
Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models



Abstract
1. Introduction
2. Taurine Sources
3. Taurine Essentiality and Requirement
4. Taurine Absorption and Metabolism
- the renal adaptive regulation of TauT depends on taurine availability and controls the body pool of Tau;
- renal Tau accumulation is affected by the ionic environment, electrochemical charge and pH;
- Tau transfer across the cell membrane is dependent on TauT and controlled by PKC phosphorylation post-translationally;
- p53 suppresses TauT expression;
- over-expression of TauT is shown to have protective effects on renal cells against oxidative stress-induced nephrotoxicity.
- high TauT expression was shown in the liver, kidney, brain, retinas and placenta of mammals [42].
5. Biological Roles of Tau
6. Antioxidant Systems of the Body
- decreasing the localised oxygen concentration;
- reducing the activity of prooxidant enzymes;
- improving the efficiency of ETC in the mitochondria and decreasing electron leakage;
- inducing various transcription factors (e.g., Nrf2, NF-κB, etc.) with increased antioxidant response element-related synthesis of various antioxidants;
- binding metal ions (metal-binding proteins) and metal chelating;
- decomposing peroxides;
- chain breaking by scavenging intermediate radicals, including peroxyl and alkoxyl radicals;
- binding reactive products of peroxidation, such as MDA, 4-hydroxynoinenal, etc.
- repairing and removing damaged molecules;
- maintaining optimal redox status;
- maintaining redox signalling and vitagene activation with synthesis and increased expression of protective molecules;
- providing antioxidant recycling mechanisms, including vitamin E recycling;
- inducing protein glutathionylation as a way to prevent its irreversible oxidation;
- activating apoptosis/ferroptosis with the removal of terminally damaged cells and restricting mutagenesis.
7. Antioxidant Properties of Tau
- direct free radical scavenging;
- maintenance of the integrity of electron-transport chain of mitochondria in stress conditions;
- stabilizing biological membranes;
- inhibiting ROS-generating specific enzymes;
- modulating transcription factors responsible for upregulation and increased synthesis of AO enzymes;
- up-regulating vitagenes (SOD, HSP, thioredoxin, TR and sirtuins), leading to improved adaptation to stress.
7.1. Direct Free Radical Scavenging
7.2. Protective Effects of Tau on Mitochondria
- the electron-transport chain (ETC), including complexes I, II and III;
- pyruvate dehydrogenase and glycerol-3-phosphate dehydrogenase;
- α-ketoglutarate dehydrogenase and aconitase;
- cytochrome b5 reductase;
- dihydroorotate dehydrogenase;
- the monoamine oxidases A and B.
7.3. Membrane-Stabilizing Activity of Tau
7.4. Inhibition of Free-Radical Producing Enzymes by Tau
7.4.1. Xanthine Oxidase
7.4.2. NADPH Oxidase
7.5. Antioxidant Protective Properties of Tau in Prevention of Toxic Effects of Various Chemicals
7.5.1. Arsenic
7.5.2. Carbon Tetrachloride
7.5.3. Thioacetamide
7.5.4. Cisplatin
- Tau is able to abrogate decreased activities of antioxidant enzymes (SOD, GPx, GR, and catalase) and non-enzymatic antioxidants (GSH) in CP-treated animals and maintain optimal redox status in stress conditions;
- Oxidative stress imposed by CP treatment was associated with DNA oxidation and apoptosis, and Tau was shown to be protective;
- CP treatment affected Tau metabolism and transport, while the overexpression of TauT in CP-treated animals showed protective effects;
- Tau can ameliorate increased NF-κB expression caused by CP-treatment and prevent excessive inflammation.
7.5.5. Doxorubicin
- DOX compromised the antioxidant defence system by decreasing antioxidant enzyme (SOD, CAT, GST, GPx and GR) activities and non-enzymatic (GSH) antioxidant concentrations, and Tau treatment was able to ameliorate these detrimental changes;
- the mechanisms of Tau-induced cytoprotection were suggested to be also associated with the activation of specific survival signals and PI3-K/Akt, as well as the suppression of p53, JNK, p38 and NF-κB.
7.5.6. Streptozotocin
- antioxidant enzyme activities (SOD, GPx, GR, GST, Catalase) in animal tissues (brain, testes, kidney, liver, aorta, heart, plasma and RBC) are shown to be compromised by STZ treatment;
- Tau supplementation of STZ-treated animals in most cases can prevent oxidative stress through the restoration of antioxidant enzyme activities and redox balance in cells/tissues, leading to a reduction in lipid and DNA oxidation;
- Tau also showed anti-inflammatory action in STZ-treated animals, probably due to decreasing NF-κB expression in various tissues.
7.5.7. Fluoride
7.5.8. Other Toxicants
- SOD—main adaptive enzyme of the first level of antioxidant defence, participating in the detoxification of superoxide radicals. It is regulated at the vitagene level [359];
- GSH—an endogenous antioxidant tripeptide considered as one of the most important cellular antioxidants. GSH can directly detoxify ROS/RNS or contribute indirectly to AO defences as a cofactor for various antioxidant enzymes, such as GPx and GST, participating in maintaining redox balance in various tissues of the body [1];
- GR—an essential component of the glutathione system responsible for recycling oxidised glutathione back into the reduced form, involved in redox balance regulation [1];
- GSTs—a superfamily of multifunctional isoenzymes participating in the cellular detoxification of various endogenous and exogenous toxic compounds, including insecticides, drugs, and organic hydroperoxides. GSTs play a crucial role in protection against various types of cellular oxidative damage [363];
- CAT—an important enzyme of the first level of the antioxidant defence network, providing detoxification of H2O2 produced by SOD [364].
8. Protective Effects of Tau in Stress Conditions of Poultry Production
9. Conclusions
- First, in some tissues (e.g., heart and eye), Tau can directly scavenge free radicals.
- Secondly, Tau participates in maintaining the integrity of electron-transport chain of mitochondria, the main sources of ROS in biological systems.
- Thirdly, Tau is shown to inhibit the activities of ROS-producing enzymes, including XO and NADP oxidase.
- Fourthly, by interacting with transcription factors (e.g., Nrf2 and NF-κB) and inducing AO enzymes, Tau can maintain the optimal redox status of the cell.
- Finally, by activating vitagenes, including SOD, HSP, thioredoxin, sirtuins, etc., Tau can provide additional protection in stress conditions and help in the creation of an effective adaptative response to stresses.
- Tau plays a vital role in the AO defence network. However, the direct antioxidant effects of Tau in biological systems are limited.
- The stabilizing effects of Tau on mitochondria under various stress conditions, including commercial animal and poultry production, deserve more attention.
- A range of toxicological models clearly showed the protective antioxidant-related effects of Tau.
- Until now, major Tau applications have been related to human health, including the prevention/treatment of various diseases or decreasing the detrimental effects of various essential drugs used in human medicine.
- Furthermore, there are a range of publications confirming the protective effects of Tau (alone or in combination with other antioxidants) in various stress conditions of commercial meat and egg production.
- Anti-inflammatory and immunomodulatory properties of Tau are of great importance for veterinary medicine.
- Indirect antioxidant activities of Tau due to the modulation of transcription factors and vitagenes leading to the upregulation of the antioxidant defence network are likely to be major molecular mechanisms of Tau’s antioxidant and anti-inflammatory activities, and they deserve more attention and further investigations.
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKT | protein kinase B |
| ALP | alkaline phosphatase |
| ALT | alanine aminotransferase |
| AST | aspartate aminotrasferase |
| AO | antioxidant |
| ALX | alloxan |
| BAD | BCL2 associated agonist of cell death |
| BCL2 | B-cell lymphoma-2 |
| CAT | catalase |
| COX2 | cyclooxygenase-2 |
| CP | cisplatin |
| CTSB | cathepsin B |
| CYP2E1 | cytochrome P450 family 2 subfamily E member 1 |
| 8-OHdG | 8 hydroxy 2 deoxyguanosine |
| DOX | doxorubicin |
| DPPH | 1,1-diphenyl-2-picrylhydrazyl |
| EIF2α | eukaryotic initiation factor 2 |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| ETC | electron-transport chain |
| FOXP3 | Forkhead box protein P3 |
| GGT | gamma-glutamyl transferase |
| GR | glutathione reductase |
| GRP78 | 78-kDa glucose-regulated protein |
| Grx | glutaredoxin |
| GSH | reduced glutathione |
| GPx | glutathione peroxidase |
| GST | glutathione S-transferase |
| HO-1 | heme oxygenase 1 |
| HSP | heat-shock protein |
| IκB | inhibitor of κB |
| iNOS | inducible nitric oxide synthases |
| IKK | IκB kinase |
| IFN-α | interferon alpha |
| IL | interleukin |
| JNK | c-Jun N-terminal kinase |
| Keap1 | Kelch-like erythroid cell-derived protein with CNC homology (ECH)-associated protein 1 |
| LDH | lactate dehydrogenase |
| LPO | lipid peroxidation |
| LPS | lipopolysaccharide |
| MAPK | mitogen-activated protein kinase |
| MDA | malondialdehyde |
| MTO1 | a mitochondrial protein |
| NF-κB | nuclear factor-κB |
| NLR | NOD-like receptor |
| NLRP3 | NLR family pyrin domain containing 3 |
| Nox | NADPH oxidase |
| NQO1 | NAD(P)H:quinone dehydrogenase 1 |
| Nrf2 | nuclear factor-erythroid-2 (NF-E2) and related factor 2 |
| PAT1 | DNA topoisomerase 2-associated protein |
| PI3-K | phosphoinositide 3-kinase |
| PPAR | peroxisome proliferator-activated receptor |
| PUFAs | polyunsaturated fatty acids |
| p53 | tumour protein p53 |
| RNS | reactive nitrogen species |
| RONS | reactive oxygen and nitrogen species |
| ROS | reactive oxygen species |
| SIL | silymarin |
| SOD | superoxide dismutase |
| STZ | streptozotocin |
| TAA | thioacetamide |
| Tau | taurine |
| TauCl | taurine chloramine |
| TauT | taurine transporter |
| TCA | tricarboxylic acid |
| TGF-β | transforming growth factor beta |
| TNF-α | tumour necrosis factor |
| TR | thioredoxin reductase |
| Trx | thioredoxin |
| UPR | unfolded protein response |
| XO | xanthine oxidase |
References
- Surai, P.F. Vitagenes in Avian Biology and Poultry Health; Wageningen Academic Publishers: Wageningen, The Netherlands, 2020. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Kochish, I.I.; Kidd, M.T. Redox Homeostasis in Poultry: Regulatory Roles of NF-κB. Antioxidants 2021, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Fisinin, V.I. Vitagenes in poultry production. Part 1. Technological and environmental stresses. Worlds Poult. Sci. J. 2016, 72, 721–733. [Google Scholar] [CrossRef]
- Surai, P.F.; Fisinin, V.I. Vitagenes in poultry production. Part 2. Nutritional and internal stresses. Worlds Poult. Sci. J. 2016, 72, 761–772. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Taverne, Y.J.; Merkus, D.; Bogers, A.J.; Halliwell, B.; Duncker, D.J.; Lyons, T.W. Reactive Oxygen Species: Radical Factors in the Evolution of Animal Life: A molecular timescale from Earth’s earliest history to the rise of complex life. Bioessays 2018, 40, 3. [Google Scholar] [CrossRef]
- Surai, P.F. Carnitine Enigma: From Antioxidant Action to Vitagene Regulation. Part 1. Absorption, Metabolism and Antioxidant Activities. J. Veter. Sci. Med. 2015, 3, 14. [Google Scholar] [CrossRef]
- Surai, P.F. Carnitine Enigma: From Antioxidant Action to Vitagene Regulation. Part 2. Transcription Factors and Practical Applications. J. Veter. Sci. Med. 2015, 3, 17. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidant Action of Carnitine: Molecular Mechanisms and Practical Applications. EC Vet. Sci. 2015, 2, 66–84. [Google Scholar]
