
Application of Polypodiopsida Class in Nanotechnology–Potential towards Development of More Effective Bioactive Solutions

 ,
,  ,
, 

Abstract
1. Introduction
2. The Polypodiopsida Class
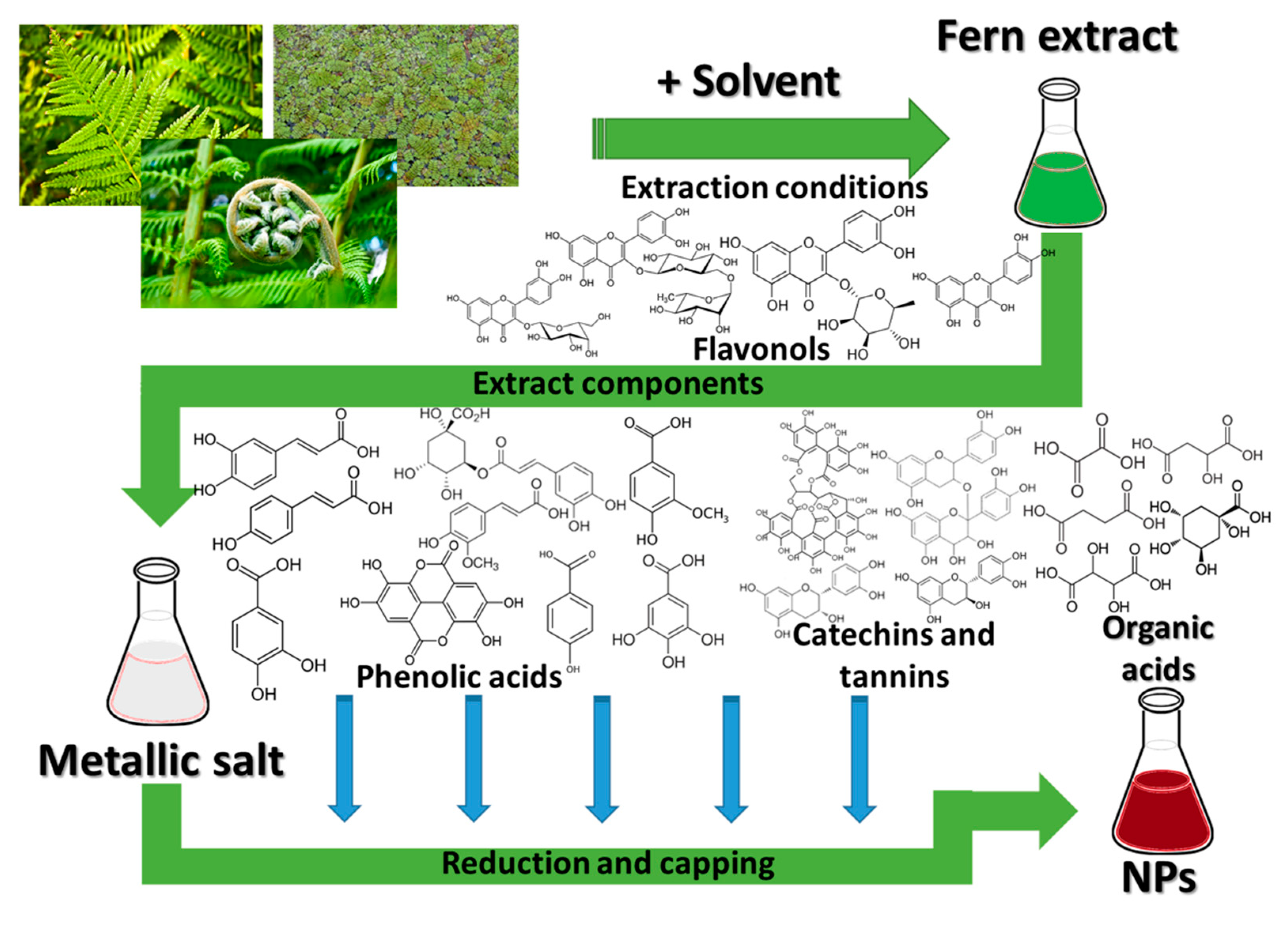
3. Polypodiopsida and the Nanomaterials
- -
- factors related to the vegetal extract used: the intrinsic properties of the plants, related to their phytocomponents, the part of the plant used, extraction procedure, used solvents, the vegetal material to solvent ratio, plant pre-treatment, etc.;
- -
- factors related to the phytosynthesis process: concentration of the metallic salt precursors, reaction conditions (temperature, pH, reaction time), extract to metallic salt ratio, etc.
3.1. Nanoparticle Phytosynthesis Using Ferns
- -
- silver ions can bind to form different insoluble precipitates, which can negatively affect their properties [65];
- -
- particularly for the case of phytosynthesized nanoparticles, the use of different phytocomponents as capping agents can not only contribute to an increase in their antimicrobial or antioxidant potential (for example) [18], but can decrease their toxic potential against non-target organisms [18], which is actually lower for NPs, compared with silver ions [67];
- -
- the large surface area to volume ratio of nanoparticles (an element common for all types of NPs) provides better contact with microorganisms, thus increasing their antimicrobial potential, as well as contributing to their successful application in other areas [68].

{kind=link}
{kind=link}
| Fern | Family | Plant Part | Extraction Conditions | NP Characteristics | Intended Application | Ref. |
|---|---|---|---|---|---|---|
| Terrestrial ferns | ||||||
| Asplenium scolopendrium L. | Aspleniaceae | Leaves, rhizomes | Ethanol, V.M.S.R. = 1:10, 48 h, R.T. | AgNPs, <50 nm | Antioxidant, cytotoxic | [47] |
| Asplenium scolopendrium L. | Aspleniaceae | Leaves | Water:ethanol, V.M.S.R. = 1:10, temperature extraction (2 h, 80 °C), microwave extraction (80 °C for 20 min., with magnetic stirring) | AgNPs, spherical, 12 nm (classical extraction), 10 nm (microwave extraction) | Antimicrobial, cytotoxic, phytotoxic | [17] |
| Diplazium esculentum (Retz.) Sw. | Athyriaceae | Leaves, powder | Dried powder directly applied in the metallic salt solution | AgNPs, spherical, oval, triangular, 10–45 nm | Photocatalytic, anticoagulative | [71] |
| Diplazium esculentum (Retz.) Sw. | Athyriaceae | Leaves, powder | Water, V.M.S.R. = 1:4.5, 70 °C, 20 min. | AgNPs, spherical, 10–25 nm (increased with extract concentration) AuNPs—spherical, triangular and decahedral shapes, 35–75 nm (dependent on reaction temperature and extract concentration) | Antimicrobial | [75] |
| Pteridium aquilinum (L.) Kuhn | Dennstaedtiaceae | Leaves | Water, V.M.S.R. = 1:10, boiling, 5 min. | AgNPs, spherical, 35–65 nm | Mosquitocidal, antiplasmodial | [90] |
| Dryopteris crassirhizoma Nakai (1920) | Dryopteridaceae | Rhizomes | Water, V.M.S.R. = 1:2.5, boiling, 30 min. | AgNPs, spherical, 5–60 nm (pH, extract, metallic salt and light exposure dependant) | Antibacterial | [91] |
| Equisetum arvense L. | Equisetaceae | Leaves | Water, V.M.S.R. = 1:25, microwave extraction (100 W for 3 min, 180 W for 1 min) | AgNPs, nearly spherical, 10–60 nm (phytosynthesis temperature dependant) | Antimicrobial | [72] |
| Equisetum arvense L. | Equisetaceae | Leaves | Water, V.M.S.R. = 1:4.58, boiled for 20 min under stirring. | AgNPs, spherical, hydrodynamic diameter, 170.5 nm | Cytotoxicity, antidiabetic, antioxidant, antibacterial | [73] |
| Equisetum giganteum L. | Equisetaceae | Leaves | Water, liquid-solid extraction, V.M.S.R. = 1:10, 80 °C, 3 min | AgNPs, spherical, 20 nm | Coating biodeterioration control | [92] |
| Dicranopteris linearis (Burm.f.) Underw | Gleicheniaceae | Leaves | Water, V.M.S.R. = 1:10, boiled for 5 min. | AgNPs, spherical, 40–60 nm | Mosquito oviposition deterrents | [93] |
| Gleichenella pectinata (Willd.) Ching | Gleicheniaceae | Leaves | Powdered sample soaked in methanol for 72 h at RT, with frequent manual agitation. V.M.S.R. = 1:10 | AgNPs, spherical, 7.51 ± 2.88 nm. | Antimicrobial | [94] |
| Nephrolepis biserrata (Sw.) Schott | Nephrolepidaceae | Leaves | Methanol, V.M.S.R. = 1:8, 50 °C, 10 min. | RuNPs, hexagonal, amorphous, ~26 nm | Antifungal, antioxidant | [78] |
| Nephrolepis cordifolia (L.) K. Presl | Nephrolepidaceae | Tubers | Water, V.M.S.R. = 1:10, boiled for 15 min under stirring | Au–Ag@AgCl nanocomposites, spherical, 10–50 nm (average 30 nm) | Nanocatalysts | [81] |
| Nephrolepis cordifolia (L.) K. Presl | Nephrolepidaceae | Tubers | Water, V.M.S.R. = 1:20, boiled for 10 min | SiO2@Au–Ag nanocomposites, Spherical SiO2 decorated with AuNPs and AgNPs, 200–246 nm (SiO2), 3 nm (AgNPs/AuNPs) | Nanocatalysts, antibacterial, cytotoxic | [82] |
| Nephrolepis cordifolia (L.) K. Presl | Nephrolepidaceae | Not declared | Water, V.M.S.R. = 3:100, 80 °C for 80 min. | FeNPs, amorphous, spherical 40–70 nm, other iron oxides | Cr (VI) removal | [79] |
| Adiantum philippense L. | Pteridaceae | Aerial parts | Dried plant material, water, V.M.S.R. = 1:20 | Anisotropic, AgNP, 13 nm, AuNP, 11 nm | Proposed for biomedical applications | [75] |
| Adiantum philippense L. | Pteridaceae | Aerial parts | Dried plant material, water, V.M.S.R. = 1:50, 25 °C for 72 h., under continuous shaking. | AuNPs, spherical and triangular, average particle size 33.9 ± 14.0 nm. | Antibacterial | [76] |
| Adiantum philippense L. | Pteridaceae | Whole plant | Dried plant material, water, V.M.S.R. = 1:20 | AgNPs, quasi-spherical, 10–60 nm | Antimicrobial | [77] |
| Adiantum philippense L. | Pteridaceae | Whole plant | Dried plant material, water, V.M.S.R. = 1:20 | CuONPs, quasi-spherical, 1–20 nm | Plant defence booster | [80] |
| Pteris tripartita Sw. | Pteridaceae | Leaves | Water, V.M.S.R. = 1:20, boiling for 5 min. | AgNPs, hexagonal, spherical, and rod-shaped, 32 nm | Anticancer, toxicity studies (on Zebra fish) | [95] |
| Adiantum raddianum C. Presl | Pteridaceae | Leaves | Dried leaf powder, water, V.M.S.R. = 1:10, 3 h., magnetic stirring | AgNPs, mostly spherical, with cubic morphologies, 9.69–13.9 nm | Mosquitocidal | [96] |
| Adiantum capillus-veneris L. | Pteridaceae | Leaves | Dried leaf powder, water, 70 °C, 15 min. | AgNPs, spherical, 18.4 nm | Antibacterial | [97] |
| Adiantum capillus-veneris L. | Pteridaceae | Leaves | Dried leaf powder, water, M.S.R. = 1:10, 60 °C, 10 min. | AuNPs | Antioxidant, antibacterial, antifungal | [98] |
| Adiantum sp. | Pteridaceae | Leaves | Aqueous extract | AgNPs, AuNPs | Anticancer | [99] |
| Pteris quadriaurita Retz. | Pteridaceae | Leaves | Dried leaf powder, water, M.S.R. = 1:10, 60 °C, 10 min. | AuNPs | Antioxidant, antibacterial, antifungal | [98] |
| Aquatic ferns | ||||||
| Marsilea quadrifolia L. | Marsileaceae | Leaves | Water, V.M.S.R. = 1:20, 90 °C, 1 h., under stirring (400 rpm) | AgNPs, spherical, 9–42 nm | Antibacterial, anticancer | [83] |
| Marsilea quadrifolia L. | Marsileaceae | Leaves | Dried leaf powder, methanol, V.M.S.R. = 1:10, R.T., 72 h. | AuNPs, spherical, 17–40 nm | Antidiabetic | [84] |
| Marsilea quadrifolia L. | Marsileaceae | Leaves | Dried leaf powder, water, V.M.S.R. = 1:25, 55 °C, 15 min. | AuNPs, spherical, 10–40 nm | Antioxidant, cytotoxic | [85] |
| Leptochilus pteropus (Blume) Fraser-Jenk | Polypodiaceae | Leaves | Methanol, V.M.S.R. = ~1:6, 24 h., under magnetic stirring | AgNPs | Antioxidant | [100] |
| Azolla pinnata R.Br. | Salviniaceae | Whole plant | Dried powder, hydroalcoholic solution (70%/96%), percolation (48 h.), V.M.S.R. = 1:2.5 | AgNPs, spherical, average size 6.5 nm | No application proposed | [101] |
| Azolla pinnata R.Br. | Salviniaceae | In vivo formation | Formation of NPs in root cells | PbNPs, spherical, rod-shaped, 12–80 nm dependent on the formation site | No application proposed | [87] |
