Typical and Aberrant Functional Brain Flexibility: Lifespan Development and Aberrant Organization in Traumatic Brain Injury and Dyslexia

 ,
,  ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Participants
2.2. Data Recording and Preprocessing
2.3. Dynamic Functional Connectivity
2.3.1. Intra and Inter-Frequency Coupling Estimators and Statistical Filtering
2.3.2. Amplitude Envelope Correlation (AEC)
2.3.3. Phase-to-Amplitude Cross-Frequency Coupling (Cross-Frequency iPLV)
2.3.4. Intra-Frequency Phase-to-Phase Coupling (Same-Frequency iPLV)
2.3.5. Cross-Frequency Interactions via Delay Symbolic Transfer Entropy (dSTE)
2.3.6. Phase interactions: Directed Phase Lag Index (dPLI)
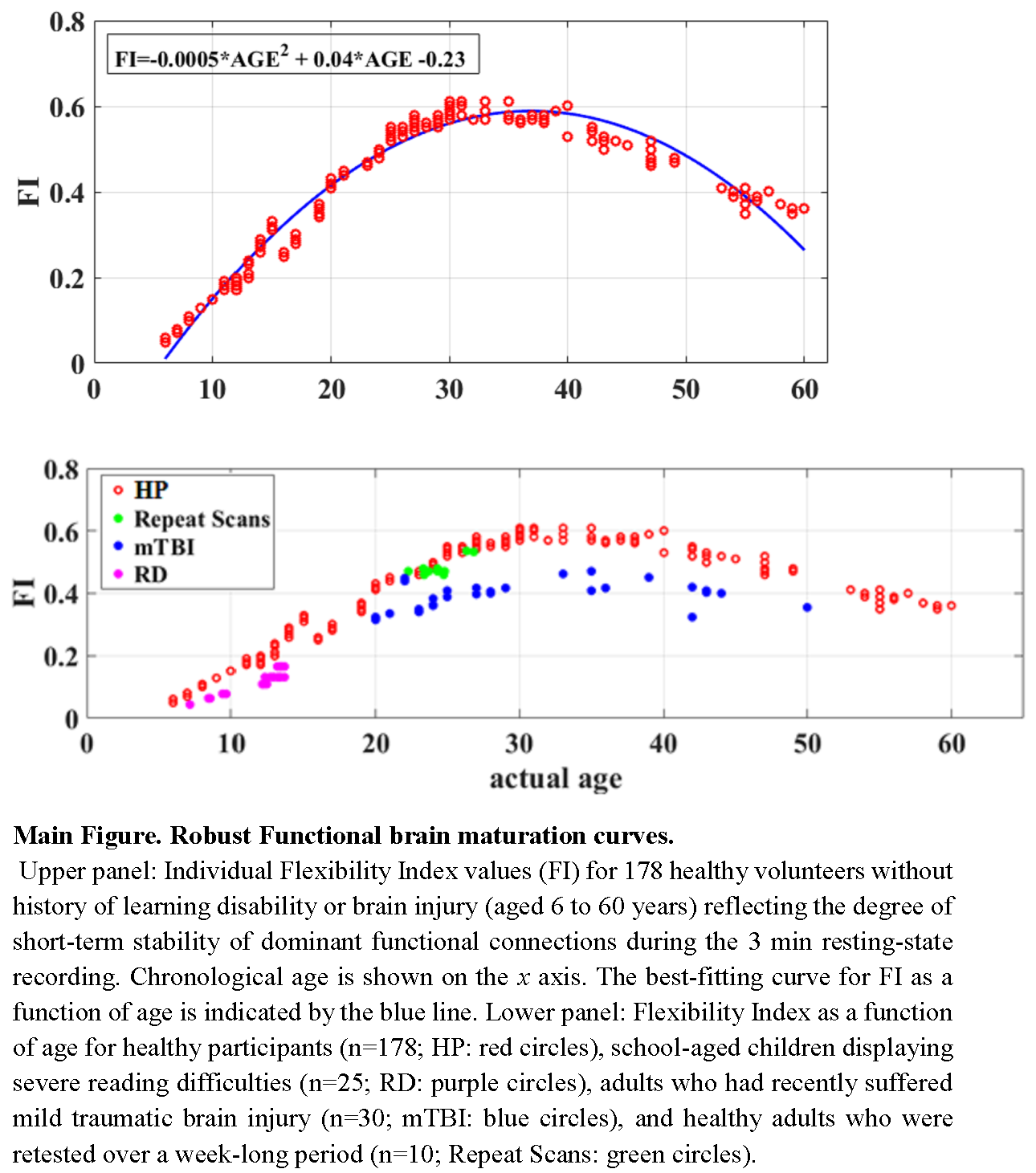
2.3.7. Identifying the Dominant Intrinsic Coupling Mode (dICM) for A Given Pair of Sensors
2.3.8. Topological Filtering based on Orthogonal Minimal Spanning Tress (OMSTs)
2.3.9. Identifying the Dominant Type of Inter- and Intra-Hemispheric Interactions for Groups of Neighbouring Sensors
2.3.10. Flexibility Index (FI) Based on Dominant Intrinsic Coupling Modes
2.4. Modelling Participant Age through Individual FI values
2.5. Deriving Age-Related Neuromagnetic Features
2.6. Software for Analyses
3. Results
3.1. Age-Related Differences in dominant Intrinsic Coupling Modes (dICMs)
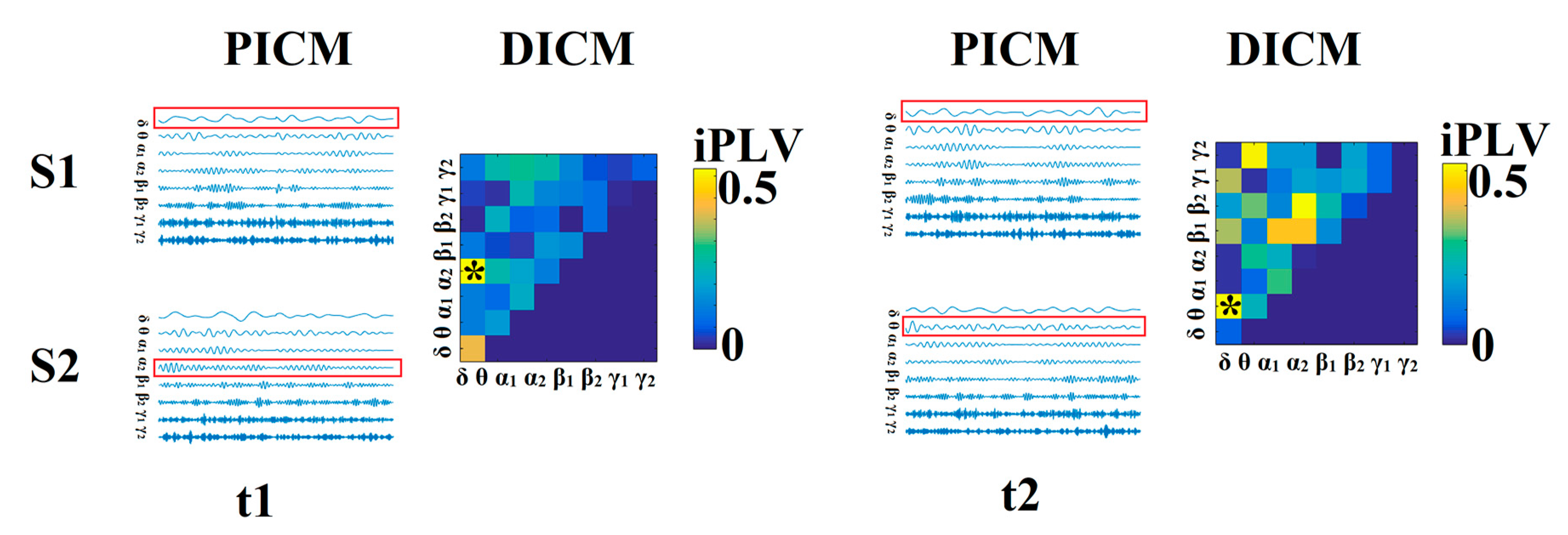
3.2. Identifying Age-Related Neuromagnetic Features
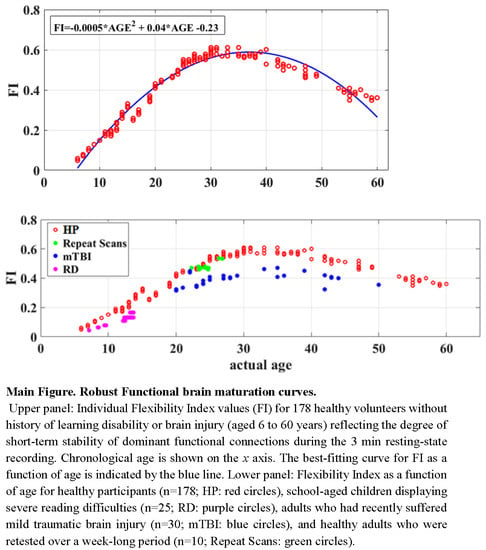
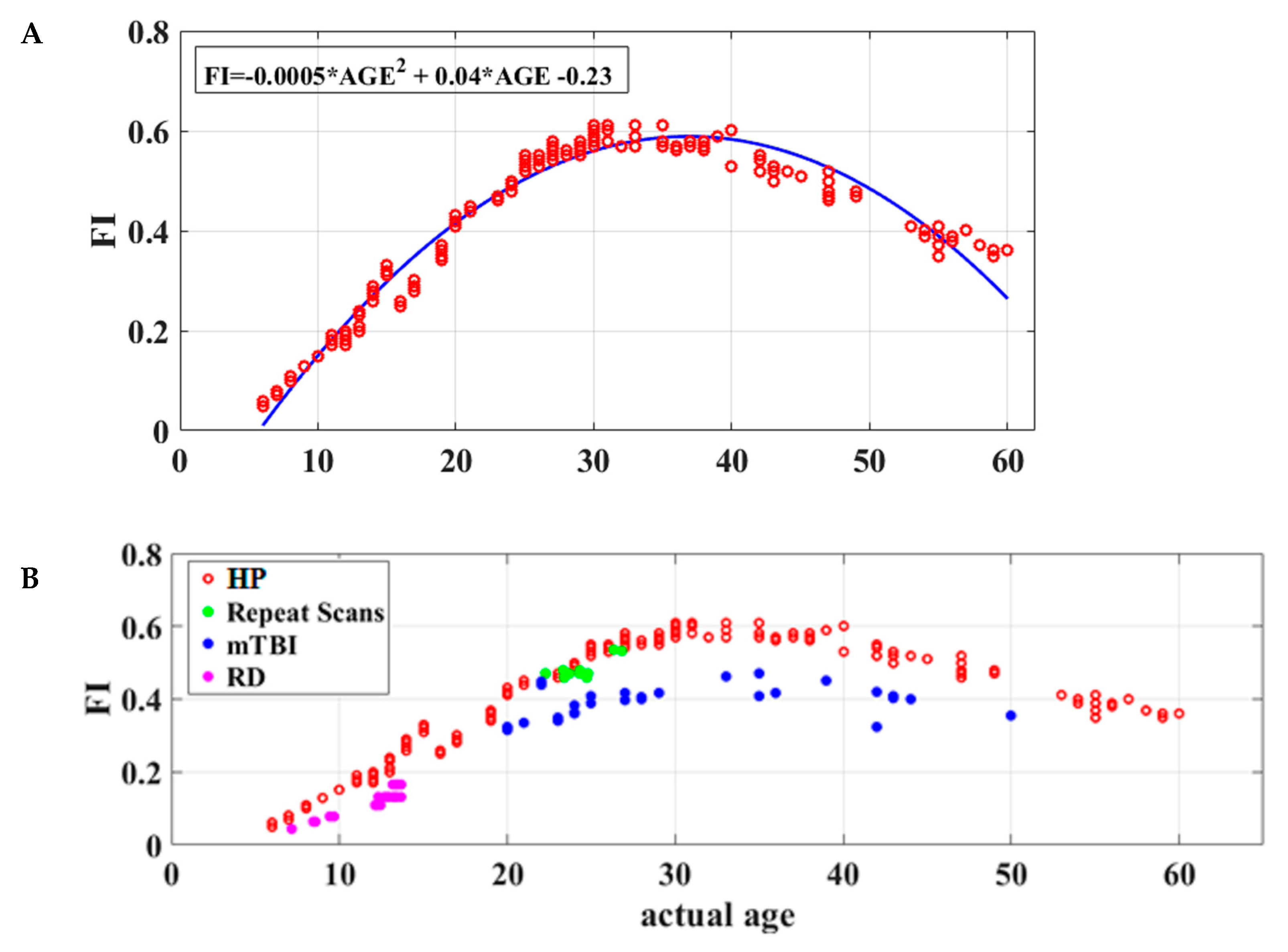
3.3. Maturation Patterns
3.4. Reliability and Clinical Validity of the Flexibility Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kessler, D.; Angstadt, M.; Sripada, C. Growth charting of brain connectivity networks and the identification of attention impairment in youth. JAMA Psychiatry 2016, 73, 481–489. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W. Oscillations and neuronal dynamics in schizophrenia: The search for basic symptoms and translational opportunities. Biol. Psychiat. 2015, 77, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.K.; Fries, P.; Singer, W. Dynamic predictions: Oscillations and synchrony in top-down processing. Nat. Rev. Neurosci. 2001, 2, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Friston, K. A theory of cortical responses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 815–836. [Google Scholar] [CrossRef]