- Seidel, U.; Huebbe, P.; Rimbach, G. Taurine: A Regulator of Cellular Redox-Homeostasis and Skeletal Muscle Function. Mol. Nutr. Food Res. 2019, 63, e1800569. [Google Scholar] [CrossRef]
- Ribeiro, R.A.; Bonfleur, M.L.; Batista, T.M.; Borck, P.C.; Carneiro, E.M. Regulation of glucose and lipid metabolism by the pancreatic and extra-pancreatic actions of taurine. Amino Acids 2018, 50, 1511–1524. [Google Scholar] [CrossRef]
- Schaffer, S.; Kim, H.W. Effects and Mechanisms of Taurine as a Therapeutic Agent. Biomol. Ther. 2018, 26, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.L.; Gao, Y.; Ma, H.T.; Zheng, L.H.; Dai, B.; Miao, J.F.; Zhang, Y.S. Effects of taurine and housing density on renal function in laying hens. J. Zhejiang Univ. Sci. B 2016, 17, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Dietary taurine supplementation improves breast meat quality in chronic heat-stressed broilers via activating the Nrf2 pathway and protecting mitochondria from oxidative attack. J. Sci. Food Agric. 2019, 99, 1066–1072. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Mi, Y.; Liu, L.; Lv, C.; Zeng, W.; Zhang, C.; Li, J. Taurine regulates mucosal barrier function to alleviate lipopolysaccharide-induced duodenal inflammation in chicken. Amino Acids 2018, 50, 1637–1646. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Kidd, M.T. Taurine in poultry nutrition. Anim. Feed Sci. Technol. 2020, 260, 114339. [Google Scholar]
- Surai, P.F. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef]
- Tiedemann, F.; Gmelin, L. Einige neue Bestandtheile der Galle des Ochsen. Ann. Der Phys. 1827, 85, 326–337. [Google Scholar] [CrossRef]
- Huxtable, R.J. Taurine. Past, present, and future. Adv. Exp. Med. Biol. 1996, 403, 641–650. [Google Scholar]
- Kataoka, H.; Ohnishi, N. Occurrence of taurine in plants. Agric. Biol. Chem. 1986, 50, 1887–1888. [Google Scholar]
- Edgar, S.E.; Kirk, C.A.; Rogers, Q.R.; Morris, J.G. Taurine status in cats is not maintained by dietary cysteinesulfinic acid. J. Nutr. 1998, 128, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Froger, N.; Moutsimilli, L.; Cadetti, L.; Jammoul, F.; Wang, Q.P.; Fan, Y.; Gaucher, D.; Rosolen, S.G.; Neveux, N.; Cynober, L.; et al. Taurine: The comeback of a neutraceutical in the prevention of retinal degenerations. Prog. Retin. Eye Res. 2014, 41, 44–63. [Google Scholar] [CrossRef]
- Lambert, I.H.; Kristensen, D.M.; Holm, J.B.; Mortensen, O.H. Physiological role of taurine—From organism to organelle. Acta Physiol. 2015, 213, 191–212. [Google Scholar] [CrossRef] [PubMed]
- Mizina, T.I.; Dokshina, G.A. Effect of taurine on the functional status of the insular apparatus and adrenal cortex of the rat with experimental diabetes. Probl. Endokrinol. 1987, 33, 63–66. [Google Scholar]
- Pasantes-Morales, H.; Quesada, O.; Morán, J. Taurine: An osmolyte in mammalian tissues. Adv. Exp. Med. Biol. 1998, 442, 209–217. [Google Scholar]
- Schaffer, S.W.; Ito, T.; Azuma, J. Clinical significance of taurine. Amino Acids 2014, 46, 1–5. [Google Scholar] [CrossRef]
- Kadam, S.U.; Prabhasankar, P. Marine foods as functional ingredients in bakery and pasta products. Food Res. Int. 2014, 43, 1975–1980. [Google Scholar] [CrossRef]
- Qvartskhava, N.; Jin, C.; Buschmann, T.; Monhasery, N.; Oenarto, J.; Bidmon, H.; Görg, B.; Häussinger, D. Taurine deficiency induces hyperammonemia and oxidative stress and senescence in mouse liver. Z. Gastroenterol. 2018, 56, A3–A18. [Google Scholar] [CrossRef]
- Inam-U-Llah; Piao, F.; Aadil, R.M.; Suleman, R.; Li, K.; Zhang, M.; Wu, P.; Shahbaz, M.; Ahmed, Z. Ameliorative effects of taurine against diabetes: A review. Amino Acids 2018, 50, 487–502. [Google Scholar] [CrossRef]
- Sarkar, P.; Basak, P.; Ghosh, S.; Kundu, M.; Sil, P.C. Prophylactic role of taurine and its derivatives against diabetes mellitus and its related complications. Food Chem. Toxicol. 2017, 110, 109–121. [Google Scholar] [CrossRef]
- Chen, W.; Guo, J.; Zhang, Y.; Zhang, J. The beneficial effects of taurine in preventing metabolic syndrome. Food Funct. 2016, 7, 1849–1863. [Google Scholar] [CrossRef] [PubMed]
- De Luca, A.; Pierno, S.; Camerino, D.C. Taurine: The appeal of a safe amino acid for skeletal muscle disorders. J. Transl. Med. 2015, 13, 243. [Google Scholar] [CrossRef]
- Murakami, S. Taurine and atherosclerosis. Amino Acids 2014, 46, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Matsuzaki, Y. Taurine and liver diseases: A focus on the heterogeneous protective properties of taurine. Amino Acids 2014, 46, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Menzie, J.; Pan, C.; Prentice, H.; Wu, J.Y. Taurine and central nervous system disorders. Amino Acids 2014, 46, 31–46. [Google Scholar] [CrossRef]
- Sved, D.W.; Godsey, J.L.; Ledyard, S.L.; Mahoney, A.P.; Stetson, P.L.; Ho, S.; Myers, N.R.; Resnis, P.; Renwick, A.G. Absorption, tissue distribution, metabolism and elimination of taurine given orally to rats. Amino Acids 2007, 32, 459–466. [Google Scholar] [CrossRef]
- Ghandforoush-Sattari, M.; Mashayekhi, S.; Krishna, C.V.; Thompson, J.P.; Routledge, P.A. Pharmacokinetics of oral taurine in healthy volunteers. J. Amino Acids 2010, 2010, 346237. [Google Scholar] [CrossRef]
- Nielsen, C.U.; Bjerg, M.; Ulaganathan, N.; Holm, R. Oral and intravenous pharmacokinetics of taurine in sprague-dawley rats: The influence of dose and the possible involvement of the proton-coupled amino acid transporter, PAT1, in oral taurine absorption. Physiol. Rep. 2017, 5, e13467. [Google Scholar] [CrossRef]
- Han, X.; Chesney, R.W. Mechanisms of regulation of taurine transporter activity: A complex interplay of regulatory systems. Adv. Exp. Med. Biol. 2006, 583, 79–90. [Google Scholar]
- Han, X.; Patters, A.B.; Jones, D.P.; Zelikovic, I.; Chesney, R.W. The taurine transporter: Mechanisms of regulation. Acta Physiol. 2006, 187, 61–73. [Google Scholar] [CrossRef]
- Chen, N.H.; Reith, M.E.; Quick, M.W. Synaptic uptake and beyond: The sodium- and chloride-dependent neurotransmitter transporter family SLC6. Pflug. Arch. 2004, 447, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Chesney, R.W. Stress-responsive gene TauT and acute kidney injury. J. Biomed. Sci. 2010, 17 (Suppl. S1), S28. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Howard, A.; Walters, J.R.; Ganapathy, V.; Thwaites, D.T. Taurine uptake across the human intestinal brush-border membrane is via two transporters: H+-coupled PAT1 (SLC36A1) and Na+- and Cl(-)-dependent TauT (SLC6A6). J. Physiol. 2009, 587, 731–744. [Google Scholar] [CrossRef]
- Voaden, M.J.; Oraedu, A.C.I.; Marshall, J.; Lake, N. Taurine in the retina. In The Effects of Tuurine on Excitable Tissues; Schaffer, S.W., Baskin, S.I., Kocsis, J.J., Eds.; Spectrum Publications: New York, NY, USA, 1981; pp. 145–160. [Google Scholar]
- Schuller-Levis, G.B.; Park, E. Taurine and its chloramine: Modulators of immunity. Neurochem. Res. 2004, 29, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Kuzmina, V.V.; Gavrovskaya, L.K.; Ryzhova, O.V. Taurine. Effect on exotrophia and metabolism in mammals and fish. J. Evol. Biochem. Physiol. 2010, 46, 19–27. [Google Scholar] [CrossRef]
- Hansen, S.H.; Andersen, M.L.; Birkedal, H.; Cornett, C.; Wibrand, F. The important role of taurine in oxidative metabolism. Adv. Exp. Med. Biol. 2006, 583, 129–135. [Google Scholar]
- Jacobsen, J.G.; Smith, L.H. Biochemistry and physiology of taurine and taurine derivatives. Physiol. Rev. 1968, 48, 424–511. [Google Scholar] [CrossRef]
- Kocsis, J.J.; Kostos, V.J.; Baskin, S.I. Taurine levels in the heart tissues of various species. In Taurine; Raven Press: New York, NY, USA, 1976; pp. 145–153. [Google Scholar]
- Sturman, J.A.; Hepner, G.W.; Hofmann, A.F.; Thomas, P.J. Metabolism of [35S]taurine in man. J. Nutr. 1975, 105, 1206–1214. [Google Scholar] [CrossRef]
- Grove, R.Q.; Karpowicz, S.J. Reaction of hypotaurine or taurine with superoxide produces the organic peroxysulfonic acid peroxytaurine. Free Radic. Biol. Med. 2017, 108, 575–584. [Google Scholar] [CrossRef]
- Patel, S.N.; Pandya, K.; Clark, G.J.; Parikh, M.C.; Lau-Cam, C.A. Comparison of taurine and pantoyltaurine as antioxidants in vitro and in the central nervous system of diabetic rats. Exp. Toxicol. Pathol. 2016, 68, 103–112. [Google Scholar] [CrossRef]
- Cheong, S.H.; Moon, S.H.; Lee, S.J.; Kim, S.H.; Chang, K.J. Antioxidant and DNA protection effects of taurine by electron spin resonance spectroscopy. Adv. Exp. Med. Biol. 2013, 776, 167–177. [Google Scholar] [PubMed]
- Zhang, Z.; Liu, D.; Yi, B.; Liao, Z.; Tang, L.; Yin, D.; He, M. Taurine supplementation reduces oxidative stress and protects the liver in an iron-overload murine model. Mol. Med. Rep. 2014, 10, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.W.; Azuma, J.; Madura, J.D. Mechanisms underlying taurine-mediated alterations in membrane function. Amino Acids 1995, 8, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Baliou, S.; Adamaki, M.; Ioannou, P.; Pappa, A.; Panayiotidis, M.I.; Spandidos, D.A.; Christodoulou, I.; Kyriakopoulos, A.M.; Zoumpourlis, V. Protective role of taurine against oxidative stress (Review). Mol. Med. Rep. 2021, 24, 605. [Google Scholar] [CrossRef] [PubMed]
- Jong, C.J.; Azuma, J.; Schaffer, S. Mechanism underlying the antioxidant activity of taurine: Prevention of mitochondrial oxidant production. Amino Acids 2012, 42, 2223–2232. [Google Scholar] [CrossRef]
- Hansen, S.H.; Andersen, M.L.; Cornett, C.; Gradinaru, R.; Grunnet, N. A role for taurine in mitochondrial function. J. Biomed. Sci. 2010, 17 (Suppl. S1), S23. [Google Scholar] [CrossRef]
- Jong, C.J.; Sandal, P.; Schaffer, S.W. The Role of Taurine in Mitochondria Health: More Than Just an Antioxidant. Molecules 2021, 26, 4913. [Google Scholar] [CrossRef]
- Homma, K.; Toda, E.; Osada, H.; Nagai, N.; Era, T.; Tsubota, K.; Okano, H.; Ozawa, Y. Taurine rescues mitochondria-related metabolic impairments in the patient-derived induced pluripotent stem cells and epithelial-mesenchymal transition in the retinal pigment epithelium. Redox Biol. 2021, 41, 101921. [Google Scholar] [CrossRef]
- Liu, Y.; Li, F.; Zhang, L.; Wu, J.; Wang, Y.; Yu, H. Taurine alleviates lipopolysaccharide-induced liver injury by anti-inflammation and antioxidants in rats. Mol. Med. Rep. 2017, 16, 6512–6517. [Google Scholar] [CrossRef]
- Belal, S.A.; Kang, D.; Cho, E.S.R.; Park, G.H.; Shim, K.S. Taurine Reduces Heat Stress by Regulating the Expression of Heat Shock Proteins in Broilers Exposed to Chronic Heat. Braz. J. Poult. Sci. 2018, 20, 479–486. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I. Antioxidant systems in poultry biology: Nutritional modulation of vitagenes. Europ. Poult. Sci. 2017, 81, 1612–9199. [Google Scholar]
- Kang, I.S.; Kim, C. Taurine chloramine administered in vivo increases NRF2-regulated antioxidant enzyme expression in murine peritoneal macrophages. Adv. Exp. Med. Biol. 2013, 775, 259–267. [Google Scholar]
- Ciaula, A.; Garruti, G.; Lunardi Baccetto, R.; Molina-Molina, E.; Bonfrate, L.; Wang, D.Q.; Portincasa, P. Bile Acid Physiology. Ann. Hepatol. 2017, 16 (Suppl. S1), s4–s14. [Google Scholar] [CrossRef] [PubMed]
- Kevresan, S.; Kuhajda, K.; Kandrac, J.; Fawcett, J.P.; Mikov, M. Biosynthesis of bile acids in mammalian liver. Eur. J. Drug Metab. Pharm. 2006, 31, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Bouckenooghe, T.; Remacle, C.; Reusens, B. Is taurine a functional nutrient? Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.R.; Barnes, S.; Kwakye, J.B.; Diasio, R.B. Purification and characterization of bile acid-CoA:amino acid N-acyltransferase from human liver. J. Biol. Chem. 1991, 266, 10227–10233. [Google Scholar] [CrossRef]
- Wang, Y.; Matye, D.; Nguyen, N.; Zhang, Y.; Li, T. HNF4α Regulates CSAD to Couple Hepatic Taurine Production to Bile Acid Synthesis in Mice. Gene Expr. 2018, 18, 187–196. [Google Scholar] [CrossRef]
