
A Systematic Review of Serum γ-Glutamyltransferase as a Prognostic Biomarker in Patients with Genitourinary Cancer



Abstract
1. Introduction
2. Structure, Functions, and Expression of GGT
2.1. GGT Family Enzymes and Structure
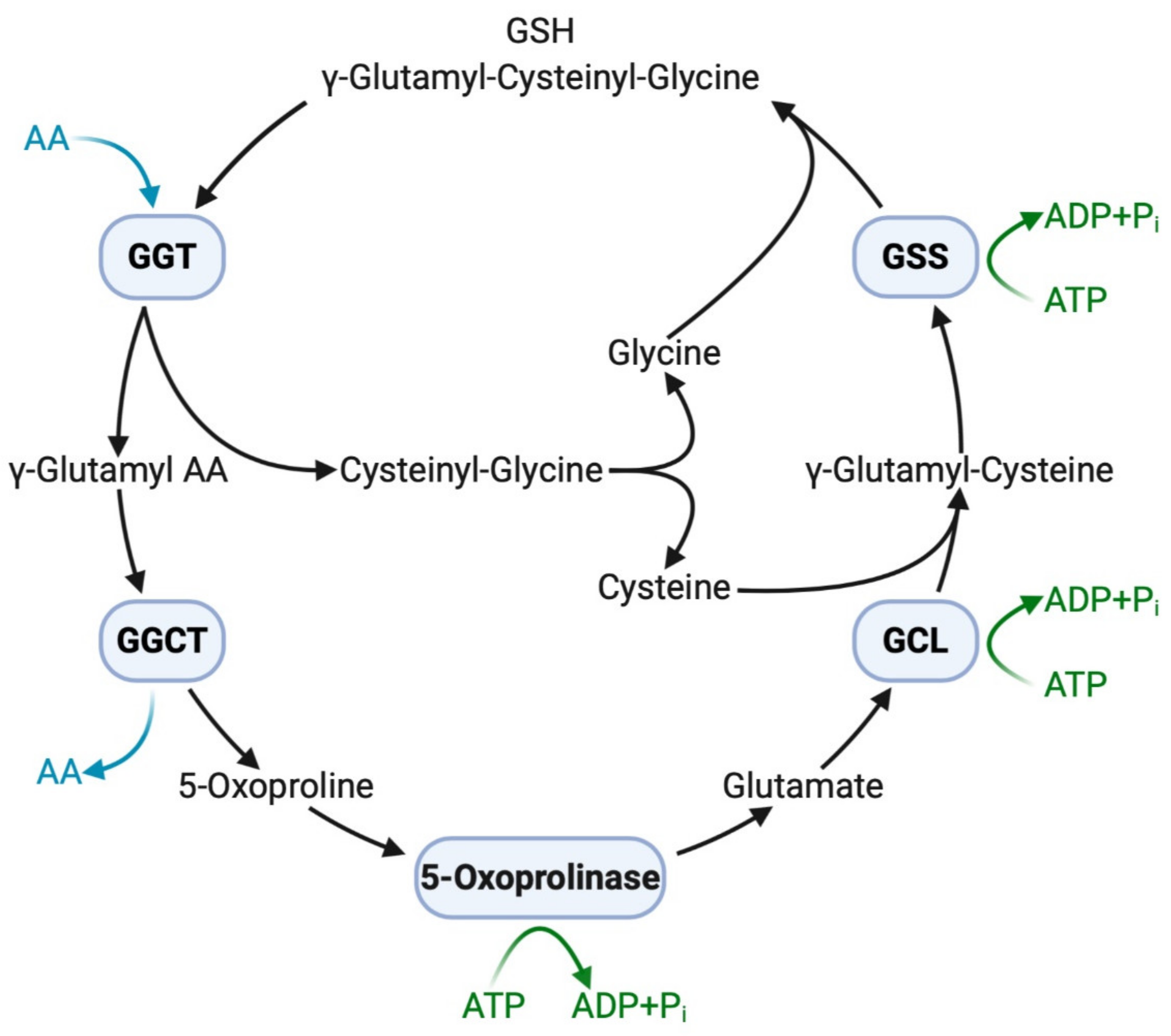
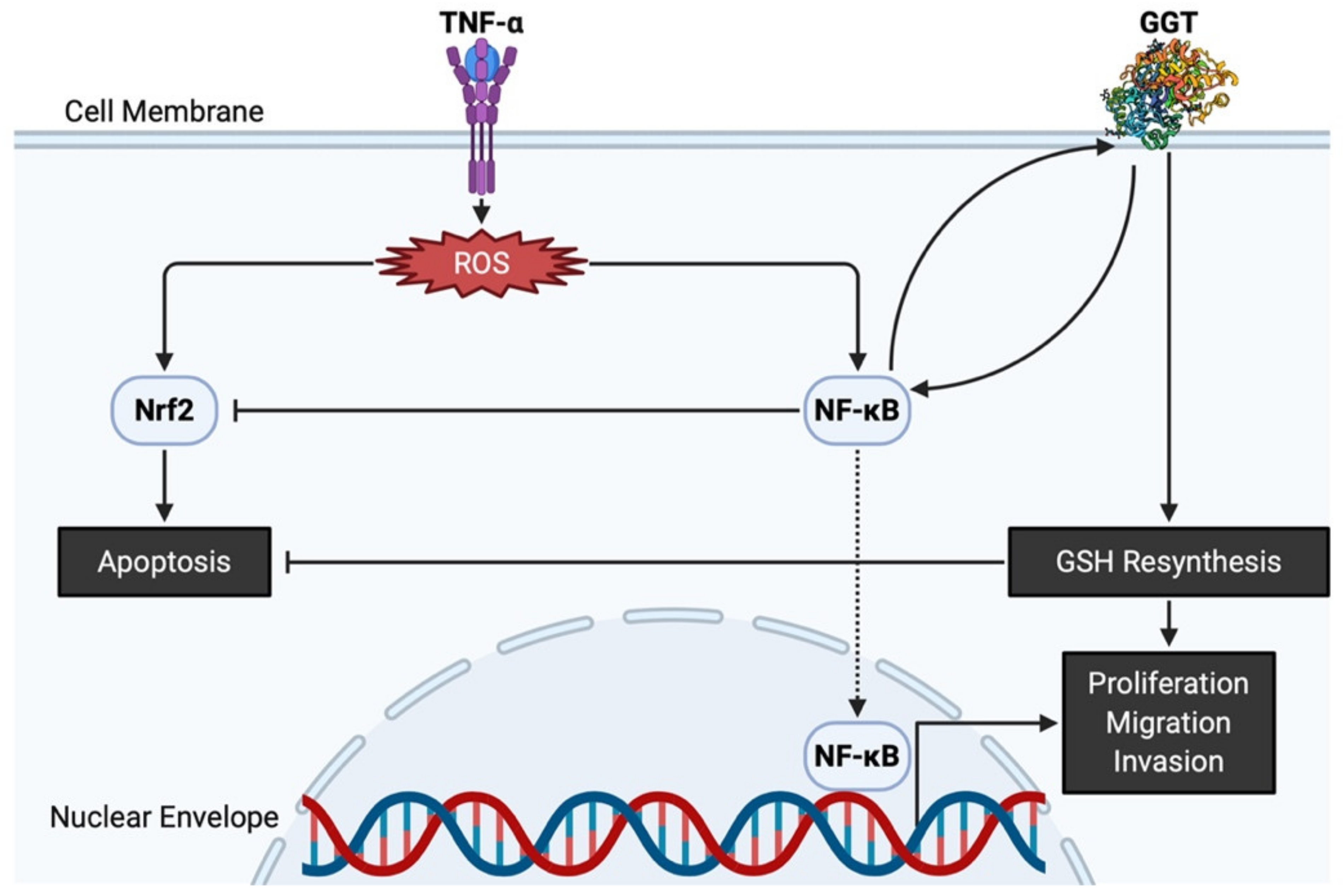
2.2. Functions of GGT
2.3. Expression of GGT in Normal Cells and Cancer Cells in Urogenital Organs
3. Potential Clinical Application of GGT Inhibitors
3.1. Competitive GGT Inhibitors
3.2. Uncompetitive GGT Inhibitors
4. Elevation of Serum GGT in Patients with GU Cancer
4.1. Sources of Serum GGT
4.2. Elevation of Serum GGT in Patients with GU Cancer
4.3. Serum GGT as a Prognostic Biomarker in Patients with Cancer
5. A Systematic Review of Serum GGT in Patients with GU Cancer
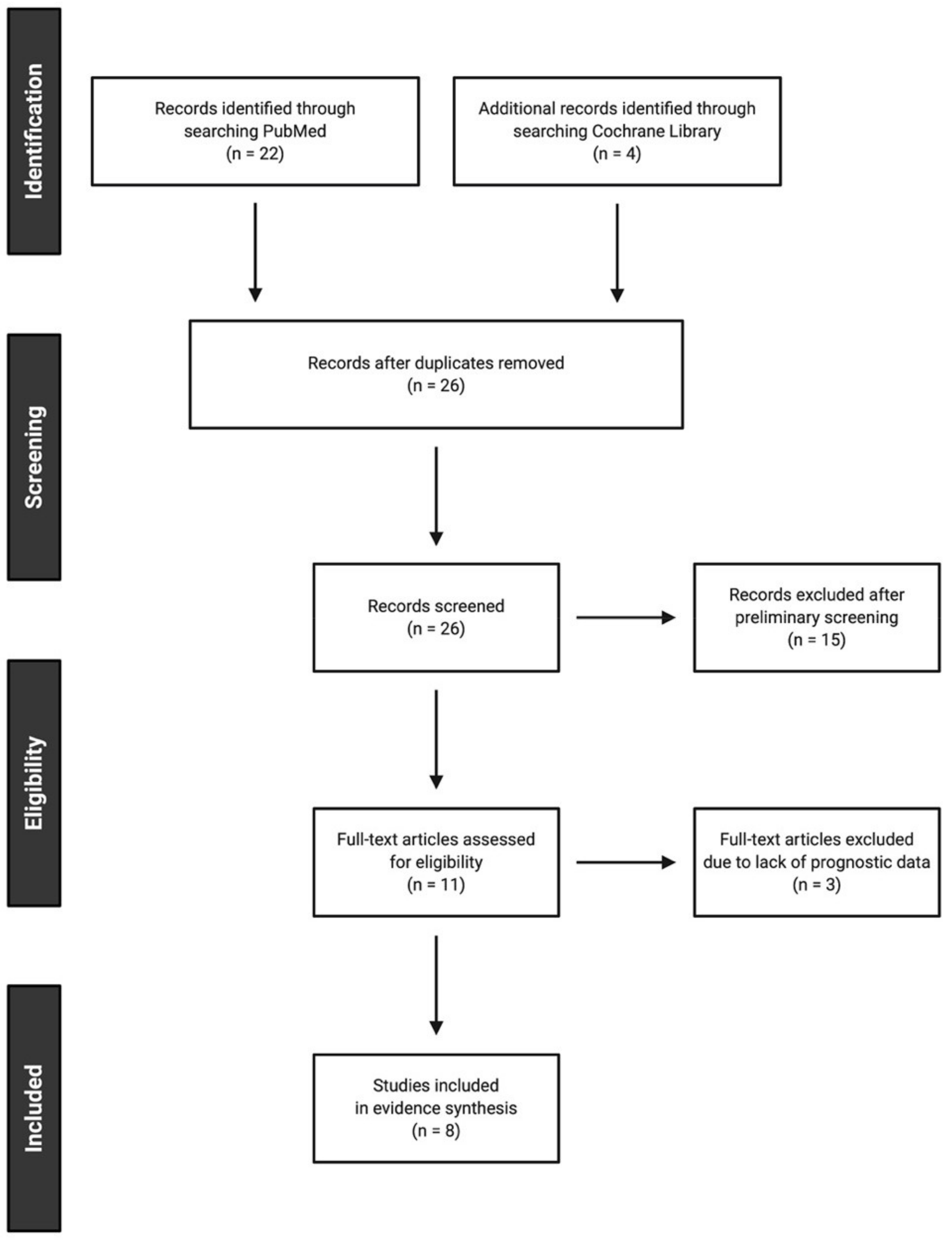
5.1. Study Selection
5.2. Study Characteristics
5.3. Evidence Synthesis
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meister, A.; Anderson, M.E. Glutathione. Annu Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Hagen, T.M.; Aw, T.Y.; Jones, D.P. Glutathione uptake and protection against oxidative injury in isolated kidney cells. Kidney Int. 1988, 34, 74–81. [Google Scholar] [CrossRef]
- Meister, A. On the enzymology of amino acid transport. Science 1973, 180, 33–39. [Google Scholar] [CrossRef]
- Koenig, G.; Seneff, S. Gamma-glutamyltransferase: A predictive biomarker of cellular antioxidant inadequacy and disease risk. Dis. Markers 2015, 2015, 818570. [Google Scholar] [CrossRef]
- Ou, Y.; Huang, J.; Yang, L. The prognostic significance of pretreatment serum gamma-glutamyltranspeptidase in primary liver cancer: A meta-analysis and systematic review. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Sun, P.; Li, Y.; Chang, L.; Tian, X. Prognostic and clinicopathological significance of Gamma-Glutamyltransferase in patients with hepatocellular carcinoma: A PRISMA-compliant meta-analysis. Medicine 2019, 98, e15603. [Google Scholar] [CrossRef]
- Ramankulov, A.; Lein, M.; Kristiansen, G.; Meyer, H.A.; Loening, S.A.; Jung, K. Elevated plasma osteopontin as marker for distant metastases and poor survival in patients with renal cell carcinoma. J. Cancer Res. Clin. Oncol. 2007, 133, 643–652. [Google Scholar] [CrossRef]
- Hofbauer, S.L.; Stangl, K.I.; De Martino, M.; Lucca, I.; Haitel, A.; Shariat, S.F.; Klatte, T. Pretherapeutic gamma-glutamyltransferase is an independent prognostic factor for patients with renal cell carcinoma. Br. J. Cancer 2014, 111, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Dalpiaz, O.; Pichler, M.; Mrsic, E.; Reitz, D.; Krieger, D.; Venturino, L.; Bezan, A.; Stojakovic, T.; Pummer, C.; Zigeuner, R.; et al. Preoperative serum-gamma-glutamyltransferase (GGT) does not represent an independent prognostic factor in a European cohort of patients with non-metastatic renal cell carcinoma. J. Clin. Pathol. 2015, 68, 547–551. [Google Scholar] [CrossRef]
- Luo, C.; Xu, B.; Fan, Y.; Yu, W.; Zhang, Q.; Jin, J. Preoperative gamma-glutamyltransferase is associated with cancer-specific survival and recurrence-free survival of nonmetastatic renal cell carcinoma with venous tumor thrombus. Biomed. Res. Int. 2017, 2017, 3142926. [Google Scholar] [CrossRef] [PubMed]
- Takemura, K.; Yuasa, T.; Inamura, K.; Amori, G.; Koga, F.; Board, P.G.; Yonese, J. Impact of serum γ-glutamyltransferase on overall survival in patients with metastatic renal cell carcinoma in the era of targeted therapy. Target. Oncol. 2020, 15, 347–356. [Google Scholar] [CrossRef]
- Ishiyama, Y.; Kondo, T.; Tachibana, H.; Ishihara, H.; Fukuda, H.; Yoshida, K.; Takagi, T.; Iizuka, J.; Tanabe, K. Predictive role of γ-glutamyltransferase in patients receiving nivolumab therapy for metastatic renal cell carcinoma. Int. J. Clin. Oncol. 2021, 26, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Takemura, K.; Ito, M.; Nakanishi, Y.; Kataoka, M.; Sakamoto, K.; Suzuki, H.; Tobisu, K.-I.; Koga, F. Serum γ-glutamyltransferase as a prognostic biomarker in metastatic castration-resistant prostate cancer treated with enzalutamide. Anticancer Res. 2019, 39, 5773–5780. [Google Scholar] [CrossRef] [PubMed]