- Gautama, T.; Mandic, D.P.; Van Hulle, M.M. The delay vector variance method for detecting determinism and nonlinearity in time series. Phys. D Nonlinear Phenom. 2004, 190, 167–176. [Google Scholar] [CrossRef]
- Arnal, L.H.; Giraud, A.L. Cortical oscillations and sensory predictions. Trends Cogn. Sci. 2012, 16, 390–398. [Google Scholar] [CrossRef]
- Steriade, M.; Amzica, F.; Contreras, D. Synchronization of fast (30–40 Hz) spontaneous cortical rhythms during brain activation. J. Neurosci. 1996, 16, 392–417. [Google Scholar] [CrossRef] [PubMed]
- Contreras, D.; Destexhe, A.; Sejnowski, T.J.; Steriade, M. Control of spatiotemporal coherence of a thalamic oscillation by corticothalamic feedback. Science 1996, 274, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Destexhe, A.; Contreras, D.; Steriade, M. Cortically-induced coherence of a thalamic-generated oscillation. Neuroscience 1999, 92, 427–444. [Google Scholar] [CrossRef]
- Dosenbach, N.U.; Nardos, B.; Cohen, A.L.; Fair, D.A.; Power, J.D.; Church, J.A.; Nelson, S.M.; Wig, G.S.; Vogel, A.C.; Lessov-Schlaggar, C.N.; et al. Prediction of individual brain maturity using fMRI. Science 2010, 329, 1358–1361. [Google Scholar] [CrossRef] [PubMed]
- Fransson, P.; den, U.; Blennow, M.; Lagercrantz, H. The functional architecture of the infant brain as revealed by resting-state fMRI. Cereb. Cortex 2011, 21, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Chen, S.G.; Hu, D.; Zeng, L.L.; Fan, Y.M.; Chen, X.P.; Shen, H. Predicting individual brain maturity using dynamic functional connectivity. Front. Hum. Neurosci. 2015, 9, 418. [Google Scholar] [CrossRef] [PubMed]
- Betzel, R.F.; Byrge, L.; He, Y.; Goñi, J.; Zuo, X.-N.; Sporns, O. Changes in structural and functional connectivity among resting-state networks across the human life span. Neuroimage 2014, 102, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Vergun, S.; Deshpande, A.S.; Meier, T.B.; Song, J.; Tudorascu, D.L.; Nair, V.A.; Singh, V.; Biswal, B.B.; Meyerand, M.E.; Birn, R.M.; et al. Characterizing functional connectivity differences in aging adults using machine learning on resting state fMRI data. Front. Comput. Neurosci. 2013, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Fair, D.A.; Cohen, A.L.; Power, J.D.; Dosenbach, N.U.; Church, J.A.; Miezin, F.M.; Schlaggar, B.L.; Petersen, S.E. Functional brain networks develop from a “local to distributed” organization. PLoS Comput. Biol. 2009, 5, e1000381. [Google Scholar] [CrossRef]
