
Neonatal Handling Positively Modulates Anxiety, Sensorimotor Gating, Working Memory, and Cortico-Hippocampal Neuroplastic Adaptations in Two Genetically Selected Rat Strains Differing in Emotional and Cognitive Traits

 , , , , , , , ,
, , , , , , , , 

Abstract
1. Introduction
2. Material and Methods
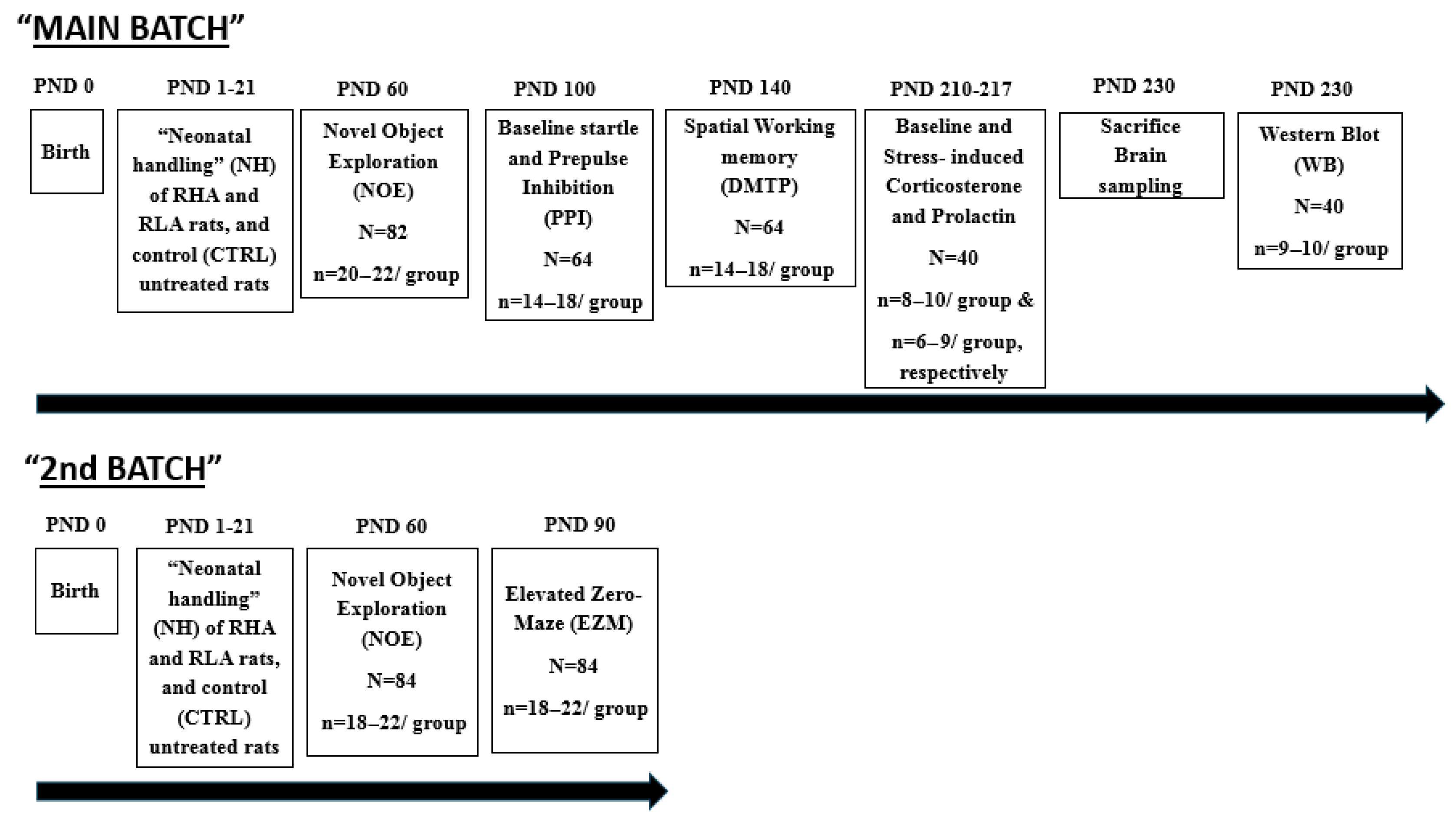
2.1. Animals and Neonatal Handling (NH) Treatment
2.2. Behavioural and Hormonal Measures
2.2.1. Novel Object Exploration (NOE) Test of Anxiety-Related Behaviour
2.2.2. Elevated Zero-Maze (EZM) Test of Anxiety
2.2.3. Baseline Startle and Prepulse Inhibition of the Acoustic Startle Response (Sensorimotor Gating, PPI)
- Five-minute habituation to the startle chamber (only background noise, 55 dB).
- Ten “pulse-alone” trials (105 dB, 40 ms), to allow some habituation of startle and more stable “baseline startle” responses.
- Then, six different trial types were administered 10 times in a random order (for a total of 60 trials). These six trial types were as follows: (i) Pulse-alone (105 dB, 40 ms), “baseline startle” trials, from which the percentage prepulse inhibition (%PPI) is calculated. (ii–v) Prepulse stimuli (20 ms) of four different intensities (65, 70, 75, 80 dB), plus a 100 ms interval followed by the acoustic startle stimulus of 105 dB (40 ms). (vi) Background noise (55 dB), i.e., no stimulus trials.
