and the underlying mechanisms. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 164, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Kinaci, M.K.; Erkasap, N.; Kucuk, A.; Koken, T.; Tosun, M. Effects of quercetin on apoptosis, NF-κB and NOS gene expression in renal ischemia/reperfusion injury. Exp. Ther. Med. 2012, 3, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Altintas, R.; Ciftci, O.; Aydin, M.; Akpolat, N.; Oguz, F.; Beytur, A. Quercetin prevents docetaxel-induced testicular damage in rats. Andrologia 2015, 47, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Piao, M.; Song, Y.; Liu, C. Quercetin suppresses AOM/DSS-induced colon carcinogenesis through its anti-inflammation effects in mice. J. Immunol. Res. 2020, 2020, 9242601. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, B.L.; Xu, C.G.; Zhou, X.J. Dihydroquercetin protects against renal fibrosis by activating the Nrf2 pathway. Phytomedicine 2020, 69, 153185. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, P.; Zhang, Q.; Fu, Y.; Hou, Y.; Wei, Y.; Zheng, X.; Feng, W. Taxifolin improves disorders of glucose metabolism and water-salt metabolism in kidney via PI3K/AKT signaling pathway in metabolic syndrome rats. Life Sci. 2020, 263, 118713. [Google Scholar] [CrossRef]
- Xu, M.X.; Wang, M.; Yang, W.W. Gold-quercetin nanoparticles prevent metabolic endotoxemia-induced kidney injury by regulating TLR4/NF-κB signaling and Nrf2 pathway in high fat diet fed mice. Int. J. Nanomed. 2017, 12, 327–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Sun, N.; Mo, N.; Lu, S.; Song, E.; Ren, C.; Li, Z. Quercetin inhibits kidney fibrosis and the epithelial to mesenchymal transition of the renal tubular system involving suppression of the Sonic Hedgehog signaling pathway. Food Funct. 2019, 10, 3782–3797. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, J.; Liu, C.; Wang, X.; Pan, J. Hyperoside alleviated N-acetyl-para-amino-phenol-induced acute hepatic injury via Nrf2 activation. Int. J. Clin. Exp. Pathol. 2019, 12, 64–76. [Google Scholar]
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer’s disease. Life Sci. 2019, 224, 109–119. [Google Scholar] [CrossRef]
- Morales, A.I.; Vicente-Sánchez, C.; Jerkic, M.; Santiago, J.M.; Sánchez-González, P.D.; Pérez-Barriocanal, F.; López-Novoa, J.M. Effect of quercetin on metallothionein, nitric oxide synthases and cyclooxygenase-2 expression on experimental chronic cadmium nephrotoxicity in rats. Toxicol. Appl. Pharmacol. 2006, 210, 128–135. [Google Scholar] [CrossRef]
- Sanchez-Gonzalez, P.D.; Lopez-Hernandez, F.J.; Perez-Barriocanal, F.; Morales, A.I.; Lopez-Novoa, J.M. Quercetin reduces cisplatin nephrotoxicity in rats without compromising its anti-tumour activity. Nephrol. Dial. Transplant. 2011, 26, 3484–3495. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Vicente, L.; González-Calle, D.; Casanova, A.G.; Hernández-Sánchez, M.T.; Prieto, M.; Rama-Merchán, J.C.; Martín-Moreiras, J.; Martín-Herrero, F.; Sánchez, P.L.; López-Hernández, F.J.; et al. Quercetin, a promising clinical candidate for the prevention of contrast-induced nephropathy. Int. J. Mol. Sci. 2019, 20, 4961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreucci, M.; Faga, T.; Pisani, A.; Serra, R.; Russo, D.; de Sarro, G.; Michael, A. Quercetin protects against radiocontrast medium toxicity in human renal proximal tubular cells. J. Cell. Physiol. 2018, 233, 4116–4125. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, S.; Pergola, P.E.; Zager, R.A.; Vaziri, N.D. Targeting the transcription factor Nrf2 to ameliorate oxidative stress and inflammation in chronic kidney disease. Kidney Int. 2013, 83, 1029–1041. [Google Scholar] [CrossRef] [Green Version]