| Azolla filiculoides Lam. | Salviniaceae | Whole plant | Dried powder, methanol, 72 h., V.M.S.R. = 1:10 | AuNPs, spherical, 17–40 nm | Antioxidant, hepatoprotective | [102] |
| Salvinia molesta D. Mitch. | Salviniaceae | Whole plant | Dried plant, water, boiling, 5 min., V.M.S.R. = 1:100 | AuNPs, various morphologies: spherical, triangular, pentagonal and nanoflower-like, dimensions ranging from 7 to 175.8 nm, dependent on the part plant used and extract to metallic salt ratio | No application proposed | [86] |
| Salvinia molesta D. Mitch. | Salviniaceae | Leaves | Water, boiling, 5 min., V.M.S.R. = 1:5 | AgNPs, spherical, average size 12.46 nm | Antimicrobial | [103] |
| Tree ferns | ||||||
| Cibotium barometz (L.) J. Sm. | Cibotiaceae | Roots | Dried powder, water, boiling, 30 min., V.M.S.R. = 1:20 | AgNPs, spherical, 5–40 nm; AuNPs, spherical, 5–20 nm, | Antimicrobial, antioxidant, cytotoxic | [88] |
| Alsophila nilgirensis (Holttum) R.M. Tryon | Cyatheaceae | Sporophytes | Water, boiling, 30 min., V.M.S.R. = 1:10 | AgNPs, spherical, 45–74 nm | Cytotoxic, phytotoxic | [89] |
3.2. Potential Applications of Phytosynthesized Nanoparticles
3.3. Development of Biogenic Nanoparticles
4. Other Applications of Ferns in Nano and Biotechnology
4.1. Phytoremediation and Metal Up-Take
4.2. Other Depollution Applications
4.3. Phytotoxicity Assays
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Song, X.; Liu, Z.; Sun, D.D. Nano gives the answer: Breaking the bottleneck of internal concentration polarization with a nanofiber composite forward osmosis membrane for a high water production rate. Adv. Mater. 2011, 23, 3256–3260. [Google Scholar] [CrossRef]
- Park, J.S.; Kyhm, J.; Kim, H.H.; Jeong, S.; Kang, J.H.; Lee, S.; Lee, K.T.; Park, K.; Barange, N.; Han, J.Y.; et al. Alternative patterning process for realization of large-area, full-color, active quantum dot display. Nano Lett. 2016, 16, 6946–6953. [Google Scholar] [CrossRef]
- Fuertes, G.; Soto, I.; Carrasco, R.; Vargas, M.; Sabattin, J.; Lagos, C. Intelligent packaging systems: Sensors and nanosensors to monitor food quality and safety. J. Sens. 2016, 2016, 4046061. [Google Scholar] [CrossRef]
- Contado, C. Nanomaterials in consumer products: A challenging analytical problem. Front. Chem. 2015, 3, 48. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kesarla, R.; Omri, A. Formulation strategies to improve the bioavailability of poorly absorbed drugs with special emphasis on self-emulsifying systems. ISRN Pharm. 2013, 2013, 848043. [Google Scholar] [CrossRef]
- Kumar, R.; Lal, S. Synthesis of Organic Nanoparticles and their Applications in Drug Delivery and Food Nanotechnology: A Review. J. Nanomater. Mol. Nanotechnol. 2014, 3, 4. [Google Scholar]
- FDA (U.S. Food Drug Administration). Guidance for Industry Safety of Nanomaterials in Cosmetic Products; U.S. Department of Health and Human Services Food and Drug Administration Center for Food Safety and Applied Nutrition: Rockville, MD, USA, 2014.
- Gupta, R.; Xie, H. Nanoparticles in daily life: Applications, toxicity and regulations. J. Environ. Pathol. Toxicol. Oncol. 2018, 37, 209–230. [Google Scholar] [CrossRef]
- Elwerfalli, A.M.; Al-Kinani, A.; Alany, R.G.; ElShaer, A. Nano-engineering chitosan particles to sustain the release of promethazine from orodispersables. Carbohydr. Polym. 2015, 131, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.; Ghormade, V.; Kolge, H.; Paknikar, K.M. Dual effect of chitosan-based nanoparticles on the inhibition of β-amyloid peptide aggregation and disintegration of the preformed fibrils. J. Mater. Chem. B 2019, 7, 3362–3373. [Google Scholar] [CrossRef]
- Li, X.Y.; Li, Y.C.; Yu, D.G.; Liao, Y.Z.; Wang, X. Fast disintegrating quercetin-loaded drug delivery systems fabricated using coaxial electrospinning. Int. J. Mol. Sci. 2013, 14, 21647–21659. [Google Scholar] [CrossRef] [PubMed]
- Ando, J.; Yano, T.; Fujita, K.; Kawata, S. Metal nanoparticles for nano-imaging and nano-analysis. Phys. Chem. Chem. Phys. 2013, 15, 13713–13722. [Google Scholar] [CrossRef]
- Das, S.; Kotcherlakota, R.; Patra, C.R. Noninvasive imaging techniques of metal nanoparticles and their future diagnostic applications. In Medical Imaging Methods; Shukla, A., Ed.; Springer: Singapore, 2019; pp. 119–141. [Google Scholar]
- Yaqoob, A.A.; Ahmad, H.; Parveen, T.; Ahmad, A.; Oves, M.; Ismail, I.M.I.; Qari, H.A.; Umar, K.; Mohamad, I.M.N. Recent advances in metal decorated nanomaterials and their various biological applications: A review. Front. Chem. 2020, 8, 341. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.D.; Gwenin, V.V.; Gwenin, C.D. Magnetic functionalized nanoparticles for biomedical, drug delivery and imaging applications. Nanoscale Res. Lett. 2019, 14, 188. [Google Scholar] [CrossRef]
- Mauricio, M.D.; Guerra-Ojeda, S.; Marchio, P.; Valles, S.L.; Aldasoro, M.; Escribano-Lopez, I.; Herance, J.R.; Rocha, M.; Vila, J.M.; Victor, V.M. Nanoparticles in medicine: A focus on vascular oxidative stress. Oxid. Med. Cell. Longev. 2018, 2018, 6231482. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, R.C.; Fierascu, I.; Lungulescu, E.M.; Nicula, N.; Somoghi, R.; Diţu, L.M.; Ungureanu, C.; Sutan, A.N.; Drăghiceanu, O.A.; Paunescu, A.; et al. Phytosynthesis and radiation-assisted methods for obtaining metal nanoparticles. J. Mater. Sci. 2020, 55, 1915–1932. [Google Scholar] [CrossRef]
- Fierascu, I.; Fierascu, I.C.; Brazdis, R.I.; Baroi, A.M.; Fistos, T.; Fierascu, R.C. Phytosynthesized metallic nanoparticles-between nanomedicine and toxicology. A brief review of 2019′s findings. Materials 2020, 13, 574. [Google Scholar] [CrossRef]
- Fierascu, I.; Fierascu, I.C.; Dinu-Pirvu, C.E.; Fierascu, R.C.; Anuta, V.; Velescu, B.S.; Jinga, M.; Jinga, V. A short overview of recent developments on antimicrobial coatings based on phytosynthesized metal nanoparticles. Coatings 2019, 9, 787. [Google Scholar] [CrossRef]
- Salouti, M.; Derakhshan, F.K. Phytosynthesis of nanoscale materials. In Advances in Phytonanotechnology. From Synthesis to Application; Ghorbanpour, M., Wani, S.H., Eds.; Academic Press: London, UK, 2019; pp. 45–121. [Google Scholar]
- Husen, A.; Siddiqi, K.S. Phytosynthesis of nanoparticles: Concept, controversy and application. Nanoscale Res. Lett. 2014, 9, 229. [Google Scholar] [CrossRef]
- Scott, D. The early history of the land flora. Nature 1922, 110, 638–640. [Google Scholar] [CrossRef]
- Bomfleur, B.; McLoughlin, S.; Vajda, V. Fossilized nuclei and chromosomes reveal 180 million years of genomic stasis in royal ferns. Science 2014, 343, 1376–1377. [Google Scholar] [CrossRef]
- Maroyi, A. Not just minor wild edible forest products: Consumption of pteridophytes in sub-Saharan Africa. J. Ethnobiol. Ethnomed. 2014, 10, 78. [Google Scholar] [CrossRef]
- Andrade, J.M.M.; Passos, C.S.; Dresch, R.R.; Kieling-Rubio, M.A.; Moreno, P.R.H.; Henriques, A.T. Chemical analysis, antioxidant, antichemotactic and monoamine oxidase inhibition effects of some pteridophytes from Brazil. Pharm. Mag. 2014, 10, S100–S109. [Google Scholar] [CrossRef]
- Ho, R.; Teai, T.; Bianchini, J.P.; Lafont, R.; Raharivelomanana, P. Ferns: From traditional uses to pharmaceutical development, chemical identification of active principles. In Working with Ferns: Issues and Applications; Fernández, H., Kumar, A., Revilla, A., Eds.; Springer Science: New York, NY, USA, 2010; pp. 321–346. [Google Scholar]
- Patil, M.P.; Kim, G.D. Eco-friendly approach for nanoparticles synthesis and mechanism behind antibacterial activity of silver and anticancer activity of gold nanoparticles. Appl. Microbiol. Biotechnol. 2017, 101, 79–92. [Google Scholar] [CrossRef]
