
Tissue Antioxidant Status and Lipid Peroxidation Are Related to Dietary Intake of n-3 Polyunsaturated Acids: A Rabbit Model

 ,
,  ,
,  , ,
, ,  ,
, 
 ,
,  , ,
, , 
Abstract
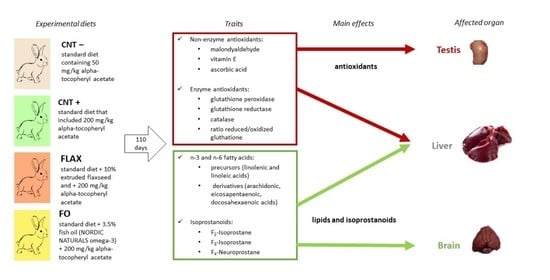
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
- The negative control (CNT−) group was fed ad libitum a standard diet containing 50 mg/kg alpha-tocopheryl acetate (vitamin–mineral premix).
- The positive control (CNT+) group was fed ad libitum a standard diet that also included 200 mg/kg alpha-tocopheryl acetate (50 mg/kg contained in the vitamin–mineral premix + 150 mg/kg of alpha-tocopheryl acetate).
- The FLAX group was fed a standard diet supplemented with 10% extruded flaxseed and 200 mg/kg alpha-tocopheryl acetate (50 mg/kg contained in the vitamin–mineral premix + 150 mg/kg of alpha-tocopheryl acetate).
- The FO group was fed a standard diet supplemented with 3.5% fish oil (NORDIC NATURALS omega-3®; Watsonville, CA, USA) and 200 mg/kg alpha-tocopheryl acetate (50 mg/kg contained in the vitamin–mineral premix + 150 mg/kg of alpha-tocopheryl acetate).
2.2. Sampling of Rabbit Organs
2.3. Analytical Determinations
2.3.1. Determination of the Fatty Acid Content in Rabbit Organs
2.3.2. Determination of the Vitamin E, Ascorbic Acid, and Catalase Levels in Rabbit Organs
2.3.3. Determination of the Glutathione-Related Enzyme Activities and Glutathione Molecular Forms in Rabbit Organs
2.3.4. Determination of Malondialdehyde, F2-Isoprostane, F3-Isoprostane, and F4-Neuroprostane Content in Rabbit Organs
2.4. Statistical Analysis
3. Results
3.1. Oxidative Status and Antioxidant Content of Organs
3.1.1. Non-enzymatic Antioxidants
3.1.2. Enzymatic Antioxidants
3.2. Fatty Acid Profile and Generation of Isoprostanoids
3.2.1. Precursors and Derivatives of n-6 and n-3 Polyunsaturated Fatty Acids
3.2.2. F2-Isoprostane, F3-Isoprostane, and F4-Neuroprostane Generation
3.3. Discriminant Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rodríguez, M.; Rebollar, P.G.; Mattioli, S.; Castellini, C. n-3 PUFA Sources (Precursor/Products): A Review of Current Knowledge on Rabbit. Animals 2019, 9, 806. [Google Scholar] [CrossRef] [PubMed]
- Rambjør, G.S.; Windsor, S.L.; Harris, W.S. Eicosapentaenoic acid is primarily responsible for hypotriglyceridemic effect of fish oil in humans. Lipids 1996, 31, S45–S49. [Google Scholar] [CrossRef]
- Willett, W.C. Dietary fats and coronary heart disease. J. Intern. Med. 2012, 272, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Leaf, A. The electrophysiologic basis for the antiarrhythmic and anticonvulsant effects of n-3 polyunsaturated fatty acids: Heart and brain. Lipids 2001, 36, S107–S110. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D. Fatty acids and immune responses—A new perspective in searching for clues to mechanism. Annu. Rev. Nutr. 2000, 20, 431–456. [Google Scholar] [CrossRef]
- Storlien, L.H.; Jenkins, A.B.; Chisholm, D.J.; Pascoe, W.S.; Khouri, S.; Kraegen, E.W. Influence of Dietary Fat Composition on Development of Insulin Resistance in Rats: Relationship to Muscle Triglyceride and -3 Fatty Acids in Muscle Phospholipid. Diabetes 1991, 40, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Litman, B.; Kim, H.-Y.; Gawrisch, K. Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids 2001, 36, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Signorini, C.; Moretti, E.; Collodel, G. Role of isoprostanes in human male infertility. Syst. Biol. Reprod. Med. 2020, 66, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Collodel, G.; Castellini, C.; Lee, J.C.-Y.; Signorini, C. Relevance of Fatty Acids to Sperm Maturation and Quality. Oxidative Med. Cell. Longev. 2020, 2020, 7038124. [Google Scholar] [CrossRef] [PubMed]
