
Bidirectional Role of Pericytes in Ischemic Stroke

Abstract
1. Introduction
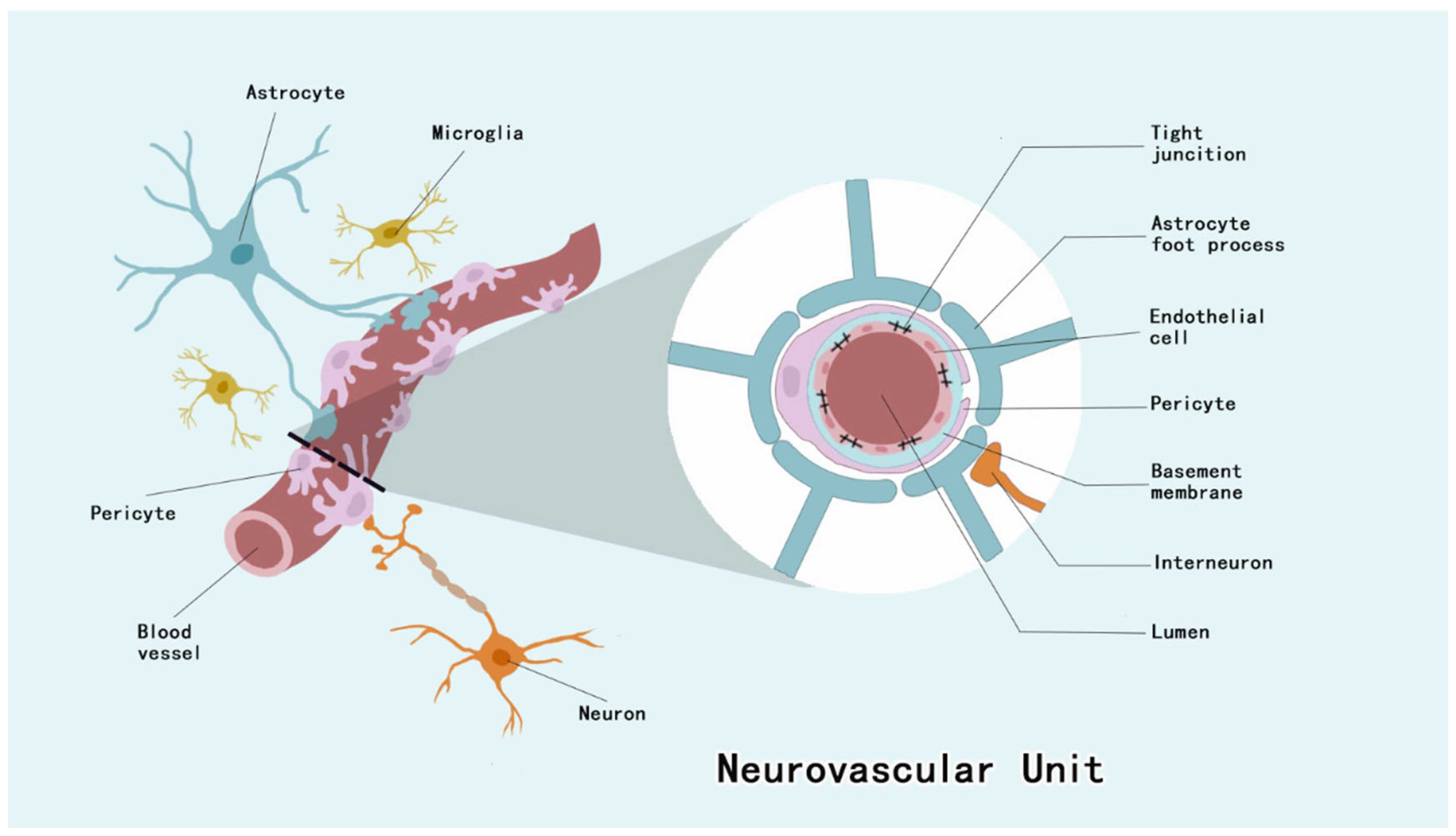
2. Pericytes
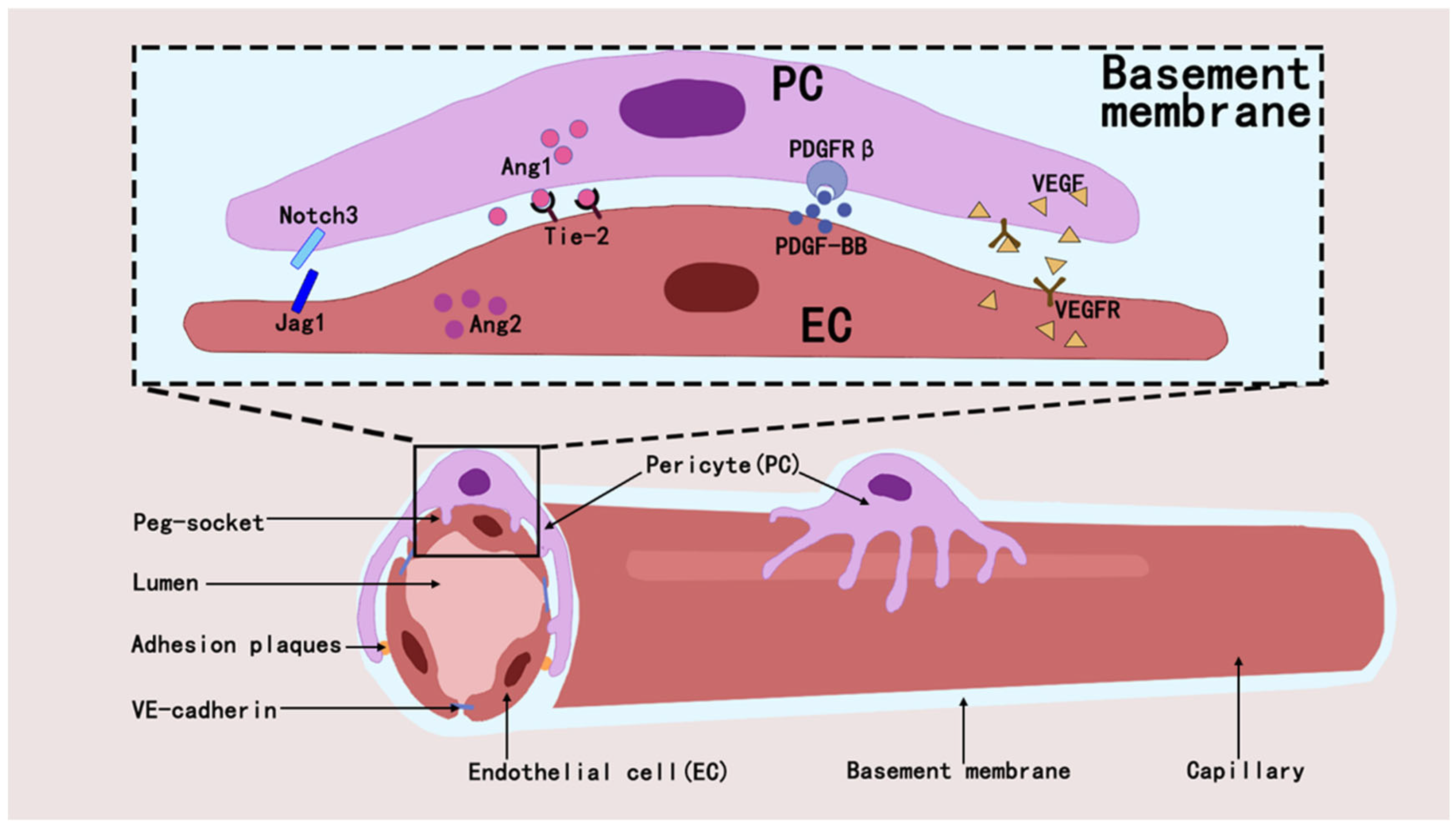
2.1. Pericytes Form the BBB and Maintain CNS Homeostasis
2.2. Pericytes and Angiogenesis and Blood Flow Regulation
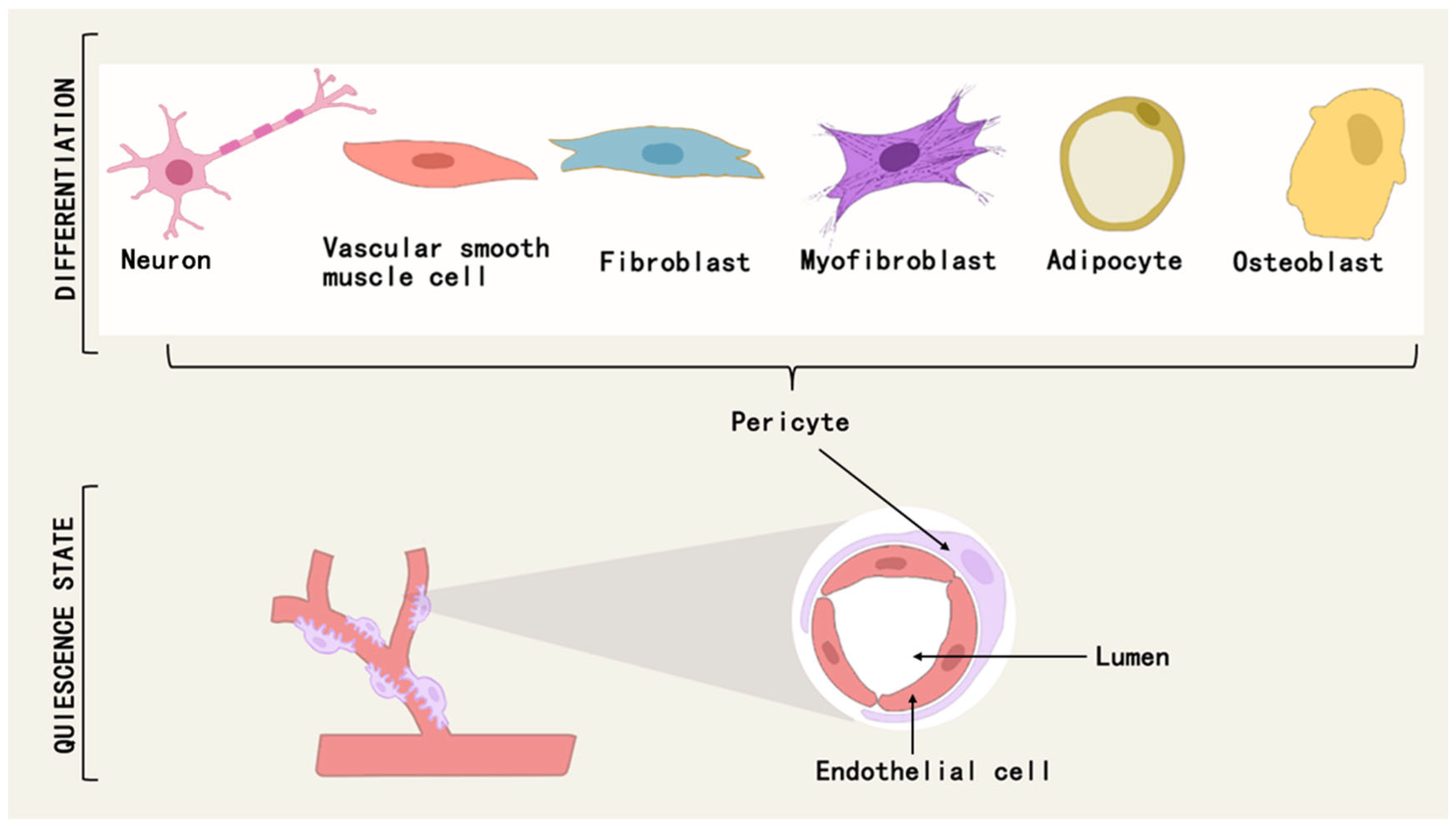
2.3. Pericyte Differentiation and Tissue Repair and Fibrosis
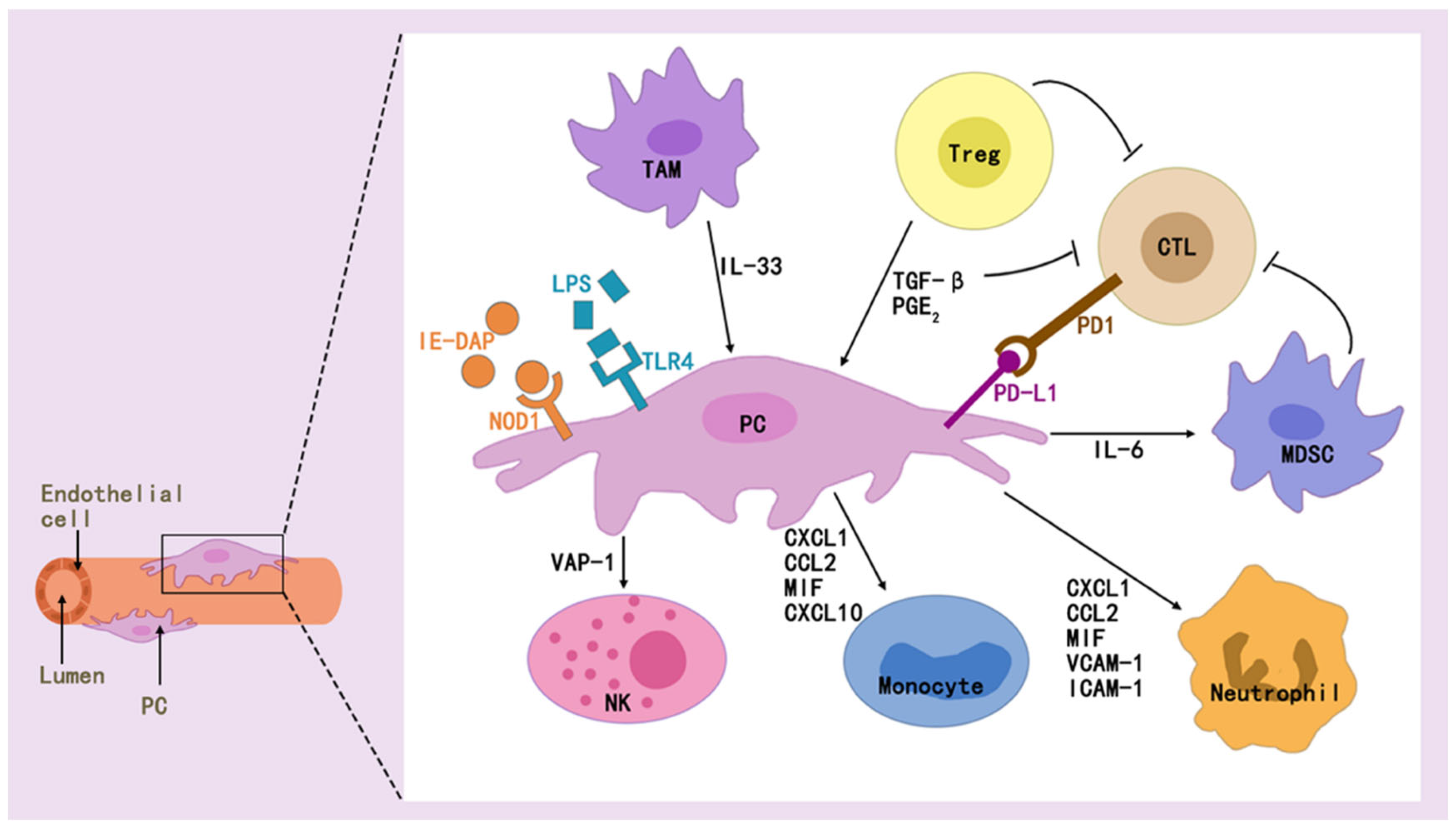
2.4. Pericytes and Immune Regulation
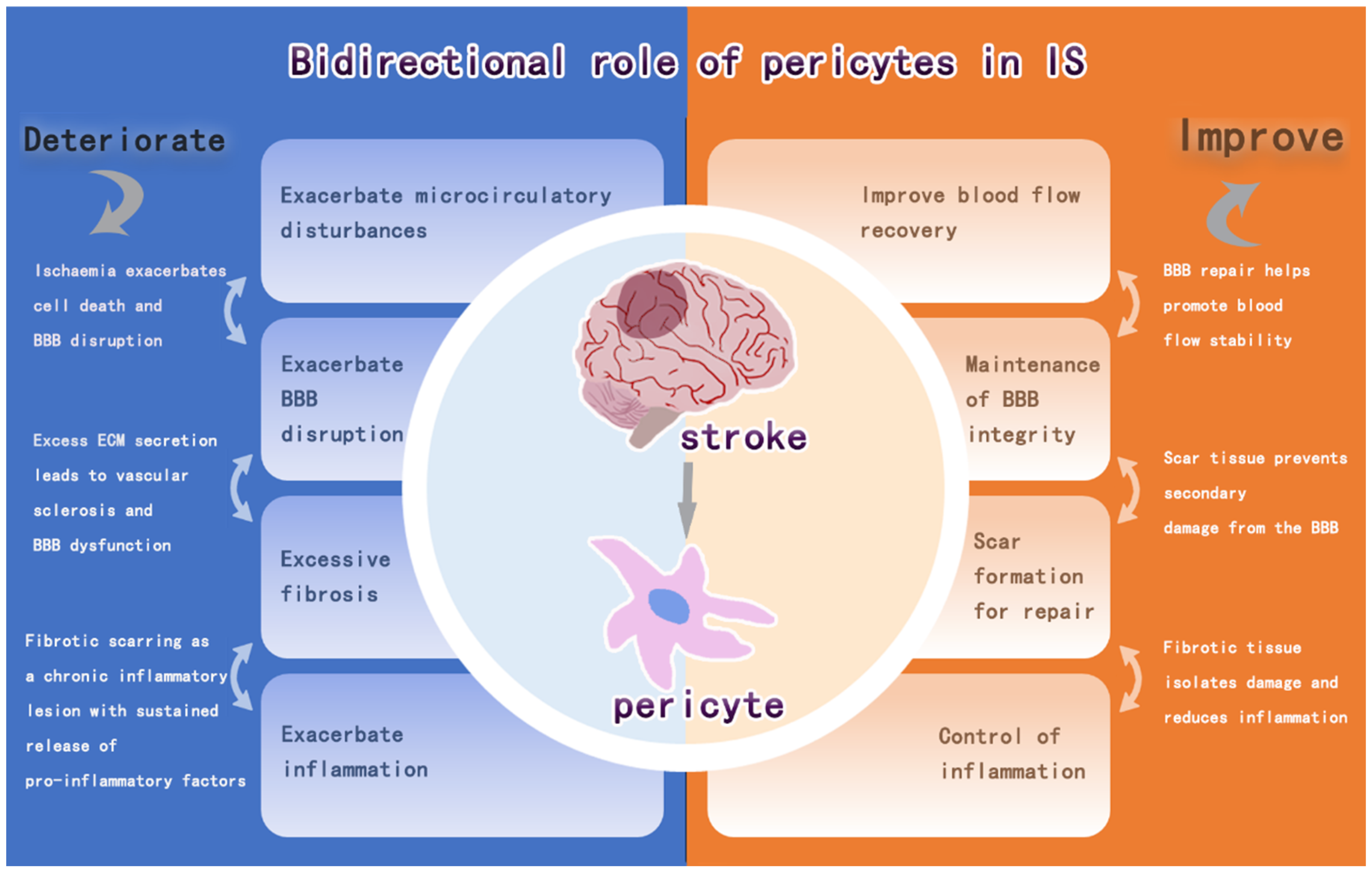
3. Pericytes and IS
3.1. Pericytes and Cerebrovascular Regulation and Regeneration After IS
3.2. Bidirectional Action of Pericytes on the BBB Under Conditions of Cerebral Hypoxia
3.3. Pericytes Bi-Directionally Regulate Intracerebral Immunity After IS
3.4. Bifacial Role of Pericytes in Intracerebral Scar Formation and Fibrosis After IS
4. Targeted Pericyte Therapy for IS

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Direction of Action | Treatment | Participants | Mechanism of Action |
|---|---|---|---|
| Regulation of blood flow and blood vessels | Adenosine [165] | In vitro, Mice | Activation of ATP-sensitive K+ channels (Gαs/cAMP/PKA pathway) in CNS pericytes results in pericyte relaxation, dilation of capillaries, and increased cerebral blood flow (CBF). |
| Iptakalim (IPT) [166] | Mice, In vitro | By preventing the formation of the SUR2/EPAC1 complex, it promotes the opening of K(ATP) channels, thereby inhibiting pericyte contraction. | |
| PI3Kδ inhibitor CAL-101 [167] | Mice, In vitro | Inhibition of TNF α-induced TRPV2 expression in OGD/R-treated PERICYTEs results in inhibition of Ca2+ uptake and PERICYTE shrinkage. | |
| Rapamycin [168] | Mice, In vitro | Decreased pericyte contraction, increased the diameter of subpericyte capillaries and increased the number of open capillaries 30 min after recanalization, improving cerebral reperfusion after stroke. | |
| PDGF-D [191] | Mice, In vitro | PDGF-D is a specific ligand for PDGFRβ, (PDGFR) β controls pericyte survival, migration, and interaction with brain endothelial cells, and PDGF-D promotes stable neovascularization of injured tissue and improves cerebral perfusion by acting on pericytes. | |
| Salvia miltiorrhiza (SAL), Panax notoginseng (PNS) [169] | In vitro | Suppressed oxidative stress and apoptosis, and simultaneously enhanced the release of pericyte-derived pro-angiogenic regulators, which are related to the PI3K/AKT/mTOR and JNK/ERK/P38 signaling pathways. | |
| Ginsenoside F1 (GF1) [170] | Rats, Zebrafish, In vitro | Activation of the Insulin-like Growth Factor 1 (IGF-1)/Insulin-like Growth Factor 1 Receptor (IGF1R) axis promotes the recruitment of pericytes into newly formed vessels to stabilize and induce maturation of the vascular system. | |
| Salvianolic acids (SALS) [171] | Mice | Activation of the Janus kinase 2/signal transducer and activator of transcription 3 (JAK2/STAT3) signaling pathway increases pericyte coverage in the peri-infarct area. | |
| Mitigation of BBB damage | Edaravone (Eda.B) [172] | Rats | Facilitates the multiplication of pericytes, augments the extent to which pericytes envelop ECs, and diminishes the generation of MMP-9. |
| Cilostazol [173] | Rats | Avoid the dissociation of pericytes from microvessels, promotes pericyte multiplication, and restrains the synthesis of MMP-9. Enhances pericyte coverage in the brain. | |
| Dimethyl malonate (DMM) [192] | Mice | Surge in pericyte numbers. | |
| Sigma-1 Receptor [174] | Mice, In vitro | Through the combined function of PDGFRβ and integrin α5β1, the recruitment of pericytes is regulated, which helps sustain and repair the BBB post-IS. | |
| Perlecan [175] | Mice | Through the combined function of PDGFRβ and integrin α5β1, the recruitment of pericytes is regulated, which helps sustain and repair the BBB post-IS. | |
| Estradiol [176,177] | In vitro | 17β-estradiol down-regulates miR-638 via ER-β to eliminate TNFα-induced pericyte migration in human cerebral vasculature. | |
| Electroacupuncture Serum [178] | Rats, In vitro | Prevents OGD/R-induced BBB damage in vitro by attenuating pericyte apoptosis and migration and enhancing pericyte viability. | |
| 3K3A-activated protein C (APC) [179] | Mice | Maintains pericyte coverage and reduces ischaemia-induced pericyte contraction. Inhibits nuclear factor-κB-mediated BBB degradation of MMP9 enzyme expression, inhibits inflammatory cytokines, and exerts direct neuroprotective activity. | |