- Sessa, E.B. Evolution and classification of ferns and lycophytes. In Current Advances in Fern Research; Fernández, H., Ed.; Springer: Cham, Switzerland, 2018; pp. 179–200. [Google Scholar]
- PPG, I. A community-derived classification for extant lycophytes and ferns The Pteridophyte Phylogeny Group. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- Ehrendorfer, F. Pteridophyta. In Strasburger—Lehrbuch der Botanik; Sitte, P., Ziegler, H., Ehrendorfer, F., Bresinsky, A., Eds.; Spektrum Akademischer: Berlin, Germany, 1999; pp. 652–684. [Google Scholar]
- Chen, X.; Chen, Z.; Huang, W.; Fu, H.; Wang, Q.; Wang, Y.; Cao, J. Proteomic analysis of gametophytic sex expression in the fern Ceratopteris thalictroides. PLoS ONE 2019, 14, e0221470. [Google Scholar] [CrossRef]
- Simpson, M.G. Evolution and diversity of vascular plants. In Plant Systematics; Simpson, M.G., Ed.; Academic Press: Burlington, MA, USA, 2010; pp. 73–128. [Google Scholar]
- Moran, R. Diversity, biogeography, and floristics. In Biology and Evolution of Ferns and Lycophytes; Ranker, T.A., Haufler, C.H., Eds.; Cambridge University Press: New York, NY, USA, 2008; pp. 367–394. [Google Scholar]
- Aldasoro, J.J.; Cabezas, F.; Aedo, C. Diversity and distribution of ferns in sub-Saharan Africa, Madagascar and some islands of the South Atlantic. J. Biogeogr. 2004, 31, 1579–1604. [Google Scholar] [CrossRef]
- Pérez-Atilano, Y.; Sánchez-González, A.; Tejero Díez, J.D. Species Richness, Distribution, and Morphological Variation of Lycophytes and Monilophytes in a Semi-arid Region of Mexico. Am. Fern J. 2015, 105, 238–256. [Google Scholar] [CrossRef]
- Liu, Y.; Wujisguleng, W.; Long, C. Food uses of ferns in China: A review. Acta Soc. Bot. Pol. 2012, 81, 263–270. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Chase, M.W. Trends and concepts in fern classification. Ann. Bot. 2014, 113, 571–594. [Google Scholar] [CrossRef]
- Gifford, E.M.; Foster, A.S. Morphology and Evolution of Vascular Plants, 3rd ed.; W. H. Freeman and Co.: New York, NY, USA, 1988. [Google Scholar]
- Vasco, A.; Moran, R.C.; Ambrose, B.A. The evolution, morphology, and development of fern leaves. Front. Plant Sci. 2013, 4, 345. [Google Scholar] [CrossRef]
- Davies, K.L. A brief comparative survey of aerophore structure within the Filicopsida. Bot. J. Linn. Soc. 1991, 107, 115–137. [Google Scholar] [CrossRef]
- Farrar, D.R.; Dassler, C.; Watkins, J.E., Jr.; Skelton, C. Gametophyte ecology. In Biology and Evolution of Ferns and Lycophytes; Ranker, T.A., Haufler, C.H., Eds.; Cambridge University Press: New York, NY, USA, 2008; pp. 222–256. [Google Scholar]
- Schneller, J.J. Antheridiogens. In Biology and Evolution of Ferns and Lycophytes; Ranker, T.A., Haufler, C.H., Eds.; Cambridge University Press: New York, NY, USA, 2008; pp. 134–158. [Google Scholar]
- Chiou, W.L.; Farrar, D.R. Antheridiogen production and response in Polypodiaceae species. Am. J. Bot. 1997, 84, 633–640. [Google Scholar] [CrossRef]
- Haufler, C.H.; Pryer, K.M.; Schuettpelz, E.; Sessa, E.B.; Farrar, D.R.; Moran, R.; Schneller, J.J.; Watkins, J.E., Jr.; Windham, M.D. Sex and the single gametophyte: Revising the homosporous vascular plant life cycle in light of contemporary research. BioScience 2016, 66, 928–937. [Google Scholar] [CrossRef]
- Razafin-Drabazo, F.; Donno, D.; Tombozara, N.; Razafindrakoto, Z.R.; Rajaonarison, J.F.; Andrianjara, C.; Ramanitrahasimbola, D.; Beccaro, J.L. Phyto-compounds and pharmacological activities of Lygodium lanceolatum Desv. (Schizaeaceae). S. Afr. J. Bot. 2020, 135, 225–232. [Google Scholar] [CrossRef]
- Nguyen, K.H. Assessment of Anti-Diabetic Effect of Vietnamese Herbal Drugs. Ph.D. Thesis, Department of Molecular Medicine and Surgery, Karolinska Universitetssjukhuset, Solna, Sweden, 2005. [Google Scholar]
- Sutan, N.A.; Fierascu, I.; Fierascu, R.C.; Manolescu, D.S.; Soare, L.C. Comparative analytical characterization and in vitro citogenotoxic activity evaluation of Asplenium scolopendrium L. leaves and rhizome extracts prior to and after Ag nanoparticles phytosynthesis. Ind. Crops Prod. 2016, 83C, 379–386. [Google Scholar] [CrossRef]
- Soare, L.C.; Ferdeş, M.; Stefanov, S.; Denkova, Z.; Nicolova, R.; Denev, P.; Bejan, C.; Păunescu, A. Antioxidant activity, polyphenols content and antimicrobial activity of several native pteridophytes of Romania. Not. Bot. Horti Agrobot. 2012, 40, 53–57. [Google Scholar] [CrossRef]
- Alonso-Amelot, M.E.; Oliveros, A.; Calcagno-Pisarelli, M.P. Phenolics and condensed tannins in relation to altitude in neotropical Pteridium spp. a field study in the Venezuelan Andes. Biochem. Syst. Ecol. 2004, 32, 969–981. [Google Scholar] [CrossRef]
- Alonso-Amelot, M.E.; Oliveros-Bastidas, A.; Calcagno-Pisarelli, M.P. Phenolics and condensed tannins of high altitude Pteridium arachnoideum in relation to sunlight exposure, elevation, and rain regime. Biochem. Syst. Ecol. 2007, 35, 1–10. [Google Scholar] [CrossRef]
- Vetter, V. Secondary Metabolites of Ferns. In Current Advances in Fern Research; Fernández, H., Ed.; Springer: Cham, Switzerland, 2018; pp. 305–327. [Google Scholar]
- Durdevic, L.; Mitrovic, M.; Pavlovic, P.; Bojovic, S.; Jaric, S.; Oberan, L.; Gajic, G.; Kostic, O. Total phenolics and phenolic acids content in leaves, rhizomes and rhizosphere soil under Ceterach officinarum D.C., Asplenium trichomanes L., and A. adiantum-nigrum L. in The Gorge of Sicevo (Serbia). Ekológia 2007, 26, 164–173. [Google Scholar]
- Lai, H.Y.; Lim, Y.Y. Antioxidant properties of some Malaysian ferns. In Proceedings of the 3rd International Conference on Chemical, Biological and Environmental Engineering, Singapore, 23–25 September 2011; Volume 20, pp. 8–12. [Google Scholar]
- Bancuta, O.R.; Chilian, A.; Bancuta, I.; Ion, R.M.; Setnescu, R.; Setnescu, T.; Gheboianu, A.; Lungulescu, M. FT-IR and UV-Vis characterization of grape extracts used as antioxidants in polymers. Rev. Roum. Chim. 2015, 60, 571–577. [Google Scholar]
- Jipa, S.; Zaharescu, T.; Kappel, W.; Dumitrescu, C.; Maris, M.; Mantsch, A.; Lungulescu, M. Scavenger capacity of natural phenolics in some selected labiatae herbs. Optoelectron. Adv. Mater. 2008, 2, 669–673. [Google Scholar]
- Soare, L.C.; Visoiu, E.; Bejan, C.; Dobrescu, C.M.; Fierascu, I.; Iosub, I.; Paunescu, A. Research on the in vitro bioaccumulation capacity of lead in some pteridophyte species of the Romanian flora. Rev. Chim. 2015, 66, 2017–2020. [Google Scholar]
- Lafont, R.; Ho, R.; Raharivelomanana, P.; Dinan, L. Ecdysteroids. In Working with Ferns: Issues and Applications; Fernández, H., Kumar, A., Revilla, A., Eds.; Springer Science: New York, NY, USA, 2011; pp. 305–319. [Google Scholar]
- Dinan, L.; Lafont, R. Effects and applications of arthropod steroid hormones (ecdysteroids) in mammals. J. Endocrinol. 2006, 191, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shuvalov, O.; Fedorova, O.; Tananykina, E.; Gnennaya, Y.; Daks, A.; Petukhov, A.; Barlev, N.A. An arthropod hormone, ecdysterone, inhibits the growth of breast cancer cells via different mechanisms. Front. Pharmacol. 2020, 11, 561537. [Google Scholar] [CrossRef]
- De Leeuw, J.W.; Versteegh, G.J.M.; Van Bergen, P.F. Biomacromelcules of algae and plants and their fossil analogues. Plant Ecol. 2006, 182, 209–233. [Google Scholar] [CrossRef]
- Mackenzie, G.; Beckett, S.; Atkin, S.; Taboad, A.D. Pollen and spore shells-nature’s microcapsules. In Microencapsulation in the Food Industry; Gaonkar, A.G., Vasisht, N., Khare, A.R., Sobel, R., Eds.; Academic Press: Amsterdam, The Netherlands, 2014; pp. 283–297. [Google Scholar]
- Warne, T.R.; Hickok, L.G. Control of sexual development in gametophytes of Ceratopteris richardii: Antheridiogen and abscisic acid. Bot. Gaz. 1991, 152, 148–153. [Google Scholar] [CrossRef]