- Takemura, K.; Fukushima, H.; Ito, M.; Kataoka, M.; Nakanishi, Y.; Sakamoto, K.; Suzuki, H.; Tobisu, K.-I.; Koga, F. Prognostic significance of serum γ-glutamyltransferase in patients with advanced urothelial carcinoma. Urol. Oncol. 2019, 37, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Heisterkamp, N.; Groffen, J.; Warburton, D.; Sneddon, T.P. The human gamma-glutamyltransferase gene family. Hum. Genet. 2008, 123, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Heisterkamp, N.; Rajpert-De Meyts, E.; Uribe, L.; Forman, H.J.; Groffen, J. Identification of a human gamma-glutamyl cleaving enzyme related to, but distinct from, gamma-glutamyl transpeptidase. Proc. Natl. Acad. Sci. USA 1991, 88, 6303–6307. [Google Scholar] [CrossRef]
- Hanigan, M.H.; Gillies, E.M.; Wickham, S.; Wakeham, N.; Wirsig-Wiechmann, C.R. Immunolabeling of gamma-glutamyl transferase 5 in normal human tissues reveals that expression and localization differ from gamma-glutamyl transferase 1. Histochem. Cell Biol. 2015, 143, 505–515. [Google Scholar] [CrossRef]
- Wickham, S.; West, M.B.; Cook, P.F.; Hanigan, M.H. Gamma-glutamyl compounds: Substrate specificity of gamma-glutamyl transpeptidase enzymes. Anal. Biochem. 2011, 414, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.O.; Williams, P.; Wagner, E.; Chang, D.S.; Wild, K.; Colwell, R.E.; Wolff, J.A. Mice with genetic gamma-glutamyl transpeptidase deficiency exhibit glutathionuria, severe growth failure, reduced life spans, and infertility. J. Biol. Chem. 1997, 272, 12560–12567. [Google Scholar] [CrossRef]
- Levasseur, R.; Barrios, R.; Elefteriou, F.; Glass, D.A.; Lieberman, M.W.; Karsenty, G. Reversible skeletal abnormalities in gamma-glutamyl transpeptidase-deficient mice. Endocrinology 2003, 144, 2761–2764. [Google Scholar] [CrossRef][Green Version]
- Shi, Z.Z.; Han, B.; Habib, G.M.; Matzuk, M.M.; Lieberman, M.W. Disruption of gamma-glutamyl leukotrienase results in disruption of leukotriene D(4) synthesis in vivo and attenuation of the acute inflammatory response. Mol. Cell Biol. 2001, 21, 5389–5395. [Google Scholar] [CrossRef] [PubMed]
- Darin, N.; Leckstrom, K.; Sikora, P.; Lindgren, J.; Almen, G.; Asin-Cayuela, J. Gamma-glutamyl transpeptidase deficiency caused by a large homozygous intragenic deletion in GGT1. Eur. J. Hum. Genet. 2018, 26, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Grimm, C.; Hofstetter, G.; Aust, S.; Mutz-Dehbalaie, I.; Bruch, M.; Heinze, G.; Rahhal-Schupp, J.; Reinthaller, A.; Concin, N.; Polterauer, S. Association of gamma-glutamyltransferase with severity of disease at diagnosis and prognosis of ovarian cancer. Br. J. Cancer 2013, 109, 610–614. [Google Scholar] [CrossRef]
- Bui, T.T.; Nitta, R.T.; Kahn, S.A.; Razavi, S.M.; Agarwal, M.; Aujla, P.; Gholamin, S.; Recht, L.; Li, G. γ-Glutamyl transferase 7 is a novel regulator of glioblastoma growth. BMC Cancer 2015, 15, 225. [Google Scholar] [CrossRef]
- Hanigan, M.H.; Frierson, H.F. Immunohistochemical detection of gamma-glutamyl transpeptidase in normal human tissue. J. Histochem. Cytochem. 1996, 44, 1101–1108. [Google Scholar] [CrossRef]
- Oster, T.; Visvikis, A.; Thioudellet, C.; Fournel-Gigleux, S.; Wellman, M.; Siest, G. Establishment of a V79 transfected cell line highly producing recombinant human gamma-glutamyltransferase. Toxicology 1993, 82, 151–167. [Google Scholar] [CrossRef]
- West, M.B.; Chen, Y.; Wickham, S.; Heroux, A.; Cahill, K.; Hanigan, M.H.; Mooers, B.H. Novel insights into eukaryotic γ-glutamyltranspeptidase 1 from the crystal structure of the glutamate-bound human enzyme. J. Biol. Chem. 2013, 288, 31902–31913. [Google Scholar] [CrossRef] [PubMed]