- Engel, A.K.; Gerloff, C.; Hilgetag, C.C.; Nolte, G. Intrinsic coupling modes: Multiscale interactions in ongoing brain activity. Neuron 2013, 80, 867–886. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, C.B.; Morgan, B.R.; Ye, A.X.; Taylor, M.J.; Doesburg, S.M. Oscillations, networks, and their development: MEG connectivity changes with age. Hum. Brain Mapp. 2014, 35, 5249–5261. [Google Scholar] [CrossRef]
- Cabral, J.; Luckhoo, H.; Woolrich, M.; Joensson, M.; Mohseni, H.; Baker, A.; Kringelbach, M.L.; Deco, G. Exploring mechanisms of spontaneous functional connectivity in MEG: How delayed network interactions lead to structured amplitude envelopes of band-pass filtered oscillations. Neuroimage 2004, 90, 423–435. [Google Scholar] [CrossRef]
- Canolty, R.T.; Edwards, E.; Dalal, S.S.; Soltani, M.; Nagarajan, S.S.; Kirsch, H.E. High gamma power is phase-locked to theta oscillations in human neocortex. Science 2006, 313, 1626–1628. [Google Scholar] [CrossRef]
- Antonakakis, M.; Dimitriadis, S.I.; Zervakis, M.; Micheloyannis, S.; Rezaie, R.; Babajani-Feremi, A.; Zouridakis, G.; Papanicolaou, A.C. Altered cross-frequency coupling in resting-state MEG after mild traumatic brain injury. Int. J. Psychophysiol. 2016, 102, 1–11. [Google Scholar] [CrossRef]
- Dimitriadis, S.I.; Laskaris, N.A.; Simos, P.G.; Fletcher, J.M.; Papanicolaou, A.C. Greater Repertoire and Temporal Variability of Cross-Frequency Coupling (CFC) Modes in Resting-State Neuromagnetic Recordings among Children with Reading Difficulties. Front. Hum. Neurosci. 2016, 10, 163. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, W.; Liu, Z.; Zhang, K.; Lei, X.; Yao, Y.; Becker, B.; Liu, Y.; Kendrick, K.M.; Lu, G.; et al. Neural, electrophysiological and anatomical basis of brain-network variability and its characteristic changes in mental disorders. Brain 2016, 139, 2307–2321. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Wymbs, N.F.; Porter, M.A.; Mucha, P.J.; Carlson, J.M.; Grafton, S.T. Dynamic reconfiguration of human brain networks during learning. Proc. Natl. Acad. Sci. USA 2011, 108, 7641–7646. [Google Scholar] [CrossRef]