- Casanova, A.G.; Prieto, M.; Colino, C.I.; Gutiérrez-Millán, C.; Ruszkowska-Ciastek, B.; de Paz, E.; Martín, Á.; Morales, A.I.; López-Hernández, F.J. A micellar formulation of quercetin prevents cisplatin nephrotoxicity. Int. J. Mol. Sci. 2021, 22, 729. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
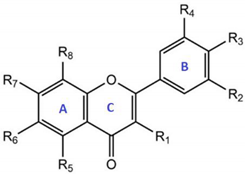
| Quercetin Derivatives | Radicals | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | ||
| Aglycone | Quercetin | OH | H | OH | OH | OH | H | OH | H |
| Reduced aglycone | Dihydroquercetin (taxifolin) | (OH)1 | H | OH | OH | OH | H | OH | H |
| Glycosides | Quercetin 3-O-rhamnoside (quercitrin) | O-Rham2 | H | OH | OH | OH | H | OH | H |
| Quercetin 7-O-rhamnoside (Vincetoxicoside B) | OH | H | OH | OH | OH | H | O-Rham | H | |
| Quercetin 3-O-rhamnoglucoside (rutin) | O-RG3 | H | OH | OH | OH | H | OH | H | |
| Quercetin 3-O-glucoside (isoquercetin) | O-Gls4 | H | OH | OH | OH | H | OH | H | |
| Quercetin 3-O-galactoside (hyperoside) | O-Gal5 | H | OH | OH | OH | H | OH | H | |
| Ethers | Quercetin 3′-methylether (isorhamnetin) | OH | H | OH | O-Met6 | OH | H | OH | H |
| Quercetin 4′-methylether (tamarixetin) | OH | H | O-Met | OH | OH | H | OH | H | |
| Quercetin 7-methylether (rhamnetin) | OH | H | OH | OH | OH | H | O-Met | H | |
| Prenylated | 8-prenylquercetin | OH | H | OH | OH | OH | H | OH | Pren7 |
| 6,5′-di-C-prenylquercetin | OH | H | OH | Pren | OH | Pren | OH | H | |
| Sulfated | Quercetin 3,7,3′,4′-tetrasulfate | OSO3-8 | H | OSO3- | OSO3- | OH | H | OSO3- | H |
| Quercetin 3-O-sulfate | OSO3- | H | OH | OH | OH | H | OH | H | |
| Quercetin 3′-O-sulfate | OH | H | OH | OSO3- | OH | H | OH | H | |
| Quercetin 4′-O-sulfate | OH | H | OSO3- | OH | OH | H | OH | H | |
| Quercetin 7-O-sulfate | OH | H | OH | OH | OH | H | OSO3- | H | |
| Glucuronides | Quercetin 3-O-glucuronide | O-Glc9 | H | OH | OH | OH | H | OH | H |
| Quercetin 3′-O-glucuronide | OH | H | OH | O-Glc | OH | H | OH | H | |
| Quercetin 4′-O-glucuronide | OH | H | O-Glc | OH | OH | H | OH | H | |
| Quercetin 7-O-glucuronide | OH | H | OH | OH | OH | H | O-Glc | H | |
| Quercetin diglucuronide | O-Glc | H | OH | OH | OH | H | O-Glc | H | |
| Quercetin Conjugates (%) | Data from [20] | |
|---|---|---|
| Plasma | Urine | |
| Quercetin monoglucuronide | 27.6 | 21.4 |
| Quercetin diglucuronide | 4.6 | 17.2 |
| Methylated quercetin monoglucuronide | 8.8 | 19.3 |
| Methylated quercetin diglucuronide | - | 11.1 |
| Quercetin glucoside monoglucuronide | - | 1.3 |
| Quercetin glucoside sulfate | - | 9.4 |
| Quercetin monoglucuronide sulfate | 10.5 | 20.3 |
| Quercetin 3′-sulfate | 48.5 | Trace |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Reyes, D.; Morales, A.I.; Prieto, M. Transit and Metabolic Pathways of Quercetin in Tubular Cells: Involvement of Its Antioxidant Properties in the Kidney. Antioxidants 2021, 10, 909. https://doi.org/10.3390/antiox10060909
Muñoz-Reyes D, Morales AI, Prieto M. Transit and Metabolic Pathways of Quercetin in Tubular Cells: Involvement of Its Antioxidant Properties in the Kidney. Antioxidants. 2021; 10(6):909. https://doi.org/10.3390/antiox10060909
Chicago/Turabian StyleMuñoz-Reyes, Daniel, Ana I. Morales, and Marta Prieto. 2021. "Transit and Metabolic Pathways of Quercetin in Tubular Cells: Involvement of Its Antioxidant Properties in the Kidney" Antioxidants 10, no. 6: 909. https://doi.org/10.3390/antiox10060909
APA StyleMuñoz-Reyes, D., Morales, A. I., & Prieto, M. (2021). Transit and Metabolic Pathways of Quercetin in Tubular Cells: Involvement of Its Antioxidant Properties in the Kidney. Antioxidants, 10(6), 909. https://doi.org/10.3390/antiox10060909