- Terzyan, S.S.; Burgett, A.W.; Heroux, A.; Smith, C.A.; Mooers, B.H.; Hanigan, M.H. Human γ-glutamyl transpeptidase 1: Structures of the free enzyme, inhibitor-bound tetrahedral transition states, and glutamate-bound enzyme reveal novel movement within the active site during catalysis. J. Biol. Chem. 2015, 290, 17576–17586. [Google Scholar] [CrossRef]
- Meister, A.; Tate, S.S.; Griffith, O.W. Gamma-glutamyl transpeptidase. Methods Enzymol. 1981, 77, 237–253. [Google Scholar] [PubMed]
- Lewis, R.A.; Austen, K.F.; Soberman, R.J. Leukotrienes and other products of the 5-lipoxygenase pathway. Biochemistry and relation to pathobiology in human diseases. N. Engl. J. Med. 1990, 323, 645–655. [Google Scholar]
- Angeli, V.; Tacito, A.; Paolicchi, A.; Barsacchi, R.; Franzini, M.; Baldassini, R.; Vecolid, C.; Pompella, A.; Bramanti, E.; Bramanti, E. A kinetic study of gamma-glutamyltransferase (GGT)-mediated S-nitrosoglutathione catabolism. Arch. Biochem. Biophys. 2009, 481, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Meister, A. The Liver: Biology and Pathobiology, 2nd ed.; Raven Press: New York, NY, USA, 1988. [Google Scholar]
- Obrador, E.; Carretero, J.; Ortega, A.; Medina, I.; Rodilla, V.; Pellicer, J.A.; Estrela, J.M. Gamma-Glutamyl transpeptidase overexpression increases metastatic growth of B16 melanoma cells in the mouse liver. Hepatology 2002, 35, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Giommarelli, C.; Corti, A.; Supino, R.; Favini, E.; Paolicchi, A.; Pompella, A.; Zunino, F. Cellular response to oxidative stress and ascorbic acid in melanoma cells overexpressing gamma-glutamyltransferase. Eur. J. Cancer 2008, 44, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Franzini, M.; Corti, A.; Lorenzini, E.; Paolicchi, A.; Pompella, A.; De Cesare, M.; Perego, P.; Gatti, L.; Leone, R.; Apostoli, P.; et al. Modulation of cell growth and cisplatin sensitivity by membrane gamma-glutamyltransferase in melanoma cells. Eur. J. Cancer 2006, 42, 2623–2630. [Google Scholar] [CrossRef] [PubMed]
- Mungrue, I.N.; Pagnon, J.; Kohannim, O.; Gargalovic, P.S.; Lusis, A.J. CHAC1/MGC4504 is a novel proapoptotic component of the unfolded protein response, downstream of the ATF4-ATF3-CHOP cascade. J. Immunol. 2009, 182, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Tikoo, S.; Maity, S.; Sengupta, S.; Kaur, A.; Bachhawat, A.K. Mammalian proapoptotic factor ChaC1 and its homologues function as γ-glutamyl cyclotransferases acting specifically on glutathione. EMBO Rep. 2012, 13, 1095–1101. [Google Scholar] [CrossRef]
- Reuter, S.; Schnekenburger, M.; Cristofanon, S.; Buck, I.; Teiten, M.H.; Daubeuf, S.; Serge, E.; Dicato, M.; Aggarwal, B.B.; Visvikis, A.; et al. Tumor necrosis factor alpha induces gamma-glutamyltransferase expression via nuclear factor-kappaB in cooperation with Sp1. Biochem. Pharmacol. 2009, 77, 397–411. [Google Scholar] [CrossRef]
- Dominici, S.; Visvikis, A.; Pieri, L.; Paolicchi, A.; Valentini, M.A.; Comporti, M.; Pompella, A. Redox modulation of NF-kappaB nuclear translocation and DNA binding in metastatic melanoma. The role of endogenous and gamma-glutamyl transferase-dependent oxidative stress. Tumori 2003, 89, 426–433. [Google Scholar] [CrossRef]
- Kabel, A.M.; Atef, A.; Estfanous, R.S. Ameliorative potential of sitagliptin and/or resveratrol on experimentally-induced clear cell renal cell carcinoma. Biomed. Pharmacother. 2018, 97, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Huseby, N.E.; Ravuri, C.; Moens, U. The proteasome inhibitor lactacystin enhances GSH synthesis capacity by increased expression of antioxidant components in an Nrf2-independent, but p38 MAPK-dependent manner in rat colorectal carcinoma cells. Free Radic. Res. 2016, 50, 1–13. [Google Scholar] [CrossRef]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NFkappaB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef]