- Dimitriadis, S.I.; Tarnanas, I.; Wiederholdg, M.; Wiederholdh, B.; Tsolaki, M.; Fleische, E. Mnemonic strategy training of the elderly at risk for dementia enhances integration of information processing via cross-frequency coupling. Alzheimer’s Dement. 2016, 2, 241–249. [Google Scholar] [CrossRef]
- Dimitriadis, S.I.; Sun, Y.; Thakor, N.V.; Bezerianos, A. Causal interactions between frontalθ–parieto-occipitalα2 predict performance on a mental arithmetic task. Front. Hum. Neurosci. 2016, 10, 454. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, S.I.; Sallis, C.; Tarnanas, I.; Linden, D.E. Topological Filtering of Dynamic Functional Brain Networks Unfolds Informative Chronnectomics: A novel data-driven thresholding scheme based on Orthogonal Minimal Spanning Trees (OMSTs). Front. Neuroinform. 2017, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, S.I.; Antonakakis, M.; Simos, P.G.; Fletcher, J.; Paanicolaou, A. Data-driven topological filtering based on orthogonal minimal spanning trees: Application to multi-group MEG resting-state connectivity. Brain Connect. 2017, 7, 661–670. [Google Scholar] [CrossRef]
- Dimitriadis, S.I.; Salis, C.I. Mining Time-Resolved Functional Brain Graphs to an EEG-Based Chronnectomic Brain Aged Index (CBAI). Front. Hum. Neurosci. 2017, 11, 423. [Google Scholar] [CrossRef]
- Dimitriadis, S.I.; Simos, P.G.; Fletcher, J.Μ.; Papanicolaou, A.C. Aberrant resting-state functional brain networks in dyslexia: Symbolic mutual information analysis of neuromagnetic signals. Int. J. Psychophysiol. 2018, 126, 20–29. [Google Scholar] [CrossRef]
- Dimitriadis, S.I. Complexity of Brain Activity and Connectivity in Functional Neuroimaging. J. Neurosci. Res. 2018, 96, 1741–1757. [Google Scholar] [CrossRef]
- Niso, G.; Rogers, C.; Moreau, J.T.; Chen, L.Y.; Madjar, C.; Das, S.; Bock, E.; Tadel, F.; Evans, A.C.; Jolicoeur, P.; et al. OMEGA: The Open MEG archive. Neuroimage 2016, 124, 1182–1187. [Google Scholar] [CrossRef]
- Theiler, J.; Eubank, S.; Longtin, A.; Galdrikian, B.; Farmer, J.D. Testing for nonlineaity in time series the method of surrogate data. Phys. D 1992, 85, 77–94. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate-a Practical and Powerful Approach to Multiple Testing. J. Royal Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Dimitriadis, S.; Laskaris, N.; Simos, P.; Micheloyannis, S.; Fletcher, J.; Rezaie, R.; Papanicolaou, A. Altered temporal correlations in resting-state connectivity fluctuations in children with reading difficulties detected via MEG. Neuroimage 2013, 83, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Brookes, M.J.; Woolrich, M.W.; Barnes, G.R. Measuring functional connectivity in MEG: A multivariate approach insensitive to linear source leakage. Neuroimage 2012, 63, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G. Neural syntax: Cell assemblies, synapsembles, and readers. Neuron 2010, 68, 362–385. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G.; Logothetis, N.; Singer, W. Scaling brain size, keeping timing: Evolutionary preservation of brain rhythms. Neuron 2013, 80, 751–764. [Google Scholar] [CrossRef]
- Cohen, M.X. Assessing transient cross-frequency coupling in EEG data. J. Neurosci. Methods 2008, 168, 494–499. [Google Scholar] [CrossRef]
- Voytek, B.; Canolty, R.T.; Shestyuk, A.; Crone, N.E.; Parvizi, J.; Knight, R.T. Shifts in gamma phase-amplitude coupling frequency from theta to alpha over posterior cortex during visual tasks. Front. Hum. Neurosci. 2010, 4, 191. [Google Scholar] [CrossRef]
- Lachaux, J.P.; Rodriguez, E.; Martinerie, J.; Varela, F.J. Measuring phase synchrony in brain signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef]
- Nolte, G.; Bai, O.; Wheaton, L.; Mari, Z.; Vorbach, S.; Hallett, M. Identifying true brain interaction from EEG data using the imaginary part of coherency. Clin. Neurophysiol. 2004, 115, 2292–2307. [Google Scholar] [CrossRef] [PubMed]
- Chávez, M.; Martinerie, J.; Le Van Quyen, M. Statistical assessment of nonlinear causality: Application to epileptic EEG signals. J. Neurosci. Methods 2003, 124, 113–128. [Google Scholar] [CrossRef]
- Staniek, M.; Lehnertz, K. Symbolic transfer entropy. Phys. Rev. Lett. 2008, 100, 158101:1–158101:4. [Google Scholar] [CrossRef] [PubMed]
- Martinetz, T.M.; Berkovich, S.G.; Schulten, K.J. Neural-Gas Network for Vector Quantization and Its Application to Time-Series Prediction. IEEE Trans. Neural Networ. 1993, 4, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, S.I.; Kanatsouli, K.; Laskaris, N.A.; Tsirka, V.; Vourkas, M.; Micheloyannis, S. Surface EEG shows that functional segregation via phase coupling contributes to the neural substrate of mental calculations. Brain Cogn. 2012, 80, 45–52. [Google Scholar] [CrossRef]