- Jakaria, M.; Azam, S.; Haque, M.E.; Jo, S.H.; Uddin, M.S.; Kim, I.S.; Choi, D.K. Taurine and its analogs in neurological disorders: Focus on therapeutic potential and molecular mechanisms. Redox Biol. 2019, 24, 101223. [Google Scholar] [CrossRef]
- Henry, E.F.; MacCormack, T.J. Taurine protects cardiac contractility in killifish, Fundulus heteroclitus, by enhancing sarcoplasmic reticular Ca(2+) cycling. J. Comp. Physiol. B 2018, 188, 89–99. [Google Scholar] [CrossRef]
- Berry, T.M.; Moustafa, A.A. Osteoporosis and the effect of dysregulation of the transsulfuration pathway via taurine on intracellular calcium homeostasis, vitamin D absorption and vitamin K absorption. Clin. Nutr. ESPEN 2021, 43, 191–196. [Google Scholar] [CrossRef]
- Fukuda, N.; Yoshitama, A.; Sugita, S.; Fujita, M.; Murakami, S. Dietary taurine reduces hepatic secretion of cholesteryl ester and enhances fatty acid oxidation in rats fed a high-cholesterol diet. J. Nutr. Sci. Vitaminol. 2011, 57, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sebastian, B.M.; Tang, H.; McMullen, M.M.; Axhemi, A.; Jacobsen, D.W.; Nagy, L.E. Taurine supplementation prevents ethanol-induced decrease in serum adiponectin and reduces hepatic steatosis in rats. Hepatology 2009, 49, 1554–1562. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, F.G.; Brandao, C.; Batitucci, G.; Souza, A.O.; Ferrari, G.D.; Alberici, L.C.; Muñoz, V.R.; Pauli, J.R.; De Moura, L.P.; Ropelle, E.R.; et al. Taurine supplementation associated with exercise increases mitochondrial activity and fatty acid oxidation gene expression in the subcutaneous white adipose tissue of obese women. Clin. Nutr. 2021, 40, 2180–2187. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Li, F.; Zhang, L.; Duan, Y.; Guo, Q.; Wang, W.; He, S.; Li, J.; Yin, Y. Taurine is Involved in Energy Metabolism in Muscles, Adipose Tissue, and the Liver. Mol. Nutr. Food Res. 2019, 63, 2. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.W.; Shimada-Takaura, K.; Jong, C.J.; Ito, T.; Takahashi, K. Impaired energy metabolism of the taurine-deficient heart. Amino Acids 2016, 48, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Doss, H.M.; Kim, H.J.; Yang, H.I. Taurine Stimulates Thermoregulatory Genes in Brown Fat Tissue and Muscle without an Influence on Inguinal White Fat Tissue in a High-Fat Diet-Induced Obese Mouse Model. Foods 2020, 9, 688. [Google Scholar] [CrossRef] [PubMed]
- Warskulat, U.; Heller-Stilb, B.; Oermann, E.; Zilles, K.; Haas, H.; Lang, F.; Häussinger, D. Phenotype of the taurine transporter knockout mouse. Methods Enzymol. 2007, 428, 439–458. [Google Scholar]
- Cuisinier, C.; Michotte De Welle, J.; Verbeeck, R.K.; Poortmans, J.R.; Ward, R.; Sturbois, X.; Francaux, M. Role of taurine in osmoregulation during endurance exercise. Eur. J. Appl. Physiol. 2002, 87, 489–495. [Google Scholar] [CrossRef]
- Page, L.K.; Jeffries, O.; Waldron, M. Acute taurine supplementation enhances thermoregulation and endurance cycling performance in the heat. Eur. J. Sport. Sci. 2019, 19, 1101–1109. [Google Scholar] [CrossRef]
- Frosini, M.; Sesti, C.; Saponara, S.; Ricci, L.; Valoti, M.; Palmi, M.; Machetti, F.; Sgaragli, G. A specific taurine recognition site in the rabbit brain is responsible for taurine effects on thermoregulation. Br. J. Pharmacol. 2003, 139, 487–494. [Google Scholar] [CrossRef]
- Frosini, M.; Sesti, C.; Saponara, S.; Donati, A.; Palmi, M.; Valoti, M.; Machetti, F.; Sgaragli, G. Effects of taurine and some structurally related analogues on the central mechanism of thermoregulation: A structure-activity relationship study. Adv. Exp. Med. Biol. 2000, 483, 273–282. [Google Scholar]
- Qvartskhava, N.; Jin, C.J.; Buschmann, T.; Albrecht, U.; Bode, J.G.; Monhasery, N.; Oenarto, J.; Bidmon, H.J.; Görg, B.; Häussinger, D. Taurine transporter (TauT) deficiency impairs ammonia detoxification in mouse liver. Proc. Natl. Acad. Sci. USA 2019, 116, 6313–6318. [Google Scholar] [CrossRef] [PubMed]
- Vanitha, M.K.; Baskaran, K.; Periyasamy, K.; Selvaraj, S.; Ilakkia, A.; Saravanan, D.; Venkateswari, R.; Revathi Mani, B.; Anandakumar, P.; Sakthisekaran, D. Modulatory Effect of Taurine on 7,12-Dimethylbenz(a)Anthracene-Induced Alterations in Detoxification Enzyme System, Membrane Bound Enzymes, Glycoprotein Profile and Proliferative Cell Nuclear Antigen in Rat Breast Tissue. J. Biochem. Mol. Toxicol. 2016, 30, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.L.; Viswanathan, P.; Anuradha, C.V. Taurine enhances the metabolism and detoxification of ethanol and prevents hepatic fibrosis in rats treated with iron and alcohol. Environ. Toxicol. Pharmacol. 2009, 27, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Kilb, W.; Fukuda, A. Taurine as an Essential Neuromodulator during Perinatal Cortical Development. Front. Cell Neurosci. 2017, 11, 328. [Google Scholar] [CrossRef]
- Hu, X.Y.; Ma, P.S.; Wu, W.; Zhou, R.; Hao, Y.J.; Niu, Y.; Sun, T.; Li, Y.X.; Yu, J.Q. Neuroprotective actions of taurine on hypoxic-ischemic brain damage in neonatal rats. Brain Res. Bull. 2016, 124, 295–305. [Google Scholar]
- Wu, H.; Jin, Y.; Wei, J.; Jin, H.; Sha, D.; Wu, J.Y. Mode of action of taurine as a neuroprotector. Brain Res. 2005, 1038, 123–131. [Google Scholar] [CrossRef]
- Pradhan, L.K.; Sahoo, P.K.; Aparna, S.; Sargam, M.; Biswal, A.K.; Polai, O.; Chauhan, N.R.; Das, S.K. Suppression of bisphenol A-induced oxidative stress by taurine promotes neuroprotection and restores altered neurobehavioral response in zebrafish (Danio rerio). Environ. Toxicol. 2021, 36, 2342–2353. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino Acids 2014, 46, 7–20. [Google Scholar] [CrossRef]
- Kim, C.; Cha, Y.N. Taurine chloramine produced from taurine under inflammation provides anti-inflammatory and cytoprotective effects. Amino Acids 2014, 46, 89–100. [Google Scholar] [CrossRef]
- Faghfouri, A.H.; Seyyed Shoura, S.M.; Fathollahi, P.; Shadbad, M.A.; Papi, S.; Ostadrahimi, A.; Faghfuri, E. Profiling inflammatory and oxidative stress biomarkers following taurine supplementation: A systematic review and dose-response meta-analysis of controlled trials. Eur. J. Clin. Nutr. 2021, in press. [Google Scholar] [CrossRef]
- Lan, R.; Wan, Z.; Xu, Y.; Wang, Z.; Fu, S.; Zhou, Y.; Lin, X.; Han, X.; Luo, Z.; Miao, J.; et al. Taurine Reprograms Mammary-Gland Metabolism and Alleviates Inflammation Induced by Streptococcus uberis in Mice. Front. Immunol. 2021, 12, 696101. [Google Scholar] [CrossRef] [PubMed]
- Walczewska, M.; Ciszek-Lenda, M.; Surmiak, M.; Kozlowska, A.; Jozefowski, S.; Marcinkiewicz, J. Impact of Taurine on Innate and Adaptive Immunity as the Result of HOCl Neutralization. Adv. Exp. Med. Biol. 2015, 803, 109–120. [Google Scholar] [PubMed]
- Marcinkiewicz, J. Taurine bromamine (TauBr)—Its role in immunity and new perspectives for clinical use. J. Biomed. Sci. 2010, 17 (Suppl. S1), S3. [Google Scholar] [CrossRef]
- Zhou, J.; Lu, Y.; Wu, W.; Feng, Y. Taurine promotes the production of CD4+CD25+FOXP3+ Treg cells through regulating IL-35/STAT1 pathway in a mouse allergic rhinitis model. Allergy Asthma Clin. Immunol. 2021, 17, 59. [Google Scholar] [CrossRef]
- Ramos-Pinto, L.; Machado, M.; Calduch-Giner, J.; Pérez-Sánchez, J.; Dias, J.; Conceição, L.; Silva, T.S.; Costas, B. Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status. Animals 2021, 11, 1193. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.W.; Jong, C.J.; Ramila, K.C.; Azuma, J. Physiological roles of taurine in heart and muscle. J. Biomed. Sci. 2010, 17 (Suppl. S1), S2. [Google Scholar] [CrossRef]
- Ito, T.; Murakami, S.; Schaffer, S. Pathway Analysis of a Transcriptome and Metabolite Profile to Elucidate a Compensatory Mechanism for Taurine Deficiency in the Heart of Taurine Transporter Knockout Mice. J 2018, 1, 7. [Google Scholar] [CrossRef]
- Ito, T.; Oishi, S.; Takai, M.; Kimura, Y.; Uozumi, Y.; Fujio, Y.; Schaffer, S.W.; Azuma, J. Cardiac and skeletal muscle abnormality in taurine transporter-knockout mice. J. Biomed. Sci. 2010, 17 (Suppl. S1), S20. [Google Scholar] [CrossRef]
- Takahashi, Y.; Hatta, H. Effects of taurine administration on exercise-induced fatigue and recovery. J. Phys. Fit. Sports Med. 2017, 6, 33–39. [Google Scholar] [CrossRef]
- Jong, C.J.; Ito, T.; Azuma, J.; Schaffer, S. Taurine Depletion Decreases GRP78 Expression and Downregulates Perk-Dependent Activation of the Unfolded Protein Response. Adv. Exp. Med. Biol. 2015, 803, 571–579. [Google Scholar]
- Jong, C.J.; Azuma, J.; Schaffer, S.W. Role of mitochondrial permeability transition in taurine deficiency-induced apoptosis. Exp. Clin. Cardiol. 2011, 16, 125–128. [Google Scholar] [PubMed]
- Jong, C.J.; Ito, T.; Schaffer, S.W. The ubiquitin-proteasome system and autophagy are defective in the taurine-deficient heart. Amino Acids 2015, 47, 2609–2622. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Castelli, V.; Paladini, A.; d’Angelo, M.; Allegretti, M.; Mantelli, F.; Brandolini, L.; Cocchiaro, P.; Cimini, A.; Varrassi, G. Taurine and oxidative stress in retinal health and disease. CNS Neurosci. Ther. 2021, 27, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I.; Halliwell, B.; Hoey, B.M.; Butler, J. The antioxidant action of taurine, hypotaurine and their metabolic precursors. Biochem. J. 1988, 256, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Chahine, R.; Aftimos, G.; Nader, M.; Mounayar, A.; Esseily, F.; Chamat, S. Protective effect of taurine against free radicals damage in the rat myocardium. Exp. Toxicol. Pathol. 2004, 56, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Mehta, T.R.; Dawson, R., Jr. Taurine is a weak scavenger of peroxynitrite and does not attenuate sodium nitroprusside toxicity to cells in culture. Amino Acids 2001, 20, 419–433. [Google Scholar] [CrossRef]
- Shi, X.; Flynn, D.C.; Porter, D.W.; Leonard, S.S.; Vallyathan, V.; Castranova, V. Efficacy of taurine-based compounds as hydroxyl radical scavengers in silica induced peroxidation. Ann. Clin. Lab. Sci. 1997, 27, 365–374. [Google Scholar]
- Du, H.; You, J.S.; Zhao, X.; Park, J.Y.; Kim, S.H.; Chang, K.J. Antiobesity and hypolipidemic effects of lotus leaf hot water extract with taurine supplementation in rats fed a high fat diet. J. Biomed. Sci. 2010, 17 (Suppl. S1), S42. [Google Scholar] [CrossRef]
- Kilic, F.; Bhardwaj, R.; Caulfeild, J.; Trevithick, J.R. Modelling cortical cataractogenesis 22: Is in vitro reduction of damage in model diabetic rat cataract by taurine due to its antioxidant activity? Exp. Eye Res. 1999, 69, 291–300. [Google Scholar] [CrossRef]
- Oliveira, M.W.; Minotto, J.B.; de Oliveira, M.R.; Zanotto-Filho, A.; Behr, G.A.; Rocha, R.F.; Moreira, J.C.; Klamt, F. Scavenging and antioxidant potential of physiological taurine concentrations against different reactive oxygen/nitrogen species. Pharmacol. Rep. 2010, 62, 185–193. [Google Scholar] [CrossRef]
- Messina, S.A.; Dawson, R., Jr. Attenuation of oxidative damage to DNA by taurine and taurine analogs. Adv. Exp. Med. Biol. 2000, 483, 355–367. [Google Scholar]
- Gupta, R.C. Taurine analogues and taurine transport: Therapeutic advantages. Adv. Exp. Med. Biol. 2006, 583, 449–467. [Google Scholar] [PubMed]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D. The sites and topology of mitochondrial superoxide production. Exp. Gerontol. 2010, 45, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef]
- Wong, H.S.; Dighe, P.A.; Mezera, V.; Monternier, P.A.; Brand, M.D. Production of superoxide and hydrogen peroxide from specific mitochondrial sites under different bioenergetic conditions. J. Biol. Chem. 2017, 292, 16804–16809. [Google Scholar] [CrossRef]