- Wijendran, V.; Hayes, K. Dietary n-6 and n-3 Fatty Acid Balance and Cardiovascular Health. Annu. Rev. Nutr. 2004, 24, 597–615. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health Implications of High Dietary Omega-6 Polyunsaturated Fatty Acids. J. Nutr. Metab. 2012, 2012, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Human requirement for N-3 polyunsaturated fatty acids. Poult. Sci. 2000, 79, 961–970. [Google Scholar] [CrossRef]
- Joint, F.A.O.; Consultation, W.E. Fats and fatty acids in human nutrition. In Proceedings of the Joint FAO/WHO Expert Consultation, Geneva, Switzerland, 10–14 November 2008; Volume 55. [Google Scholar]
- Fard, S.G.; Linderborg, K.M.; Turchini, G.M.; Sinclair, A.J. Comparison of the bioavailability of docosapentaenoic acid (DPA, 22:5n-3) and eicosapentaenoic acid (EPA, 20:5n-3) in the rat. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Ricoult, S.J.H.; Manning, B.D. The multifaceted role of mTORC1 in the control of lipid metabolism. EMBO Rep. 2013, 14, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, S.; Ji, Y.; Chen, H.; Zhang, H.; Chen, W.; Gu, Z.; Chen, Y.Q. Dietary intake of n-3 PUFAs modifies the absorption, distribution and bioavailability of fatty acids in the mouse gastrointestinal tract. Lipids Health Dis. 2017, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The omega-6/omega-3 fatty acid ratio: Health implications. Oléagineux Corps Gras Lipides 2010, 17, 267–275. [Google Scholar] [CrossRef]
- Tang, M.; Zhang, M.; Cai, H.; Li, H.; Jiang, P.; Dang, R.; Liu, Y.; He, X.; Xue, Y.; Cao, L.; et al. Maternal diet of polyunsaturated fatty acid altered the cell proliferation in the dentate gyrus of hippocampus and influenced glutamatergic and serotoninergic systems of neonatal female rats. Lipids Health Dis. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Du Bois, T.; Deng, C.; Bell, W.; Huang, X.-F. Fatty acids differentially affect serotonin receptor and transporter binding in the rat brain. Neuroscience 2006, 139, 1397–1403. [Google Scholar] [CrossRef]
- Lafourcade, M.; Larrieu, T.; Mato, S.; Duffaud, A.; Sepers, M.; Matias, I.; De Smedt-Peyrusse, V.; Labrousse, V.F.; Bretillon, L.; Matute, C.; et al. Nutritional omega-3 deficiency abolishes endocannabinoid-mediated neuronal functions. Nat. Neurosci. 2011, 14, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Castellini, C.; Mattioli, S.; Signorini, C.; Cotozzolo, E.; Noto, D.; Moretti, E.; Brecchia, G.; Dal Bosco, A.; Belmonte, G.; Durand, T.; et al. Effect of Dietary n-3 Source on Rabbit Male Reproduction. Oxidative Med. Cell. Longev. 2019, 2019, 3279670. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, S.; Bosco, A.D.; Maranesi, M.; Petrucci, L.; Rebollar, P.G.; Castellini, C. Dietary fish oil and flaxseed for rabbit does: Fatty acids distribution and Δ6-desaturase enzyme expression of different tissues. Animal 2019, 13, 1934–1942. [Google Scholar] [CrossRef] [PubMed]