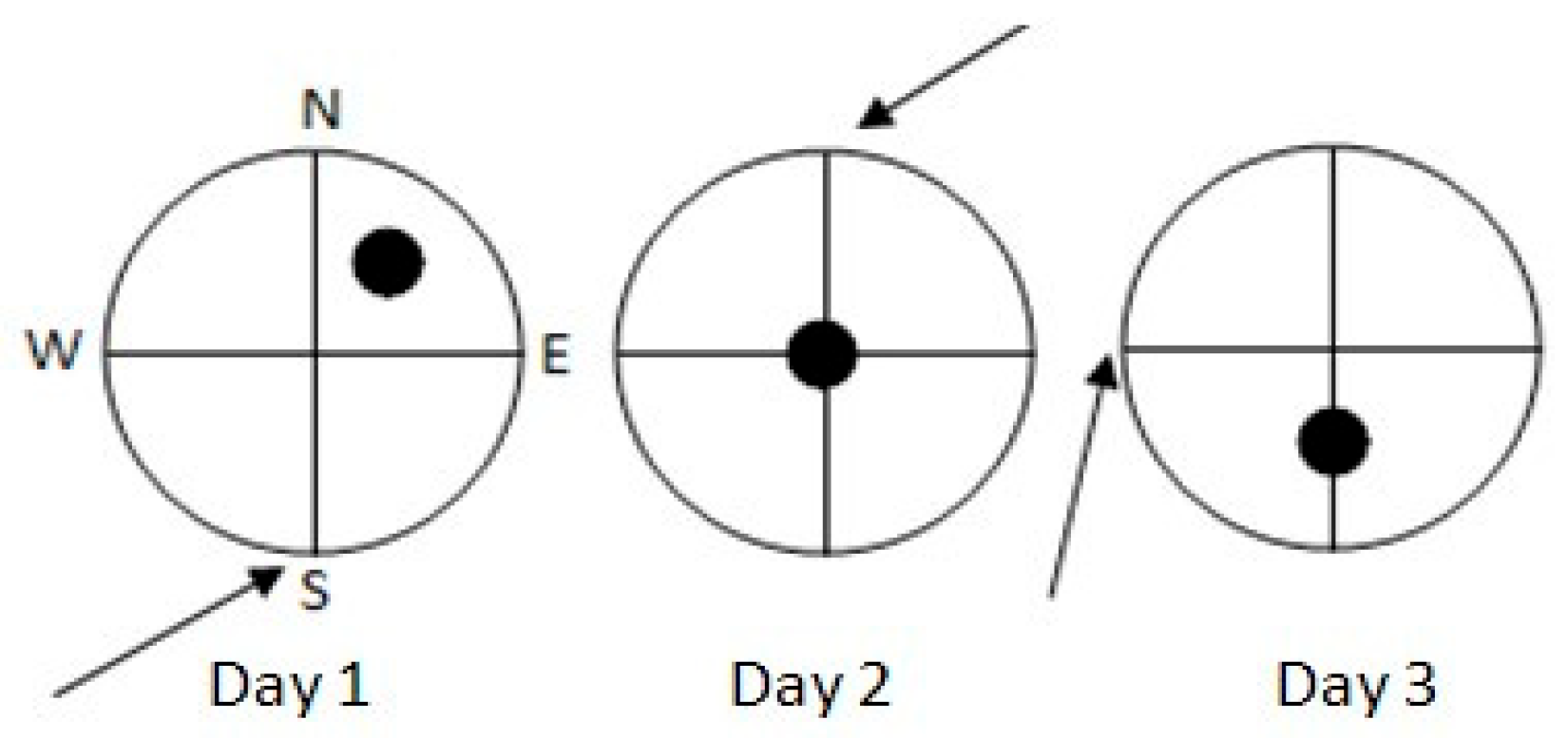
2.2.4. Morris Water Maze: Delayed Matching-to-Place Task (DMTP) of Spatial Working Memory
2.2.5. Hormone Measurements
2.3. Brain Sampling and Molecular Measures
Western Blot (WB) Assay
2.4. Statistical Analyses
3. Results
3.1. Novel Object Exploration (NOE) and Elevated Zero-Maze (EZM) Tests
3.2. Baseline Startle Response and Prepulse Inhibition of the Startle Response (PPI)
3.3. Delayed Matching-to-Place (DMPT) Task of Spatial Working Memory
3.4. Baseline and Post-Stress Corticosterone and Prolactin Responses
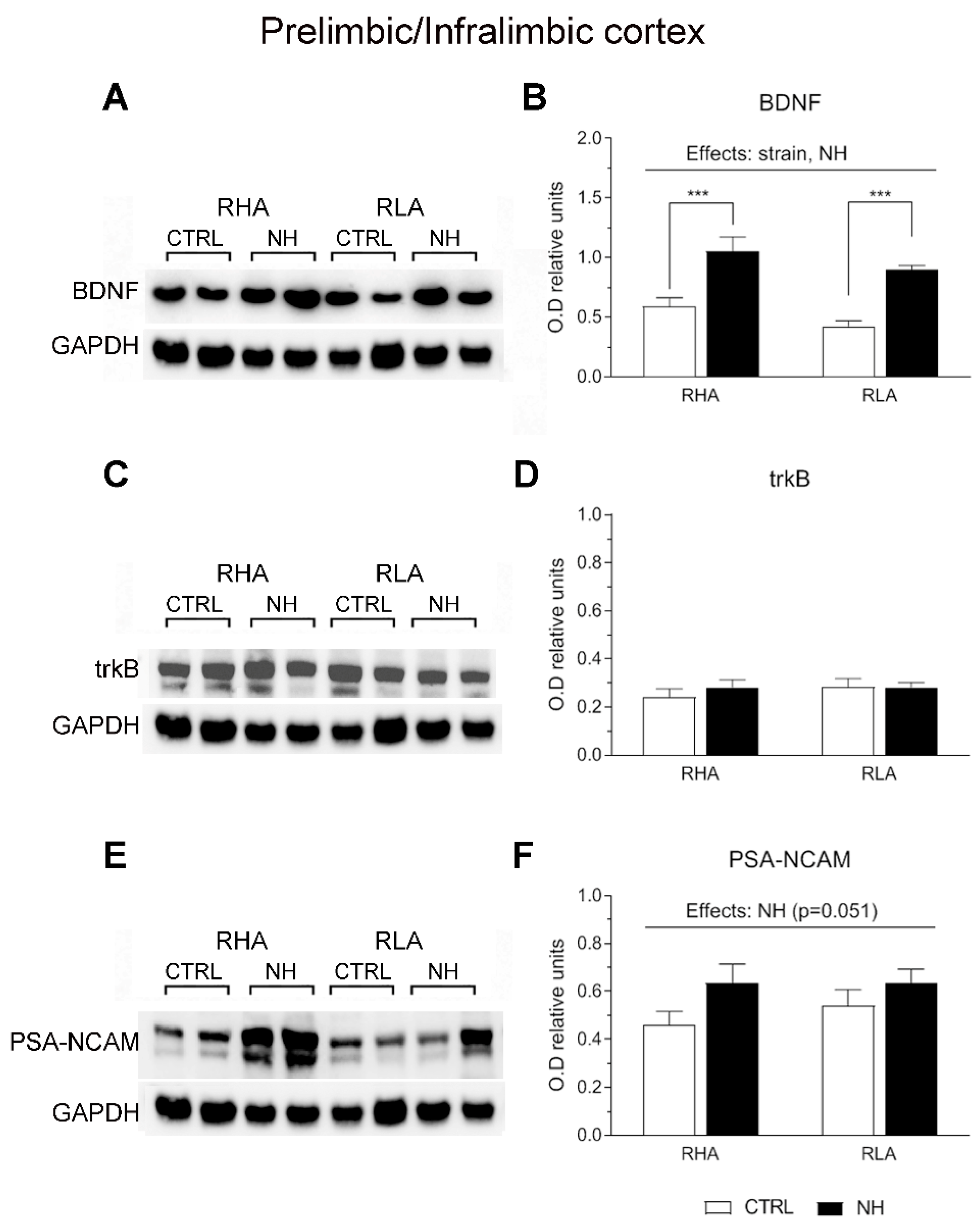
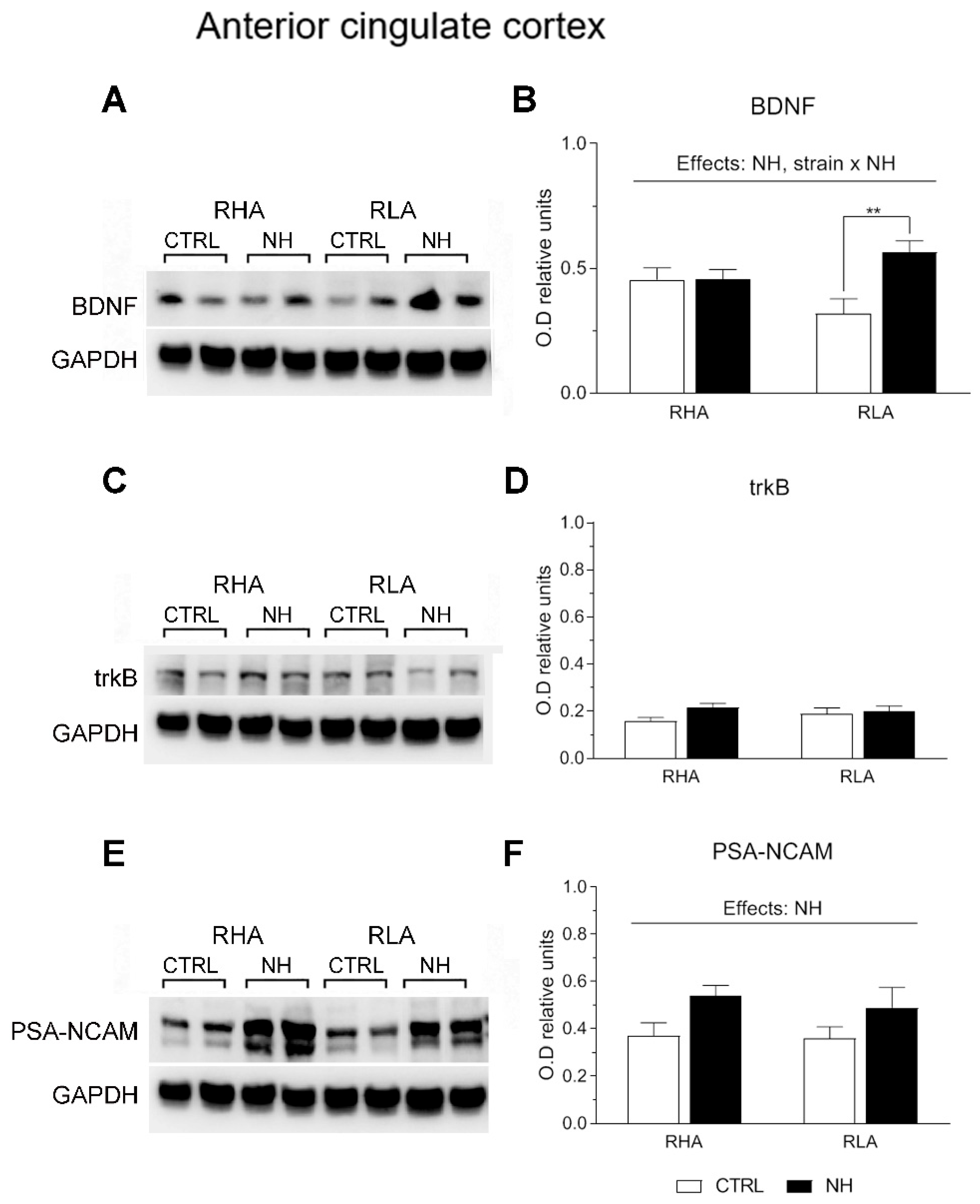
3.5. BDNF, trkB, and PSA-NCAM Protein Levels in PFC (Prelimbic/Infralimbic) and ACg of RHA and RLA Rats: Effects of Neonatal Handling
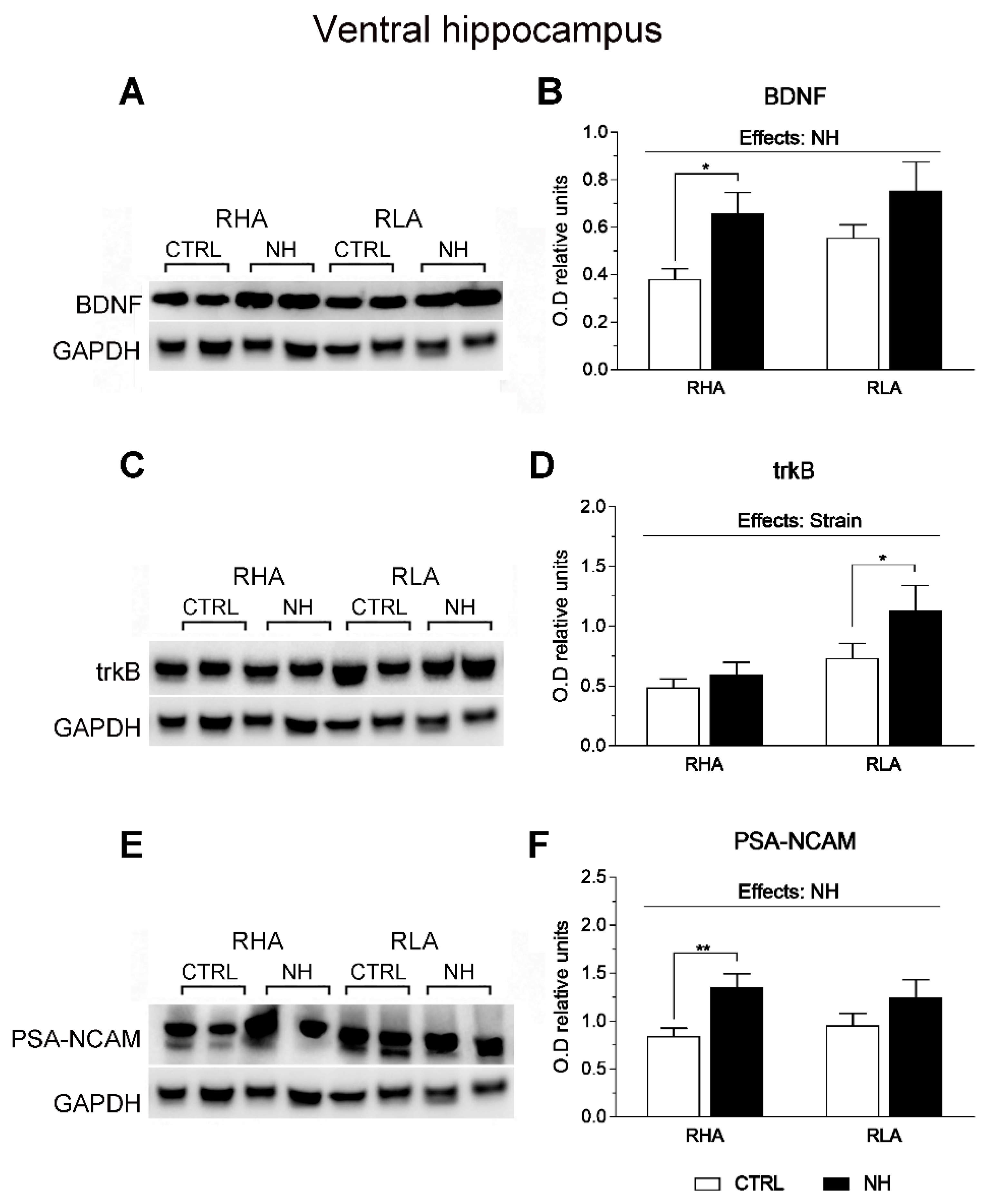
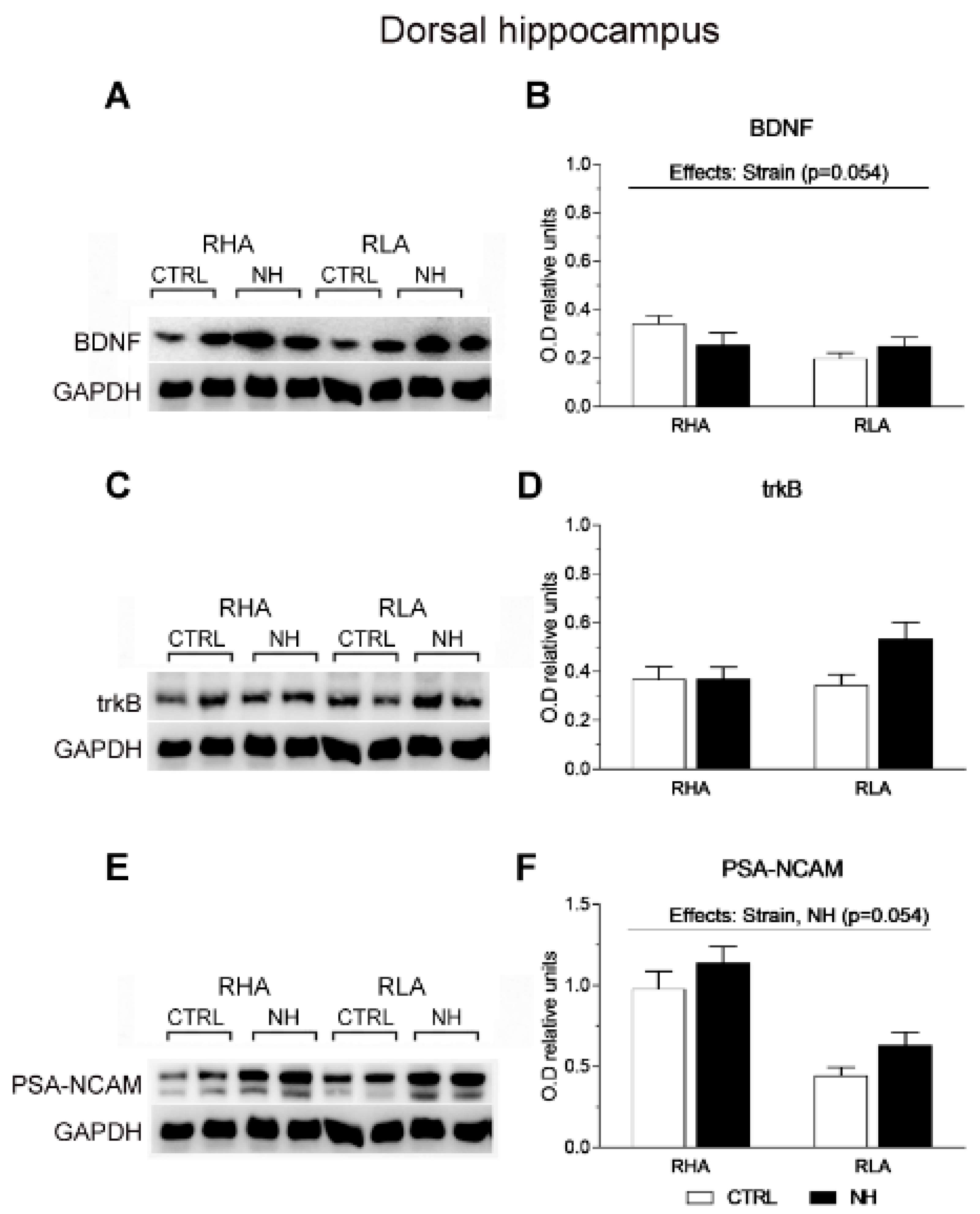
3.6. BDNF, trkB, and PSA-NCAM Protein Levels in vHPC and dHPC of RHA and RLA Rats: Effects of Neonatal Handling
3.7. Correlational Analyses: Relations Among BDNF, trkB, and PSA-NCAM Protein Levels in the Different Areas, and Between Specific Protein Levels and Behaviour, Hormonal, and Cognitive Measures
3.8. Exploratory Factor Analysis of Protein Levels, Behaviour, and Stress Hormone Values
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bignami, G. Selection for High Rates and Low Rates of Avoidance Conditioning in the Rat. Anim. Behav. 1965, 13, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Teruel, A.; Oliveras, I.; Cañete, T.; Rio-Álamos, C.; Tapias-Espinosa, C.; Sampedro-Viana, D.; Sánchez-González, A.; Sanna, F.; Torrubia, R.; González-Maeso, J.; et al. Neurobehavioral and Neurodevelopmental Profiles of a Heuristic Genetic Model of Differential Schizophrenia- and Addiction-Relevant Features: The RHA vs. RLA Rats. Neurosci. Biobehav. Rev. 2021, 131, 597–617. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Teruel, A.; Cañete, T.; Sampedro-Viana, D.; Oliveras, I.; Torrubia, R.; Tobeña, A. Contribution of the Roman rat lines/strains to personality neuroscience: Neurobehavioral modeling of internalizing/externalizing psychopathologies. Personal. Neurosci. 2023, 6, e8. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, O.; Corda, M.G.; Fernández-Teruel, A. A Genetic Model of Impulsivity, Vulnerability to Drug Abuse and Schizophrenia-Relevant Symptoms with Translational Potential: The Roman High- vs. Low-Avoidance Rats. Front Behav Neurosci 2019, 13, 145. [Google Scholar] [CrossRef] [PubMed]
- Castanon, N.; Dulluc, J.; Le Moal, M.; Mormède, P. Maturation of the Behavioral and Neuroendocrine Differences between the Roman Rat Lines. Physiol. Behav. 1994, 55, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Gentsch, C.; Lichtsteiner, M.; Driscoll, P.; Feer, H. Differential Hormonal and Physiological Responses to Stress in Roman High- and Low-Avoidance Rats. Physiol. Behav. 1982, 28, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Papini, M.R.; Fuchs, P.N.; Torres, C. Behavioral Neuroscience of Psychological Pain. Neurosci. Biobehav. Rev. 2015, 48, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Steimer, T.; Python, A.; Schulz, P.E.; Aubry, J.M. Plasma Corticosterone, Dexamethasone (DEX) Suppression and DEX/CRH Tests in a Rat Model of Genetic Vulnerability to Depression. Psychoneuroendocrinology 2007, 32, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Steimer, T.; Driscoll, P. Divergent Stress Responses and Coping Styles in Psychogenetically Selected Roman High-(RHA) and Low-(RLA) Avoidance Rats: Behavioural, Neuroendocrine and Developmental Aspects. Stress 2003, 6, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Dimiziani, A.; Añó, L.B.; Tsartsalis, S.; Millet, P.; Herrmann, F.; Ginovart, N. Differential Involvement of D2 and D3 Receptors during Reinstatement of Cocaine-Seeking Behavior in the Roman High- and Low-Avoidance Rats. Behav. Neurosci. 