| Metformin [180] | Mice | Inhibition of JNK p38 MAPK signaling activation in ischaemia/reperfusion injury prevents pericyte apoptosis and promotes nerve regeneration. | |
| Atorvastatin [181] | Rats | Promotes pericyte-mediated endothelial TJ formation, which plays an important role in remodeling the vascular system. | |
| immunomodulation | Bellidifolin [183] | In vitro | Suppresses the assembly of pro-inflammatory NLRP3/ASC/Caspase-1 complexes, thus safeguarding pericytes from undergoing cell death. |
5. Summary and Outlook
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| IS | Ischemic stroke |
| BBB | blood–brain barrier |
| EVT | endovascular therapy |
| NVU | neurovascular unit |
| ECs | endothelial cells |
| CNS | central nervous system |
| α-SMA | α-smooth muscle actin |
| PDGFRβ | platelet-derived growth factor receptor β |
| ANGPT1 | Angiopoietin1 |
| TGF-β | transforming growth factor-β |
| PTC | peritubular capillary |
| TME | tumor microenvironment TME |
| MSC | mesenchymal stem cell |
| CXCL12 | C-X-C motif chemokine ligand 12 |
| ECM | extracellular matrix |
| HSC | Hepatic stellate cells |
| CAF | cancer-associated fibroblasts |
| PMT | pericyte–myofibroblast transformation |
| AMPK | adenosine monophosphate AMP -activated protein kinase |
| PKM2 | Pericyte pyruvate kinase M2 |
| HKII | Hexokinase II |
| S1P | sphingosine 1-phosphate |
| Spns2 | spinster homolog 2 |
| MHC | major histocompatibility complex |
| PD-L1/2 | programmed cell death ligand 1/2 |
| LTBP1 | transforming growth factor β-binding protein 1 |
| TCMR | T-cell-mediated acute rejection |
| ABMR | antibody-mediated rejection |
| γ-PCs IFN | γ-activated pericyte |
| IDO1 | indoleamine 2,3-dioxygenase 1 |
| ABMR | Antibody-mediated rejection |
| ILC2 | type 2 innate lymphocytes |
| DCs | dendritic cells |
| GB | Glioblastoma |
| CMA | chaperone-mediated autophagy |
| RCC | renal cell carcinoma |
| MMP | metalloproteinases |
| FLT1 | fms-like tyrosine kinase 1 |
| TJs | tight junctions |
| HIF-1 | hypoxia-inducible factor-1 |
| Slc22a8 | solute carrier family 22 member 8 |
| IL6 | interleukin 6 |
| MGs/MΦs | microglia/macrophages |
| SEMA4D | Semaphorin 4D |
| DAMPs | damage-associated molecular patterns |
| GLAST | glutamate–aspartate transporter |
References
- Duan, H.; Li, S.; Hao, P.; Hao, F.; Zhao, W.; Gao, Y.; Qiao, H.; Gu, Y.; Lv, Y.; Bao, X.; et al. Activation of endogenous neurogenesis and angiogenesis by basic fibroblast growth factor-chitosan gel in an adult rat model of ischemic stroke. Neural Regen. Res. 2024, 19, 409–415. [Google Scholar] [CrossRef]
- Ding, J.S.; Zhang, Y.; Wang, T.Y.; Li, X.; Ma, C.; Xu, Z.M.; Sun, Q.; Xu, X.; Chen, G. Therapeutic applications of hydrogen sulfide and novel donors for cerebral ischemic stroke: A narrative review. Med. Gas Res. 2023, 13, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qi, T.; Li, L.; Pan, Y.; Huang, L.; Zhu, L.; Zhang, D.; Ma, X.; Qin, Y. MRPS9-Mediated Regulation of the PI3K/Akt/mTOR Pathway Inhibits Neuron Apoptosis and Protects Ischemic Stroke. J. Mol. Neurosci. 2024, 74, 23. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.W.; Yoon, C.E.; Kwon, I.; Lee, K.O.; Kim, J.; Kim, Y.D.; Heo, J.H.; Nam, H.S. Mild hypercapnia before reperfusion reduces ischemia-reperfusion injury in hyperacute ischemic stroke rat model. J. Cereb. Blood Flow Metab. 2025, 45, 664–676. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.Y.; Guo, Z.N.; Zhang, D.H.; Qu, Y.; Jin, H. The Role of Pericytes in Ischemic Stroke: Fom Cellular Functions to Therapeutic Targets. Front. Mol. Neurosci. 2022, 15, 866700. [Google Scholar] [CrossRef]
- Sun, M.; Chen, X.; Yin, Y.X.; Gao, Y.; Zhang, L.; Chen, B.; Ji, Y.; Fukunaga, K.; Han, F.; Lu, Y.M. Role of pericyte-derived SENP1 in neuronal injury after brain ischemia. CNS Neurosci. Ther. 2020, 26, 815–828. [Google Scholar] [CrossRef]
- Zhang, Y.; Mu, B.R.; Ran, Z.; Zhu, T.; Huang, X.; Yang, X.; Wang, D.M.; Ma, Q.H.; Lu, M.H. Pericytes in Alzheimer’s disease: Key players and therapeutic targets. Exp. Neurol. 2024, 379, 114825. [Google Scholar] [CrossRef]
- Klouda, T.; Kim, Y.; Baek, S.H.; Bhaumik, M.; Li, Y.; Liu, Y.; Wu, J.C.; Raby, B.A.; Perez, V.J.; Yuan, K. Specialized pericyte subtypes in the pulmonary capillaries. EMBO J. 2025, 44, 1074–1106. [Google Scholar] [CrossRef]
- Holm, A.; Heumann, T.; Augustin, H.G. Microvascular Mural Cell Organotypic Heterogeneity and Functional Plasticity. Trends Cell Biol. 2018, 28, 302–316. [Google Scholar] [CrossRef]