- Joffre, C.; Grégoire, S.; De Smedt, V.; Acar, N.; Bretillon, L.; Nadjar, A.; Layé, S. Modulation of brain PUFA content in different experimental models of mice. Prostaglandins Leukot. Essent. Fat. Acids 2016, 114, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Niki, E.; Yoshida, Y.; Saito, Y.; Noguchi, N. Lipid peroxidation: Mechanisms, inhibition, and biological effects. Biochem. Biophys. Res. Commun. 2005, 338, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Milne, G.L.; Dai, Q.; Roberts, L.J. The isoprostanes—25 years later. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2015, 1851, 433–445. [Google Scholar] [CrossRef]
- Ahmed, O.S.; Galano, J.-M.; Pavlickova, T.; Revol-Cavalier, J.; Vigor, C.; Lee, J.C.-Y.; Oger, C.; Durand, T. Moving forward with isoprostanes, neuroprostanes and phytoprostanes: Where are we now? Essays Biochem. 2020, 64, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Richardson, R.I.; Nute, G.R.; Fisher, A.V.; Campo, M.M.; Kasapidou, E.; Sheard, P.R.; Enser, M. Effects of fatty acids on meat quality: A review. Meat Sci. 2004, 66, 21–32. [Google Scholar] [CrossRef]
- Bosco, A.D.; Castellini, C.; Bernardini, M. Nutritional Quality of Rabbit Meat as Affected by Cooking Procedure and Dietary Vitamin E. J. Food Sci. 2001, 66, 1047–1051. [Google Scholar] [CrossRef]
- Candellone, A.; Peiretti, P.G.; Binello, A.; Bergagna, S.; Meineri, G. Effect of linseed diet and hazelnut skin supplementation on oxidative status and blood serum metabolites in rabbits. Prog. Nutr. 2019, 21, 631–640. [Google Scholar]
- Cardinali, R.; Cullere, M.; Bosco, A.D.; Mugnai, C.; Ruggeri, S.; Mattioli, S.; Castellini, C.; Marinucci, M.T.; Zotte, A.D. Oregano, rosemary and vitamin E dietary supplementation in growing rabbits: Effect on growth performance, carcass traits, bone development and meat chemical composition. Livest. Sci. 2015, 175, 83–89. [Google Scholar] [CrossRef]
- Mattioli, S.; Duarte, J.M.M.; Castellini, C.; D’Amato, R.; Regni, L.; Proietti, P.; Businelli, D.; Cotozzolo, E.; Rodrigues, M.; Bosco, A.D. Use of olive leaves (whether or not fortified with sodium selenate) in rabbit feeding: Effect on performance, carcass and meat characteristics, and estimated indexes of fatty acid metabolism. Meat Sci. 2018, 143, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Bosco, A.D.; Castellini, C.; Bianchi, L.; Mugnai, C. Effect of dietary α-linolenic acid and vitamin E on the fatty acid composition, storage stability and sensory traits of rabbit meat. Meat Sci. 2004, 66, 407–413. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Gerencsér, Z.; Szendro, Z.; Mugnai, C.; Cullere, M.; Kovàcs, M.; Ruggeri, S.; Mattioli, S.; Castellini, C.; Dalle Zotte, A. Effect of dietary supplementation of Spirulina (Arthrospira platensis) and Thyme (Thymus vulgaris) on rabbit meat appearance, oxidative stability and fatty acid profile during retail display. Meat Sci. 2014, 96, 114–119. [Google Scholar] [CrossRef]
- Bosco, A.D.; Castellini, C.; Martino, M.; Mattioli, S.; Marconi, O.; Sileoni, V.; Ruggeri, S.; Tei, F.; Benincasa, P. The effect of dietary alfalfa and flax sprouts on rabbit meat antioxidant content, lipid oxidation and fatty acid composition. Meat Sci. 2015, 106, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.; Barbas, C. Vitamin E: Action, metabolism and perspectives. J. Physiol. Biochem. 2001, 57, 43–56. [Google Scholar] [CrossRef]