- Sekine, S.; Ichijo, H. Mitochondrial proteolysis: Its emerging roles in stress responses. Biochim. Biophys. Acta 2015, 1850, 274–280. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Larosa, V.; Remacle, C. Insights into the respiratory chain and oxidative stress. Biosci. Rep. 2018, 38, BSR2017149. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Stella, A.M.; Calabrese, E.J. Cellular stress responses, mitostress and carnitine insufficiencies as critical determinants in aging and neurodegenerative disorders: Role of hormesis and vitagenes. Neurochem. Res. 2010, 35, 1880–1915. [Google Scholar] [CrossRef] [PubMed]
- Lobo, M.V.; Alonso, F.J.; Martin del Rio, R. Immunocytochemical localization of taurine in different muscle cell types of the dog and rat. Histochem. J. 2000, 32, 53–61. [Google Scholar] [CrossRef]
- Lobo, M.V.; Alonso, F.J.; del Río, R.M. Immunohistochemical localization of taurine in the male reproductive organs of the rat. J. Histochem. Cytochem. 2000, 48, 313–320. [Google Scholar] [CrossRef]
- Jong, C.J.; Ito, T.; Mozaffari, M.; Azuma, J.; Schaffer, S. Effect of beta-alanine treatment on mitochondrial taurine level and 5-taurinomethyluridine content. J. Biomed. Sci. 2010, 17 (Suppl. S1), S25. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Suzuki, T.; Wada, T.; Saigo, K.; Watanabe, K. Taurine as a constituent of mitochondrial tRNAs: New insights into the functions of taurine and human mitochondrial diseases. EMBO J. 2002, 21, 6581–6589. [Google Scholar] [CrossRef] [PubMed]
- Kirino, Y.; Yasukawa, T.; Ohta, S.; Akira, S.; Ishihara, K.; Watanabe, K.; Suzuki, T. Codon-specific translational defect caused by a wobble modification deficiency in mutant tRNA from a human mitochondrial disease. Proc. Natl. Acad. Sci. USA 2004, 101, 15070–15075. [Google Scholar] [CrossRef]
- Fakruddin, M.; Wei, F.Y.; Suzuki, T.; Asano, K.; Kaieda, T.; Omori, A.; Izumi, R.; Fujimura, A.; Kaitsuka, T.; Miyata, K.; et al. Defective Mitochondrial tRNA Taurine Modification Activates Global Proteostress and Leads to Mitochondrial Disease. Cell Rep. 2018, 22, 482–496. [Google Scholar] [CrossRef]
- Schaffer, S.W.; Azuma, J.; Mozaffari, M. Role of antioxidant activity of taurine in diabetes. Can. J. Physiol. Pharmacol. 2009, 87, 91–99. [Google Scholar] [CrossRef]
- Warskulat, U.; Borsch, E.; Reinehr, R.; Heller-Stilb, B.; Mönnighoff, I.; Buchczyk, D.; Donner, M.; Flögel, U.; Kappert, G.; Soboll, S.; et al. Chronic liver disease is triggered by taurine transporter knockout in the mouse. FASEB J. 2006, 20, 574–576. [Google Scholar] [CrossRef]
- Rikimaru, M.; Ohsawa, Y.; Wolf, A.M.; Nishimaki, K.; Ichimiya, H.; Kamimura, N.; Nishimatsu, S.; Ohta, S.; Sunada, Y. Taurine ameliorates impaired the mitochondrial function and prevents stroke-like episodes in patients with MELAS. Int. Med. 2012, 51, 3351–3357. [Google Scholar] [CrossRef]
- Hansen, S.H.; Grunnet, N. Taurine, glutathione and bioenergetics. Adv. Exp. Med. Biol. 2013, 776, 3–12. [Google Scholar]
- Hansen, S.H.; Birkedal, H.; Wibrand, F.; Grunnet, N. Taurine and regulation of mitochondrial metabolism. Adv. Exp. Med. Biol. 2015, 803, 397–405. [Google Scholar] [PubMed]
- Mézes, M.; Surai, P.; Sályi, G.; Speake, B.K.; Gaál, T.; Maldjian, A. Nutritional metabolic diseases of poultry and disorders of the biological antioxidant defence system. Acta Vet. Hung. 1997, 45, 349–360. [Google Scholar] [PubMed]
- El Idrissi, A. Taurine increases mitochondrial buffering of calcium: Role in neuroprotection. Amino Acids 2008, 34, 321–328. [Google Scholar] [CrossRef]
- El Idrissi, A.; Trenkner, E. Growth factors and taurine protect against excitotoxicity by stabilizing calcium homeostasis and energy metabolism. J. Neurosci. 1999, 19, 9459–9568. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.W.; Ramila, K.C.; Jong, C.J.; Shetewy, A.; Shimada, K.; Ito, T.; Azuma, J.; Cioffi, E. Does taurine prolong lifespan by improving heart function? Adv. Exp. Med. Biol. 2015, 803, 555–570. [Google Scholar] [PubMed]
- Chang, L.; Xu, J.X.; Zhao, J.; Pang, Y.Z.; Tang, C.S.; Qi, Y.F. Taurine antagonized oxidative stress injury induced by homocysteine in rat vascular smooth muscle cells. Acta Pharmacol. Sin. 2004, 25, 341–346. [Google Scholar] [PubMed]
- Parvez, S.; Tabassum, H.; Banerjee, B.D.; Raisuddin, S. Taurine prevents tamoxifen-induced mitochondrial oxidative damage in mice. Basic Clin. Pharmacol. Toxicol. 2008, 102, 382–387. [Google Scholar] [CrossRef]
- Ahmadi, N.; Ghanbarinejad, V.; Ommati, M.M.; Jamshidzadeh, A.; Heidari, R. Taurine prevents mitochondrial membrane permeabilization and swelling upon interaction with manganese: Implication in the treatment of cirrhosis-associated central nervous system complications. J. Biochem. Mol. Toxicol. 2018, 32, e22216. [Google Scholar] [CrossRef]
- Niu, X.; Zheng, S.; Liu, H.; Li, S. Protective effects of taurine against inflammation, apoptosis, and oxidative stress in brain injury. Mol. Med. Rep. 2018, 18, 4516–4522. [Google Scholar] [CrossRef]
- Cai, J.G.; Luo, L.M.; Tang, H.; Zhou, L. Cytotoxicity of Malondialdehyde and Cytoprotective Effects of Taurine via Oxidative Stress and PGC-1α Signal Pathway in C2C12 Cells. Mol. Biol. 2018, 52, 616–627. [Google Scholar] [CrossRef]
- Estévez, M.; Xiong, Y. Intake of Oxidized Proteins and Amino Acids and Causative Oxidative Stress and Disease: Recent Scientific Evidences and Hypotheses. J. Food. Sci. 2019, 84, 387–396. [Google Scholar] [CrossRef]
- Hirata, H.; Ueda, S.; Ichiseki, T.; Shimasaki, M.; Ueda, Y.; Kaneuji, A.; Kawahara, N. Taurine Inhibits Glucocorticoid-Induced Bone Mitochondrial Injury, Preventing Osteonecrosis in Rabbits and Cultured Osteocytes. Int. J. Mol. Sci. 2020, 21, 6892. [Google Scholar] [CrossRef] [PubMed]
- Rezaee-Tazangi, F.; Zeidooni, L.; Rafiee, Z.; Fakhredini, F.; Kalantari, H.; Alidadi, H.; Khorsandi, L. Taurine effects on Bisphenol A-induced oxidative stress in the mouse testicular mitochondria and sperm motility. JBRA Assist. Reprod. 2020, 24, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Huxtable, R.; Bressler, R. Effect of taurine on a muscle intracellular membrane. Biochim. Biophys. Acta 1973, 323, 573–583. [Google Scholar] [CrossRef]
- Pasantes-Morales, H.; Cruz, C. Protective effect of taurine and zinc on peroxidation-induced damage in photoreceptor outer segments. J. Neurosci. Res. 1984, 11, 303–311. [Google Scholar] [CrossRef]
- Pasantes-Morales, H.; Wright, C.E.; Gaull, G.E. Protective effect of taurine, zinc and tocopherol on retinol-induced damage in human lymphoblastoid cells. J. Nutr. 1984, 114, 2256–2261. [Google Scholar] [CrossRef]
- Pasantes-Morales, H.; Cruz, C. Taurine: A physiological stabilizer of photoreceptor membranes. Prog. Clin. Biol. Res. 1985, 179, 371–381. [Google Scholar]
- Kulakowski, E.C.; Maturo, J. Hypoglycemic properties of taurine: Not mediated by enhanced insulin release. Biochem. Pharmacol. 1984, 33, 2835–2838. [Google Scholar] [CrossRef]
- Pasantes-Morales, H.; Cruz, C. Taurine and hypotaurine inhibit light-induced lipid peroxidation and protect rod outer segment structure. Brain Res. 1985, 330, 154–157. [Google Scholar] [CrossRef]
- Lombardini, J.B. Inhibition by taurine of the phosphorylation of rat retinal membranes. Prog. Clin. Biol. Res. 1985, 179, 383–393. [Google Scholar] [PubMed]
- Liebowitz, S.M.; Lombardini, J.B.; Allen, C.I. Effects of aminocycloalkanesulfonic acid analogs of taurine on ATP-dependent calcium ion uptake and protein phosphorylation. Biochem. Pharmacol. 1988, 37, 1303–1309. [Google Scholar] [CrossRef]
- Hamaguchi, T.; Azuma, J.; Schaffer, S. Interaction of taurine with methionine: Inhibition of myocardial phospholipid methyltransferase. J. Cardiovasc. Pharmacol. 1991, 18, 224–230. [Google Scholar] [CrossRef]
- Chen, C.; Xia, S.; He, J.; Lu, G.; Xie, Z.; Han, H. Roles of taurine in cognitive function of physiology, pathologies and toxication. Life Sci. 2019, 231, 116584. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; He, M.; Zhong, M.; Li, L.; Lu, Y.; Zhang, Y.; Zhang, L.; Yu, Z.; Zhou, Z. The neuroprotective effects of taurine against nickel by reducing oxidative stress and maintaining mitochondrial function in cortical neurons. Neurosci. Lett. 2015, 590, 52–57. [Google Scholar] [CrossRef] [PubMed]
- El Idrissi, A. Taurine Regulation of Neuroendocrine Function. Adv. Exp. Med. Biol. 2019, 1155, 977–985. [Google Scholar]
- Surai, P.F. Vitamin E in avian reproduction. Poult. Avian Biol. Rev. 1999, 10, 1–60. [Google Scholar]
- Surai, P.F.; Speake, B.K.; Sparks, N.H.C. Comparative Aspects of Lipid Peroxidation and Antioxidant Protection in Avian Semen. In Male Fertility and Lipid Metabolism; De Vriese, S., Christophe, A., Eds.; AOCS Press: Champaign, IL, USA, 2003; Chapter 15; pp. 211–249. [Google Scholar]
- Alvarez, J.G.; Storey, B.T. Taurine, hypotaurine, epinephrine and albumin inhibit lipid peroxidation in rabbit spermatozoa and protect against loss of motility. Biol. Reprod. 1983, 29, 548–555. [Google Scholar] [CrossRef]
- Michael, A.J.; Alexopoulos, C.; Pontiki, E.A.; Hadjipavlou-Litina, D.J.; Saratsis, P.; Ververidis, H.N.; Boscos, C.M. Effect of antioxidant supplementation in semen extenders on semen quality and reactive oxygen species of chilled canine spermatozoa. Anim. Reprod. Sci. 2009, 112, 119–135. [Google Scholar] [CrossRef]
- Ozasa, H.; Gould, K.G. Protective effect of taurine from osmotic stress on chimpanzee spermatozoa. Arch. Androl. 1982, 9, 121–126. [Google Scholar] [CrossRef]
- Perumal, P.; Vupru, K.; Rajkhowa, C. Effect of Addition of Taurine on the Liquid Storage (5 °C) of Mithun (Bos frontalis) Semen. Vet. Med. Int. 2013, 2013, 165348. [Google Scholar] [CrossRef] [PubMed]
- Shiva Shankar Reddy, N.; Jagan Mohanarao, G.; Atreja, S.K. Effects of adding taurine and trehalose to a tris-based egg yolk extender on buffalo (Bubalus bubalis) sperm quality following cryopreservation. Anim. Reprod. Sci. 2010, 119, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Singh, V.K.; Chhillar, S.; Atreja, S.K. Effect of supplementation of taurine or trehalose in extender on immunolocalization of tyrosine phosphoproteins in buffalo and cattle (Karan Fries) cryopreserved spermatozoa. Reprod. Domest. Anim. 2013, 48, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Chhillar, S.; Singh, V.K.; Kumar, R.; Atreja, S.K. Effects of Taurine or Trehalose supplementation on functional competence of cryopreserved Karan Fries semen. Anim. Reprod. Sci. 2012, 135, 1–7. [Google Scholar] [CrossRef]
- Bottrel, M.; Acha, D.; Ortiz, I.; Hidalgo, M.; Gósalvez, J.; Camisão, J.; Dorado, J. Cryoprotective effect of glutamine, taurine, and proline on post-thaw semen quality and DNA integrity of donkey spermatozoa. Anim. Reprod. Sci. 2018, 189, 128–135. [Google Scholar] [CrossRef]
- Martins-Bessa, A.; Rocha, A.; Mayenco-Aguirre, A. Effects of taurine and hypotaurine supplementation and ionophore concentrations on post-thaw acrosome reaction of dog spermatozoa. Theriogenology 2009, 71, 248–253. [Google Scholar] [CrossRef]
- Partyka, A.; Rodak, O.; Bajzert, J.; Kochan, J.; Niżański, W. The Effect of L-Carnitine, Hypotaurine, and Taurine Supplementation on the Quality of Cryopreserved Chicken Semen. Biomed. Res. Int. 2017, 2017, 7279341. [Google Scholar] [CrossRef]
- Sánchez-Partida, L.G.; Setchell, B.P.; Maxwell, W.M. Epididymal compounds and antioxidants in diluents for the frozen storage of ram spermatozoa. Reprod. Fertil. Dev. 1997, 9, 689–696. [Google Scholar] [CrossRef]
- Banday, M.N.; Lone, F.A.; Rasool, F.; Rashid, M.; Shikari, A. Use of antioxidants reduce lipid peroxidation and improve quality of crossbred ram sperm during its cryopreservation. Cryobiology 2017, 74, 25–30. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Sohail, T.; Kang, Y.; Niu, H.; Sun, X.; Ji, D.; Li, Y. Effects of Taurine on Sperm Quality during Room Temperature Storage in Hu Sheep. Animals 2021, 11, 2725. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.G.; Fang, Q.; Liu, Q.; Du, R.R.; Yang, G.S.; Wang, L.Q.; Hu, J.H. Supplemental effect of different levels of taurine in Modena on boar semen quality during liquid preservation at 17 °C. Anim. Sci. J. 2017, 88, 1692–1699. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, A.; Ducharme, R. Effect of various extenders and taurine on survival of stallion sperm cooled to 5 degrees C. Theriogenology 1995, 44, 1039–1050. [Google Scholar] [CrossRef]