2019, 133, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Oliveras, I.; Río-Álamos, C.; Cañete, T.; Blázquez, G.; Martínez-Membrives, E.; Giorgi, O.; Corda, M.G.; Tobeña, A.; Fernández-Teruel, A. Prepulse Inhibition Predicts Spatial Working Memory Performance in the Inbred Roman High- and Low-Avoidance Rats and in Genetically Heterogeneous NIH-HS Rats: Relevance for Studying Pre-Attentive and Cognitive Anomalies in Schizophrenia. Front. Behav. Neurosci. 2015, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Vallejo, N.; Cañete, T.; Sampedro-Viana, D.; Güell-Falgueras, P.; Río-Álamos, C.; Oliveras, I.; Tobeña, A.; Aznar, S.; Fernández-Teruel, A. Neonatal Handling Enhances Behavioural and Attentional Domains, and Frontocortical Synaptic Maturation in Rat Models of Schizophrenia-like Behaviour and Anxiety-Related Responses. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2025, 139, 111364. [Google Scholar] [CrossRef] [PubMed]
- Tournier, B.B.; Steimer, T.; Millet, P.; Moulin-Sallanon, M.; Vallet, P.; Ibañez, V.; Ginovart, N. Innately Low D2 Receptor Availability Is Associated with High Novelty-Seeking and Enhanced Behavioural Sensitization to Amphetamine. Int. J. Neuropsychopharmacol. 2013, 16, 1819–1834. [Google Scholar] [CrossRef] [PubMed]
- Serra, M.P.; Poddighe, L.; Boi, M.; Sanna, F.; Piludu, M.A.; Sanna, F.; Corda, M.G.; Giorgi, O.; Quartu, M. Effect of Acute Stress on the Expression of BDNF, TrkB, and PSA-NCAM in the Hippocampus of the Roman Rats: A Genetic Model of Vulnerability/Resistance to Stress-Induced Depression. Int. J. Mol. Sci. 2018, 19, 3745. [Google Scholar] [CrossRef] [PubMed]
- Serra, M.P.; Sanna, F.; Boi, M.; Trucas, M.; Fernández-Teruel, A.; Corda, M.G.; Giorgi, O.; Quartu, M. Effects of Tail Pinch on BDNF and TrkB Expression in the Hippocampus of Roman Low- (RLA) and High-Avoidance (RHA) Rats. Int J Mol Sci 2023, 24, 9498. [Google Scholar] [CrossRef] [PubMed]
- Sønderstrup, M.; Batiuk, M.Y.; Mantas, P.; Tapias-Espinosa, C.; Oliveras, I.; Cañete, T.; Sampedro-Viana, D.; Brudek, T.; Rydbirk, R.; Khodosevich, K.; et al. A Maturational Shift in the Frontal Cortex Synaptic Transcriptional Landscape Underlies Schizophrenia-Relevant Behavioural Traits: A Congenital Rat Model. Eur. Neuropsychopharmacol. 2023, 74, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Meyza, K.Z.; Boguszewski, P.M.; Nikolaev, E.; Zagrodzka, J. Diverse Sensitivity of RHA/Verh and RLA/Verh Rats to Emotional and Spatial Aspects of a Novel Environment as a Result of a Distinct Pattern of Neuronal Activation in the Fear/Anxiety Circuit. Behav. Genet. 2009, 39, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Río-Álamos, C.; Piludu, M.A.; Gerbolés, C.; Barroso, D.; Oliveras, I.; Sánchez-González, A.; Cañete, T.; Tapias-Espinosa, C.; Sampedro-Viana, D.; Torrubia, R.; et al. Volumetric Brain Differences between the Roman Rat Strains: Neonatal Handling Effects, Sensorimotor Gating and Working Memory. Behav. Brain Res. 2019, 361, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Steimer, T.; Driscoll, P.; Schulz, P.E. Brain Metabolism of Progesterone, Coping Behaviour and Emotional Reactivity in Male Rats from Two Psychogenetically Selected Lines. J. Neuroendocrinol. 