- Viola, V.; Pilolli, F.; Piroddi, M.; Pierpaoli, E.; Orlando, F.; Provinciali, M.; Betti, M.; Mazzini, F.; Galli, F. Why tocotrienols work better: Insights into the in vitro anti-cancer mechanism of vitamin E. Genes Nutr. 2011, 7, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Cappai, M.G.; Pudda, F.; Wolf, P.; Accioni, F.; Boatto, G.; Pinna, W. Variation of Hematochemical Profile and Vitamin E Status in Feral Giara Horses from Free Grazing in the Wild to Hay Feeding during Captivity. J. Equine Veter. Sci. 2020, 94, 103220. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [CrossRef]
- Schüep, W.; Rettenmaier, R. Analysis of vitamin E homologs in plasma and tissue: High-performance liquid chromatography. Methods Enzymol. 1994, 234, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.A. Determination of ascorbic acid and uric acid in plasma by high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1994, 657, 197–200. [Google Scholar] [CrossRef]
- Johansson, L.H.; Borg, L.A.H. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef]
- Cribb, A.E.; Leeder, J.; Spielberg, S.P. Use of a microplate reader in an assay of glutathione reductase using 5,5′-dithiobis(2-nitrobenzoic acid). Anal. Biochem. 1989, 183, 195–196. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. [12] Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Nencini, C.; Giorgi, G.; Micheli, L. Protective effect of silymarin on oxidative stress in rat brain. Phytomedicine 2007, 14, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Shara, M.A.; Dickson, P.H.; Bagchi, D.; Stohs, S.J. Excretion of formaldehyde, malondialdehyde, acetaldehyde and acetone in the urine of rats in response to 2,3,7,8-tetrachlorodibenzo-p-dioxin, paraquat, endrin and carbon tetrachloride. J. Chromatogr. B Biomed. Sci. Appl. 1992, 576, 221–233. [Google Scholar] [CrossRef]
- Longini, M.; Moretti, E.; Signorini, C.; Noto, D.; Iacoponi, F.; Collodel, G. Relevance of seminal F2-dihomo-IsoPs, F2-IsoPs and F4-NeuroPs in idiopathic infertility and varicocele. Prostaglandins Other Lipid Mediat. 2020, 149, 106448. [Google Scholar] [CrossRef] [PubMed]
- Signorini, C.; Cardile, V.; Pannuzzo, G.; Graziano, A.C.E.; Durand, T.; Galano, J.-M.; Oger, C.; Leoncini, S.; Cortelazzo, A.; Lee, J.C.-Y.; et al. Increased isoprostanoid levels in brain from murine model of Krabbe disease—Relevance of isoprostanes, dihomo-isoprostanes and neuroprostanes to disease severity. Free. Radic. Biol. Med. 2019, 139, 46–54. [Google Scholar] [CrossRef]
- StataCorp, L.P. StataCorp Stata Statistical Software: Release 14; StataCorp LP.: College Station, TX, USA, 2015. [Google Scholar]
- Garson, D.G. Factor Analysis: Statnotes. In From North Carolina State University Public Administration Program; North Carolina University Publishing: Chapel Hill, NC, USA, 2008; ISBN 0028-4793. [Google Scholar]
- Agradi, S.; Curone, G.; Negroni, D.; Vigo, D.; Brecchia, G.; Bronzo, V.; Panseri, S.; Chiesa, L.M.; Peric, T.; Danes, D.; et al. Determination of Fatty Acids Profile in Original Brown Cows Dairy Products and Relationship with Alpine Pasture Farming System. Animals 2020, 10, 1231. [Google Scholar] [CrossRef]