- Bansal, A.; Sanchez, D.J.; Nimgaonkar, V.; Sanchez, D.; Riscal, R.; Skuli, N.; Simon, M.C. Gamma-glutamyltransferase 1 promotes clear cell renal cell carcinoma initiation and progression. Mol. Cancer Res. 2019, 17, 1881–1892. [Google Scholar] [CrossRef] [PubMed]
- Frierson, H.F.; Theodorescu, D.; Mills, S.E.; Hanigan, M.H. Gamma-Glutamyl transpeptidase in normal and neoplastic prostate glands. Mod. Pathol. 1997, 10, 1–6. [Google Scholar]
- Tsuda, H.; Inoue, T.; Asamoto, M.; Fukushima, S.; Ito, N.; Okamura, T.; Ohtaguro, K.; Washida, H.; Satoh, K.; Amelizad, Z.; et al. Comparison of enzyme phenotypes in human bladder tumours and experimentally induced hyperplastic and neoplastic lesions of the rat urinary bladder. A combined histochemical and immunohistochemical approach. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1989, 56, 307–316. [Google Scholar] [CrossRef]
- Hanigan, M.H.; Frierson, H.F.; Swanson, P.E.; De Young, B.R. Altered expression of gamma-glutamyl transpeptidase in human tumors. Hum. Pathol. 1999, 30, 300–305. [Google Scholar] [CrossRef]
- Ripple, M.O.; Pickhardt, P.A.; Wilding, G. Alteration in gamma-glutamyl transpeptidase activity and messenger RNA of human prostate carcinoma cells by androgen. Cancer Res. 1997, 57, 2428–2433. [Google Scholar]
- Lan, Z.J.; Palladino, M.A.; Rudolph, D.B.; Labus, J.C.; Hinton, B.T. Identification, expression, and regulation of the transcriptional factor polyomavirus enhancer activator 3, and its putative role in regulating the expression of gamma-glutamyl transpeptidase mRNA-IV in the rat epididymis. Biol. Reprod. 1997, 57, 186–193. [Google Scholar] [CrossRef][Green Version]
- Priolo, C.; Khabibullin, D.; Reznik, E.; Filippakis, H.; Ogórek, B.; Kavanagh, T.R.; Nijmeh, J.; Herbert, Z.T.; Asara, J.M.; Kwiatkowski, D.J.; et al. Impairment of gamma-glutamyl transferase 1 activity in the metabolic pathogenesis of chromophobe renal cell carcinoma. Proc. Natl. Acad. Sci. USA 2018, 115, E6274–E6282. [Google Scholar] [CrossRef] [PubMed]
- Tate, S.S.; Meister, A. Affinity labeling of gamma-glutamyl transpeptidase and location of the gamma-glutamyl binding site on the light subunit. Proc. Natl. Acad. Sci. USA 1977, 74, 931–935. [Google Scholar] [CrossRef]
- Fischer, P.H.; Pamukcu, R.; Bittner, G.; Willson, J.K. Enhancement of the sensitivity of human colon cancer cells to growth inhibition by acivicin achieved through inhibition of nucleic acid precursor salvage by dipyridamole. Cancer Res. 1984, 44, 3355–3359. [Google Scholar] [PubMed]
- Mena, S.; Benlloch, M.; Ortega, A.; Carretero, J.; Obrador, E.; Asensi, M.; Petschen, I.; Brown, B.D.; Estrela, J.M. Bcl-2 and glutathione depletion sensitizes B16 melanoma to combination therapy and eliminates metastatic disease. Clin. Cancer Res. 2007, 13, 2658–2666. [Google Scholar] [CrossRef]
- McGovren, J.P.; Pratt, E.A.; Belt, R.J.; Taylor, S.A.; Benjamin, R.S.; Ardalan, B.; Ohnuma, T. Pharmacokinetic and biochemical studies on acivicin in phase I clinical trials. Cancer Res. 1985, 45, 4460–4463. [Google Scholar] [PubMed]
- Willson, J.K.; Fischer, P.H.; Tutsch, K.; Alberti, D.; Simon, K.; Hamilton, R.D.; Bruggink, J.; Koeller, J.M.; Tormey, D.C.; Earhart, R.H.; et al. Phase I clinical trial of a combination of dipyridamole and acivicin based upon inhibition of nucleoside salvage. Cancer Res. 1988, 48, 5585–5590. [Google Scholar]
- Earhart, R.H.; Khandekar, J.D.; Faraggi, D.; Schinella, R.A.; Davis, T.E. Phase II trial of continuous drug infusions in advanced ovarian carcinoma: Acivicin versus vinblastine. Investig. New Drugs. 1989, 7, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Falkson, G.; Cnaan, A.; Simson, I.W.; Dayal, Y.; Falkson, H.; Smith, T.J.; Haller, D.G. A randomized phase II study of acivicin and 4’deoxydoxorubicin in patients with hepatocellular carcinoma in an Eastern Cooperative Oncology Group study. Am. J. Clin. Oncol. 1990, 13, 510–515. [Google Scholar] [CrossRef]