- Du, Y.; Liu, H.; Zhang, M.; Zhang, S.; Hu, J.; Wu, G.; Yang, J. Taurine Increases Spermatozoa Quality and Function in Asthenospermia Rats Impaired by Ornidazole. Adv. Exp. Med. Biol. 2019, 1155, 507–520. [Google Scholar] [PubMed]
- Seify, M.; Zarabadipour, M.; Ghaleno, L.R.; Alizadeh, A.; Rezazadeh Valojerdi, M. The anti-oxidant roles of Taurine and Hypotaurine on acrosome integrity, HBA and HSPA2 of the human sperm during vitrification and post warming in two different temperature. Cryobiology 2019, 90, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Kutluyer, F.; Öğretmen, F.; Inanan, B.E. Cryopreservation of Goldfish (Carassius auratus) spermatozoa: Effects of extender supplemented with taurine on sperm motility and DNA damage. Cryo. Lett. 2016, 37, 41–46. [Google Scholar]
- Martínez-Páramo, S.; Diogo, P.; Dinis, M.T.; Soares, F.; Sarasquete, C.; Cabrita, E. Effect of two sulfur-containing amino acids, taurine and hypotaurine in European sea bass (Dicentrarchus labrax) sperm cryopreservation. Cryobiology 2013, 66, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Labat-Robert, J.; Robert, L. Longevity and aging. Role of free radicals and xanthine oxidase. A review. Pathol. Biol. 2014, 62, 61–66. [Google Scholar] [CrossRef]
- Doehner, W.; Landmesser, U. Xanthine oxidase and uric acid in cardiovascular disease: Clinical impact and therapeutic options. Semin. Nephrol. 2011, 31, 433–440. [Google Scholar] [CrossRef]
- Das, J.; Vasan, V.; Sil, P.C. Taurine exerts hypoglycemic effect in alloxan-induced diabetic rats, improves insulin-mediated glucose transport signaling pathway in heart and ameliorates cardiac oxidative stress and apoptosis. Toxicol. Appl. Pharmacol. 2012, 258, 296–308. [Google Scholar] [CrossRef]
- Das, J.; Sil, P.C. Taurine ameliorates alloxan-induced diabetic renal injury, oxidative stress-related signaling pathways and apoptosis in rats. Amino Acids 2012, 43, 1509–1523. [Google Scholar] [CrossRef]
- Ahmad, M.K.; Mahmood, R. Protective effect of taurine against potassium bromate-induced hemoglobin oxidation, oxidative stress, and impairment of antioxidant defense system in blood. Environ. Toxicol. 2016, 31, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Sun, F.; Gao, Y.; Yang, J.; Wu, G.; Lin, S.; Hu, J. Taurine decreased uric acid levels in hyperuricemic rats and alleviated kidney injury. Biochem. Biophys. Res. Commun. 2017, 489, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Filip-Ciubotaru, F.; Manciuc, C.; Stoleru, G.; Liliana, F.O.I.A. Nadph oxidase: Structure and activation mechanisms (review). Note I. Rev. Med. Chir. Soc. Med. Nat. Iasi. 2016, 120, 29–33. [Google Scholar]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Mozaffari, M.S.; Tan, B.H.; Lucia, M.A.; Schaffer, S.W. Effect of drug-induced taurine depletion on cardiac contractility and metabolism. Biochem. Pharamacol. 1986, 35, 985–989. [Google Scholar] [CrossRef]
- Li, Y.; Arnold, J.M.; Pampillo, M.; Babwah, A.V.; Peng, T. Taurine prevents cardiomyocyte death by inhibiting NADPH oxidase-mediated calpain activation. Free Radic. Biol. Med. 2009, 46, 51–61. [Google Scholar] [CrossRef]
- Winiarska, K.; Szymanski, K.; Gorniak, P.; Dudziak, M.; Bryla, J. Hypoglycaemic, antioxidative and nephroprotective effects of taurine in alloxan diabetic rabbits. Biochimie 2009, 91, 261–270. [Google Scholar] [CrossRef]
- Miao, J.; Zhang, J.; Zili, M.; Zheng, L. The role of NADPH oxidase in taurine attenuation of Stereptococcus uberis -induced mastitis in rats. Int. Immunopharmacol. 2013, 16, 429–435. [Google Scholar] [CrossRef]
- Han, Z.; Gao, L.Y.; Lin, Y.H.; Chang, L.; Wu, H.Y.; Luo, C.X.; Zhu, D.Y. Neuroprotection of taurine against reactive oxygen species is associated with inhibiting NADPH oxidases. Eur. J. Pharmacol. 2016, 777, 129–135. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, S.; Gu, Y.; Wang, J.; Liu, Z.; Zhang, Z. Taurine Attenuates Calpain-2 Induction and a Series of Cell Damage via Suppression of NOX-Derived ROS in ARPE-19 Cells. Oxid. Med. Cell Longev. 2018, 2018, 4596746. [Google Scholar] [CrossRef]
- Che, Y.; Hou, L.; Sun, F.; Zhang, C.; Liu, X.; Piao, F.; Zhang, D.; Li, H.; Wang, Q. Taurine protects dopaminergic neurons in a mouse Parkinson’s disease model through inhibition of microglial M1 polarization. Cell Death Dis. 2018, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Che, Y.; Sun, F.; Wang, Q. Taurine protects noradrenergic locus coeruleus neurons in a mouse Parkinson’s disease model by inhibiting microglial M1polarization. Amino Acids 2018, 50, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Scicchitano, B.M.; Sica, G. The Beneficial Effects of Taurine to Counteract Sarcopenia. Curr. Protein Pept. Sci. 2018, 19, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Ingawale, D.K.; Mandlik, S.K.; Naik, S.R. Models of hepatotoxicity and the underlying cellular, biochemical and immunological mechanism(s): A critical discussion. Environ. Toxicol. Pharmacol. 2014, 37, 118–133. [Google Scholar] [CrossRef]
- Sriset, Y.; Chatuphonprasert, W.; Jarukamjorn, K. A Review on the Mechanism ff Oxidative Stress-Induction in the Liver by Xenobiotics. Isan J. Pharm. Sci. 2018, 14, 1–12. [Google Scholar]
- Sheikh, M.A.; Tembhre, M. Liver Toxicity and its Amelioration by Natural Antioxidants–A Review. Asian J. Exp. Sci. 2018, 32, 35–43. [Google Scholar]
- Upadhyay, M.K.; Shukla, A.; Yadav, P.; Srivastava, S. A review of arsenic in crops, vegetables, animals and food products. Food Chem. 2019, 276, 608–618. [Google Scholar] [CrossRef]
- Chandravanshi, L.P.; Gupta, R.; Shukla, R.K. Developmental Neurotoxicity of Arsenic: Involvement of Oxidative Stress and Mitochondrial Functions. Biol. Trace Elem. Res. 2018, 186, 185–198. [Google Scholar] [CrossRef]
- Flora, S.J.; Mittal, M.; Mehta, A. Heavy metal induced oxidative stress & its possible reversal by chelation therapy. Indian J. Med. Res. 2008, 128, 501–523. [Google Scholar]
- Ghosh, J.; Das, J.; Manna, P.; Sil, P.C. Taurine prevents arsenic-induced cardiac oxidative stress and apoptotic damage: Role of NF-kappa B, p38 and JNK MAPK pathway. Toxicol. Appl. Pharmacol. 2009, 240, 73–87. [Google Scholar] [CrossRef]
- Roy, A.; Manna, P.; Sil, P.C. Prophylactic role of taurine on arsenic mediated oxidative renal dysfunction via MAPKs/NF-kappaB and mitochondria dependent pathways. Free Radic. Res. 2009, 43, 995–1007. [Google Scholar] [CrossRef]
- Das, J.; Ghosh, J.; Manna, P.; Sinha, M.; Sil, P.C. Arsenic-induced oxidative cerebral disorders: Protection by taurine. Drug Chem. Toxicol. 2009, 32, 93–102. [Google Scholar] [CrossRef]
- Das, J.; Ghosh, J.; Manna, P.; Sinha, M.; Sil, P.C. Taurine protects rat testes against NaAsO(2)-induced oxidative stress and apoptosis via mitochondrial dependent and independent pathways. Toxicol. Lett. 2009, 187, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Yao, X.; Jiang, L.; Qiu, T.; Liu, S.; Qi, B.; Zheng, Y.; Kong, Y.; Yang, G.; Chen, M.; et al. Taurine protects against As2O3-induced autophagy in pancreas of rat offsprings through Nrf2/Trx pathway. Biochimie 2016, 123, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Manna, P.; Sil, P.C. Taurine, a conditionally essential amino acid, ameliorates arsenic-induced cytotoxicity in murine hepatocytes. Toxicol. Vitr. 2007, 21, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Pei, P.; Yao, X.; Jiang, L.; Wei, S.; Wang, Z.; Bai, J.; Yang, G.; Gao, N.; Yang, L.; et al. Taurine attenuates arsenic-induced pyroptosis and nonalcoholic steatohepatitis by inhibiting the autophagic-inflammasomal pathway. Cell Death Dis. 2018, 9, 946. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Yao, X.; Jiang, L.; Yang, L.; Qiu, T.; Wang, Z.; Pei, P.; Yang, G.; Liu, X.; Sun, X. Taurine improves low-level inorganic arsenic-induced insulin resistance by activating PPARγ-mTORC2 signalling and inhibiting hepatic autophagy. J. Cell Physiol. 2019, 234, 5143–5152. [Google Scholar] [CrossRef]
- Zheng, Y.; Qu, H.; Wang, D.; Li, S.; Zhang, C.; Piao, F. Protection of Taurine against Arsenic-Induced DNA Damage of Mice Kidneys. Adv. Exp. Med. Biol. 2017, 975, 917–927. [Google Scholar]
- Zhang, C.; Liu, X.; Li, S.; Guo, W.; Chen, M.; Yan, X.; Jiang, L.; Piao, F. Taurine Normalizes the Levels of Se, Cu, Fe in Mouse Liver and Kidney Exposed to Arsenic Subchronically. Adv. Exp. Med. Biol. 2017, 975, 843–853. [Google Scholar]
- Ma, N.; Sasoh, M.; Kawanishi, S.; Sugiura, H.; Piao, F. Protection effect of taurine on nitrosative stress in the mice brain with chronic exposure to arsenic. J. Biomed. Sci. 2010, 17 (Suppl. S1), S7. [Google Scholar] [CrossRef]
- Piao, F.; Zhang, Y.; Yang, L.; Zhang, C.; Shao, J.; Liu, X.; Li, Y.; Li, S. Taurine Attenuates As(2)O(3)-Induced Autophagy in Cerebrum of Mouse Through Nrf2 Pathway. Adv. Exp. Med. Biol. 2017, 975, 863–870. [Google Scholar]
- Wang, Y.; Piao, F.; Li, Y.; Wang, X.; Guang, H. Protective effect of taurine on down-regulated expression of thyroid hormone receptor genes in brains of mice exposed to arsenic. Adv. Exp. Med. Biol. 2013, 775, 155–166. [Google Scholar]
- Li, S.; Yang, L.; Zhang, Y.; Zhang, C.; Shao, J.; Liu, X.; Li, Y.; Piao, F. Taurine Ameliorates Arsenic-Induced Apoptosis in the Hippocampus of Mice Through Intrinsic Pathway. Adv. Exp. Med. Biol. 2017, 975, 183–192. [Google Scholar]
- Li, S.; Wei, B.K.; Wang, J.; Dong, G.; Wang, X. Taurine Supplementation Ameliorates Arsenic-Induced Hepatotoxicity and Oxidative Stress in Mouse. Adv. Exp. Med. Biol. 2019, 1155, 463–470. [Google Scholar] [PubMed]
- Zhang, Q.; Bai, J.; Yao, X.; Jiang, L.; Wu, W.; Yang, L.; Gao, N.; Qiu, T.; Yang, G.; Habtemariam Hidru, T.; et al. Taurine rescues the arsenic-induced injury in the pancreas of rat offsprings and in the INS-1 cells. Biomed. Pharmacother. 2019, 109, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Rashid, K.; Sinha, K.; Sil, P.C. An update on oxidative stress-mediated organ pathophysiology. Food Chem. Toxicol. 2013, 62, 584–600. [Google Scholar] [CrossRef] [PubMed]
- Manibusan, M.K.; Odin, M.; Eastmond, D.A. Postulated carbon tetrachloride mode of action: A review. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2007, 25, 185–209. [Google Scholar] [CrossRef]
- Jiang, L.; Huang, J.; Wang, Y.; Tang, H. Metabonomic analysis reveals the CCl4-induced systems alterations for multiple rat organs. J. Proteome Res. 2012, 11, 3848–3859. [Google Scholar] [CrossRef]
- Raj, S.; Gothandam, K.M. Hepatoprotective effect of polyphenols rich methanolic extract of Amorphophallus commutatus var. Wayanadensis against CCl4 induced hepatic injury in swiss albino mice. Food Chem. Toxicol. 2014, 67, 105–112. [Google Scholar]
- Alkreathy, H.M.; Khan, R.A.; Khan, M.R.; Sahreen, S. CCl4 induced genotoxicity and DNA oxidative damages in rats: Hepatoprotective effect of Sonchus arvensis. BMC Complement. Altern. Med. 2014, 14, 452. [Google Scholar] [CrossRef]
- Krishnappa, P.; Venkatarangaiah, K.; Shivamogga, V.; Rajanna, S.K.; Prakash, K.; Gupta, R. Antioxidant and prophylactic effects of Delonix elata L., stem bark extracts, and flavonoid isolated quercetin against carbon tetrachloride-induced hepatotoxicity in rats. Biomed. Res. Int. 2014, 2014, 507851. [Google Scholar] [CrossRef] [PubMed]
- Shaker, E.; Mahmoud, H.; Mnaa, S. Silymarin, the antioxidant component and Silybum marianum extracts prevent liver damage. Food Chem. Toxicol. 2010, 48, 803–806. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Taniko, T.; Kuriyama, K. Therapeutic effect of taurine administration on carbon tetrachloride-induced hepatic injury. Jpn. J. Pharmacol. 1982, 32, 583–589. [Google Scholar] [CrossRef]