1997, 9, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Roozendaal, B.; Wiersma, A.; Driscoll, P.; Koolhaas, J.M.; Bohus, B. Vasopressinergic Modulation of Stress Responses in the Central Amygdala of the Roman High-Avoidance and Low-Avoidance Rat. Brain Res. 1992, 596, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Steimer, T.; Driscoll, P. Inter-Individual vs Line/Strain Differences in Psychogenetically Selected Roman High-(RHA) and Low-(RLA) Avoidance Rats: Neuroendocrine and Behavioural Aspects. Neurosci. Biobehav. Rev. 2005, 29, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, F.; Brenè, S.; Mathé, A.A. BDNF in Schizophrenia, Depression and Corresponding Animal Models. Mol. Psychiatry 2005, 10, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Boulle, F.; Van Den Hove, D.L.A.; Jakob, S.B.; Rutten, B.P.; Hamon, M.; Van Os, J.; Lesch, K.P.; Lanfumey, L.; Steinbusch, H.W.; Kenis, G. Epigenetic Regulation of the BDNF Gene: Implications for Psychiatric Disorders. Mol. Psychiatry 2012, 17, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Thome, J.; Foley, P.; Riederer, P. Neurotrophic Factors and the Maldevelopmental Hypothesis of Schizophrenic Psychoses. Review Article. J. Neural. Transm. 1998, 105, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Gliwińska, A.; Czubilińska-Łada, J.; Więckiewicz, G.; Świętochowska, E.; Badeński, A.; Dworak, M.; Szczepańska, M. The Role of Brain-Derived Neurotrophic Factor (BDNF) in Diagnosis and Treatment of Epilepsy, Depression, Schizophrenia, Anorexia Nervosa and Alzheimer’s Disease as Highly Drug-Resistant Diseases: A Narrative Review. Brain Sci. 2023, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Altar, C.A.; Laeng, P.; Jurata, L.W.; Brockman, J.A.; Lemire, A.; Bullard, J.; Bukhman, Y.V.; Young, T.A.; Charles, V.; Palfreyman, M.G. Electroconvulsive Seizures Regulate Gene Expression of Distinct Neurotrophic Signaling Pathways. J. Neurosci. 2004, 24, 2667–2677. [Google Scholar] [CrossRef] [PubMed]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and TrkB MRNA in Rat Brain by Chronic Electroconvulsive Seizure and Antidepressant Drug Treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef] [PubMed]
- Siuciak, J.A.; Lewis, D.R.; Wiegand, S.J.; Lindsay, R.M. Antidepressant-like Effect of Brain-Derived Neurotrophic Factor (BDNF). Pharmacol. Biochem. Behav. 1997, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Makino, S.; Kvetňanský, R.; Post, R.M. Effects of Stress on Neurotrophic Factor Expression in the Rat Brain. Ann. N. Y Acad. Sci. 1995, 771, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, T.; Kawai, Y.; Nemoto, K.; Sekimoto, M.; Toné, S.; Senba, E. Immobilization Stress Reduced the Expression of Neurotrophins and Their Receptors in the Rat Brain. Neurosci. Res. 1997, 28, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.J. Down-Regulation of the Trk-B Signal Pathway: The Possible Pathogenesis of Major Depression. Med. Hypotheses 2004, 62, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Favalli, G.; Li, J.; Belmonte-de-Abreu, P.; Wong, A.H.C.; Daskalakis, Z.J. The Role of BDNF in the Pathophysiology and Treatment of Schizophrenia. J. Psychiatr. Res. 2012, 46, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Durany, N.; Michel, T.; Zöchling, R.; Boissl, K.W.; Cruz-Sánchez, F.F.; Riederer, P.; Thome, J. Brain-Derived Neurotrophic Factor and Neurotrophin 3 in Schizophrenic Psychoses. Schizophr. Res. 2001, 52, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, V.; Bove, S.E.; Volfson, D.; Lewis, D.A.; Kleiman, R.J.; Lanz, T.A. Evaluation of TrkB and BDNF Transcripts in Prefrontal Cortex, Hippocampus, and Striatum from Subjects with Schizophrenia, Bipolar Disorder, and Major Depressive Disorder. Neurobiol. Dis. 2015, 77, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.J.; Yu, Y.W.Y.; Lin, C.H.; Tsai, S.J. An Association Study of a Brain-Derived Neurotrophic Factor Val66Met Polymorphism and Clozapine Response of Schizophrenic Patients. Neurosci. Lett. 2003, 349, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Penadés, R.; López-Vílchez, I.; Catalán, R.; Arias, B.; González-Rodríguez, A.; García-Rizo, C.; Masana, G.; Ruíz, V.; Mezquida, G.; Bernardo, M. BDNF as a Marker of Response to Cognitive Remediation in Patients with Schizophrenia: A Randomized and Controlled Trial. Schizophr. Res. 2018, 197, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Li, W.; Lv, L.; Zhang, Z.; Zhan, X. BDNF as a biomarker in diagnosis and evaluation of treatment for schizophrenia and depression. Discov Med. 2018, 26, 127–136. [Google Scholar] [PubMed]