- Taylor, S.A.; Crowley, J.; Pollock, T.W.; Eyre, H.J.; Jaeckle, C.; Hynes, H.E.; Stephens, R.L. Objective antitumor activity of acivicin in patients with recurrent CNS malignancies: A Southwest Oncology Group trial. J. Clin. Oncol. 1991, 9, 1476–1479. [Google Scholar] [CrossRef] [PubMed]
- Bonomi, P.; Finkelstein, D.; Chang, A. Phase II trial of acivicin versus etoposide-cisplatin in non-small cell lung cancer. An Eastern Cooperative Oncology Group study. Am. J. Clin. Oncol. 1994, 17, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Baruchel, S.; Bernstein, M.; Whitehead, V.M.; Devine, S.; Bell, B.; Dubowy, R.; Grier, H.; Kretschmar, C.; Langevin, A.-M.; Vietti, T. A phase I study of acivicin in refractory pediatric solid tumors. A Pediatric Oncology Group study. Investig. New Drugs. 1995, 13, 211–216. [Google Scholar] [CrossRef]
- Hidalgo, M.; Rodriguez, G.; Kuhn, J.G.; Brown, T.; Weiss, G.; MacGovren, J.P.; Von Hoff, D.D.; Rowinsky, E.K. A Phase I and pharmacological study of the glutamine antagonist acivicin with the amino acid solution aminosyn in patients with advanced solid malignancies. Clin. Cancer Res. 1998, 4, 2763–2770. [Google Scholar]
- Kreuzer, J.; Bach, N.C.; Forler, D.; Sieber, S.A. Target discovery of acivicin in cancer cells elucidates its mechanism of growth inhibition†Electronic supplementary information (ESI) available: Synthesis, cloning, protein expression, purification and biochemical assays. Chem. Sci. 2014, 6, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Tate, S.S.; Meister, A. Serine-borate complex as a transition-state inhibitor of gamma-glutamyl transpeptidase. Proc. Natl. Acad. Sci. USA 1978, 75, 4806–4809. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Hiratake, J.; Kamiyama, A.; Sakata, K. Design, synthesis, and evaluation of gamma-phosphono diester analogues of glutamate as highly potent inhibitors and active site probes of gamma-glutamyl transpeptidase. Biochemistry 2007, 46, 1432–1447. [Google Scholar] [CrossRef] [PubMed]
- Lherbet, C.; Keillor, J.W. Probing the stereochemistry of the active site of gamma-glutamyl transpeptidase using sulfur derivatives of l-glutamic acid. Org. Biomol. Chem. 2004, 2, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Terzyan, S.S.; Cook, P.F.; Heroux, A.; Hanigan, M.H. Structure of 6-diazo-5-oxo-norleucine-bound human gamma-glutamyl transpeptidase 1, a novel mechanism of inactivation. Protein Sci. 2017, 26, 1196–1205. [Google Scholar] [CrossRef]
- King, J.B.; West, M.B.; Cook, P.F.; Hanigan, M.H. A novel, species-specific class of uncompetitive inhibitors of gamma-glutamyl transpeptidase. J. Biol. Chem. 2009, 284, 9059–9065. [Google Scholar] [CrossRef]
- Wickham, S.; Regan, N.; West, M.B.; Thai, J.; Cook, P.F.; Terzyan, S.S.; Kai Li, P.; Hanigan, M.H. Inhibition of human γ-glutamyl transpeptidase: Development of more potent, physiologically relevant, uncompetitive inhibitors. Biochem. J. 2013, 450, 547–557. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Russo, G.L.; Russo, M.; Castellano, I.; Napolitano, A.; Palumbo, A. Ovothiol isolated from sea urchin oocytes induces autophagy in the Hep-G2 cell line. Mar. Drugs 2014, 12, 4069–4085. [Google Scholar] [CrossRef]
- Milito, A.; Brancaccio, M.; Lisurek, M.; Masullo, M.; Palumbo, A.; Castellano, I. Probing the interactions of sulfur-containing histidine compounds with human gamma-glutamyl transpeptidase. Mar. Drugs 2019, 17, 650. [Google Scholar] [CrossRef]
- Brancaccio, M.; Russo, M.; Masullo, M.; Palumbo, A.; Russo, G.L.; Castellano, I. Sulfur-containing histidine compounds inhibit γ-glutamyl transpeptidase activity in human cancer cells. J. Biol. Chem. 2019, 294, 14603–14614. [Google Scholar] [CrossRef]
- Selinger, M.J.; Matloff, D.S.; Kaplan, M.M. Gamma-Glutamyl transpeptidase activity in liver disease: Serum elevation is independent of hepatic GGTP activity. Clin. Chim. Acta 1982, 125, 283–290. [Google Scholar] [CrossRef]