- Nakashima, T.; Takino, T.; Kuriyama, K. Pretreatment with taurine facilitates hepatic lipid peroxide formation associated with carbon tetrachloride intoxication. Jpn. J. Pharmacol. 1983, 33, 515–523. [Google Scholar] [CrossRef]
- Waterfield, C.J.; Mesquita, M.; Parnham, P.; Timbrell, J.A. Cytoprotective effects of taurine in isolated rat hepatocytes. Toxicol. Vitr. 1994, 8, 573–575. [Google Scholar] [CrossRef]
- Timbrell, J.A.; Seabra, V.; Watereld, C.J. The in vivo and in vitro protective properties of taurine. Gen. Pharmacol. 1995, 26, 453–462. [Google Scholar] [CrossRef]
- Wu, Q.D.; Wang, J.H.; Fennessy, F.; Redmond, H.P.; Bouchier-Hayes, D. Taurine prevents high-glucose-induced human vascular endothelial cell apoptosis. Am. J. Physiol. 1999, 277, C1229–C1238. [Google Scholar]
- Vohra, B.P.; Hui, X. Taurine protects against carbon tetrachloride toxicity in the cultured neurons and in vivo. Arch. Physiol. Biochem. 2001, 109, 90–94. [Google Scholar] [CrossRef]
- Timbrell, J.A.; Waterfield, C.J. Changes in taurine as an indicator of hepatic dysfunction and biochemical perturbations. Studies in vivo and in vitro. Adv. Exp. Med. Biol. 1996, 403, 125–134. [Google Scholar]
- Waterfield, C.J.; Turton, J.A.; Scales, M.D.; Timbrell, J.A. Taurine, a possible urinary marker of liver damage: A study of taurine excretion in carbon tetrachloride-treated rats. Arch. Toxicol. 1991, 65, 548–555. [Google Scholar] [CrossRef]
- Waterfield, C.J.; Turton, J.A.; Scales, M.D.; Timbrell, J.A. Reduction of liver taurine in rats by beta-alanine treatment increases carbon tetrachloride toxicity. Toxicology 1993, 77, 7–20. [Google Scholar] [CrossRef]
- Zhou, J.; Zhou, J.; Zhong, X.; Chen, L.; Yang, X. Abnormal expressions of hepatocellular proteins and extracellular matrix in CCL4-induced liver injury in rats. Chin. Med. J. 1996, 109, 366–371. [Google Scholar] [PubMed]
- Dincer, S.; Ozenirler, S.; Oz, E.; Akyol, G.; Ozogul, C. The protective effect of taurine pretreatment on carbon tetrachloride-induced hepatic damage—A light and electron microscopic study. Amino Acids 2002, 22, 417–426. [Google Scholar] [PubMed]
- Erman, F.; Balkan, J.; Cevikbas, U.; Kocak-Toker, N.; Uysal, M. Betaine or taurine administration prevents fibrosis and lipid peroxidation induced by rat liver by ethanol plus carbon tetrachloride intoxication. Amino Acids 2004, 27, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Karube, M.; Matsuzaki, Y.; Ikegami, T.; Doy, M.; Tanaka, N.; Bouscarel, B. Taurine inhibits oxidative damage and prevents fibrosis in carbon tetrachloride-induced hepatic fibrosis. J. Hepatol. 2005, 43, 117–125. [Google Scholar] [CrossRef]
- Tasci, I.; Mas, N.; Mas, M.R.; Tuncer, M.; Comert, B. Ultrastructural changes in hepatocytes after taurine treatment in CCl4 induced liver injury. World J. Gastroenterol. 2008, 14, 4897–4902. [Google Scholar] [CrossRef]
- Miyazaki, T.; Bouscarel, B.; Ikegami, T.; Honda, A.; Matsuzaki, Y. The protective effect of taurine against hepatic damage in a model of liver disease and hepatic stellate cells. Adv. Exp. Med. Biol. 2009, 643, 293–303. [Google Scholar]
- Abd-Elhakim, Y.M.; Ghoneim, M.H.; Ebraheim, L.L.M.; Imam, T.S. Taurine and hesperidin rescues carbon tetrachloride-triggered testicular and kidney damage in rats via modulating oxidative stress and inflammation. Life Sci. 2020, 254, 117782. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.M.; Al-Kahtani, M.A.; El-Kersh, M.A.; Al-Omair, M.A. Free Radical-Scavenging, Anti-Inflammatory/Anti-Fibrotic and Hepatoprotective Actions of Taurine and Silymarin against CCl4 Induced Rat Liver Damage. PLoS ONE 2015, 10, e0144509. [Google Scholar]
- Cao, W.; Li, Y.; Li, M.; Zhang, X.; Liao, M. Txn1, Ctsd and Cdk4 are key proteins of combination therapy with taurine, epigallocatechin gallate and genistein against liver fibrosis in rats. Biomed. Pharmacother. 2017, 85, 611–619. [Google Scholar] [CrossRef]
- Mustafa, H.N.; El Awdan, S.A.; Hegazy, G.A. Protective role of antioxidants on thioacetamide-induced acute hepatic encephalopathy: Biochemical and ultrastructural study. Tissue Cell 2013, 45, 350–362. [Google Scholar] [CrossRef]
- Fazal, Y.; Fatima, S.N.; Shahid, S.M.; Mahboob, T. Effects of curcumin on angiotensin-converting enzyme gene expression, oxidative stress and anti-oxidant status in thioacetamide-induced hepatotoxicity. J. Renin Angiotensin Aldosterone Syst. 2015, 16, 1046–1051. [Google Scholar] [CrossRef] [PubMed]
- Balkan, J.; Doğru-Abbasoğlu, S.; Kanbağli, O.; Cevikbaş, U.; Aykaç-Toker, G.; Uysal, M. Taurine has a protective effect against thioacetamide-induced liver cirrhosis by decreasing oxidative stress. Hum. Exp. Toxicol. 2001, 20, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Doğru-Abbasoğlu, S.; Kanbağli, O.; Balkan, J.; Cevikbaş, U.; Aykaç-Toker, G.; Uysal, M. The protective effect of taurine against thioacetamide hepatotoxicity of rats. Hum. Exp. Toxicol. 2001, 20, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Hammes, T.O.; Pedroso, G.L.; Hartmann, C.R.; Escobar, T.D.; Fracasso, L.B.; da Rosa, D.P.; Marroni, N.P.; Porawski, M.; da Silveira, T.R. The effect of taurine on hepatic steatosis induced by thioacetamide in zebrafish (Danio rerio). Dig. Dis. Sci. 2012, 57, 675–682. [Google Scholar] [CrossRef]
- Jamshidzadeh, A.; Heidari, R.; Abasvali, M.; Zarei, M.; Ommati, M.M.; Abdoli, N.; Khodaei, F.; Yeganeh, Y.; Jafari, F.; Zarei, A.; et al. Taurine treatment preserves brain and liver mitochondrial function in a rat model of fulminant hepatic failure and hyperammonemia. Biomed. Pharmacother. 2017, 86, 514–520. [Google Scholar] [CrossRef]
- Younis, N.S.; Ghanim, A.M.H.; Elmorsy, M.A.; Metwaly, H.A. Taurine ameliorates thioacetamide induced liver fibrosis in rats via modulation of toll like receptor 4/nuclear factor kappa B signaling pathway. Sci. Rep. 2021, 11, 12296. [Google Scholar] [CrossRef]
- Ghanim, A.; Farag, M.; Anwar, M.; Ali, N.; Hawas, M.; Elsallab, H.; Elhendawy, W.; Basyouni, L.; Refaey, O.; Zaki, K.; et al. Taurine alleviates kidney injury in a thioacetamide rat model by mediating Nrf2/HO-1, NQO-1 and MAPK/NF-κB signaling pathways. Can. J. Physiol. Pharmacol. 2021, in press. [Google Scholar] [CrossRef]
- Dos Santos, N.A.; Carvalho Rodrigues, M.A.; Martins, N.M.; dos Santos, A.C. Cisplatin-induced nephrotoxicity and targets of nephroprotection: An update. Arch. Toxicol. 2012, 86, 1233–1250. [Google Scholar] [CrossRef]
- Mansour, H.H.; Hafez, H.F.; Fahmy, N.M. Silymarin modulates Cisplatin-induced oxidative stress and hepatotoxicity in rats. J. Biochem. Mol. Biol. 2006, 39, 656–661. [Google Scholar] [CrossRef]
- El-Awady, S.E.; Moustafa, Y.M.; Abo-Elmatty, D.M.; Radwan, A. Cisplatin-induced cardiotoxicity: Mechanisms and cardioprotective strategies. Eur. J. Pharmacol. 2011, 650, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Sierra, T.; Eugenio-Pérez, D.; Sánchez-Chinchillas, A.; Pedraza-Chaverri, J. Role of food-derived antioxidants against cisplatin induced-nephrotoxicity. Food Chem. Toxicol. 2018, 120, 230–242. [Google Scholar] [CrossRef]
- Saad, S.Y.; Al-Rikabi, A.C. Protection effects of Taurine supplementation against cisplatin-induced nephrotoxicity in rats. Chemotherapy 2002, 48, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Shalby, A.B.; Assaf, N.; Ahmed, H.H. Possible mechanisms for N-acetyl cysteine and taurine in ameliorating acute renal failure induced by cisplatin in rats. Toxicol. Mech. Methods. 2011, 21, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Rovetta, F.; Ferroni, M.; Corsetti, G.; Lavazza, A.; Sberveglieri, G.; Aleo, M.F. Taurine rescues cisplatin-induced muscle atrophy in vitro: A morphological study. Oxid. Med. Cell Longev. 2014, 2014, 840951. [Google Scholar] [CrossRef]
- Chowdhury, S.; Sinha, K.; Banerjee, S.; Sil, P.C. Taurine protects cisplatin induced cardiotoxicity by modulating inflammatory and endoplasmic reticulum stress responses. Biofactors 2016, 42, 647–664. [Google Scholar] [CrossRef]
- Abdel-Wahab, W.M.; Moussa, F.I.; Saad, N.A. Synergistic protective effect of N-acetylcysteine and taurine against cisplatin-induced nephrotoxicity in rats. Drug Des. Devel. Ther. 2017, 11, 901–908. [Google Scholar] [CrossRef]
- Owoeye, O.; Adedara, I.A.; Farombi, E.O. Pretreatment with taurine prevented brain injury and exploratory behaviour associated with administration of anticancer drug cisplatin in rats. Biomed. Pharmacother. 2018, 102, 375–384. [Google Scholar] [CrossRef]
- Han, X.; Chesney, R.W. Regulation of TauT by cisplatin in LLC-PK1 renal cells. Pediatr. Nephrol. 2005, 20, 1067–1072. [Google Scholar] [CrossRef]
- Han, X.; Yue, J.; Chesney, R.W. Functional TauT protects against acute kidney injury. J. Am. Soc. Nephrol. 2009, 20, 1323–1332. [Google Scholar] [CrossRef]
- Baliou, S.; Kyriakopoulos, A.M.; Goulielmaki, M.; Panayiotidis, M.I.; Spandidos, D.A.; Zoumpourlis, V. Significance of taurine transporter (TauT) in homeostasis and its layers of regulation (Review). Mol. Med. Rep. 2020, 22, 2163–2173. [Google Scholar] [CrossRef]
- Tsunekawa, M.; Wang, S.; Kato, T.; Yamashita, T.; Ma, N. Taurine Administration Mitigates Cisplatin Induced Acute Nephrotoxicity by Decreasing DNA Damage and Inflammation: An Immunocytochemical Study. Adv. Exp. Med. Biol. 2017, 975, 703–716. [Google Scholar]
- Kato, T.; Tsunekawa, M.; Wang, S.; Yamashita, T.; Ma, N. Effect of Taurine on iNOS-Mediated DNA Damage in Drug-Induced Renal Injury. Adv. Exp. Med. Biol. 2017, 975, 717–727. [Google Scholar]
- Azab, S.S.; Kamel, I.; Ismail, N.N.; El Din Hosni, H.; El Fatah, M.A. The defensive role of taurine against gonadotoxicity and testicular apoptosis effects induced by cisplatin in rats. J. Infect. Chemother. 2020, 26, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lu, R.; Huang, C.; Lin, D. Taurine Protects C2C12 Myoblasts from Impaired Cell Proliferation and Myotube Differentiation Under Cisplatin-Induced ROS Exposure. Front. Mol. Biosci. 2021, 8, 685362. [Google Scholar] [CrossRef] [PubMed]
- Cappetta, D.; De Angelis, A.; Sapio, L.; Prezioso, L.; Illiano, M.; Quaini, F.; Rossi, F.; Berrino, L.; Naviglio, S.; Urbanek, K. Oxidative Stress and Cellular Response to Doxorubicin: A Common Factor in the Complex Milieu of Anthracycline Cardiotoxicity. Oxid. Med. Cell Longev. 2017, 2017, 1521020. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Gu, J.; Chi, M.; Li, M.; Lei, S.; Wang, G. Activation of PI3K-Akt through taurine is critical for propofol to protect rat cardiomyocytes from doxorubicin-induced toxicity. Can. J. Physiol. Pharmacol. 2014, 92, 155–161. [Google Scholar] [CrossRef]
- Hosseini, A.; Sahebkar, A. Reversal of Doxorubicin-induced Cardiotoxicity by Using Phytotherapy: A Review. J. Pharmacopuncture 2017, 20, 243–256. [Google Scholar]
- Hamaguchi, T.; Azuma, J.; Awata, N.; Ohta, H.; Takihara, K.; Harada, H.; Kishimoto, S.; Sperelakis, N. Reduction of doxorubicin-induced cardiotoxicity in mice by taurine. Res. Commun. Chem. Pathol. Pharmacol. 1988, 59, 21–30. [Google Scholar]
- Ito, T.; Muraoka, S.; Takahashi, K.; Fujio, Y.; Schaffer, S.W.; Azuma, J. Beneficial effect of taurine treatment against doxorubicin-induced cardiotoxicity in mice. Adv. Exp. Med. Biol. 2009, 643, 65–74. [Google Scholar]
- Das, J.; Ghosh, J.; Manna, P.; Sil, P.C. Taurine suppresses doxorubicin-triggered oxidative stress and cardiac apoptosis in rat via up-regulation of PI3-K/Akt and inhibition of p53, p38-JNK. Biochem. Pharmacol. 2011, 81, 891–909. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, E.K.; Hwang, J.W.; Kim, W.S.; Shin, W.B.; Natarajan, S.B.; Moon, S.H.; Jeon, B.T.; Park, P.J. Taurine Attenuates Doxorubicin-Induced Toxicity on B16F10 Cells. Adv. Exp. Med. Biol. 2017, 975, 1179–1190. [Google Scholar]