- van Splunder, H.; Villacampa, P.; Martínez-Romero, A.; Graupera, M. Pericytes in the disease spotlight. Trends Cell Biol. 2024, 34, 58–71. [Google Scholar] [CrossRef]
- Baek, S.H.; Maiorino, E.; Kim, H.; Glass, K.; Raby, B.A.; Yuan, K. Single Cell Transcriptomic Analysis Reveals Organ Specific Pericyte Markers and Identities. Front. Cardiovasc. Med. 2022, 9, 876591. [Google Scholar] [CrossRef] [PubMed]
- Muhl, L.; Mocci, G.; Pietilä, R.; Liu, J.; He, L.; Genové, G.; Leptidis, S.; Gustafsson, S.; Buyandelger, B.; Raschperger, E.; et al. A single-cell transcriptomic inventory of murine smooth muscle cells. Dev. Cell 2022, 57, 2426–2443. [Google Scholar] [CrossRef] [PubMed]
- Muhl, L.; Genové, G.; Leptidis, S.; Liu, J.; He, L.; Mocci, G.; Sun, Y.; Gustafsson, S.; Buyandelger, B.; Chivukula, I.V.; et al. Single-cell analysis uncovers fibroblast heterogeneity and criteria for fibroblast and mural cell identification and discrimination. Nat. Commun. 2020, 11, 3953. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.C.; Vest, R.T.; Kern, F.; Lee, D.P.; Agam, M.; Maat, C.A.; Losada, P.M.; Chen, M.B.; Schaum, N.; Khoury, N.; et al. A human brain vascular atlas reveals diverse mediators of Alzheimer’s risk. Nature 2022, 603, 885–892. [Google Scholar] [CrossRef]
- Winkler, E.A.; Kim, C.N.; Ross, J.M.; Garcia, J.H.; Gil, E.; Oh, I.; Chen, L.Q.; Wu, D.; Catapano, J.S.; Raygor, K.; et al. A single-cell atlas of the normal and malformed human brain vasculature. Science 2022, 375, eabi7377. [Google Scholar] [CrossRef]
- Sziraki, A.; Zhong, Y.; Neltner, A.M.; Niedowicz, D.M.; Rogers, C.B.; Wilcock, D.M.; Nehra, G.; Neltner, J.H.; Smith, R.R.; Hartz, A.M.; et al. A high-throughput single-cell RNA expression profiling method identifies human pericyte markers. Neuropathol. Appl. Neurobiol. 2023, 49, e12942. [Google Scholar] [CrossRef]
- Yin, G.N.; Ryu, J.K. Role of pericytes in regulating penile angiogenesis and nerve regeneration. Asian J. Androl. 2025, 27, 13–19. [Google Scholar] [CrossRef]
- Tang, Y.; Frisendahl, C.; Piltonen, T.T.; Arffman, R.K.; Lalitkumar, P.G.; Gemzell-Danielsson, K. Human Endometrial Pericytes: A Comprehensive Overview of Their Physiological Functions and Implications in Uterine Disorders. Cells 2024, 13, 1510. [Google Scholar] [CrossRef]
- Lin, L.; Hu, X.; Hong, W.; Pan, T.; Wang, Z.; Wang, E.; Wu, G. A novel animal model of spontaneous epilepsy: Cdk5 knockout in pericyte-specific mice. Front. Cell. Neurosci. 2024, 18, 1474231. [Google Scholar] [CrossRef]
- Villaseñor, R.; Kuennecke, B.; Ozmen, L.; Ammann, M.; Kugler, C.; Grüninger, F.; Loetscher, H.; Freskgård, P.O.; Collin, L. Region-specific permeability of the blood-brain barrier upon pericyte loss. J. Cereb. Blood Flow Metab. 2017, 37, 3683–3694. [Google Scholar] [CrossRef]
- Moro, M.; Balestrero, F.C.; Grolla, A. A Pericytes: Jack-of-all-trades in cancer-related inflammation. Front. Pharmacol. 2024, 15, 1426033. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, G.; Spitz, S.; Pramotton, F.M.; Tsai, A.; Li, B.M.; Wang, X.; Barr, O.M.; Ko, E.C.; Zhang, S.; Ashley, S.J.; et al. Engineered 3D human neurovascular model of Alzheimer’s disease to study vascular dysfunction. Biomaterials 2025, 314, 122864. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Huang, L.; Qu, Y.; Xiao, D.; Mu, D. Pericytes in Cerebrovascular Diseases: An Emerging Therapeutic Target. Front. Cell. Neurosci. 2019, 13, 519. [Google Scholar] [CrossRef] [PubMed]
- Faal, T.; Phan, D.T.T.; Davtyan, H.; Scarfone, V.M.; Varady, E.; Blurton-Jones, M.; Hughes, C.C.W.; Inlay, M.A. Induction of Mesoderm and Neural Crest-Derived Pericytes from Human Pluripotent Stem Cells to Study Blood-Brain Barrier Interactions. Stem Cell Rep. 2019, 12, 451–460. [Google Scholar] [CrossRef]
- Guo, X.; Xia, S.; Ge, T.; Lin, Y.; Hu, S.; Wu, H.; Xie, X.; Zhang, B.; Zhang, S.; Zeng, J.; et al. Atp13a5 Marker Reveals Pericyte Specification in the Mouse Central Nervous System. J. Neurosci. 2024, 44, e0727242024. [Google Scholar] [CrossRef]
- Alarcon-Martinez, L.; Yemisci, M.; Dalkara, T. Pericyte morphology and function. Histol. Histopathol. 2021, 36, 633–643. [Google Scholar]
- Cashion, J.M.; Brown, L.S.; Morris, G.P.; Fortune, A.J.; Courtney, J.M.; Makowiecki, K.; Premilovac, D.; Cullen, C.L.; Young, K.M.; Sutherland, B.A. Pericyte ablation causes hypoactivity and reactive gliosis in adult mice. Brain Behav. Immun. 2025, 123, 681–696. [Google Scholar] [CrossRef]