- Franzini, M.; Bramanti, E.; Ottaviano, V.; Ghiri, E.; Scatena, F.; Barsacchi, R.; Pompella, A.; Donato, L.; Emdin, M.; Paolicchi, A. A high performance gel filtration chromatography method for gamma-glutamyltransferase fraction analysis. Anal. Biochem. 2008, 374, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Franzini, M.; Bramanti, E.; Ottaviano, V.; Ghiri, E.; Scatena, F.; Barsacchi, R.; Pompella, A.; Donato, L.; Emdin, M.; Paolicchi, A. Cultured human cells release soluble gamma-glutamyltransferase complexes corresponding to the plasma b-GGT. Biomarkers 2009, 14, 486–492. [Google Scholar] [CrossRef]
- Wang, Q.; Shu, X.; Dong, Y.; Zhou, J.; Teng, R.; Shen, J.; Chen, Y.; Dong, M.; Zhang, W.; Huang, Y.; et al. Tumor and serum gamma-glutamyl transpeptidase, new prognostic and molecular interpretation of an old biomarker in gastric cancer. Oncotarget 2017, 8, 36171–36184. [Google Scholar] [CrossRef] [PubMed]
- Kazemi-Shirazi, L.; Endler, G.; Winkler, S.; Schickbauer, T.; Wagner, O.; Marsik, C. Gamma glutamyltransferase and long-term survival: Is it just the liver? Clin. Chem. 2007, 53, 940–946. [Google Scholar] [CrossRef]
- Van Hemelrijck, M.; Jassem, W.; Walldius, G.; Fentiman, I.S.; Hammar, N.; Lambe, M.; Garmo, H.; Jungner, I.; Holmberg, L. Gamma-glutamyltransferase and risk of cancer in a cohort of 545,460 persons - the Swedish AMORIS study. Eur J. Cancer. 2011, 47, 2033–2041. [Google Scholar] [CrossRef]
- Bosco, C.; Garmo, H.; Hammar, N.; Walldius, G.; Jungner, I.; Malmström, H.; Holmberg, L.; Van Hemelrijck, M. Glucose, lipids and gamma-glutamyl transferase measured before prostate cancer diagnosis and secondly diagnosed primary tumours: A prospective study in the Swedish AMORIS cohort. BMC Cancer 2018, 18, 205. [Google Scholar] [CrossRef] [PubMed]
- Kunutsor, S.K.; Laukkanen, J.A. Gamma-glutamyltransferase and risk of prostate cancer: Findings from the KIHD prospective cohort study. Int. J. Cancer 2017, 140, 818–824. [Google Scholar] [CrossRef]
- Kawakami, K.; Fujita, Y.; Matsuda, Y.; Arai, T.; Horie, K.; Kameyama, K.; Kato, T.; Masunaga, K.; Kasuya, Y.; Tanaka, M.; et al. Gamma-glutamyltransferase activity in exosomes as a potential marker for prostate cancer. BMC Cancer 2017, 17, 316. [Google Scholar] [CrossRef]
- Simic, T.; Dragicevic, D.; Savic-Radojevic, A.; Cimbaljevic, S.; Tulic, C.; Mimic-Oka, J. Serum gamma glutamyl-transferase is a sensitive but unspecific marker of metastatic renal cell carcinoma. Int J. Urol. 2007, 14, 289–293. [Google Scholar] [CrossRef]
- Polterauer, S.; Hofstetter, G.; Grimm, C.; Rahhal, J.; Mailath-Pokorny, M.; Kohl, M.; Concin, N.; Tempfer, C.; Mart, C.; Reinthaller, A. Relevance of gamma-glutamyltransferase—A marker for apoptotic balance--in predicting tumor stage and prognosis in cervical cancer. Gynecol. Oncol. 2011, 122, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Seebacher, V.; Polterauer, S.; Grimm, C.; Rahhal, J.; Hofstetter, G.; Bauer, E.M.; Husslein, H.; Leipold, H.; Marth, C.; Reinthaller, A.; et al. Prognostic significance of gamma-glutamyltransferase in patients with endometrial cancer: A multi-centre trial. Br. J. Cancer 2012, 106, 1551–1555. [Google Scholar] [CrossRef]
- Edlinger, M.; Concin, N.; Concin, H.; Nagel, G.; Ulmer, H.; Gobel, G. Lifestyle-related biomarkers and endometrial cancer survival: Elevated gamma-glutamyltransferase as an important risk factor. Cancer Epidemiol. 2013, 37, 156–161. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, A.J.; Wu, D.B.; Shen, Z.; Chen, G.; Shi, Y.Y.; Wu, H.; Wang, J. Prognostic significance of the pretreatment serum gamma-glutamyltransferase levels in Chinese patients with non-metastatic cervical cancer. Oncotarget 2017, 8, 115701–115708. [Google Scholar] [CrossRef] [PubMed]
- Schwameis, R.; Grimm, C.; Brodowicz, T.; Petru, E.; Hefler-Frischmuth, K.; Staudigl, C.; Reinthaller, A.; Heinze, G.; Polterauer, S.; Polterauer, M. Gamma-glutamyltransferase as novel biomarker in patients with uterine leiomyosarcoma. Sci. Rep. 2016, 6, 33757. [Google Scholar] [CrossRef] [PubMed]