- Granger, C.W. Investigating causal relations by econometric models and cross-spectral methods. Econometrica 1969, 37, 424–438. [Google Scholar] [CrossRef]
- Verdes, P.F. Assessing causality from multivariate time series. Phys. Rev. E 2005, 72, 026222. [Google Scholar] [CrossRef]
- Wibral, M.; Pampu, N.; Priesemann, V.; Siebenhühner, F.; Seiwert, H.; Lindner, M.; Vicente, R. Measuring information-transfer delays. PLoS ONE 2013, 8, e55809. [Google Scholar] [CrossRef]
- Lizier, J.T.; Heinzle, J.; Horstmann, A.; Haynes, J.-D.; Prokopenko, M. Multivariate information-theoretic measures reveal directed information structure and task relevant changes in fMRI connectivity. J. Comput. Neurosci. 2011, 30, 85–107. [Google Scholar] [CrossRef]
- Vicente, R.; Wibral, M.; Lindner, M.; Pipa, G. Transfer entropy—A model-free measure of effective connectivity for the neurosciences. J. Comput. Neurosci. 2011, 30, 45–67. [Google Scholar] [CrossRef]
- Stam, C.J.; van Straaten, E.C.W. Go with the flow: Use of a directed phase lag index (dPLI) to characterize patterns of phase relations in a large-scale model of brain dynamics. Neuroimage 2012, 62, 1415–1428. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Bullmore, E.T. Human brain networks in health and disease. Curr. Opin. Neurol. 2009, 22, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Meyer-Lindenberg, A.; Achard, S.; Duke, T.; Bullmore, E. Adaptive reconfiguration of fractal small-world human brain functional networks. Proc. Natl. Acad. Sci. USA 2006, 103, 19518–19523. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.G.; Dingwell, J.B. Differential Changes with Age in Multiscale Entropy of Electromyography Signals from Leg Muscles during Treadmill Walking. PLoS ONE 2016, 11, e0162034. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W.J. Developmental Changes in Neuronal Oscillations and Synchrony: Evidence for a Late Critical Period. Proc. Natl. Acad. Sci. USA 2011, 106, 9866–9871. [Google Scholar] [CrossRef]
- Buzsaki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Cobb, S.R.; Buhl, E.H.; Halasy, K.; Paulsen, O.; Somogyi, P. Synchronization of neuronal activity in hippocampus by individual GABAergic interneurons. Nature 1995, 378, 75–78. [Google Scholar] [CrossRef]