- Török, O.; Schreiner, B.; Schaffenrath, J.; Tsai, H.C.; Maheshwari, U.; Stifter, S.A.; Welsh, C.; Amorim, A.; Sridhar, S.; Utz, S.G.; et al. Pericytes regulate vascular immune homeostasis in the CNS. Proc. Natl. Acad. Sci. USA 2021, 118, e2016587118. [Google Scholar] [CrossRef]
- Girolamo, F.; Errede, M.; Bizzoca, A.; Virgintino, D.; Ribatti, D. Central Nervous System Pericytes Contribute to Health and Disease. Cells 2022, 11, 1707. [Google Scholar] [CrossRef]
- Fu, J.; Liang, H.; Yuan, P.; Wei, Z.; Zhong, P. Brain pericyte biology: From physiopathological mechanisms to potential therapeutic applications in ischemic stroke. Front. Cell. Neurosci. 2023, 17, 1267785. [Google Scholar] [CrossRef]
- Wang, T.; Li, M.Y.; Pei, Z.; Chen, Q.X.; Cheng, Q.S.; Li, Z. Down-regulation of platelet-derived growth factor receptor β in pericytes increases blood-brain barrier permeability and significantly enhances α-synuclein in a Parkinson’s Disease 3D cell model in vitro under hyperglycemic condition. Tissue Cell 2025, 93, 102751. [Google Scholar] [CrossRef] [PubMed]
- Huang, H. Pericyte-endothelial interactions in the retinal microvasculature. Int. J. Mol. Sci. 2020, 21, 7413. [Google Scholar] [CrossRef] [PubMed]
- Alimperti, S.; Mirabella, T.; Bajaj, V.; Polacheck, W.; Pirone, D.M.; Duffield, J.; Eyckmans, J.; Assoian, R.K.; Chen, C.S. Three-dimensional biomimetic vascular model reveals a RhoA, Rac1, and N-cadherin balance in mural cell-endothelial cell-regulated barrier function. Proc. Natl. Acad. Sci. USA 2017, 114, 8758–8763. [Google Scholar] [CrossRef] [PubMed]
- Campisi, M.; Shin, Y.; Osaki, T.; Hajal, C.; Chiono, V.; Kamm, R.D. 3D self-organized microvascular model of the human blood-brain barrier with endothelial cells, pericytes and astrocytes. Biomaterials 2018, 180, 117–129. [Google Scholar] [CrossRef]
- Li, G.; Gao, J.; Ding, P.; Gao, Y. The Role of Endothelial Cell–Pericyte Interactions in Vascularization and Diseases. J. Adv. Res. 2025, 67, 269–288. [Google Scholar] [CrossRef]
- Dave, J.M.; Chakraborty, R.; Agyemang, A.; Ntokou, A.; Saito, J.; Ballabh, P.; Martin, K.A.; Greif, D.M. Loss of TGFβ-Mediated Repression of Angiopoietin-2 in Pericytes Underlies Germinal Matrix Hemorrhage Pathogenesis. Stroke 2024, 55, 2340–2352. [Google Scholar] [CrossRef]
- Zhang, Y.; Neng, L.; Sharma, K.; Hou, Z.; Johnson, A.; Song, J.; Dabdoub, A.; Shi, X. Pericytes control vascular stability and auditory spiral ganglion neuron survival. eLife 2023, 12, e83486. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, W.; Ma, J.; He, H.; Liu, Z.; Zhu, X.; He, X.; He, J.; Chen, Z.; Jin, X.; et al. Pericyte signaling via soluble guanylate cyclase shapes the vascular niche and microenvironment of tumors. EMBO J. 2024, 43, 1519–1544. [Google Scholar] [CrossRef]
- Tefft, J.B.; Bays, J.L.; Lammers, A.; Kim, S.; Eyckmans, J.; Chen, C.S. Notch1 and Notch3 coordinate for pericyte-induced stabilization of vasculature. Am. J. Physiol. Cell Physiol. 2022, 322, C185–C196. [Google Scholar] [CrossRef]
- Bhowmick, S.; D’Mello, V.; Caruso, D.; Wallerstein, A.; Muneer, P.M.A. Impairment of pericyte-endothelium crosstalk leads to blood-brain barrier dysfunction following traumatic brain injury. Exp. Neurol. 2019, 317, 260–270. [Google Scholar] [CrossRef]
- Yin, G.N.; Jin, H.R.; Choi, M.J.; Limanjaya, A.; Ghatak, K.; Minh, N.N.; Ock, J.; Kwon, M.H.; Song, K.M.; Park, H.J.; et al. Pericyte-Derived Dickkopf2 Regenerates Damaged Penile Neurovasculature Through an Angiopoietin-1-Tie2 Pathway. Diabetes 2018, 67, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cui, Y.; Li, B.; Weng, J.; Wang, W.; Zhang, S.; Huang, X.; Guo, X.; Huang, Q. Advanced glycation end products induce immature angiogenesis in in vivo and ex vivo mouse models. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H519–H533. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Z.; Duan, J.; Gui, L.; Li, H.; Liang, X.; Tian, X.; Liu, K.; Li, Y.; Yang, J. H2O2-responsive VEGF/NGF gene co-delivery nano-system achieves stable vascularization in ischemic hindlimbs. J. Nanobiotechnol. 2022, 20, 145. [Google Scholar] [CrossRef] [PubMed]
- Burganova, G.; Bridges, C.; Thorn, P.; Landsman, L. The role of vascular cells in pancreatic beta-cell function. Front. Endocrinol. 2021, 12, 667170. [Google Scholar] [CrossRef]