- Froissard, D.; Rapior, S.; Bessière, J.M.; Buatois, B.; Fruchier, A.; Sol, V.; Fons, F. Asplenioideae species as a reservoir of volatile organic compounds with potential therapeutic properties. Nat. Prod. Commun. 2015, 10, 1079–1083. [Google Scholar] [CrossRef]
- Cerón-Carpio, A.B.; Pérez-Garcí, B.; Monribot Villanuev, J.L.; Kiel-Martínez, A.L.; Espinosa-Matias, S.; Guerrero-Analco, J.A.; Mehltreter, K. Chemical composition and micromorphological structure of cuticular leaf waxes of eight tropical fern species of Mexico. Biochem. Syst. Ecol. 2019, 85, 13–20. [Google Scholar] [CrossRef]
- Kędziora, A.; Speruda, M.; Krzyżewska, E.; Rybka, J.; Łukowiak, A.; Bugla-Płoskońska, G. Similarities and differences between silver ions and silver in nanoforms as antibacterial agents. Int. J. Mol. Sci. 2018, 19, 444. [Google Scholar] [CrossRef]
- Li, W.R.; Sun, T.L.; Zhou, S.L.; Ma, Y.K.; Shi, Q.S.; Xie, X.B.; Huang, X.M. A comparative analysis of antibacterial activity, dynamics and effects of silver ions and silver nanoparticles against four bacterial strains. Int. Biodeterior. Biodegrad. 2017, 123, 304–310. [Google Scholar]
- Velicogna, J.R.; Ritchie, E.E.; Scroggins, R.P.; Princz, J.I. A comparison of the effects of silver nanoparticles and silver nitrate on a suite of soil dwelling organisms in two field soils. Nanotoxicology 2016, 10, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Yadav, A.; Gade, A. Silver nanoparticles as a new generation of antimicrobials. Biotechnol. Adv. 2009, 27, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Perde-Schrepler, M.; Florea, A.; Brie, I.; Virag, P.; Fischer-Fodor, E.; Vâlcan, A.; Gurzău, E.; Lisencu, C.; Maniu, A. Size-dependent cytotoxicity and genotoxicity of silver nanoparticles in cochlear cells. In Vitro J. Nanomat. 2019, 2019, 6090259. [Google Scholar] [CrossRef]
- Sinha, T.; Ahmaruzzaman, M. Indigenous north eastern India fern mediated fabrication of spherical silver and anisotropic gold nano structured materials and their efficacy for the abatement of perilous organic compounds from waste water-A green approach. RSC Adv. 2016, 6, 21076–21089. [Google Scholar] [CrossRef]
- Paul, B.; Bhuyan, B.; Purkayastha, D.D.; Dhar, S.S. Green synthesis of silver nanoparticles using dried biomass of Diplazium esculentum (retz.) sw. and studies of their photocatalytic and anticoagulative activities. J. Mol. Liq. 2015, 212, 813–817. [Google Scholar] [CrossRef]
- Miljković, M.; Lazić, V.; Davidović, S.; Milivojević, A.; Papan, J.; Fernandes, M.M.; Lanceros-Mendez, S.; Ahrenkiel, S.P.; Nedeljković, J.M. Selective antimicrobial performance of biosynthesized silver nanoparticles by horsetail extract against E. coli. J. Inorg. Organomet. Polym. Mater. 2020, 30, 2598–2607. [Google Scholar] [CrossRef]
- Das, G.; Patra, J.K.; Shin, H.S. Biosynthesis, and potential effect of fern mediated biocompatible silver nanoparticles by cytotoxicity, antidiabetic, antioxidant and antibacterial, studies. Mater. Sci. Eng. C 2020, 114, 111011. [Google Scholar] [CrossRef]
- Mukherji, S.; Bharti, S.; Shukla, G.; Mukherji, S. Synthesis and characterization of size- and shape-controlled silver nanoparticles. Phys. Sci. Rev. 2018, 4, 20170082. [Google Scholar]
- Sant, D.G.; Gujarathi, T.R.; Harne, S.R.; Ghosh, S.; Kitture, R.; Kale, S.; Chopade, B.A.; Pardesi, K.R. Adiantum philippense L. frond assisted rapid green synthesis of gold and silver nanoparticles. J. Nanoparticles 2013, 2013, 182320. [Google Scholar] [CrossRef]
- Kalita, S.; Kandimalla, R.; Sharma, K.K.; Kataki, A.C.; Deka, M.; Kotoky, J. Amoxicillin functionalized gold nanoparticles reverts MRSA resistance. Mater. Sci. Eng. C 2016, 61, 720–727. [Google Scholar] [CrossRef]
- Chatterjee, A.; Khatua, S.; Acharya, K.; Sarkar, J. A green approach for the synthesis of antimicrobial bio-surfactant silver nanoparticles by using a fern. Dig. J. Nanomater. Biostruct. 2019, 14, 479–490. [Google Scholar]
- Gupta, P.K.; Ranganath, K.V.S.; Dubey, N.K.; Mishra, L. Green synthesis, characterization and biological activity of synthesized ruthenium nanoparticles using fishtail fern, sago palm, rosy periwinkle and holy basil. Curr. Sci. 2019, 117, 1308–1317. [Google Scholar] [CrossRef]
- Yi, Y.; Tu, G.; Tsang, P.E.; Xiao, S.; Fang, Z. Green synthesis of iron-based nanoparticles from extracts of Nephrolepis auriculata and applications for Cr(VI) removal. Mater. Lett. 2019, 234, 388–391. [Google Scholar] [CrossRef]
- Sarkar, J.; Chakraborty, N.; Chatterjee, A.; Bhattacharjee, A.; Dasgupta, D.; Acharya, K. Green synthesized copper oxide nanoparticles ameliorate defence and antioxidant enzymes in Lens culinaris. Nanomaterials 2020, 10, 312. [Google Scholar] [CrossRef]
- Sapkota, K.; Han, S.S. A novel environmentally sustainable synthesis of Au-Ag@AgCl nanocomposites and their application as an efficient and recyclable catalyst for quinoline synthesis. New J. Chem. 2017, 41, 5395–5402. [Google Scholar] [CrossRef]
- Sapkota, K.; Chaudhary, P.; Han, S.S. Environmentally sustainable route to SiO2@Au-Ag nanocomposites for biomedical and catalytic applications. RSC Adv. 2018, 8, 31311–31321. [Google Scholar] [CrossRef]
- Maji, A.; Beg, M.; Mandal, A.K.; Das, S.; Jha, P.K.; Kumar, A.; Sarwar, S.; Hossain, M.; Chakrabarti, P. Spectroscopic interaction study of human serum albumin and human hemoglobin with Mersilea quadrifolia leaves extract mediated silver nanoparticles having antibacterial and anticancer activity. J. Mol. Struct. 2017, 1141, 584–592. [Google Scholar] [CrossRef]
- Chowdhury, A.; Kunjiappan, S.; Bhattacharjee, C.; Somasundaram, B.; Panneerselvam, T. Biogenic synthesis of Marsilea quadrifolia gold nanoparticles: A study of improved glucose utilization efficiency on 3T3-L1 adipocytes. In Vitro Cell Dev. Biol. Anim. 2017, 53, 483–493. [Google Scholar] [CrossRef]
- Balashanmugam, P.; Mosachristas, K.; Kowsalya, E. In vitro cytotoxicity and antioxidant evaluation of biogenic synthesized gold nanoparticles from Marsilea quadrifolia on lung and ovarian cancer cells. Int. J. Appl. Pharmaceut. 2018, 10, 153–158. [Google Scholar]
- Abbasi, T.; Anuradha, J.; Abbasi, S.A. Utilization of the pernicious aquatic weed salvinia (Salvinia molesta D S Mitchell) in generating gold nanoparticles. Indian J. Biotechnol. 2016, 15, 382–391. [Google Scholar]
- Chumpol, J.; Siri, S. In vivo formation of spherical and rod lead nanoparticles in root cells of water velvet (Azolla pinnata). Biotechnol. Appl. Biochem. 2020, 67, 991–999. [Google Scholar] [CrossRef]
- Wang, D.; Markus, J.; Wang, C.; Kim, Y.J.; Mathiyalagan, R.; Aceituno, V.C.; Ahn, S.; Yang, D.C. Green synthesis of gold and silver nanoparticles using aqueous extract of Cibotium barometz root. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Santhanam, A.; Thangaiah, S.; Narayanan, J. Green synthesis of silver nanoparticles using Cyathea nilgirensis Holttum and their cytotoxic and phytotoxic potentials. Part. Sci. Technol. 2018, 36, 578–582. [Google Scholar]
- Panneerselvam, C.; Murugan, K.; Roni, M.; Aziz, A.T.; Suresh, U.; Rajaganesh, R.; Madhiyazhagan, P.; Subramaniam, J.; Dinesh, D.; Nicoletti, M.; et al. Fern-synthesized nanoparticles in the fight against malaria: LC/MS analysis of Pteridium aquilinum leaf extract and biosynthesis of silver nanoparticles with high mosquitocidal and antiplasmodial activity. Parasitol. Res. 2016, 115, 997–1013. [Google Scholar] [CrossRef]
- Lee, J.H.; Lim, J.M.; Velmurugan, P.; Park, Y.J.; Park, Y.J.; Bang, K.S.; Oh, B.T. Photobiologic-mediated fabrication of silver nanoparticles with antibacterial activity. J. Photochem. Photobiol. B 2016, 162, 93–99. [Google Scholar] [CrossRef]
- Barberia-Roque, L.; Gámez-Espinosa, E.; Viera, M.; Bellotti, N. Assessment of three plant extracts to obtain silver nanoparticles as alternative additives to control biodeterioration of coatings. Int. Biodeterior. Biodegrad. 2019, 141, 52–61. [Google Scholar] [CrossRef]