- He, W.Z.; Guo, G.F.; Yin, C.X.; Jiang, C.; Wang, F.; Qiu, H.J.; Chen, X.-X.; Rong, R.-M.; Zhang, B.; Xia, L.-P. Gamma-glutamyl transpeptidase level is a novel adverse prognostic indicator in human metastatic colorectal cancer. Colorectal Dis. 2013, 15, e443–e452. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, S.; Yang, H.; Luo, K.; Wen, J.; Hu, Y.; Hu, R.; Huang, Q.; Chen, J.; Fu, J. Prognostic significance of gamma-glutamyltransferase in patients with resectable esophageal squamous cell carcinoma. Dis. Esophagus. 2015, 28, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, X.P.; Li, X.H.; Chen, H.; Zheng, X.; Lin, J.H.; Kang, T.; Zhang, L.; Chen, P.S. Prognostic value of pretreatment serum alanine aminotransferase/aspartate aminotransferase (ALT/AST) ratio and gamma glutamyltransferase (GGT) in patients with esophageal squamous cell carcinoma. BMC Cancer 2017, 17, 544. [Google Scholar] [CrossRef]
- Staudigl, C.; Concin, N.; Grimm, C.; Pfeiler, G.; Nehoda, R.; Singer, C.F.; Polterauer, S. Prognostic relevance of pretherapeutic gamma-glutamyltransferase in patients with primary metastatic breast cancer. PLoS ONE 2015, 10, e0125317. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yin, W.; Wu, Z.; Wang, Y.; Lu, J. The predictive value of pre-therapeutic serum gamma-glutamyl transferase in efficacy and adverse reactions to neoadjuvant chemotherapy among breast cancer patients. J. Breast Cancer 2020, 23, 509–520. [Google Scholar] [CrossRef]
- Xu, X.S.; Miao, R.C.; Zhang, L.Q.; Wang, R.T.; Qu, K.; Pang, Q.; Liu, C. Model based on alkaline phosphatase and gamma-glutamyltransferase for gallbladder cancer prognosis. Asian Pac. J. Cancer Prev. 2015, 16, 6255–6259. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wen, Y.F.; Yang, X.Z.; Zeng, L.S.; Peng, H.H.; Huang, W.J.; Cai, L.M.; Zhou, T.-C.; Lin, X.D. Prognostic impact of pretherapeutic gamma-glutamyltransferase on patients with nasopharyngeal carcinoma. PLoS ONE 2017, 12, e0172345. [Google Scholar] [CrossRef]
- Luo, M.; Sun, W.; Wu, C.; Zhang, L.; Liu, D.; Li, W.; Mei, Q.; Hu, G. High pretreatment serum gamma-glutamyl transpeptidase predicts an inferior outcome in nasopharyngeal carcinoma. Oncotarget 2017, 8, 67651–67662. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, S.; He, X.; Liu, Y.; Ding, X.; Jiang, H.; Tan, Y.; Lu, H. Prognostic significance of serum uric acid and gamma-glutamyltransferase in patients with advanced gastric cancer. Dis. Markers 2019, 2019, 1415421. [Google Scholar] [CrossRef]
- Bozkaya, Y.; Yazıcı, O. Prognostic significance of gamma-glutamyl transferase in patients with metastatic non-small cell lung cancer. Expert Rev. Mol. Diagn. 2019, 19, 267–272. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, H.; Lu, J.; Li, D.; Xu, C.; Risch, H.A. Serum gamma-glutamyltransferase and the overall survival of metastatic pancreatic cancer. BMC Cancer 2019, 19, 1020. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Knoll, T.; Omar, M.I.; Maclennan, S.; Hernandez, V.; Canfield, S.; Yuan, Y.; Bruins, M.; Marconi, L.; Van Poppel, H.; N’Dow, J.; et al. Key steps in conducting systematic reviews for underpinning clinical practice guidelines: Methodology of the European Association of Urology. Eur. Urol. 2018, 73, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Horie, K.; Kawakami, K.; Fujita, Y.; Matsuda, Y.; Arai, T.; Suzui, N.; Miyazaki, T.; Koie, T.; Mizutani, K.; Ito, M. Serum exosomal gamma-glutamyltransferase activity increased in patients with renal cell carcinoma with advanced clinicopathological features. Oncology 2020, 98, 734–742. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author (Year) | Country | Participants (P) | Intervention (I) | Comparator (C), Cut-off of Serum GGT (U/L) | Outcomes (O) | Reference |
|---|---|---|---|---|---|---|