- Aplin, A.C.; Aghazadeh, Y.; Mohn, O.G.; Hull-Meichle, R.L. Role of the Pancreatic Islet Microvasculature in Health and Disease. J. Histochem. Cytochem. 2024, 72, 711–728. [Google Scholar] [CrossRef]
- Shankar, A.S.; Du, Z.; Mora, H.T.; van den Bosch, T.P.P.; Korevaar, S.S.; Van den Berg-Garrelds, I.M.; Bindels, E.; Lopez-Iglesias, C.; Clahsen-van Groningen, M.C.; Gribnau, J.; et al. Human kidney organoids produce functional renin. Kidney Int. 2021, 99, 134–147. [Google Scholar] [CrossRef]
- Saifi, M.A.; Godugu, C. Inhibition of lysyl oxidase ameliorates renal injury by inhibiting CD44-mediated pericyte detachment and loss of peritubular capillaries. Life Sci. 2020, 243, 117294. [Google Scholar] [CrossRef]
- Xu, J.; Pu, J.; Chen, H.; Sun, L.; Fei, S.; Han, Z.; Tao, J.; Ju, X.; Wang, Z.; Tan, R.; et al. Role of microvascular pericyte dysfunction in antibody-mediated rejection following kidney transplantation. Ren. Fail. 2025, 47, 2458749. [Google Scholar] [CrossRef]
- Dasgupta, S.; Saha, A.; Ganguly, N.; Bhuniya, A.; Dhar, S.; Guha, I.; Ghosh, T.; Sarkar, A.; Ghosh, S.; Roy, K.; et al. NLGP regulates RGS5-TGFβ axis to promote pericyte-dependent vascular normalization during restricted tumor growth. FASEB J. 2022, 36, e22268. [Google Scholar] [CrossRef]
- Hu, X.; Ye, K.; Bo, S.; Xiao, Z.; Ma, M.; Pan, J.; Zhong, X.; Zhang, D.; Mo, X.; Yu, X.; et al. Monitoring imatinib decreasing pericyte coverage and HIF-1α level in a colorectal cancer model by an ultrahigh-field multiparametric MRI approach. J. Transl. Med. 2024, 22, 712. [Google Scholar] [CrossRef]
- Sakuma, R.; Minato, Y.; Maeda, S.; Yagi, H. Nrf2 phosphorylation contributes to acquisition of pericyte reprogramming via the PKCδ pathway. Neurobiol. Dis. 2025, 206, 106824. [Google Scholar] [CrossRef] [PubMed]
- Polat, S.; Yazir, Y.; Duruksu, G.; Kiliç, K.C.; Mert, S.; Gacar, G.; Öncel Duman, B.; Halbutoğullari, Z.S. Investigation of the differentiation potential of pericyte cells as an alternative source of mesenchymal stem cells. Acta Histochem. 2024, 126, 152145. [Google Scholar] [CrossRef] [PubMed]
- Potts, C.M.; Yang, X.; Lynes, M.D.; Malka, K.; Liaw, L. Exploration of Conserved Human Adipose Subpopulations Using Targeted Single-Nuclei RNA Sequencing Data Sets. J. Am. Heart Assoc. 2025, 14, e038465. [Google Scholar] [CrossRef] [PubMed]
- Fancello, I.; Willett, S.; Castiglioni, C.; Amer, S.; Santoleri, S.; Bragg, L.; Galli, F.; Cossu, G. TNAP expressing adventitial pericytes contribute to myogenesis during foetal development. Vascul. Pharmacol. 2025, 159, 107489. [Google Scholar] [CrossRef]
- Moriggi, M.; Torretta, E.; Cescon, M.; Russo, L.; Gregorio, I.; Braghetta, P.; Sabatelli, P.; Faldini, C.; Merlini, L.; Gargioli, C.; et al. Characterization of Proteome Changes in Aged and Collagen VI-Deficient Human Pericyte Cultures. Int. J. Mol. Sci. 2024, 25, 7118. [Google Scholar] [CrossRef]
- Alex, L.; Tuleta, I.; Hernandez, S.C.; Hanna, A.; Venugopal, H.; Astorkia, M.; Humeres, C.; Kubota, A.; Su, K.; Zheng, D.; et al. Cardiac Pericytes Acquire a Fibrogenic Phenotype and Contribute to Vascular Maturation After Myocardial Infarction. Circulation 2023, 148, 882–898. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The fate and role of the pericytes in myocardial diseases. Eur. J. Clin. Investig. 2024, 54, e14204. [Google Scholar] [CrossRef]
- Cao, Y.; Ni, Q.; Bao, C.; Cai, C.; Wang, T.; Ruan, X.; Li, Y.; Wang, H.; Wang, R.; Sun, W. The Role of Pericyte Migration and Osteogenesis in Periodontitis. J. Dent. Res. 2024, 103, 723–733. [Google Scholar] [CrossRef]
- Choi, H.; Jeong, B.C.; Hur, S.W.; Kim, J.W.; Lee, K.B.; Koh, J.T. The Angiopoietin-1 Variant COMP-Ang1 Enhances BMP2-Induced Bone Regeneration with Recruiting Pericytes in Critical Sized Calvarial Defects. PLoS ONE 2015, 10, e0140502. [Google Scholar] [CrossRef]
- Kudaibergen, G.; Mukhlis, S.; Mukhambetova, A.; Issabekova, A.; Sekenova, A.; Sarsenova, M.; Temirzhan, A.; Baidarbekov, M.; Umbayev, B.; Ogay, V. Repair of Rat Calvarial Critical-Sized Defects Using Heparin-Conjugated Fibrin Hydrogel Containing BMP-2 and Adipose-Derived Pericytes. Bioengineering 2024, 11, 437. [Google Scholar] [CrossRef]
- Dias, D.O.; Kalkitsas, J.; Kelahmetoglu, Y.; Estrada, C.P.; Tatarishvili, J.; Holl, D.; Jansson, L.; Banitalebi, S.; Amiry-Moghaddam, M.; Ernst, A.; et al. Pericyte-derived fibrotic scarring is conserved across diverse central nervous system lesions. Nat. Commun. 2021, 12, 5501. [Google Scholar] [CrossRef] [PubMed]