- Das, J.; Ghosh, J.; Manna, P.; Sil, P.C. Taurine protects rat testes against doxorubicin-induced oxidative stress as well as p53, Fas and caspase 12-mediated apoptosis. Amino Acids 2012, 42, 1839–1855. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mei, X.; Yuan, J.; Lu, W.; Li, B.; Xu, D. Taurine zinc solid dispersions attenuate doxorubicin-induced hepatotoxicity and cardiotoxicity in rats. Toxicol. Appl. Pharmacol. 2015, 289, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Fukuno, S.; Oda, A.; Konishi, H. Protective effects of taurine on doxorubicin-induced acute hepatotoxicity through suppression of oxidative stress and apoptotic responses. Anticancer Drugs 2016, 27, 17–23. [Google Scholar] [CrossRef]
- Kim, Y.S.; Sung, S.H.; Tang, Y.; Choi, E.J.; Choi, Y.J.; Hwang, Y.J.; Park, P.J.; Kim, E.K. Protective Effect of Taurine on Mice with Doxorubicin-induced Acute Kidney Injury. Adv. Exp. Med. Biol. 2017, 975, 1191–1201. [Google Scholar]
- Zhang, H.; Guo, J.; Cui, S.; Zhou, Y. Taurine Has Potential Protective Effects against the Chronic Cardiotoxicity Induced by Doxorubicin in Mice. Biol. Pharm. Bull. 2021, 44, 1732–1737. [Google Scholar] [CrossRef]
- Ito, T.; Fujio, Y.; Schaffer, S.W.; Azuma, J. Involvement of transcriptional factor TonEBP in the regulation of the taurine transporter in the cardiomyocyte. Adv. Exp. Med. Biol. 2009, 643, 523–532. [Google Scholar] [PubMed]
- Han, X.; Chesney, R.W. Knockdown of TauT expression impairs human embryonic kidney 293 cell development. Adv. Exp. Med. Biol. 2013, 776, 307–320. [Google Scholar]
- Damasceno, D.C.; Netto, A.O.; Iessi, I.L.; Gallego, F.Q.; Corvino, S.B.; Dallaqua, B.; Sinzato, Y.K.; Bueno, A.; Calderon, I.M.; Rudge, M.V. Streptozotocin-induced diabetes models: Pathophysiological mechanisms and fetal outcomes. Biomed. Res. Int. 2014, 2014, 819065. [Google Scholar] [CrossRef]
- Parihar, P.; Shetty, R.; Ghafourifar, P.; Parihar, M.S. Increase in oxidative stress and mitochondrial impairment in hypothalamus of streptozotocin treated diabetic rat: Antioxidative effect of Withania somnifera. Cell Mol. Biol. 2016, 62, 73–83. [Google Scholar]
- Surai, P.F. Natural Antioxidants in Avian Nutrition and Reproduction; Nottingham University Press: Nottingham, UK, 2002. [Google Scholar]
- Obrosova, I.G.; Fathallah, L.; Stevens, M.J. Taurine counteracts oxidative stress and nerve growth factor deficit in early experimental diabetic neuropathy. Exp. Neurol. 2001, 172, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Javed, H.; Khan, A.; Vaibhav, K.; Moshahid Khan, M.; Ahmad, A.; Ejaz Ahmad, M.; Ahmad, A.; Tabassum, R.; Islam, F.; Safhi, M.M.; et al. Taurine ameliorates neurobehavioral, neurochemical and immunohistochemical changes in sporadic dementia of Alzheimer’s type (SDAT) caused by intracerebroventricular streptozotocin in rats. Neurol. Sci. 2013, 34, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.J.; Pandya, K.; Lau-Cam, C.A. The Effect of Metformin and Taurine, Alone and in Combination, on the Oxidative Stress Caused by Diabetes in the Rat Brain. Adv. Exp. Med. Biol. 2017, 975, 353–369. [Google Scholar]
- Reeta, K.H.; Singh, D.; Gupta, Y.K. Chronic treatment with taurine after intracerebroventricular streptozotocin injection improves cognitive dysfunction in rats by modulating oxidative stress, cholinergic functions and neuroinflammation. Neurochem. Int. 2017, 108, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Caletti, G.; Herrmann, A.P.; Pulcinelli, R.R.; Steffens, L.; Morás, A.M.; Vianna, P.; Chies, J.A.B.; Moura, D.J.; Barros, H.M.T.; Gomez, R. Taurine counteracts the neurotoxic effects of streptozotocin-induced diabetes in rats. Amino Acids 2018, 50, 95–104. [Google Scholar] [CrossRef]
- Tsounapi, P.; Saito, M.; Dimitriadis, F.; Koukos, S.; Shimizu, S.; Satoh, K.; Takenaka, A.; Sofikitis, N. Antioxidant treatment with edaravone or taurine ameliorates diabetes-induced testicular dysfunction in the rat. Mol. Cell Biochem. 2012, 369, 195–204. [Google Scholar] [CrossRef]
- Abd El-Twab, S.M.; Mohamed, H.M.; Mahmoud, A.M. Taurine and pioglitazone attenuate diabetes-induced testicular damage by abrogation of oxidative stress and up-regulation of the pituitary-gonadal axis. Can J. Physiol. Pharmacol. 2016, 94, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chowdhury, S.; Das, A.K.; Sil, P.C. Taurine ameliorates oxidative stress induced inflammation and ER stress mediated testicular damage in STZ-induced diabetic Wistar rats. Food Chem. Toxicol. 2019, 124, 64–80. [Google Scholar] [CrossRef]
- Liu, H.; Lin, S.; Lv, Q.; Yang, Q.; Wu, G.; Hu, J.; Yang, J. Taurine Recovers Testicular Steroidogenesis and Spermatogenesis in Streptozotocin-Induced Diabetic Rats. Adv. Exp. Med. Biol. 2017, 975, 801–811. [Google Scholar]
- Pandya, K.; Clark, G.J.; Lau-Cam, C.A. Investigation of the Role of a Supplementation with Taurine on the Effects of Hypoglycemic-Hypotensive Therapy against Diabetes-Induced Nephrotoxicity in Rats. Adv. Exp. Med. Biol. 2017, 975, 371–400. [Google Scholar]
- Shivananjappa, M.M.; Muralidhara. Taurine attenuates maternal and embryonic oxidative stress in a streptozotocin-diabetic rat model. Reprod. Biomed. Online 2012, 24, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Mathew, E.; Barletta, M.A.; Lau-Cam, C.A. The effects of taurine and thiotaurine on oxidative stress in the aorta and heart of diabetic rats. Adv. Exp. Med. Biol. 2013, 775, 345–369. [Google Scholar] [PubMed]
- Budhram, R.; Pandya, K.G.; Lau-Cam, C.A. Protection by taurine and thiotaurine against biochemical and cellular alterations induced by diabetes in a rat model. Adv. Exp. Med. Biol. 2013, 775, 321–343. [Google Scholar]
- Patel, S.N.; Lau-Cam, C.A. The Effect of Taurine and Its Immediate Homologs on Diabetes-Induced Oxidative Stress in the Brain and Spinal Cord of Rats. Adv. Exp. Med. Biol. 2017, 975, 337–351. [Google Scholar]
- Miranda, G.H.N.; Gomes, B.A.Q.; Bittencourt, L.O.; Aragão, W.A.B.; Nogueira, L.S.; Dionizio, A.S.; Buzalaf, M.A.R.; Monteiro, M.C.; Lima, R.R. Chronic Exposure to Sodium Fluoride Triggers Oxidative Biochemistry Misbalance in Mice: Effects on Peripheral Blood Circulation. Oxid. Med. Cell Longev. 2018, 2018, 8379123. [Google Scholar] [CrossRef] [PubMed]
- da Silva Pereira, H.A.B.; Dionizio, A.S.; Araujo, T.T.; Fernandes, M.D.S.; Iano, F.G.; Buzalaf, M.A.R. Proposed mechanism for understanding the dose- and time-dependency of the effects of fluoride in the liver. Toxicol. Appl. Pharmacol. 2018, 358, 68–75. [Google Scholar] [CrossRef]
- Das, J.; Ghosh, J.; Manna, P.; Sil, P.C. Taurine provides antioxidant defense against NaF-induced cytotoxicity in murine hepatocytes. Pathophysiology 2008, 15, 181–190. [Google Scholar] [CrossRef]
- Adedara, I.A.; Ojuade, T.J.D.; Olabiyi, B.F.; Idris, U.F.; Onibiyo, E.M.; Ajeigbe, O.F.; Farombi, E.O. Taurine Ameliorates Renal Oxidative Damage and Thyroid Dysfunction in Rats Chronically Exposed to Fluoride. Biol. Trace Elem. Res. 2017, 175, 388–395. [Google Scholar] [CrossRef]
- Adedara, I.A.; Abolaji, A.O.; Idris, U.F.; Olabiyi, B.F.; Onibiyo, E.M.; Ojuade, T.D.; Farombi, E.O. Neuroprotective influence of taurine on fluoride-induced biochemical and behavioral deficits in rats. Chem. Biol. Interact. 2017, 261, 1–10. [Google Scholar] [CrossRef]
- Adedara, I.A.; Olabiyi, B.F.; Ojuade, T.D.; Idris, U.F.; Onibiyo, E.M.; Farombi, E.O. Taurine reverses sodium fluoride-mediated increase in inflammation, caspase-3 activity, and oxidative damage along the brain-pituitary-gonadal axis in male rats. Can. J. Physiol. Pharmacol. 2017, 95, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Sevgiler, Y.; Karaytug, S.; Karayakar, F. Antioxidative effects of N-acetylcysteine, lipoic acid, taurine, and curcumin in the muscle of Cyprinus carpio L. exposed to cadmium. Arh. Hig. Rada Toksikol. 2011, 62, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Manna, P.; Sil, P.C. Induction of necrosis in cadmium-induced hepatic oxidative stress and its prevention by the prophylactic properties of taurine. J. Trace Elem. Med. Biol. 2009, 23, 300–313. [Google Scholar] [CrossRef]
- Sinha, M.; Manna, P.; Sil, P.C. Taurine protects the antioxidant defense system in the erythrocytes of cadmium treated mice. BMB Rep. 2008, 41, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Manna, P.; Sil, P.C. Cadmium-induced neurological disorders: Prophylactic role of taurine. J. Appl. Toxicol. 2008, 28, 974–986. [Google Scholar] [CrossRef]
- Manna, P.; Sinha, M.; Sil, P.C. Cadmium induced testicular pathophysiology: Prophylactic role of taurine. Reprod. Toxicol. 2008, 26, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Jagadeesan, G.; Sankarsami Pillai, S. Hepatoprotective effects of taurine against mercury induced toxicity in rats. J. Environ. Biol. 2007, 28, 753–756. [Google Scholar]
- Boşgelmez, I.İ.; Güvendik, G. Beneficial Effects of N-Acetyl-L-cysteine or Taurine Pre- or Post-treatments in the Heart, Spleen, Lung, and Testis of Hexavalent Chromium-Exposed Mice. Biol. Trace Elem. Res. 2019, 190, 437–445. [Google Scholar] [CrossRef]
- Song, X.; Kenston, S.S.F.; Kong, L.; Zhao, J. Molecular mechanisms of nickel induced neurotoxicity and chemoprevention. Toxicology 2017, 392, 47–54. [Google Scholar] [CrossRef]
- Qiao, M.; Liu, P.; Ren, X.; Feng, T.; Zhang, Z. Potential protection of taurine on antioxidant system and ATPase in brain and blood of rats exposed to aluminum. Biotechnol. Lett. 2015, 37, 1579–1584. [Google Scholar] [CrossRef]
- Al Kahtani, M.A.; Abdel-Moneim, A.M.; El-Sayed, W.M. The influence of taurine pretreatment on aluminum chloride induced nephrotoxicity in Swiss albino mice. Histol. Histopathol. 2014, 29, 45–55. [Google Scholar] [PubMed]
- Gürer, H.; Ozgünes, H.; Saygin, E.; Ercal, N. Antioxidant effect of taurine against lead-induced oxidative stress. Arch. Environ. Contam. Toxicol. 2001, 41, 397–402. [Google Scholar] [CrossRef]
- Akande, M.G.; Shittu, M.; Uchendu, C.; Yaqub, L.S. Taurine ameliorated thyroid function in rats co-administered with chlorpyrifos and lead. Vet. Res. Commun. 2016, 40, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Oudit, G.Y.; Trivieri, M.G.; Khaper, N.; Husain, T.; Wilson, G.J.; Liu, P.; Sole, M.J.; Backx, P.H. Taurine supplementation reduces oxidative stress and improves cardiovascular function in an iron-overload murine model. Circulation 2004, 109, 1877–1885. [Google Scholar] [CrossRef]
- Cetiner, M.; Sener, G.; Sehirli, A.O.; Ekşioğlu-Demiralp, E.; Ercan, F.; Sirvanci, S.; Gedik, N.; Akpulat, S.; Tecimer, T.; Yeğen, B.C. Taurine protects against methotrexate-induced toxicity and inhibits leukocyte death. Toxicol. Appl. Pharmacol. 2005, 209, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, H.; Rehman, H.; Banerjee, B.D.; Raisuddin, S.; Parvez, S. Attenuation of tamoxifen-induced hepatotoxicity by taurine in mice. Clin. Chim. Acta 2006, 370, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.S.; Hafiz, N.A.; Abd El-Rahim, A.H. Protective role of taurine against genotoxic damage in mice treated with methotrexate and tamoxfine. Environ. Toxicol. Pharmacol. 2011, 31, 143–152. [Google Scholar] [CrossRef]
- Yousef, H.N.; Aboelwafa, H.R. The potential protective role of taurine against 5-fluorouracil-induced nephrotoxicity in adult male rats. Exp. Toxicol. Pathol. 2017, 69, 265–274. [Google Scholar] [CrossRef]
- Alhumaidha, K.A.; Saleh, D.O.; Abd El Fattah, M.A.; El-Eraky, W.I.; Moawad, H. Cardiorenal protective effect of taurine against cyclophosphamide-induced toxicity in albino rats. Can. J. Physiol. Pharmacol. 2016, 94, 131–139. [Google Scholar] [CrossRef]
- Hagar, H.H. The protective effect of taurine against cyclosporine A-induced oxidative stress and hepatotoxicity in rats. Toxicol. Lett. 2004, 151, 335–343. [Google Scholar] [CrossRef]