- Ahmad, R.; Azman, K.F.; Yahaya, R.; Shafin, N.; Omar, N.; Ahmad, A.H.; Zakaria, R.; Wijaya, A.; Othman, Z. Brain-Derived Neurotrophic Factor (BDNF) in Schizophrenia Research: A Quantitative Review and Future Directions. AIMS Neurosci. 2023, 10, 5–32. [Google Scholar] [CrossRef] [PubMed]
- Lencz, T.; Lipsky, R.H.; DeRosse, P.; Burdick, K.E.; Kane, J.M.; Malhotra, A.K. Molecular Differentiation of Schizoaffective Disorder from Schizophrenia Using BDNF Haplotypes. Br. J. Psychiatry 2009, 194, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Ursini, G.; Cavalleri, T.; Fazio, L.; Angrisano, T.; Iacovelli, L.; Porcelli, A.; Maddalena, G.; Punzi, G.; Mancini, M.; Gelao, B.; et al. BDNF Rs6265 Methylation and Genotype Interact on Risk for Schizophrenia. Epigenetics 2016, 11, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Ashe, P.C.; Chlan-Fourney, J.; Juorio, A.V.; Li, X.M. Brain-Derived Neurotrophic Factor (BDNF) MRNA in Rats with Neonatal Ibotenic Acid Lesions of the Ventral Hippocampus. Brain Res. 2002, 956, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Tseng, K.Y.; Chambers, R.A.; Lipska, B.K. The Neonatal Ventral Hippocampal Lesion as a Heuristic Neurodevelopmental Model of Schizophrenia. Behav. Brain Res. 2009, 204, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, F.; Molteni, R.; Roceri, M.; Bedogni, F.; Santero, R.; Fossati, C.; Gennarelli, M.; Racagni, G.; Riva, M.A. Effect of Antipsychotic Drugs on Brain-Derived Neurotrophic Factor Expression under Reduced N-Methyl-D-Aspartate Receptor Activity. J. Neurosci. Res. 2003, 72, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Teruel, A.; Giménez-Llort, L.; Escorihuela, R.M.; Gil, L.; Aguilar, R.; Steimer, T.; Tobeña, A. Early-Life Handling Stimulation and Environmental Enrichment: Are Some of Their Effects Mediated by Similar Neural Mechanisms? Pharmacol. Biochem. Behav. 2002, 73, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Levine, S. A Further Study of Infantile Handling and Adult Avoidance Learning. J. Pers. 1956, 25, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Raineki, C.; Lucion, A.B.; Weinberg, J. Neonatal Handling: An Overview of the Positive and Negative Effects. Dev. Psychobiol. 2014, 56, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Depue, R.A. A Novel Neurobehavioral Framework of the Effects of Positive Early Postnatal Experience on Incentive and Consummatory Reward Sensitivity. Neurosci. Biobehav. Rev. 2019, 107, 615–640. [Google Scholar] [CrossRef] [PubMed]
- Katsouli, S.; Stamatakis, A.; Giompres, P.; Kouvelas, E.D.; Stylianopoulou, F.; Mitsacos, A. Sexually Dimorphic Long-Term Effects of an Early Life Experience on AMPA Receptor Subunit Expression in Rat Brain. Neuroscience 2014, 257, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Meaney, M.J.; Aitken, D.H.; Van Berkel, C.; Bhatnagar, S.; Sapolsky, R.M. Effect of Neonatal Handling on Age-Related Impairments Associated with the Hippocampus. Science 1988, 239, 766–768. [Google Scholar] [CrossRef] [PubMed]