- Holl, D.; Göritz, C. Decoding fibrosis in the human central nervous system. Am. J. Physiol. Cell Physiol. 2023, 325, C1415–C1420. [Google Scholar] [CrossRef] [PubMed]
- Kuppe, C.; Kramann, R. Role of mesenchymal stem cells in kidney injury and fibrosis. Curr. Opin. Nephrol. Hypertens. 2016, 25, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Castellano, G.; Stasi, A.; Franzin, R.; Sallustio, F.; Divella, C.; Spinelli, A.; Netti, G.S.; Fiaccadori, E.; Cantaluppi, V.; Crovace, A.; et al. LPS-Binding Protein Modulates Acute Renal Fibrosis by Inducing Pericyte-to-Myofibroblast Trans-Differentiation through TLR-4 Signaling. Int. J. Mol. Sci. 2019, 20, 3682. [Google Scholar] [CrossRef]
- Castellano, G.; Franzin, R.; Stasi, A.; Divella, C.; Sallustio, F.; Pontrelli, P.; Lucarelli, G.; Battaglia, M.; Staffieri, F.; Crovace, A.; et al. Complement activation during ischemia/reperfusion injury induces pericyte-to-myofibroblast transdifferentiation regulating peritubular capillary Lumen Reduction Through pERK Signaling. Front. Immunol. 2018, 9, 1002. [Google Scholar] [CrossRef]
- Zhong, Y.; Wei, B.; Wang, W.; Chen, J.; Wu, W.; Liang, L.; Huang, X.R.; Szeto, C.C.; Yu, X.; Nikolic-Paterson, D.J.; et al. Single-cell RNA-Sequencing identifies bone marrow-derived progenitor cells as a main source of Extracellular Matrix-Producing cells across multiple organ-based Fibrotic diseases. Int. J. Biol. Sci. 2024, 20, 5027–5042. [Google Scholar] [CrossRef]
- Mierzejewski, B.; Różycka, J.; Stremińska, W.; Brągiel-Pieczonka, A.; Sidor, K.; Hoser, G.; Bartoszewicz, Z.; Gewartowska, M.; Frontczak-Baniewicz, M.; Ciemerych, M.A.; et al. The Role of Pericytes in Lipopolysaccharide-Induced Murine Acute Respiratory Distress Syndrome. Am. J. Pathol. 2024, 194, 1443–1457. [Google Scholar] [CrossRef]
- Tan, H.L.; Yap, J.Q.; Qian, Q. Acute kidney injury: Tubular markers and risk for chronic kidney disease and end-stage kidney failure. Blood Purif. 2016, 41, 144–150. [Google Scholar] [CrossRef]
- Castellano, G.; Franzin, R.; Stasi, A.; Divella, C.; Sallustio, F.; Pontrelli, P.; Lucarelli, G.; Battaglia, M.; Staffieri, F.; Crovace, A.; et al. Role of endothelial-to-mesenchymal transition induced by TGF-β1 in transplant kidney interstitial fibrosis. J. Cell. Mol. Med. 2017, 21, 2359–2369. [Google Scholar]
- Khan, M.A.; Fischer, J.; Harrer, L.; Schwiering, F.; Groneberg, D.; Friebe, A. Hepatic stellate cells in zone 1 engage in capillarization rather than myofibroblast formation in murine liver fibrosis. Sci. Rep. 2024, 14, 18840. [Google Scholar] [CrossRef]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Sererols-Viñas, L.; Garcia-Vicién, G.; Ruiz-Blázquez, P.; Lee, T.F.; Lee, Y.A.; Gonzalez-Sanchez, E.; Vaquero, J.; Moles, A.; Filliol, A.; Affò, S. Hepatic stellate cells functional heterogeneity in liver cancer. In Seminars in Liver Disease; Thieme Medical Publishers: New York, NY, USA, 2025. [Google Scholar]
- Rentsch, N.H.; Rust, R. ‘Scary’ pericytes: The fibrotic scar in brain and spinal cord lesions. Trends Neurosci. 2022, 45, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Luo, Y.; Liu, Y.C.; Chen, Y.H.; Li, Y.T.; Hu, X.Y.; You, X.Y.; Yu, S.S.; Li, Z.Y.; Chen, L.; et al. Imatinib inhibits pericyte-fibroblast transition and inflammation and promotes axon regeneration by blocking the PDGF-BB/PDGFRβ pathway in spinal cord injury. Inflamm. Regen. 2022, 42, 44. [Google Scholar] [CrossRef]
- Duan, Q.; Liu, Z.; Xing, Y.; Huang, H.; Zhu, L.; Liu, J.; He, P.; Ma, G.; Zhang, Y.; Nie, K.; et al. Edaravone dexborneol alleviates pericyte-mediated fibrosis depositing extracellular matrix through TGF-β1/IL-11 in cerebral small vessel disease. J. Transl. Med. 2025, 23, 161. [Google Scholar] [CrossRef]
- Xu, C.; Hong, Q.; Zhuang, K.; Ren, X.; Cui, S.; Dong, Z.; Wang, Q.; Bai, X.; Chen, X. Regulation of pericyte metabolic reprogramming restricts the AKI to CKD transition. Metabolism 2023, 145, 155592. [Google Scholar] [CrossRef]
- Chen, Y.; Bai, X.; Chen, J.; Huang, M.; Hong, Q.; Ouyang, Q.; Sun, X.; Zhang, Y.; Liu, J.; Wang, X.; et al. Pyruvate kinase M2 regulates kidney fibrosis through pericyte glycolysis during the progression from acute kidney injury to chronic kidney disease. Cell Prolif. 2024, 57, e13548. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Deng, Y.; Chen, J.; Huang, M.; Zhu, F.; Gao, Z.; Wu, L.; Hong, Q.; Feng, Z.; et al. The PI3K-Akt-mTOR pathway mediates renal pericyte-myofibroblast transition by enhancing glycolysis through HKII. J. Transl. Med. 2023, 21, 323. [Google Scholar] [CrossRef]