- Wang, X.J.; Buzsaki, G. Gamma oscillation by synaptic inhibition in a hippocampal interneuronal network model. J. Neurosci. 1996, 16, 6402–6413. [Google Scholar] [CrossRef]
- Hwang, K.; Velanova, K.; Luna, B. Strengthening of top-down frontal cognitive control networks underlying the development of inhibitory control: A functional magnetic resonance imaging effective connectivity study. J. Neurosci. 2010, 30, 15535–15545. [Google Scholar] [CrossRef]
- Blankenship, T.T.; Bell, M.A. Frontotemporal Coherence and Executive Functions Contribute to Episodic Memory during Middle Childhood. Dev. Neuropsychol. 2015, 40, 430–444. [Google Scholar] [CrossRef]
- Canolty, R.T.; Knight, R.T. The functional role of cross-frequency coupling. Trends Cogn. Sci. 2010, 14, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, S.I.; Zouridakis, G.; Rezaie, R.; Babajani-Feremi, A.; Papanicolaou, A.C. Functional connectivity changes detected with magnetoencephalography after mild traumatic brain injury. NeuroImage Clin. 2015, 9, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Antonakakis, M.; Dimitriadis, S.I.; Papanicolaou, A.C.; Zouridakis, G.; Zervakis, M. Improving the detection complexity analysis in resting–state magnetoencephalography. In Proceedings of the 2016 IEEE International Conference on Imaging Systems and Techniques (IST 2016), Chania, Greece, 4–6 October 2016. [Google Scholar]
- Antonakakis, M.; Dimitriadis, S.I.; Zervakis, M.; Papanicolaou, A.C.; Zouridakis, G. Altered Rich-Club and Frequency-Dependent Subnetwork Organization in Mild Traumatic Brain Injury: A MEG Resting-State Study. Front. Hum. Neurosci. 2017, 11, 416. [Google Scholar] [CrossRef] [PubMed]
- Antonakakis, M.; Dimitriadis, S.I.; Zervakis, M.; Papanicolaou, A.C.; Zouridakis, G. Alterations in Dynamic Spontaneous Network Microstates in Mild Traumatic Brain Injury: A MEG Beamformed Dynamic Connectivity Analysis. BioRxiv 2019. [Google Scholar] [CrossRef]
- Antonakakis, M.; Dimitriadis, S.I.; Zervakis, M.; Papanicolaou, A.C.; Zouridakis, G. Reconfiguration of Dominant Coupling Modes in Mild Traumatic Brain Injury Mediated by δ-band Activity: A Resting State MEG Study. Neuroscience 2017, 356, 275–286. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Topography of Regional Interaction | Frequency Band | Connectivity Metric | MSS | FO | |
|---|---|---|---|---|---|