- Vanitha, M.K.; Priya, K.D.; Baskaran, K.; Periyasamy, K.; Saravanan, D.; Venkateswari, R.; Mani, B.R.; Ilakkia, A.; Selvaraj, S.; Menaka, R.; et al. Taurine Regulates Mitochondrial Function During 7,12-Dimethyl Benz[a]anthracene Induced Experimental Mammary Carcinogenesis. J. Pharmacopunct. 2015, 18, 68–74. [Google Scholar] [CrossRef]
- Ozan, G.; Turkozkan, N.; Bircan, F.S.; Balabanli, B. Effect of taurine on brain 8-hydroxydeoxyguanosine and 3-nitrotyrosine levels in endotoxemia. Inflammation 2012, 35, 665–670. [Google Scholar] [CrossRef]
- Jeon, S.H.; Lee, M.Y.; Rahman, M.M.; Kim, S.J.; Kim, G.B.; Park, S.Y.; Hong, C.U.; Kim, S.Z.; Kim, J.S.; Kang, H.S. The antioxidant, taurine reduced lipopolysaccharide (LPS)-induced generation of ROS, and activation of MAPKs and Bax in cultured pneumocytes. Pulm. Pharmacol. Ther. 2009, 22, 562–566. [Google Scholar] [CrossRef]
- Heidari, R.; Babaei, H.; Eghbal, M.A. Amodiaquine-induced toxicity in isolated rat hepatocytes and the cytoprotective effects of taurine and/or N-acetyl cysteine. Res. Pharm. Sci. 2014, 9, 97–105. [Google Scholar] [PubMed]
- Heidari, R.; Rasti, M.; Shirazi Yeganeh, B.; Niknahad, H.; Saeedi, A.; Najibi, A. Sulfasalazine-induced renal and hepatic injury in rats and the protective role of taurine. Bioimpacts 2016, 6, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Heidari, R.; Jamshidzadeh, A.; Keshavarz, N.; Azarpira, N. Mitigation of Methimazole-Induced Hepatic Injury by Taurine in Mice. Sci. Pharm. 2014, 83, 143–158. [Google Scholar] [CrossRef]
- Sener, G.; Ozer Sehirli, A.; Ipci, Y.; Cetinel, S.; Cikler, E.; Gedik, N.; Alican, I. Taurine treatment protects against chronic nicotine-induced oxidative changes. Fundam. Clin. Pharmacol. 2005, 19, 155–164. [Google Scholar] [CrossRef]
- Yang, Q.; Yang, J.; Wu, G.; Feng, Y.; Lv, Q.; Lin, S.; Hu, J. Effects of taurine on myocardial cGMP/cAMP ratio, antioxidant ability, and ultrastructure in cardiac hypertrophy rats induced by isoproterenol. Adv. Exp. Med. Biol. 2013, 776, 217–229. [Google Scholar] [PubMed]
- Motawi, T.K.; Abd Elgawad, H.M.; Shahin, N.N. Modulation of indomethacin-induced gastric injury by spermine and taurine in rats. J. Biochem. Mol. Toxicol. 2007, 21, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Erdem, A.; Gündoğan, N.U.; Usubütün, A.; Kilinç, K.; Erdem, S.R.; Kara, A.; Bozkurt, A. The protective effect of taurine against gentamicin-induced acute tubular necrosis in rats. Nephrol. Dial. Transplant. 2000, 15, 1175–1182. [Google Scholar] [CrossRef]
- Shao, X.; Hu, Z.; Hu, C.; Bu, Q.; Yan, G.; Deng, P.; Lv, L.; Wu, D.; Deng, Y.; Zhao, J.; et al. Taurine protects methamphetamine-induced developmental angiogenesis defect through antioxidant mechanism. Toxicol. Appl. Pharmacol. 2012, 260, 260–270. [Google Scholar] [CrossRef]
- Ahmed, M.A. Amelioration of nandrolone decanoate-induced testicular and sperm toxicity in rats by taurine: Effects on steroidogenesis, redox and inflammatory cascades, and intrinsic apoptotic pathway. Toxicol. Appl. Pharmacol. 2015, 282, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Acharya, M.; Lau-Cam, C.A. Comparative evaluation of the effects of taurine and thiotaurine on alterations of the cellular redox status and activities of antioxidant and glutathione-related enzymes by acetaminophen in the rat. Adv. Exp. Med. Biol. 2013, 776, 199–215. [Google Scholar] [PubMed]
- Balasubramanian, T.; Somasundaram, M.; Felix, A.J. Taurine prevents Ibuprofen-induced gastric mucosal lesions and influences endogenous antioxidant status of stomach in rats. Sci. World J. 2004, 4, 1046–1054. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhou, J.; Li, Y.; Yan, G.; Bu, Q.; Lv, L.; Yang, Y.; Zhao, J.; Shao, X.; Deng, Y.; Zhu, R.; et al. Protective role of taurine against morphine-induced neurotoxicity in C6 cells via inhibition of oxidative stress. Neurotox. Res. 2011, 20, 334–342. [Google Scholar] [CrossRef]
- Ozden, S.; Catalgol, B.; Gezginci-Oktayoglu, S.; Karatug, A.; Bolkent, S.; Alpertunga, B. Acute effects of methiocarb on oxidative damage and the protective effects of vitamin E and taurine in the liver and kidney of Wistar rats. Toxicol. Ind. Health. 2013, 29, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Das, T.; Paramanik, A.; Choudhury, S.M. Taurine improves lambda cyhalothrin induced biochemical alterations in Wistar rat liver. Int. J. Bioassays 2016, 5, 5059–5065. [Google Scholar] [CrossRef][Green Version]
- Turkez, H.; Aydin, E. The effects of taurine on permethrininduced cytogenetic and oxidative damage in cultured human lymphocytes. Arh. Hig. Rada. Toksikol. 2012, 63, 27–34. [Google Scholar] [CrossRef]
- Aly, H.A.; Khafagy, R.M. Taurine reverses endosulfan-induced oxidative stress and apoptosis in adult rat testis. Food Chem. Toxicol. 2014, 64, 1–9. [Google Scholar] [CrossRef]
- Akande, M.G.; Ahmed, U.S. Taurine abated subacute dichlorvos toxicity. Toxicol. Rep. 2017, 4, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Sill, P.C. Tertiary butyl hydroperoxide induced oxidative damage in mice erythrocytes: Protection by taurine. Pathophysiology 2012, 19, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Kocak-Toker, N.; Giris, M.; Tülübas, F.; Uysal, M.; Aykac-Toker, G. Peroxynitrite induced decrease in Na+, K+-ATPase activity is restored by taurine. World J. Gastroenterol. 2005, 11, 3554–3557. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Cha, Y.N.; Kim, C. Taurine chloramine protects RAW 264.7 macrophages against hydrogen peroxide-induced apoptosis by increasing antioxidants. J. Clin. Biochem. Nutr. 2011, 49, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Arancibia, S.; Dorado-Martínez, C.; Borgonio-Pérez, G.; Hiriart-Urdanivia, M.; Verdugo-Diaz, L.; Durán-Vázquez, A.; Colin-Baranque, L.; Avila-Costa, M.R. Effects of taurine on ozone-induced memory deficits and lipid peroxidation levels in brains of young, mature, and old rats. Environ. Res. 2000, 82, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Aydın, A.F.; Çoban, J.; Doğan-Ekici, I.; Betül-Kalaz, E.; Doğru-Abbasoğlu, S.; Uysal, M. Carnosine and taurine treatments diminished brain oxidative stress and apoptosis in D-galactose aging model. Metab. Brain Dis. 2016, 31, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Nandhini, A.T.; Thirunavukkarasu, V.; Ravichandran, M.K.; Anuradha, C.V. Effect of taurine on biomarkers of oxidative stress in tissues of fructose-fed insulin-resistant rats. Singap. Med. J. 2005, 46, 82–87. [Google Scholar]
- Chang, Y.Y.; Chou, C.H.; Chiu, C.H.; Yang, K.T.; Lin, Y.L.; Weng, W.L.; Chen, Y.C. Preventive effects of taurine on development of hepatic steatosis induced by a high-fat/cholesterol dietary habit. J. Agric. Food Chem. 2011, 59, 450–457. [Google Scholar] [CrossRef]
- Rashid, K.; Das, J.; Sil, P.C. Taurine ameliorate alloxan induced oxidative stress and intrinsic apoptotic pathway in the hepatic tissue of diabetic rats. Food Chem. Toxicol. 2013, 51, 317–329. [Google Scholar] [CrossRef]
- Mahalakshmi, K.; Pushpakiran, G.; Anuradha, C.V. Taurine prevents acrylonitrile-induced oxidative stress in rat brain. Pol. J. Pharmacol. 2003, 55, 1037–1043. [Google Scholar]
- Başaran-Küçükgergin, C.; Bingül, I.; Tekkeşin, M.S.; Olgaç, V.; Doğru-Abbasoğlu, S.; Uysal, M. Effects of carnosine, taurine, and betaine pretreatments on diethylnitrosamine-induced oxidative stress and tissue injury in rat liver. Toxicol. Ind. Health. 2016, 32, 1405–1413. [Google Scholar] [CrossRef]
- Li, Y.; Piao, F.; Liu, X. Protective effect of taurine on triorthocresyl phosphate (TOCP)-induced cytotoxicity in C6 glioma cells. Adv. Exp. Med. Biol. 2013, 776, 231–240. [Google Scholar] [PubMed]
- Tadros, M.G.; Khalifa, A.E.; Abdel-Naim, A.B.; Arafa, H.M. Neuroprotective effect of taurine in 3-nitropropionic acid-induced experimental animal model of Huntington’s disease phenotype. Pharmacol. Biochem. Behav. 2005, 82, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F. Antioxidant systems in poultry biology: Superoxide dismutase. J. Anim. Res. Nutr. 2016, 1, 8. [Google Scholar] [CrossRef]
- Surai, P.F. Selenium in Poultry Nutrition and Health; Wageningen Academic Publishers: Wageningen, The Netherlands, 2018. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I. Glutathione peroxidases in poultry biology: Part 1. Classification and mechanisms of action. World Poult. Sci. J. 2018, 74, 185–198. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I. Glutathione peroxidases in poultry biology: Part 2. Modulation of enzymatic activities. World Poult. Sci. J. 2018, 74, 239–250. [Google Scholar] [CrossRef]
- Kim, Y.; Cha, S.J.; Choi, H.J.; Kim, K. Omega Class Glutathione S-Transferase: Antioxidant Enzyme in Pathogenesis of Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2017, 2017, 5049532. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell Longev. 2019, 2019, 961309. [Google Scholar] [CrossRef]
- Han, H.L.; Zhang, J.F.; Yan, E.F.; Shen, M.M.; Wu, J.M.; Gan, Z.D.; Wei, C.H.; Zhang, L.L.; Wang, T. Effects of taurine on growth performance, antioxidant capacity, and lipid metabolism in broiler chickens. Poult. Sci. 2020, 99, 5707–5717. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, L.; Li, J.; Xing, T.; Jiang, Y.; Gao, F. Dietary taurine supplementation ameliorates muscle loss in chronic heat stressed broilers via suppressing the perk signaling and reversing endoplasmic reticulum-stress-induced apoptosis. J. Sci. Food Agric. 2021, 101, 2125–2134. [Google Scholar] [CrossRef]
- Huang, R.S.; Peng, Z.L. Effect of diet type and dietary taurine supplementation on growth performance of weaning pigs. Cereal Feed Ind. 2008, 9, 44–45. [Google Scholar]
- Tang, Z.; Liu, J.; Sun, Z.; Li, J.; Sun, W.; Mao, J.; Wang, Y. Protective effects of taurine on growth performance and intestinal epithelial barrier function in weaned piglets challenged without or with lipopolysaccharide. Anim. Prod. Sci. 2018, 58, 2011–2020. [Google Scholar] [CrossRef]
- Xu, M.; Che, L.; Gao, K.; Wang, L.; Yang, X.; Wen, X.; Jiang, Z.; Wu, D. Effects of Dietary Taurine Supplementation to Gilts during Late Gestation and Lactation on Offspring Growth and Oxidative Stress. Animals 2019, 9, 220. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Guo, Q.; Wang, W.; Duan, Y.; Zhang, L.; Li, J.; He, S.; Chen, W.; Li, F. Taurine Alleviates Intestinal Injury by Mediating Tight Junction Barriers in Diquat-Challenged Piglet Models. Front. Physiol. 2020, 11, 449. [Google Scholar] [CrossRef] [PubMed]


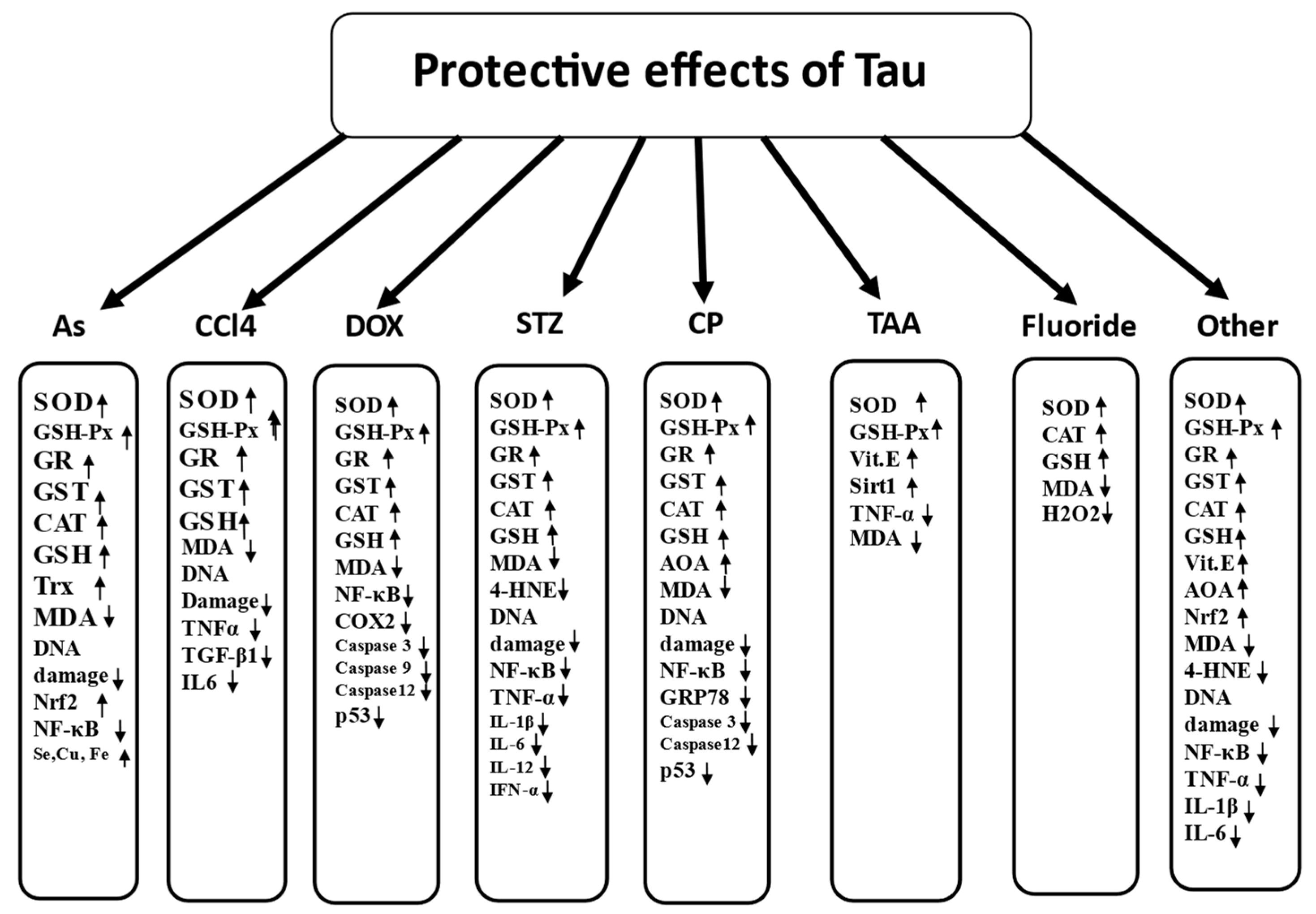{kind=link}
| Function | Recent References |
|---|---|
| Antioxidant | [1,17,52,53,54] |
| Membrane stabilisation | [55,56,57] |
| Mitochondrial integrity maintenance | [11,13,58,59,60,61] |
| Vitagene activation | [1,62,63,64,65] |
| Bile acid conjugation | [66,67,68,69,70] |
| Ca homeostasis | [13,71,72,73] |
| FA metabolism/oxidation | [74,75,76] |
| Energy metabolism | [77,78,79] |
| Osmoregulation | [13,80,81] |
| Thermoregulation | [82,83,84] |
| Detoxification | [85,86,87] |
| Neuroprotection | [71,88,89,90,91] |
| Anti-inflammatory | [11,92,93,94,95] |
| Immunomodulation | [46,96,97,98,99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surai, P.F.; Earle-Payne, K.; Kidd, M.T. Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models. Antioxidants 2021, 10, 1876. https://doi.org/10.3390/antiox10121876
Surai PF, Earle-Payne K, Kidd MT. Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models. Antioxidants. 2021; 10(12):1876. https://doi.org/10.3390/antiox10121876
Chicago/Turabian StyleSurai, Peter F., Katie Earle-Payne, and Michael T. Kidd. 2021. "Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models" Antioxidants 10, no. 12: 1876. https://doi.org/10.3390/antiox10121876
APA StyleSurai, P. F., Earle-Payne, K., & Kidd, M. T. (2021). Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models. Antioxidants, 10(12), 1876. https://doi.org/10.3390/antiox10121876