- Meaney, M.J.; Aitken, D.H.; Viau, V.; Sharma, S.; Sarrieau, A. Neonatal Handling Alters Adrenocortical Negative Feedback Sensitivity and Hippocampal Type II Glucocorticoid Receptor Binding in the Rat. Neuroendocrinology 1989, 50, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Panagiotaropoulos, T.; Pondiki, S.; Papaioannou, A.; Alikaridis, F.; Stamatakis, A.; Gerozissis, K.; Stylianopoulou, F. Neonatal Handling and Gender Modulate Brain Monoamines and Plasma Corticosterone Levels Following Repeated Stressors in Adulthood. Neuroendocrinology 2004, 80, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, A.; Gerozissis, K.; Prokopiou, A.; Bolaris, S.; Stylianopoulou, F. Sex Differences in the Effects of Neonatal Handling on the Animal’s Response to Stress and the Vulnerability for Depressive Behaviour. Behav. Brain Res. 2002, 129, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Pondiki, S.; Stamatakis, A.; Fragkouli, A.; Philippidis, H.; Stylianopoulou, F. Effects of Neonatal Handling on the Basal Forebrain Cholinergic System of Adult Male and Female Rats. Neuroscience 2006, 142, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Toutountzi, E.; Fragioudaki, K.; Kouvelas, E.D.; Stylianopoulou, F.; Mitsacos, A. Selective Effects of Neonatal Handling on Rat Brain N-Methyl-d-Aspartate Receptors. Neuroscience 2009, 164, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Taschetto Vey, L.; Zuquetto Rosa, H.; Cristine Silva Barcelos, R.; Tironi Dias, V.; Izabel Ugalde Marques da Rocha, M.; Escobar Burger, M. Neonatal Handling Increases Neurogenesis, BDNF and GR in the Hippocampus Favoring Memory Acquisition in Rats. Brain Res 2020, 1745, 146921. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Palkovits, M. Punch Sampling Biopsy Technique. Methods Enzymol. 1983, 103, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Antoniazzi, C.T.D.; Metz, V.G.; Roversi, K.; Freitas, D.L.; Vey, L.T.; Dias, V.T.; Segat, H.J.; Duarte, M.M.M.F.; Burger, M.E. Tactile Stimulation during Different Developmental Periods Modifies Hippocampal BDNF and GR, Affecting Memory and Behavior in Adult Rats. Hippocampus 2017, 27, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Garoflos, E.; Panagiotaropoulos, T.; Pondiki, S.; Stamatakis, A.; Philippidis, E.; Stylianopoulou, F. Cellular Mechanisms Underlying the Effects of an Early Experience on Cognitive Abilities and Affective States. Ann. Gen. Psychiatry 2005, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, V.; Lamarque, S.; Le Moal, M.; Piazza, P.V.; Abrous, D.N. Postnatal Stimulation of the Pups Counteracts Prenatal Stress-Induced Deficits in Hippocampal Neurogenesis. Biol. Psychiatry 2006, 59, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Kline, P. An Easy Guide to Factor Analysis; Routledge: London, UK, 1994. [Google Scholar]
- Oliveras, I.; Soria-Ruiz, O.J.; Sampedro-Viana, D.; Cañete, T.; Río-Álamos, C.; Tobeña, A.; Fernández-Teruel, A. Different Maturation Patterns for Sensorimotor Gating and Startle Habituation Deficits in Male and Female RHA vs RLA Rats. Behav. Brain Res. 2022, 434, 114021. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Stamatakis, A.; Pondiki, S.; Stamatakis, A.; Pondiki, S.; Kitraki, E.; Diamantopoulou, A.; Panagiotaropoulos, T.; Raftogianni, A.; Stylianopoulou, F. Effect of Neonatal Handling on Adult Rat Spatial Learning and Memory Following Acute Stress. Stress 2008, 11, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Willig, F.; M’Harzi, M.; Bardelay, C.; Viet, D.; Delacour, J. Roman Strains as a Psychogenetic Model for the Study of Working Memory: Behavioral and Biochemical Data. Pharmacol. Biochem. Behav. 1991, 40, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.C.; Bunce, J.G.; Barbas, H. Prefrontal-Hippocampal Pathways Underlying Inhibitory Control over Memory. Neurobiol. Learn. Mem. 2016, 134 Pt A, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.P.; Robbins, T.W. The Role of Prefrontal Cortex in Cognitive Control and Executive Function. Neuropsychopharmacology 2022, 47, 72–89. [Google Scholar] [CrossRef] [PubMed]