| Frontal | Cross-hemispheric | δ Amplitude | Envelop Correlation | 4 | 20 |
| Frontal–Temporal | Within and cross-hemispheric | θ Phase → γ2 Amplitude | Phase-Amplitude Coupling | 3 | 6 |
| Frontal–Parietal | Within and cross-hemispheric | Θ → α2 Amplitude | delay Symbolic Transfer Entropy | 1 | 2 |
| Parieto-Occipital | Cross-hemispheric | α1 Phase | imaginary Phase Locking | 15 | 14 |
| Frontal | Within hemispheres | θ Phase | imaginary Phase Locking | 13 | 16 |
| L Temporal–Frontal | Cross-hemispheric | δ Phase → β Amplitude | Phase-Amplitude Coupling | 5 | 18 |
| R Temporal–Frontal | Within and cross-hemispheric | δ Phase → γ2 Amplitude | Phase-Amplitude Coupling | 7 | - |
| L Parietal–Parieto-Occipital | Within and cross-hemispheric | A1 Phase | imaginary Phase Locking | 12 | 17 |
| Parieto-Occipital | Cross-hemispheric | β Amplitude | Envelope Correlation | 8 | 21 |
| R Temporal-Parieto-Occipital | Within and cross-hemispheric | γ1 Phase | imaginary Phase Locking | 10 | 11 |
| Temporal | Cross-hemispheric | β Amplitude | Envelope Correlation | 9 | 22 |
| Occipital | Cross-hemispheric | α2 Phase → γ1 Amplitude | Phase-Amplitude Coupling | 19 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimitriadis, S.I.; Simos, P.G.; Fletcher, J.Μ.; Papanicolaou, A.C. Typical and Aberrant Functional Brain Flexibility: Lifespan Development and Aberrant Organization in Traumatic Brain Injury and Dyslexia. Brain Sci. 2019, 9, 380. https://doi.org/10.3390/brainsci9120380
Dimitriadis SI, Simos PG, Fletcher JΜ, Papanicolaou AC. Typical and Aberrant Functional Brain Flexibility: Lifespan Development and Aberrant Organization in Traumatic Brain Injury and Dyslexia. Brain Sciences. 2019; 9(12):380. https://doi.org/10.3390/brainsci9120380
Chicago/Turabian StyleDimitriadis, Stavros I., Panagiotis G. Simos, Jack Μ. Fletcher, and Andrew C. Papanicolaou. 2019. "Typical and Aberrant Functional Brain Flexibility: Lifespan Development and Aberrant Organization in Traumatic Brain Injury and Dyslexia" Brain Sciences 9, no. 12: 380. https://doi.org/10.3390/brainsci9120380
APA StyleDimitriadis, S. I., Simos, P. G., Fletcher, J. Μ., & Papanicolaou, A. C. (2019). Typical and Aberrant Functional Brain Flexibility: Lifespan Development and Aberrant Organization in Traumatic Brain Injury and Dyslexia. Brain Sciences, 9(12), 380. https://doi.org/10.3390/brainsci9120380