- Maniaci, A.; Briglia, M.; Allia, F.; Montalbano, G.; Romano, G.L.; Zaouali, M.A.; H’mida, D.; Gagliano, C.; Malaguarnera, R.; Lentini, M.; et al. The Role of Pericytes in Inner Ear Disorders: A Comprehensive Review. Biology 2024, 13, 802. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Andreeva, E.R.; Eremin, I.I.; Markin, A.M.; Nadelyaeva, I.I.; Orekhov, A.N.; Melnichenko, A.A. The Role of Pericytes in Regulation of Innate and Adaptive Immunity. Biomedicines 2023, 11, 600. [Google Scholar] [CrossRef]
- Tanaka, S. Targeting inflammation in perivascular cells and neuroimmune interactions for treating kidney disease. Clin. Exp. Nephrol. 2024, 28, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, D.K.; Bhattacharya, A.; Lozinski, B.M.; Wee Yong, V. Pericytes as mediators of infiltration of macrophages in multiple sclerosis. J. Neuroinflamm. 2021, 18, 301. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.F.; Mittelsteadt, K.L.; Brauer, R.; McKinney, B.L.; Hallstrand, T.S.; Parks, W.C.; Chen, P.; Schnapp, L.M.; Liles, W.C.; Duffield, J.S.; et al. Lung pericyte-like cells are functional interstitial immune sentinel cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L556–L567. [Google Scholar] [CrossRef] [PubMed]
- Cembran, A.; Eisenbaum, M.; Paris, D.; Mullan, M.; Crawford, F.; Ferguson, S.; Bachmeier, C. Contribution of brain pericytes to neuroinflammation following repetitive head trauma. J. Inflamm. 2025, 22, 11. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, B.; Sheng, C.; Yao, C.; Liu, Y.; Wang, J.; Zeng, Q.; Mao, Y.; Bei, J.; Zhu, B.; et al. SHISA3 Reprograms Tumor-Associated Macrophages Toward an Antitumoral Phenotype and Enhances Cancer Immunotherapy. Adv. Sci. 2024, 11, e2403019. [Google Scholar] [CrossRef]
- Schmidt, C.; Harit, K.; Traidl, S.; Naumann, M.; Werfel, T.; Roesner, L.M.; Nishanth, G.; Schlüter, D. Ablation of the deubiquitinating enzyme cylindromatosis (CYLD) augments STAT1-mediated M1 macrophage polarization and fosters Staphylococcus aureus control. Front. Immunol. 2025, 16, 1507989. [Google Scholar] [CrossRef]
- Domev, H.; Milkov, I.; Itskovitz-Eldor, J.; Dar, A. Immunoevasive pericytes from human pluripotent stem cells preferentially modulate induction of allogeneic regulatory T cells. Stem. Cells Transl. Med. 2014, 3, 1169–1181. [Google Scholar] [CrossRef]
- Li, A.; Li, Z.; Chiu, W.; Xiong, C.; Chen, Q.; Chen, J.; Lai, X.; Li, W.; Ke, Q.; Liu, J.; et al. Efficient Treatment of Pulpitis via Transplantation of Human Pluripotent Stem Cell-Derived Pericytes Partially through LTBP1-Mediated T Cell Suppression. Biomedicines 2023, 11, 3199. [Google Scholar] [CrossRef]
- Liu, R.; Merola, J.; Manes, T.D.; Qin, L.; Tietjen, G.T.; López-Giráldez, F.; Broecker, V.; Fang, C.; Xie, C.; Chen, P.M.; et al. Interferon-γ converts human microvascular pericytes into negative regulators of alloimmunity through induction of indoleamine 2,3-dioxygenase 1. JCI Insight 2018, 3, e97881. [Google Scholar] [CrossRef]
- Xu-Dubois, Y.; Kavvadas, P.; Keuylian, Z.; Hertig, A.; Rondeau, E.; Chatziantoniou, C. Notch3 expression in capillary pericytes predicts worse graft outcome in human renal grafts with antibody-mediated rejection. J. Cell. Mol. Med. 2022, 26, 3203–3212. [Google Scholar] [CrossRef]
- Schonblum, A.; Ali Naser, D.; Ovadia, S.; Egbaria, M.; Puyesky, S.; Epshtein, A.; Wald, T.; Mercado-Medrez, S.; Ashery-Padan, R.; Landsman, L. Beneficial islet inflammation in health depends on pericytic TLR/MyD88 signaling. J. Clin. Investig. 2024, 134, e179335. [Google Scholar] [CrossRef] [PubMed]
- Burganova, G.; Schonblum, A.; Sakhneny, L.; Epshtein, A.; Wald, T.; Tzaig, M.; Landsman, L. Pericytes modulate islet immune cells and insulin secretion through Interleukin-33 production in mice. Front. Endocrinol. 2023, 14, 1142988. [Google Scholar] [CrossRef] [PubMed]
- Dalmas, E.; Lehmann, F.M.; Dror, E.; Wueest, S.; Thienel, C.; Borsigova, M.; Stawiski, M.; Traunecker, E.; Lucchini, F.C.; Dapito, D.H.; et al. Interleukin-33-Activated islet-resident innate lymphoid cells promote insulin secretion through myeloid cell retinoic acid production. Immunity 2017, 47, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhou, J.; Li, L.; Liao, S.; He, J.; Zhou, S.; Zhou, Y. Pericytes in the tumor microenvironment. Cancer Lett. 2023, 556, 216074. [Google Scholar] [CrossRef]