- Rajaganesh, R.; Murugan, K.; Panneerselvam, C.; Jayashanthini, S.; Aziz, A.T.; Roni, M.; Suresh, U.; Trivedi, S.; Rehman, H.; Higuchi, A.; et al. Fern-synthesized silver nanocrystals: Towards a new class of mosquito oviposition deterrents? Res. Vet. Sci. 2016, 109, 40–51. [Google Scholar] [CrossRef]
- Femi-Adepoju, A.G.; Dada, A.O.; Otun, K.O.; Adepoju, A.O.; Fatoba, O.P. Green synthesis of silver nanoparticles using terrestrial fern (Gleichenia Pectinata (Willd.) C. Presl.): Characterization and antimicrobial studies. Heliyon 2019, 5, e01543. [Google Scholar] [CrossRef]
- Baskaran, X.; Vigila, A.V.G.; Parimelazhagan, T.; Muralidhara-Rao, D.; Zhang, S. Biosynthesis, characterization, and evaluation of bioactivities of leaf extract-mediated biocompatible silver nanoparticles from an early tracheophyte, Pteris tripartita Sw. Int. J. Nanomed. 2016, 11, 5789–5805. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; AlQahtani, F.S.; AlShebly, M.M.; Benelli, G. One-pot and eco-friendly synthesis of silver nanocrystals using Adiantum raddianum: Toxicity against mosquito vectors of medical and veterinary importance. J. Appl. Biomed. 2017, 15, 87–95. [Google Scholar] [CrossRef]
- Omidi, S.; Sedaghat, S.; Tahvildari, K.; Derakhshi, P.; Motiee, F. Biosynthesis of silver nanoparticles with Adiantum capillus-veneris l leaf extract in the batch process and assessment of antibacterial activity. Green Chem. Lett. Rev. 2018, 11, 544–551. [Google Scholar] [CrossRef]
- Rautray, S.; Rajananthini, A.U. Therapeutic potential of green, synthesized gold nanoparticles. BioPharm. Int. 2020, 33, 30–38. [Google Scholar]
- Zamani, S.; Idrees, D.; Jha, B.; Jha, A.K. Green synthesis of metal nanoparticles from adiantum frond: Comparative analysis on cancer cell lines. Nanosci. Nanotechnol. Asia 2020, 10, 806–816. [Google Scholar] [CrossRef]
- Chick, C.N.; Misawa-Suzuki, T.; Suzuki, Y.; Usuki, T. Preparation and antioxidant study of silver nanoparticles of Microsorum pteropus methanol extract. Bioorg. Med. Chem. Lett. 2020, 30, 127526. [Google Scholar] [CrossRef] [PubMed]
- Korbekandi, H.; Chitsazi, M.R.; Asghari, G.; Najafi, R.B.; Badii, A.; Iravani, S. Green biosynthesis of silver nanoparticles using Azolla pinnata whole plant hydroalcoholic extract. Green Process. Synth. 2014, 3, 365–373. [Google Scholar] [CrossRef]
- Kunjiappan, S.; Bhattacharjee, C.; Chowdhury, R. In vitro antioxidant and hepatoprotective potential of Azolla microphylla phytochemically synthesized gold nanoparticles on acetaminophen—Induced hepatocyte damage in Cyprinus carpio L. In Vitro Cell Dev. Biol. Anim. 2015, 51, 630–643. [Google Scholar] [CrossRef]
- Verma, D.K.; Hasan, S.H.; Banik, R.M. Photo-catalyzed and phyto-mediated rapid green synthesis of silver nanoparticles using herbal extract of Salvinia molesta and its antimicrobial efficacy. J. Photochem. Photobiol. B 2016, 155, 51–59. [Google Scholar] [CrossRef]
- Lungulescu, E.M.; Sbarcea, G.; Setnescu, R.; Nicula, N.; Ducu, R.; Lupu, A.M.; Ion, I.; Marinescu, V. Gamma radiation synthesis of colloidal silver nanoparticles. Rev. Chim. 2019, 70, 2826–2830. [Google Scholar] [CrossRef]
- Kasthuri, J.; Veerapandian, S.; Rajendiran, N. Biological synthesis of silver and gold nanoparticles using apiin as reducing agent. Colloids Surf. A 2009, 68, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K. Nanotechnology towards the 21st century. Thin Solid Films 1999, 34, 120–125. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Ortan, A.; Fierascu, I.C.; Fierascu, I. In vitro and in vivo evaluation of antioxidant properties of wild-growing plants. A short review. Curr. Opin. Food Sci. 2018, 24, 1–8. [Google Scholar] [CrossRef]
- Mazumder, K.; Biswas, B.; Raja, I.M.; Fukase, K. A review of cytotoxic plants of the Indian subcontinent and a broad-spectrum analysis of their bioactive compounds. Molecules 2020, 25, 1904. [Google Scholar] [CrossRef]
- Sairi, A.M.M.; Ismail, S.I.; Sukor, A.; Rashid, N.M.N.; Saad, N.; Jamian, S.; Abdullah, S. Cytotoxicity and anticancer activity of Donkioporiella mellea on MRC5 (normal human lung) and A549 (human lung carcinoma) cells lines. Evid. Based Complementary Altern. Med. 2020, 2020, 7415672. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, R.C.; Fierascu, I.; Ortan, A.; Georgiev, M.I.; Sieniawska, E. Innovative approaches for recovery of phytoconstituents from medicinal/aromatic plants and biotechnological production. Molecules 2020, 25, 309. [Google Scholar] [CrossRef]
- Mattos, B.D.; Rojas, O.J.; Magalhães, W.L.E. Biogenic SiO2 in colloidal dispersions via ball milling and ultrasonication. Powder Technol. 2016, 301, 58–64. [Google Scholar] [CrossRef]
- Mattos, B.D.; Rojas, O.J.; Magalhaes, W.L.E. Biogenic silica nanoparticles loaded with neem bark extract as green, slow-release biocide. J. Clean. Prod. 2017, 142, 4206–4213. [Google Scholar] [CrossRef]
- Mattos, B.D.; Gomes, G.R.; de Matos, M.; Ramos, L.P.; Magalhães, W.L.E. Consecutive production of hydroalcoholic extracts, carbohydrates derivatives and silica nanoparticles from Equisetum arvense. Waste Biomass Valor. 2018, 9, 1993–2002. [Google Scholar] [CrossRef]
- Hosseini Mohtasham, N.; Gholizadeh, M. Nano silica extracted from horsetail plant as a natural silica support for the synthesis of H3PW12O40 immobilized on aminated magnetic nanoparticles (Fe3O4@SiO2-EP-NH-HPA): A novel and efficient heterogeneous nanocatalyst for the green one-pot synthesis of pyrano [2,3-c]pyrazole derivatives. Res. Chem. Intermed. 2020, 46, 3037–3066. [Google Scholar]
- Adinarayana, T.V.S.; Mishra, A.; Singhal, I.; Reddy, D.V.R.K. Facile green synthesis of silicon nanoparticles from Equisetum arvense for fluorescence based detection of Fe(III) ions. Nanoscale Adv. 2020, 9, 4125–4132. [Google Scholar] [CrossRef]
- Sola-Rabada, A.; Sahare, P.; Hickman, G.J.; Vasquez, M.; Canham, L.T.; Perry, C.C.; Agarwal, V. Biogenic porous silica and silicon sourced from Mexican Giant Horsetail (Equisetum myriochaetum) and their application as supports for enzyme immobilization. Colloid Surf. B 2018, 166, 195–202. [Google Scholar] [CrossRef]
- Bogireddy, N.K.R.; Sahare, P.; Pal, U.; Méndez, S.F.O.; Gomez, L.M.; Agarwal, V. Platinum nanoparticle-assembled porous biogenic silica 3D hybrid structures with outstanding 4-Nitrophenol degradation performance. Chem. Eng. J. 2020, 388, 124237. [Google Scholar] [CrossRef]
- Gonzaga, M.I.S.; Santos, J.A.G.; Ma, L.Q. Arsenic phytoextraction and hyperaccumulation by fern species. Sci. Agric. 2006, 63, 90–101. [Google Scholar] [CrossRef]
- Indriolo, E.; Na, G.; Ellis, D.; Salt, D.E.; Banks, J.A. A vacuolar arsenite transporter necessary for arsenic tolerance in the arsenic hyperaccumulating fern Pteris vittatais missing in flowering plants. Plant Cell 2010, 22, 2045–2057. [Google Scholar] [CrossRef] [PubMed]
- Lew, T.T.S.; Park, M.; Cui, J.; Strano, M.S. Plant nanobionic sensors for Arsenic detection. Adv Mater. 2021, 33, 2005683. [Google Scholar] [CrossRef] [PubMed]
- Pongthornpruek, S.; Pampasit, S.; Sriprang, N.; Nabheerong, P.; Promtep, K. Heavy metal accumulation in soil and some fern species at Phu Soi Dao National Park, Phitsanulok province, Thailand. NU Sci. J. 2008, 5, 151–164. [Google Scholar]
- Huang, J.; Poynton, C.; Kochian, L.; Elless, M. Phytofiltration of Arsenic from drinking water using Arsenic-hyperaccumulating ferns. Environ. Sci. Technol. 2004, 38, 3412–3417. [Google Scholar] [CrossRef] [PubMed]
- Sune, N.; Maine, M.A.; Sánchez, G.; Caffaratti, S. Cadmium and chromium removal kinetics from solution by two aquatic macrophytes. Environ. Pollut. 2007, 145, 467–473. [Google Scholar] [CrossRef]
- Islam, M.S.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of two potential phytotoxic substances from the aquatic fern Marsilea crenata. J. Plant Biol. 2017, 60, 75–81. [Google Scholar] [CrossRef]