| Ramankulov, A. (2007) | Germany | 80 patients with non-metastatic or metastatic RCC: pN0M0, n = 32 (40%); pN1M0, n = 11 (14%); M1, and n = 37 (46%). | Surgery | 40.6 | Serum GGT increased in patients with non-bone metastases (median 25.3 U/L) in comparison to healthy controls (median 14.3 U/L). Elevated serum GGT was not a predictive factor of survival (p = 0.213) | [7] |
| Hofbauer, S.L. (2014) | Austria | 921 patients with non-metastatic or metastatic RCC: pN+, n = 29 (3%); M1, n = 118 (13%). | Surgery | 17.5, 34.5, and 181.5 | Serum GGT was an independent prognostic factor for CSS. Serum GGT increased with advanced pathological stages, Fuhrman grade, and tumor necrosis. Adding GGT to a base model increased discrimination from 0.9% to 1.8%. | [8] |
| Dalpiaz, O. (2015) | Austria | 700 patients with non-metastatic RCC | Surgery | 40 | Elevated serum GGT was associated with shorter MFS (p = 0.025) but not with OS (p = 0.108) or CSS (p = 0.242). Serum GGT failed to reach independent predictor status for MFS prediction. | [9] |
| Luo, C. (2017) | China | 179 patients with non-metastatic RCC with venous tumor thrombus | Surgery | 37.5 | Elevated serum GGT was significantly associated with an advanced tumor stage, was a significant predictor of shorter CSS (HR 2.99, p < 0.001) and RFS (HR 2.59, p < 0.001) on univariable analysis, and was an independent prognostic biomarker on multivariable analysis. | [10] |
| Takemura, K. (2019) | Japan | 125 patients with advanced UC. Inoperable cT4, n = 42 (34%); cN+, n = 92 (74%); cM+, n = 42 (34%) | Systemic chemotherapy, n = 93 (74%); surgery, n = 33 (26%); radiation, n = 6 (5%) | 60 | Elevated serum GGT at the diagnosis of advanced UC was associated with shorter OS on both univariable (HR 3.25, p < 0.001) and multivariable (HR 2.97, p < 0.001) analysis irrespective of liver metastases and hepatic comorbidities. | [14] |
| Takemura, K. (2019) | Japan | 60 patients with metastatic PC. Regional LN, n = 23 (46%); non-regional LN, n = 3 (6%); bone, n = 33 (66%); other sites, n = 12 (24%) | Enzalutamide | 40 | Elevated serum GGT was associated with shorter OS (HR 2.68, p = 0.018) and PSA–PFS (HR 3.23, p = 0.002). PSA changes after enzalutamide therapy were less evident in the elevated serum GGT group than in the normal serum GGT group (−45% versus −90%, p = 0.049). | [13] |
| Takemura, K. (2020) | Japan | 146 patients with metastatic RCC. cN+, n = 44 (30%); cM+, n = 143 (98%) | TKI | 1.5 × ULN | Elevated serum GGT was associated with shorter OS (HR 3.89, p < 0.001). Preoperative serum GGT was 29, 48, and 109 U/L in patients with RCC specimens showing negative to weak, moderate, and strong GGT expression, respectively (p = 0.004). | [11] |
| Ishiyama, Y. (2020) | Japan | 69 patients with metastatic RCC. A single metastasis, n = 24 (35%); multiple metastases, n = 45 (65%) | Nivolumab | 49 | Elevated baseline serum GGT was associated with shorter OS (p = 0.005) but not with PFS (p = 0.092). Serum GGT increase after nivolumab therapy was associated with shorter OS (p = 0.023) and PFS (p = 0.008). | [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takemura, K.; Board, P.G.; Koga, F. A Systematic Review of Serum γ-Glutamyltransferase as a Prognostic Biomarker in Patients with Genitourinary Cancer. Antioxidants 2021, 10, 549. https://doi.org/10.3390/antiox10040549
Takemura K, Board PG, Koga F. A Systematic Review of Serum γ-Glutamyltransferase as a Prognostic Biomarker in Patients with Genitourinary Cancer. Antioxidants. 2021; 10(4):549. https://doi.org/10.3390/antiox10040549
Chicago/Turabian StyleTakemura, Kosuke, Philip G. Board, and Fumitaka Koga. 2021. "A Systematic Review of Serum γ-Glutamyltransferase as a Prognostic Biomarker in Patients with Genitourinary Cancer" Antioxidants 10, no. 4: 549. https://doi.org/10.3390/antiox10040549
APA StyleTakemura, K., Board, P. G., & Koga, F. (2021). A Systematic Review of Serum γ-Glutamyltransferase as a Prognostic Biomarker in Patients with Genitourinary Cancer. Antioxidants, 10(4), 549. https://doi.org/10.3390/antiox10040549