- McNaughton, N.; Corr, P.J. A Two-Dimensional Neuropsychology of Defense: Fear/Anxiety and Defensive Distance. Neurosci. Biobehav. Rev. 2004, 28, 285–305. [Google Scholar] [CrossRef] [PubMed]
- McNaughton, N.; Gray, J.A. The Neuropsychology of Anxiety: An Enquiry into the Funtions of the Septo-Hippocampal System, 3rd ed.; Oxford University Press: Oxford, UK, 2024. [Google Scholar]
- Papini, M.R.; Green, T.A.; Mármol Contreras, Y.; Torres, C.; Ogawa, M.; Li, Z. Frustrative Nonreward: Behavior, Circuits, Neurochemistry, and Disorders. J. Neurosci. 2024, 44, e1021242024. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Odaka, H.; Adachi, N. Actions of Brain-Derived Neurotrophin Factor in the Neurogenesis and Neuronal Function, and Its Involvement in the Pathophysiology of Brain Diseases. Int. J. Mol. Sci. 2018, 19, 3650. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, R. Infantile Experience and Play Motivation. Soc. Neurosci. 2010, 5, 422–440. [Google Scholar] [CrossRef] [PubMed]
- Costela, C.; Tejedor-Real, P.; Mico, J.A.; Gibert-Rahola, J. Effect of Neonatal Handling on Learned Helplessness Model of Depression. Physiol. Behav. 1995, 57, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Tejedor-Real, P.; Costela, C.; Gibert-Rahola, J. Neonatal Handling Reduces Emotional Reactivity and Susceptibility to Learned Helplessness. Involvement of Catecholaminergic Systems. Life Sci. 1997, 62, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Tejedor-Real, P.; Sahagún, M.; Biguet, N.F.; Mallet, J. Neonatal Handling Prevents the Effects of Phencyclidine in an Animal Model of Negative Symptoms of Schizophrenia. Biol. Psychiatry 2007, 61, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.W.; Park, C.H.; Choi, S.H.; Kim, N.J.; Kim, H.S.; Choe, J.C.; Suh, Y.H. The Postnatal Environment Can Counteract Prenatal Effects on Cognitive Ability, Cell Proliferation, and Synaptic Protein Expression. FASEB J. 2003, 17, 1556–1558. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anxiety-related and stress hormone responses, PPI and spatial working memory (DMTP) in control (untreated; CTRL) and neonatally-handled (NH) Roman high- (RHA) and low-avoidance (RLA) rats. | |||||
| Means (±SEM) | |||||
| “MAIN BATCH” | RLA-CTRL | RLA-NH | RHA-CTRL | RHA-NH | EFFECTS |
| NOE-L (PND60) | 61.8 (12.5) (n = 20) | 8.5 (1.3) a (n = 20) | 10.9 (3.8) b (n = 20) | 4.4 (1.3) a (n = 22) | S, NH, S*NH |
| NOE-T | 19.7 (6.6) | 112.6 (5.9) a | 93.0 (5.8) b | 125.2 (9.5) a | S, NH, S*NH |
| (#) “2nd BATCH” | |||||
| (#) NOE-L | 49.1 (8.9) | 10.2 (1.7) a | 4.1 (0.7) b | 3.6 (0.5) | S, NH, S*NH |
| (#) NOE-T (PND60) | 13.4 (2.7) (n = 18) | 105.0 (6.4) a (n = 22) | 121.1 (3.2) b (n = 22) | 142.9 (3.0) a (n = 22) | S, NH, S*NH |
| (#) EZM-T | 68.0 (9.1) | 103.7 (9.6) a | 83.3 (9.7) | 144.2 (5.6) a | S, NH |
| (#) EZM-HD (PND90) | 6.9 (0.7) (n = 18) | 9.8 (0.8) a (n = 22) | 12.8 (0.8) b (n = 22) | 13.8 (0.7) (n = 22) | S, NH |
| “MAIN BATCH” | |||||
| Baseline startle (&) (PND100) | 2265.5 (418.8) (n = 16) | 1915.6 (295.6) (n = 18) | 1301.3 (214.3) b (n = 14) | 1806.7 (275.5) (n = 16) | |
| % PPI | |||||
| 65dB | 42.3 (7.5) | 30.3 (5.3) | 12.0 (6.8) b | 33.4 (8.4) a | S |
| 70 dB | 53.6 (5.1) | 50.1 (5.2) | 8.2 (8.3) b | 38.9 (10.2) a | NH*Intensity |
| 75 dB | 67.5 (4.7) | 56.2 (5.0) | 41.2 (5.3) b | 52.1 (7.8) | |
| 80 dB | 72.2 (3.7) | 65.0 (3.5) | 52.4 (6.1) b | 66.1 (4.2) | |
| Total %PPI | 58.9 (4.4) | 50.6 (4.2) | 28.5 (5.2) b | 47.8 (7.2) a | S, S*NH |
| DMTP task [mean (T1-T2)] (PND140) | 1229.0 (232.8) (n = 16) | 1542.1 (211.6) (n = 18) | 839.4 (404.8) (n = 14) | 1877.3 (422.9)a (n = 16) | NH |