- Azza, A.A.E.M.; Aref, E.M.; Hassanein, H.A.M. Bioaccumulation of heavy metals by the water fern Azolla pinnata. Egypt J. Agric. Res. 2011, 89, 1261–1276. [Google Scholar]
- Taghiganji, M.; Khosravi, M.; Rakhshaee, R. Biosorption of Pb, Cd, Cu and Zn from the wastewater by treated Azolla filiculoides with H2O2/MgCl2. Int. J. Environ. Sci. Technol. 2005, 1, 265–271. [Google Scholar] [CrossRef]
- Sood, A.; Uniyal, P.L.; Prasanna, R.; Ahluwalia, A.S. Phytoremediation potential of aquatic macrophyte. Azolla. Ambio 2012, 41, 122–137. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, A.J.; Zhao, F.J.; Xu, G.Z.; Duan, G.L.; Zhu, Y.G. Arsenic accumulation by the aquatic fern Azolla: Comparison of arsenate uptake, speciation and efflux by A. caroliniana and A. filiculoides. Environ. Pollut. 2008, 156, 1149–1155. [Google Scholar] [CrossRef]
- Fuentes, I.I.; Espadas-Gil, F.; Talavera-May, C.; Fuentes, G.; Santamaría, J.M. Capacity of the aquatic fern (Salvinia minima Baker) to accumulate high concentrations of nickel in its tissues, and its effect on plant physiological processes. Aquat. Toxicol. 2014, 155, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Olguin, E.J.; Sanchez-Galvan, G.; Perez-Perez, T. Assessment of the phytoremediation potential of Salvinia minima baker compared to Spirodela polyrrha in high-strength organic wastewater. Water Air Soil Pollut. 2007, 181, 135–147. [Google Scholar] [CrossRef]
- Qin, B.; Liu, W.; He, E.; Li, Y.; Liu, C.; Ruan, J.; Qiu, R.; Tang, Y. Vacuum pyrolysis method for reclamation of rare earth elements from hyperaccumulator Dicranopteris dichotoma grown in contaminated soil. J. Clean. Prod. 2019, 229, 480–488. [Google Scholar] [CrossRef]
- Ribeiro, T.H.; Rubio, J.; Smith, R.W. A dried hydrophobic aquaphyte as an oil filter for oil/water emulsions. Spill Sci. Technol. Bull. 2003, 8, 483–489. [Google Scholar] [CrossRef]
- Jacobson, M.E.; Chiang, S.Y.; Gueriguian, L.; Westholm, L.R.; Pierson, J.; Zhu, G.; Saunders, F.M. Transformation kinetics of trinitrotoluene conversion in aquatic plants. In Phytoremediation. Transformation and Control of Contaminants; McCutcheon, S.C., Schnoor, J.L., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003; pp. 409–427. [Google Scholar]
- Ena, A.; Carlozzi, P.; Pushparaj, B.; Paperi, R.; Carnevale, S.; Angelo, S. Ability of the aquatic fern Azolla to remove chemical oxygen demand and polyphenols from olive mill wastewater. Grasas Aceites 2007, 58, 34–39. [Google Scholar] [CrossRef]
- Al-Baldawi, I.A.; Sheikh Abdullah, S.R.; Suja, F.; Anuar, N.; Idris, M. Preliminary test of hydrocarbon exposure on Azolla pinnata in phytoremediation process. In Proceedings of the 2nd International Conference on Food Engineering and Biotechnology, Kuala Lumpur, Malaysia, 5–6 May 2012; IACSIT Press: Singapore, 2012; Volume 33. [Google Scholar]
- Kösesakal, T.; Ünal, M.; Kulen, O.; Memon, A.; Yüksel, B. Phytoremediation of petroleum hydrocarbons by using a freshwater fern species Azolla filiculoides Lam. Int. J. Phytoremediation 2016, 18, 467–476. [Google Scholar] [CrossRef]
- Kim, K.J.; Jeong, M.I.; Lee, D.W.; Song, J.S.; Kim, H.D.; Yoo, E.H.; Jeong, S.J.; Han, S.W. Variation in formaldehyde removal efficiency among indoor plant species. HortScience 2010, 45, 1489–1495. [Google Scholar] [CrossRef]
- Eimoori, R.; Zolala, J.; Pourmohiabadi, H.; Noroozian, E.; Mansouri, H. Contribution of Azolla filiculoides to hydrazine elimination from water. Wetl. Ecol. Manag. 2020, 28, 439–447. [Google Scholar] [CrossRef]
- Rizwana, M.; Darshan, M.; Nilesh, D. Phytoremediation of textile waste water using potential wetland plant: Eco-sustainable approach. Int. J. Interdiscip. Multidiscip. Stud. 2014, 1, 130–138. [Google Scholar]
- Torbati, S.; Movafeghi, A.; Khataee, A.R. Biodegradation of C.I. acid blue 92 by Nasturtium officinale: Study of some physiological responses and metabolic fate of dye. Int. J. Phytoremediation 2015, 17, 322–329. [Google Scholar] [CrossRef]
- Singh, J.; Viswanathan, P.N.; Gupta, M.; Devi, S. Uptake and translocation of Cd109 by two aquatic ferns in relation to relative toxic response. Bull. Environ. Contam. Toxicol. 1993, 51, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Popa, K.; Cecal, A.; Humelnicu, D.; Caraus, I.; Draghici, C. Removal of 60Co2+ and 137Cs+ ions from low radioactive solutions using Azolla caroliniana Willd. water fern. Open Chem. 2004, 2, 434–445. [Google Scholar] [CrossRef]
- Forni, C.; Nicolai, M.A.; D’Egidio, D.G. Potential of the small aquatic plants Azolla and Lemna for nitrogenous compounds removal from wastewater. Trans. Ecol. Environ. 2001, 49, 315–324. [Google Scholar]
- EFSA PPR Panel (EFSA Panel on Plant Protection Products and their Residues). Scientific Opinion addressing the state of the science on risk assessment of plant protection products for non-target terrestrial plants. EFSA J. 2014, 12, 3800. [Google Scholar] [CrossRef]
- Boutin, C.; Aya, K.L.; Carpenter, D.; Thomas, P.J.; Rowland, O. Phytotoxicity testing for herbicide regulation: Shortcomings in relation to biodiversity and ecosystem services in agrarian systems. Sci. Total Environ. 2012, 415, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Rowntree, J.K.; Sheffield, E. The effects of Asulam spraying on non-target ferns. Canad. J. Bot. 2005, 83, 1622–1629. [Google Scholar] [CrossRef]
- Newmaster, S.J.; Bell, F.W. The effects of silvicultural disturbances on cryptogam diversity in the boreal-mixed wood forest. Can. J. For. Res. 2002, 32, 38–51. [Google Scholar] [CrossRef]
- Cui, R.; Nam, S.H.; An, Y.J. Salvinia natans: A potential test species for ecotoxicity testing. Environ. Pollut. 2020, 267, 115650. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhong, G.; Zhou, J.; Liu, Y.; Pang, Y.; Cai, H.; Wu, Z. Separate and combined effects of glyphosate and copper on growth and antioxidative enzymes in Salvinia natans (L.) All. Sci. Total Environ. 2019, 655, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- De, A.K.; Ghosh, A.; Debnath, S.C.; Sarkar, B.; Saha, I.; Adak, M.K. Modulation of physiological responses with TiO2 nano-particle in Azolla pinnata R.Br. under 2,4-D toxicity. Mol. Biol. Rep. 2018, 45, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Spanò, C.; Bottega, S.; Sorce, C.; Bartoli, G.; Ruffini Castiglione, M. TiO2 nanoparticles may alleviate cadmium toxicity in co-treatment experiments on the model hydrophyte Azolla filiculoides. Environ. Sci. Pollut. Res. 2019, 26, 29872–29882. [Google Scholar] [CrossRef] [PubMed]
- Catalá, M.; Esteban, M.; Rodríguez-Gil, J.L.; Quintanilla, L.G. Development of a naturally miniaturised testing method based on the mitochondrial activity of fern spores: A new higher plant bioassay. Chemosphere 2009, 77, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.B.; White, S.A.; Klaine, S.J. Interactions of gold nanoparticles with freshwater aquatic macrophytes are size and species dependent. Environ. Toxicol. Chem. 2012, 31, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Zarate-Cruz, G.S.; Zavaleta-Mancera, H.A.; Alarcón, A.; Jiménez-García, L.F. Phytotoxicity of ZnO nanoparticles on the aquatic fern Azolla filiculoides Lam. Agrociencia 2016, 50, 677–691. [Google Scholar]
- Gómez-Garay, A.; Pintos, B.; Manzanera, J.A.; Prada, C.; Martín, L.; Gabriel-Y-Galán, J.M. Nanoceria and bulk cerium oxide effects on the germination of Asplenium adiantum-nigrum spores. For. Syst. 2016, 25, e067. [Google Scholar] [CrossRef]
- Jafarirad, S.; Hajat Ardehjani, P.; Movafeghi, A. Are the green synthesized nanoparticles safe for environment? A case study of aquatic plant Azolla filiculoides as an indicator exposed to magnetite nanoparticles fabricated using microwave hydrothermal treatment and plant extract. J. Environ. Sci. Health A 2019, 54, 506–517. [Google Scholar] [CrossRef]