- Zappaterra, M.; Menchetti, L.; Costa, L.N.; Padalino, B. Do Camels (Camelus dromedarius) Need Shaded Areas? A Case Study of the Camel Market in Doha. Animals 2021, 11, 480. [Google Scholar] [CrossRef] [PubMed]
- Lands, B. A critique of paradoxes in current advice on dietary lipids. Prog. Lipid Res. 2008, 47, 77–106. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, K.H.; Slater, T.F. An introduction to free radical biochemistry. Br. Med. Bull. 1993, 49, 481–493. [Google Scholar] [CrossRef]
- Moazamian, R.; Polhemus, A.; Connaughton, H.; Fraser, B.; Whiting, S.; Gharagozloo, P.; Aitken, R.J. Oxidative stress and human spermatozoa: Diagnostic and functional significance of aldehydes generated as a result of lipid peroxidation. Mol. Hum. Reprod. 2015, 21, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Antioxidant defence mechanisms: From the beginning to the end (of the beginning). Free. Radic. Res. 1999, 31, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Castellini, C.; Bosco, A.D.; Mattioli, S.; Davidescu, M.; Corazzi, L.; Macchioni, L.; Rimoldi, S.; Terova, G. Activity, Expression, and Substrate Preference of the Δ6-Desaturase in Slow- or Fast-Growing Rabbit Genotypes. J. Agric. Food Chem. 2016, 64, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Moallem, U. Invited review: Roles of dietary n-3 fatty acids in performance, milk fat composition, and reproductive and immune systems in dairy cattle. J. Dairy Sci. 2018, 101, 8641–8661. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C.C.R.; Caramujo, M.J. The Various Roles of Fatty Acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef]
- Morrow, J.D.; Awad, J.A.; Boss, H.J.; Blair, I.A.; Roberts, L.J. Non-cyclooxygenase-derived prostanoids (F2-isoprostanes) are formed in situ on phospholipids. Proc. Natl. Acad. Sci. USA 1992, 89, 10721–10725. [Google Scholar] [CrossRef]
- Roberts, L.J.; Morrow, J.D. Measurement of F2-isoprostanes as an index of oxidative stress in vivo. Free. Radic. Biol. Med. 2000, 28, 505–513. [Google Scholar] [CrossRef]
- Praticò, D.; Lawson, J.A.; Rokach, J.; Fitzgerald, G.A. The isoprostanes in biology and medicine. Trends Endocrinol. Metab. 2001, 12, 243–247. [Google Scholar] [CrossRef]
- Devasagayam, T.P.A.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Physicians India 2004, 52, 794–804. [Google Scholar] [PubMed]
- Harats, D.; Chevion, S.; Nahir, M.; Norman, Y.; Sagee, O.; Berry, E.M. Citrus fruit supplementation reduces lipoprotein oxidation in young men ingesting a diet high in saturated fat: Presumptive evidence for an interaction between vitamins C and E in vivo. Am. J. Clin. Nutr. 1998, 67, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Interaction of Ascorbate and α-Tocopherol. Ann. N. Y. Acad. Sci. 1987, 498, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.H.; Croft, K.D. Vitamin E metabolism. Mol. Asp. Med. 2007, 28, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. ASCORBATE AND GLUTATHIONE: Keeping Active Oxygen Under Control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding oxidants and antioxidants: Classical team with new players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef] [PubMed]
- Aslani, B.A.; Ghobadi, S. Studies on oxidants and antioxidants with a brief glance at their relevance to the immune system. Life Sci. 2016, 146, 163–173. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Units | CNT− | CNT+ | FLAX | FO |