| (##) | 1625.4 (308.7) (n = 10) | 252.4 (674.6) c (n = 12) | |||
| BASELINE CORT (ng/mL) (PND210) | 66.4 (11.3) | 46.1 (5.8) | 73.9 (9.1) | 65.6 (8.6) | |
| POST-STRESS CORT (ng/mL) | 329.2 (16.5) (n = 8) | 311.9 (21.4) (n = 8) | 265.1 (10.9) b (n = 10) | 210.9 (14.5) a (n = 9) | S, NH |
| BASELINE PRL (ng/mL) (PND210) | 14.4 (3.0) | 16.1 (3.6) | 6.0 (0.9) | 9.8 (1.1) | S |
| POST- STRESS PRL (ng/mL) | 33.1 (5.0) (n = 6) | 22.6 (1.8) a (n = 8) | 13.6 (0.9) b (n = 7) | 9.5 (1.5) (n = 9) | S, NH |
| (A) Varimax-rotated (orthogonal) factor analysis with the twelve protein parameters | |||
| COMPONENTS | |||
| 1 | 2 | 3 | |
| BDNF-PFC | 0.18 | 0.40 | 0.64 |
| trkB-PFC | −0.23 | 0.49 | 0.49 |
| PSA-NCAM-PFC | −0.06 | 0.82 | −0.08 |
| BDNF-ACg | 0.50 | −0.17 | 0.50 |
| trkB-ACg | 0.85 | −0.16 | 0.19 |
| PSA-NCAM-ACg | 0.67 | 0.09 | 0.44 |
| BDNF-vHPC | 0.83 | −0.08 | −0.05 |
| trkB-vHPC | 0.02 | 0.47 | −0.63 |
| PSA-NCAM-vHPC | 0.30 | 0.76 | 0.09 |
| BDNF-dHPC | 0.30 | −0.68 | −0.03 |
| trkB-dHPC | 0.80 | 0.11 | −0.19 |
| PSA-NCAM-dHPC | −0.12 | 0.04 | 0.72 |
| % variance explained | 29.03 | 19.65 | 14.20 |
| Total variance explained | 62.9% | ||
| (B) Obliquely rotated (oblimin direct rotation) principal components factor analysis with the selected molecular, behavioural, and hormonal measures | |||
| COMPONENTS (“Tentative names”) | |||
| 1 | 2 | 3 | |
| “Attention-stress” | “Emotion-memory” | ||
| BDNF-PFC | --- | 0.52 | --- |
| PSA-NCAM-PFC | --- | 0.74 | --- |
| trkB-ACg | --- | --- | 0.86 |
| BDNF-vHPC | --- | --- | 0.83 |
| PSA-NCAM-vHPC | --- | 0.78 | --- |
| PSA-NCAM-dHPC | 0.84 | --- | --- |
| NOE-T | --- | 0.73 | --- |
| % PPI | −0.74 | --- | --- |
| DMTP-WM | −0.51 | 0.56 | --- |
| Post-Stress-Cort | −0.79 | --- | --- |
| % explained variance | 25.4 | 22.5 | 17.6 |
| Total variance (%) | 65.5% | --- | --- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Río-Álamos, C.; Serra, M.P.; Sanna, F.; Piludu, M.A.; Boi, M.; Cañete, T.; Sampedro-Viana, D.; Oliveras, I.; Tobeña, A.; Corda, M.G.; et al. Neonatal Handling Positively Modulates Anxiety, Sensorimotor Gating, Working Memory, and Cortico-Hippocampal Neuroplastic Adaptations in Two Genetically Selected Rat Strains Differing in Emotional and Cognitive Traits. Brain Sci. 2025, 15, 776. https://doi.org/10.3390/brainsci15080776
Río-Álamos C, Serra MP, Sanna F, Piludu MA, Boi M, Cañete T, Sampedro-Viana D, Oliveras I, Tobeña A, Corda MG, et al. Neonatal Handling Positively Modulates Anxiety, Sensorimotor Gating, Working Memory, and Cortico-Hippocampal Neuroplastic Adaptations in Two Genetically Selected Rat Strains Differing in Emotional and Cognitive Traits. Brain Sciences. 2025; 15(8):776. https://doi.org/10.3390/brainsci15080776
Chicago/Turabian StyleRío-Álamos, Cristóbal, Maria P. Serra, Francesco Sanna, Maria A. Piludu, Marianna Boi, Toni Cañete, Daniel Sampedro-Viana, Ignasi Oliveras, Adolf Tobeña, Maria G. Corda, and et al. 2025. "Neonatal Handling Positively Modulates Anxiety, Sensorimotor Gating, Working Memory, and Cortico-Hippocampal Neuroplastic Adaptations in Two Genetically Selected Rat Strains Differing in Emotional and Cognitive Traits" Brain Sciences 15, no. 8: 776. https://doi.org/10.3390/brainsci15080776
APA StyleRío-Álamos, C., Serra, M. P., Sanna, F., Piludu, M. A., Boi, M., Cañete, T., Sampedro-Viana, D., Oliveras, I., Tobeña, A., Corda, M. G., Giorgi, O., Fernández-Teruel, A., & Quartu, M. (2025). Neonatal Handling Positively Modulates Anxiety, Sensorimotor Gating, Working Memory, and Cortico-Hippocampal Neuroplastic Adaptations in Two Genetically Selected Rat Strains Differing in Emotional and Cognitive Traits. Brain Sciences, 15(8), 776. https://doi.org/10.3390/brainsci15080776