- Forini, M.M.L.; Antunes, D.R.; Cavalcante, L.A.F.; Pontes, M.S.; Biscalchim, E.R.; Sanches, A.O.; Santiago, E.F.; Fraceto, L.F.; Grillo, R. Fabrication and characterization of a novel herbicide delivery system with magnetic collectability and its phytotoxic effect on photosystem II of aquatic macrophyte. J. Agric. Food Chem. 2020, 68, 11105–11113. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Nandi, P. Antibiotics from the pteridophytes. Sci. Cult. 1951, 16, 328–329. [Google Scholar]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Schepers, E.; Lum, A.; Rice, J.; Yalpani, N.; Gerber, R.; Jiménez-Juárez, N.; Haile, F.; Pascual, A.; Barry, J.; et al. Identification and evaluations of novel insecticidal proteins from plants of the class Polypodiopsida for crop protection against key lepidopteran pests. Toxins 2019, 11, 383. [Google Scholar] [CrossRef] [PubMed]
- Kamaraj, C.; Deepak, P.; Balasubramani, G.; Karthi, S.; Arul, D.; Aiswarya, D.; Amutha, V.; Vimalkumar, E.; Mathivanan, D.; Suseem, S.R.; et al. Target and non-target toxicity of fern extracts against mosquito vectors and beneficial aquatic organisms. Ecotoxicol. Environ. Saf. 2018, 161, 221–230. [Google Scholar] [CrossRef]
- Fasolo, J.M.M.A.; Vizuete, A.F.K.; Rico, E.P.; Rambo, R.B.S.; Toson, N.S.B.; Santos, E.; de Oliveira, D.L.; Gonçalves, C.A.S.; Schapoval, E.E.S.; Heriques, A.T. Anti-inflammatory effect of rosmarinic acid isolated from Blechnum brasiliense in adult zebrafish brain. Comp. Biochem. Phys. C 2021, 239, 108874. [Google Scholar]
- Chandran, G.; Smitha Grace, S.R.; Chauhan, J.B. Fern to pharma: Potential neuroameliorative properties of pteridophytes. In Plant and Human Health; Ozturk, M., Hakeem, K., Eds.; Springer: Cham, Switzerland, 2019; Volume 3, pp. 195–208. [Google Scholar]
- Islam, M.T.; Sarkar, C.; El-Kersh, D.M.; Jamaddar, S.; Uddin, S.J.; Shilpi, J.A.; Mubarak, M.S. Natural products and their derivatives against coronavirus: A review of the non-clinical and pre-clinical data. Phytother. Res. 2020, 34, 2471–2492. [Google Scholar] [CrossRef] [PubMed]
- Gardea-Torresdey, J.L.; Gomez, E.; Peralta-Videa, J.R.; Parsons, J.G.; Troiani, H.; Jose-Yacaman, M. Alfalfa sprouts: a natural source for the synthesis of silver nanoparticles. Langmuir 2003, 19, 1357–1361. [Google Scholar] [CrossRef]


| Class | Subclass | Order (Number) | Families (Number) | Genera (Number) | Species (Number) |
|---|---|---|---|---|---|
| Polypodiopsida | Equisetidae | 1 | 1 | 1 | 15 |
| Ophioglossidae | 2 | 2 | 12 | 129 | |
| Marattiidae | 1 | 1 | 6 | 111 | |
| Polypodiidae | 7 | 44 | 300 | 10,323 | |
| Extant ferns | 11 | 48 | 319 | 10,578 | |
| Fern Used | Applied NPs | Application Results | Ref. |
|---|---|---|---|
| Antimicrobial potential | |||
| Asplenium scolopendrium L. | AgNPs, 10–12 nm | Evaluated against Staphylococcus aureus, Pseudomonas aeruginosa; MIC-1/32 (against S. aureus); MCBE-1/16 (P. aeruginosa); | [17] |
| Dryopteris crassirhizoma Nakai (1920) | AgNPs, spherical, 5–60 nm | Evaluated against Bacillus cereus and P. aeruginosa; best inhibition zones (IZ): 10 mm/250 μg under green LED (B. cereus); 6 mm/250 μg under green LED (P. aeruginosa). | [91] |
| Equisetum arvense L. | AgNPs, nearly spherical, 10–60 nm | Evaluated against Escherichia coli, S. aureus, Candida albicans, commercial probiotic Saccharomyces boulardii; Selective activity against E. coli (effective at low concentrations-0.72 mg/L); | [72] |
| Equisetum arvense L. | AgNPs, spherical, 170.5 nm | evaluated against Salmonella enterica, B. cereus, Listeria monocytogenes, Enterococcus faecium, S. aureus, Aeromonas hydrophila) IZ (mm) = 11.64/10.75/12.46/9.68/12.53/10.80 | [73] |
| Equisetum giganteum L. | AgNPs, spherical, 20 nm | Evaluated against E. coli, S. aureus, Alternaria alternata, Chaetomium globosum; Fungal resistance test and antibacterial biofilm tests after incorporation in waterborne paints; active against all strains; MIC-3.3/13.3/3.3/67.5 μg/mL; paint films inhibited fungal and bacterial biofilm development | [92] |
| Dicranopteris linearis (Burm.f.) Underw | AgNPs, spherical, 40–60 nm | Evaluated against Bacillus subtilis, Klebsiella pneumoniae and Salmonella typhi; IZ (mm) = 21.01/20.1/19 at 75 ppm | [93] |
| Gleichenella pectinata (Willd.) Ching | AgNPs, spherical, 7.51 nm. | Evaluated against P. aeruginosa, E. coli, K. pneumoniae and C. albicans; IZ (mm) = 15/11/10/13 at 5 mM | [94] |
| Nephrolepis biserrata (Sw.) Schott | RuNPs, ~26 nm | Evaluated against Aspergillus flavus; 50% inhibition at 0.6 mL; | [78] |
| Nephrolepis cordifolia (L.) K. Presl | SiO2@Au–Ag composites (200–246 nm SiO2 decorated with 3-nm AuNPs/AgNPs) | Evaluated against E. coli, S. aureus; IZ (mm) = 21/14 | [82] |
| Adiantum philippense L. | AuNPs—spherical, triangular, 33.9 nm in AuNPs–amoxicillin composites | Evaluated against E. coli, S. aureus, Staphylococcus epidermis, B. subtilis, B. cereus, MRSA1, MRSA2, MRSA3, MRSA4; in vivo treatment of systemic MRSA infection : IZ (mm) = 31/30/19/35/38/16/15/12/12 MIC/MBC (mg/L) = 2/4; ½; ½; 16/32; 8/16; 16/32; 16/32; 16/32; 32/32. Survival rate at day 7 post-inoculation 96% | [76] |
| Adiantum philippense L. | AgNPs, quasi-spherical, 10–60 nm | Evaluated against B. subtilis, Listeria monocytogenes, S. aureus, E. coli, K. pneumoniae, Salmonella typhimurium; MIC = 105.41/17.55/17.85/12.36/17.84/28.77 | [77] |
| Pteris ripartite Sw. | AgNPs, different morphologies, 32 nm | Evaluated against B. subtilis, B. cereus, Bacillus megaterium, E. coli, Proteus vulgaris, Serratia marcescens, S. typhi, K. pneumoniae, Vibrio cholerae, Shigella sonnei, Enterobacter aerogenes, P. aeruginosa, A. niger, Aspergillus flavus, Fusarium oxysporum, Penicillium chrysogenum, Rhizopus oryzae; IZ (mm) = 8.33 (B. cereus) − 24.33 (P. aeruginosa) at 10 mg/mL; MIC (at 10 mg/mL, 24 h) = 0.29 (P. aeruginosa) − 1.40 (E. coli); | [95] |
| Adiantum capillus-veneris L. | AgNPs, spherical, 18.4 nm | Evaluated against E. coli and S. aureus; IZ (mm) = 30/19 applied as “thick nanoparticle suspension” | [97] |
| Adiantum capillus-veneris L. | AuNPs | Evaluated against E. coli, P. aeruginosa, Salmonella enterica, S. aureus, B. subtilis, Trichophyton rubrum, Scedosporium apiospermum, Aspergillus fumigates, A. niger, A. flavus; IZ (mm) = 16 (A. fumigates, S. apiospermum, S. enterica) − 21 (E. coli) | [98] |
| Pteris quadriaurita Retz. | AuNPs | Evaluated against E. coli, P. aeruginosa, S. enterica, S. aureus, B. subtilis, T. rubrum, S. apiospermum, A. fumigates, A. niger, A. flavus; IZ (mm) = 14 (T. rubrum) − 18 (S. aureus) | [98] |
| Marsilea quadrifolia L. | AgNPs, spherical, 9–42 nm | Evaluated against E. coli; MIC = 0.5 nM; | [83] |
| Salvinia molesta D. Mitch. | AgNPs, spherical, 12.46 nm | Evaluated against E. coli, S. aureus; IZ (mm) = 21/16 (at 50 ppm); MIC = 10.5/13 mg/L Cell viability loss = 95.8/92.6% after 8 h. at MIC | [103] |
| Cibotium barometz (L.) J. Sm. | AgNPs, spherical, 5–40 nm; | Evaluated against E. coli, S. aureus, S. enterica, P. aeruginosa; IZ (mm, AgNPs) = 16/17.5/12.5/12.5 at 45 μg/disc; | [88] |
| Antioxidant potential | |||
| Asplenium scolopendrium L. | AgNPs, < 50 nm | DPPH inhibition 81.34%/80.93% (rhizomes/leaves mediated NPs) | [47] |
| Equisetum arvense L. | AgNPs, spherical, 170.5 nm | IC0.50 (reducing power activity) = 641.24 μg/mL; IC50 (ABTS/DPPH/NOx) = 210.16/92.90/62.52 μg/mL; | [73] |
| Nephrolepis biserrata (Sw.) Schott | RuNPs, ~26 nm | IC50 (mg/mL, DPPH, ABTS, SORS, HSA assays) = 0.986/0.852/1.265/1.389 | [78] |
| Pteris tripartite Sw. | AgNPs, different morphologies, 32 nm | DPPH, chelating activity, Phosphomolybdenum, ABTS, HPSA assays: 47.90 (mg/L)/61.51 ± 0.61 (mg EDTA/g)/41.94 ± 2.29 (mg AAE/g)/8592.70 ± 614.2 (μmol Trolox/g)/16.20 ± 3.86 (%); | [95] |