|---|---|---|---|---|---|
| Dehydrated alfalfa meal | g/kg | 300 | 300 | 380 | 380 |
| Soybean meal 44% | g/kg | 150 | 150 | 100 | 150 |
| Barley meal | g/kg | 410 | 410 | 310 | 335 |
| Wheat bran | g/kg | 52 | 52 | 52 | 52 |
| Soybean oil | g/kg | 30 | 30 | - | - |
| Extruded flaxseed | g/kg | - | - | 100 | - |
| Fish oil 1 | g/kg | - | - | - | 35 |
| Beet molasses | g/kg | 20 | 20 | 10 | 10 |
| Calcium carbonate | g/kg | 7 | 7 | 7 | 7 |
| Calcium diphosphate | g/kg | 13.5 | 13.5 | 13.5 | 13.5 |
| Salt | g/kg | 7 | 7 | 7 | 7 |
| DL-methionine | g/kg | 0.5 | 0.5 | 0.5 | 0.5 |
| Vitamin–mineral premix 2 | g/kg | 10 | 10 | 10 | 10 |
| Alpha-tocopheryl acetate | mg/kg | - | 150 | 150 | 150 |
| Crude protein | g/kg | 175 | 174 | 174 | 175 |
| Ether extract | g/kg | 480 | 477 | 472 | 425 |
| Crude fibre | g/kg | 124 | 122 | 137 | 130 |
| Ash | g/kg | 89 | 89 | 84 | 90 |
| Organ. | DA1 | DA2 | ||
|---|---|---|---|---|
| Function 1 | Function 2 | Function 1 | Function 2 | |
| Liver | 6.418 | 0.260 | 3.713 | 1.241 |
| Testis | −2.546 | −1.913 | −5.483 | 0.332 |
| Brain | −4.296 | 1.334 | 1.770 | −1.574 |
| Function | ||
|---|---|---|
| 1 | 2 | |
| GPX | 0.567 * | 0.441 |
| Vitamin E | 0.340 * | −0.217 |
| Catalase | 0.326 * | −0.177 |
| GR | −0.109 | 0.427 * |
| AA | −0.096 | 0.326 * |
| Percentage of variance | 93.0% | 7.0% |
| Wilks’ lambda | 0.014 | 0.353 |
| Significance (p) | <0.001 | <0.001 |
| Function | ||
|---|---|---|
| 1 | 2 | |
| F2-IsoPs | 0.445 * | −0.373 |
| LCPn-6 | −0.199 * | −0.103 |
| F3-IsoPs | −0.089 | 0.544 * |
| F4-NeuroPs | 0.418 | 0.510 * |
| LCPn-3 | 0.119 | −0.273 * |
| Percentage of variance | 91.9% | 8.1% |
| Wilks’ lambda | 0.023 | 0.407 |
| Significance | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioli, S.; Collodel, G.; Signorini, C.; Cotozzolo, E.; Noto, D.; Cerretani, D.; Micheli, L.; Fiaschi, A.I.; Brecchia, G.; Menchetti, L.; et al. Tissue Antioxidant Status and Lipid Peroxidation Are Related to Dietary Intake of n-3 Polyunsaturated Acids: A Rabbit Model. Antioxidants 2021, 10, 681. https://doi.org/10.3390/antiox10050681
Mattioli S, Collodel G, Signorini C, Cotozzolo E, Noto D, Cerretani D, Micheli L, Fiaschi AI, Brecchia G, Menchetti L, et al. Tissue Antioxidant Status and Lipid Peroxidation Are Related to Dietary Intake of n-3 Polyunsaturated Acids: A Rabbit Model. Antioxidants. 2021; 10(5):681. https://doi.org/10.3390/antiox10050681
Chicago/Turabian StyleMattioli, Simona, Giulia Collodel, Cinzia Signorini, Elisa Cotozzolo, Daria Noto, Daniela Cerretani, Lucia Micheli, Anna Ida Fiaschi, Gabriele Brecchia, Laura Menchetti, and et al. 2021. "Tissue Antioxidant Status and Lipid Peroxidation Are Related to Dietary Intake of n-3 Polyunsaturated Acids: A Rabbit Model" Antioxidants 10, no. 5: 681. https://doi.org/10.3390/antiox10050681
APA StyleMattioli, S., Collodel, G., Signorini, C., Cotozzolo, E., Noto, D., Cerretani, D., Micheli, L., Fiaschi, A. I., Brecchia, G., Menchetti, L., Moretti, E., Oger, C., De Felice, C., & Castellini, C. (2021). Tissue Antioxidant Status and Lipid Peroxidation Are Related to Dietary Intake of n-3 Polyunsaturated Acids: A Rabbit Model. Antioxidants, 10(5), 681. https://doi.org/10.3390/antiox10050681