| Adiantum capillus-veneris L. | AuNPs | Inhibition: ~90% (DPPH)/~70% (SORS)/~85% (HPSA)/~82% (HSA); | [98] |
| Pteris quadriaurita Retz. | AuNPs | Inhibition: ~81% (DPPH)/~60% (SORS)/~77% (HPSA)/~75% (HSA); | [98] |
| Marsilea quadrifolia L. | AuNPs, spherical, 10–40 nm | IC50 (DPPH) = 50 mg/L; | [85] |
| Leptochilus pteropus (Blume) Fraser-Jenk | AgNPs | IC₅₀ = 47.0 μg/mL (DPPH)/35.8 μg/mL (HPSA) | [100] |
| Cibotium barometz (L.) J. Sm. | AgNPs, spherical, 5–40 nm; AuNPs, spherical, 5–20 nm, | IC50 (DPPH) = 1.4/1.22 mg/mL (AuNPs/AgNPs) | [88] |
| Cytotoxic potential | |||
| Asplenium scolopendrium L. | AgNPs, < 50 nm | Rhizomes extract mediated NPs-progressive time-related mitoinhibitory effect; for both NPs—increased frequency and variability of chromosomal aberrations in the Allium cepa assay | [47] |
| Asplenium scolopendrium L. | AgNPs, 10–12 nm | Significantly higher frequency of the total aberrant cells compared with the negative control sample in the Allium cepa assay | [17] |
| Equisetum arvense L. | AgNPs, nearly spherical, 10–60 nm | MTT assay (MC3T3-E1): Cytotoxic threshold: >2.25/> 4.5 mg L−1, lower for smaller NPs | [72] |
| Equisetum arvense L. | AgNPs spherical, 170.5 nm | Trypan blue exclusion test (HepG2): 20% viability (at 1 mg/mL) | [73] |
| Nephrolepis cordifolia (L.) K. Presl | SiO2@Au–Ag composites (200–246 nm SiO2 decorated with 3 nm AuNPs/AgNPs) | MTT assay (human keratinocyte cells): 95% cell viability at 500 μg/mL | [82] |
| Adiantum philippense L. | AuNPs-spherical, triangular, 33.9 nm. | MTT assay (L929): 81% viability (AuNPs), 79% viability (AuNPs–amoxicillin composites) | [76] |
| Adiantum sp. | AgNPs, AuNPs | MTT assay: Cytotoxicity against MCF-7 cells at different concentrations (2.5 to 100 μg/mL); non-cytotoxic to HEK293 cells | [99] |
| Marsilea quadrifolia L. | AgNPs, spherical, 9–42 nm | MTT assay: Cell death: 40.04% (MCF-7)/55.88% (HeLa), with NP sonication | [83] |
| Marsilea quadrifolia L. | AuNPs, spherical, 17–40 nm | MTT assay (3T3-L1): Cell viability = 71.23% (100 μM) − 84.02% (30μM); glucose uptake = 60.86% | [84] |
| Marsilea quadrifolia L. | AuNPs, spherical, 10–40 nm | MTT assay: IC50 = 45.88/52.01 mg/L (PA-1/A549) | [85] |
| Cibotium barometz (L.) J. Sm. | AgNPs, spherical, 5–40 nm; AuNPs, spherical, 5–20 nm, | MTT assay (RAW264.7 and MCF-7): AuNPs—no cell death at 0.1–10 mg/L; AgNPs—cytotoxicity at ≥ 10 mg/L against RAW264.7 | [88] |
| Alsophila nilgirensis (Holttum) R.M. Tryon | AgNPs, spherical, 45–74 nm | Hatched shrimps bioassay: LC50 = 869.4 μL/10 mL | [89] |
| Larvicidal potential | |||
| Pteridium aquilinum (L.) Kuhn | AgNPs spherical, 35–65 nm | Against Anopheles stephensi Liston, 1901 in laboratory conditions: LC50 of 7.48 ppm (larva I), 10.68 ppm (II), 13.77 ppm (III), 18.45 ppm (IV), and 31.51 ppm (pupae); Larvicidal assays in the field: complete removal of An. stephensi population after 72 h (at 10 × LC50 in water reservoir) | [90] |
| Dicranopteris linearis (Burm.f.) Underw | AgNPs spherical, 40–60 nm | Against Aedes aegypti (Linnaeus in Hasselquist, 1762); laboratory conditions: LC50 = 18.905 ppm (I)/ 20.929 ppm (II)/23.187 ppm (III)/26.312 ppm (IV)/29.328 ppm (pupae); LC90 = 32.140 ppm (I)/35.489 ppm (II)/39.696 ppm (III)/44.418 ppm (IV)/48.511 ppm (pupae) Field larvicidal activity (by application in water storage tanks), ovicidal assay, oviposition deterrent activity 100% reduction in A. aegypti larval populations at 10 × LC50 (after 72 h); No hatching observed at 25 ppm; ER = 94.29% at 30 ppm; | [93] |
| Adiantum raddianum C. Presl | AgNPs, 9.69–13.9 nm | Against mosquito larvae, in laboratory conditions (A. stephensi, A. aegypti, and Culex quinquefasciatus Say, 1823): LC50 = 10.33/11.23/12.19 mg/L Low toxicity against non-target organisms (Diplonychus indicus Venk. et Rao and Gambusia affinis (S. F. Baird and Girard, 1853)), LC50 = 517.86–35.98 mg/L | [96] |
| Phytotoxic potential | |||
| Asplenium scolopendrium L. | AgNPs, 10–12 nm | NPs led to the reduction of the phytotoxic effect of the extracts in Triticum test | [17] |
| Adiantum philippense L. | CuONPs, quasi-spherical, 1–20 nm | Effect on Lens culinaris Medik: 91.26% seed germination, SVI = 4168.43, RWC = 84.37% at 0.025 mg/mL (optimum dose); optimum dose showed highest activity of defense enzymes and total phenolics; higher concentrations of NPs retard all the parameters | [80] |
| Alsophila nilgirensis (Holttum) R.M. Tryon | AgNPs, spherical, 45–74 nm | Effect on Vigna radiata (L.) R. Wilczek and Sorghum vulgare (L.). Seeds: Germination reduction: 38.65/100% (V. radiata and S. vulgare) at 50 mg/L | [89] |
| Antidiabetic potential | |||
| Equisetum arvense L. | AgNPs, spherical, 170.5 nm | IC50 (alpha-glucosidase) = 1.73 μg/mL; | [73] |
| Anti-inflammatory potential | |||
| Pteris tripartite Sw. | AgNPs, different morphologies, 32 nm | Anti-inflammatory activity Wistar albino adult female rats using the carrageenan-induced paw oedema method = 56.36%, 24 h., 100 mg/kg b.w. | [95] |
| Hepatoprotective potential | |||
| Azolla filiculoides Lam. | AuNPs, spherical, 17–40 nm | Significant increase in cell viability compared to the acetaminophen group (hepatocytes damage); significant reduction in the levels of LDH and CAT (dose dependent); AuNPs significantly reduced the GOT and GPT levels (50/10%), significantly increased the levels of GSH-Px and SOD (60–70%), drastically reduced the formation of MDA (60%) and ROS generation | [102] |
| Catalytic properties | |||
| Diplazium esculentum (Retz.) Sw. | AgNPs, different morphologies, 10–45 nm | Degradation of MB and RhB dyes under solar light illumination: complete disappearance of the adsorption peaks after 8 min. | [71] |
| Diplazium esculentum (Retz.) Sw. | AgNPs—spherical, 10–25 nm; AuNPs-different morphologies, 35–75 nm | Degradation of MV 6B, RB, 4-nitro phenol: Ag-98.4/98/96.8%; Au-98.2/98.9/97.3% | [70] |
| Nephrolepis cordifolia (L.) K. Presl | Au–Ag@AgCl, average size 30 nm | Synthesis of quinoline derivatives via three component coupling/hydroarylation/dehydrogenation of arylaldehyde, aniline, and phenyl acetylene derivatives; 96% yield for the composite applied, reaction conditions 9h, at reflux | [81] |
| Nephrolepis cordifolia (L.) K. Presl | SiO2@Au–Ag composites (200–246 nm SiO2 decorated with 3 nm AuNPs/AgNPs) | Solvent-free amidation of carboxylic acid catalyst: 97% yield for the composite applied, reaction conditions—8 h, 100 °C | [82] |
| Other environmental applications | |||
| Nephrolepis cordifolia (L.) K. Presl | FeNPs, spherical, 40–70 nm, other types of iron oxides | Cr(VI) removal: 90.93% | [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fierascu, I.; Fierascu, R.C.; Ungureanu, C.; Draghiceanu, O.A.; Soare, L.C. Application of Polypodiopsida Class in Nanotechnology–Potential towards Development of More Effective Bioactive Solutions. Antioxidants 2021, 10, 748. https://doi.org/10.3390/antiox10050748
Fierascu I, Fierascu RC, Ungureanu C, Draghiceanu OA, Soare LC. Application of Polypodiopsida Class in Nanotechnology–Potential towards Development of More Effective Bioactive Solutions. Antioxidants. 2021; 10(5):748. https://doi.org/10.3390/antiox10050748
Chicago/Turabian StyleFierascu, Irina, Radu Claudiu Fierascu, Camelia Ungureanu, Oana Alexandra Draghiceanu, and Liliana Cristina Soare. 2021. "Application of Polypodiopsida Class in Nanotechnology–Potential towards Development of More Effective Bioactive Solutions" Antioxidants 10, no. 5: 748. https://doi.org/10.3390/antiox10050748
APA StyleFierascu, I., Fierascu, R. C., Ungureanu, C., Draghiceanu, O. A., & Soare, L. C. (2021). Application of Polypodiopsida Class in Nanotechnology–Potential towards Development of More Effective Bioactive Solutions. Antioxidants, 10(5), 748. https://doi.org/10.3390/antiox10050748