- Wu, Z.; Thierry, K.; Bachy, S.; Zhang, X.; Gamradt, P.; Hernandez-Vargas, H.; Mikaelian, I.; Tonon, L.; Pommier, R.; Zhao, Y.; et al. Pericyte stem cells induce Ly6G+ cell accumulation and immunotherapy resistance in pancreatic cancer. EMBO Rep. 2023, 24, e56524. [Google Scholar] [CrossRef]
- Li, Z.J.; He, B.; Domenichini, A.; Satiaputra, J.; Wood, K.H.; Lakhiani, D.D.; Bashaw, A.A.; Nilsson, L.M.; Li, J.; Bastow, E.R.; et al. Pericyte phenotype switching alleviates immunosuppression and sensitizes vascularized tumors to immunotherapy in preclinical models. J. Clin. Investig. 2024, 134, e179860. [Google Scholar] [CrossRef]
- Morris, A.B.; Adams, L.E.; Ford, M.L. Influence of T Cell Coinhibitory Molecules on CD8+ Recall Responses. Front. Immunol. 2018, 9, 1810. [Google Scholar] [CrossRef]
- Molina, M.L.; García-Bernal, D.; Salinas, M.D.; Rubio, G.; Aparicio, P.; Moraleda, J.M.; Martínez, S.; Valdor, R. Chaperone-Mediated Autophagy Ablation in Pericytes Reveals New Glioblastoma Prognostic Markers and Efficient Treatment Against Tumor Progression. Front. Cell Dev. Biol. 2022, 10, 797945. [Google Scholar] [CrossRef]
- Valdor, R.; García-Bernal, D.; Riquelme, D.; Martinez, C.M.; Moraleda, J.M.; Cuervo, A.M.; Macian, F.; Martinez, S. Glioblastoma ablates pericytes antitumor immune function through aberrant up-regulation of chaperone-mediated autophagy. Proc. Natl. Acad. Sci. USA 2019, 116, 20655–20665. [Google Scholar] [CrossRef]
- Lu, T.; Zhang, J.; Lu, S.; Yang, F.; Gan, L.; Wu, X.; Song, H.; Liu, S.; Xu, C.; Han, D.; et al. Correction to: Endosialin-positive tumor-derived pericytes promote tumor progression through impeding the infiltration of CD8+ T cells in clear cell renal cell carcinoma. Cancer Immunol. Immunother. 2023, 72, 1751. [Google Scholar] [CrossRef]
- Benarroch, E. What Are the Roles of Pericytes in the Neurovascular Unit and Its Disorders? Neurology 2023, 100, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Erdener, Ş.E.; Tang, J.; Kılıç, K.; Postnov, D.; Giblin, J.T.; Kura, S.; Chen, I.A.; Vayisoğlu, T.; Sakadžić, S.; Schaffer, C.B. Dynamic capillary stalls in reperfused ischemic penumbra contribute to injury: A hyperacute role for neutrophils in persistent traffic jams. J. Cereb. Blood Flow Metab. 2021, 41, 236–252. [Google Scholar] [CrossRef] [PubMed]
- Presa, J.L.; Saravia, F.; Bagi, Z.; Filosa, J.A. Vasculo-neuronal coupling and neurovascular coupling at the neurovascular unit: Impact of hypertension. Front. Physiol. 2020, 11, 584135. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, D.A.; Coelho-Santos, V.; Shih, A.Y. Pericyte Control of Blood Flow Across Microvascular Zones in the Central Nervous System. Annu. Rev. Physiol. 2022, 84, 331–354. [Google Scholar] [CrossRef]
- Dalkara, T.; Alarcon-Martinez, L.; Yemisci, M. Pericytes in ischemic stroke. Adv. Exp. Med. Biol. 2019, 1147, 189–213. [Google Scholar]
- Zhang, S.; Liao, X.J.; Wang, J.; Shen, Y.; Shi, H.F.; Zou, Y.; Ma, C.Y.; Wang, X.Q.; Wang, Q.G.; Wang, X.; et al. Temporal alterations in pericytes at the acute phase of ischemia/reperfusion in the mouse brain. Neural Regen. Res. 2022, 17, 2247–2252. [Google Scholar]
- Kunze, R.; Wacker, P.; Breuer, P.; Nasyrov, E.; Kur, I.M.; Weigert, A.; Wagner, A.H.; Marti, H.H.; Korff, T. Adequate post-ischemic reperfusion of the mouse brain requires endothelial NFAT5. Acta Neuropathol. Commun. 2024, 12, 200. [Google Scholar] [CrossRef]
- Lochhead, J.J.; Williams, E.I.; Reddell, E.S.; Dorn, E.; Ronaldson, P.T.; Davis, T.P. High Resolution Multiplex Confocal Imaging of the Neurovascular Unit in Health and Experimental Ischemic Stroke. Cells 2023, 12, 645. [Google Scholar] [CrossRef]
- Feng, Z.; Fang, C.; Ma, Y.; Chang, J. Obesity-induced blood-brain barrier dysfunction: Phenotypes and mechanisms. J. Neuroinflamm. 2024, 21, 110. [Google Scholar] [CrossRef]
- Omorphos, N.P.; Gao, C.; Tan, S.S.; Sangha, M.S. Understanding angiogenesis and the role of angiogenic growth factors in the vascularisation of engineered tissues. Mol. Biol. Rep. 2021, 48, 941–950. [Google Scholar] [CrossRef]
- Kolbinger, A.; Kestner, R.I.; Jencio, L.; Schäufele, T.J.; Vutukuri, R.; Pfeilschifter, W.; Scholich, K. Behind the Wall-Compartment-Specific Neovascularisation during Post-Stroke Recovery in Mice. Cells 2022, 11, 1659. [Google Scholar] [CrossRef]
- Ghori, A.; Prinz, V.; Nieminen-Kehlä, M.; Bayerl, S.H.; Kremenetskaia, I.; Riecke, J.; Krechel, H.; Broggini, T.; Scherschinski, L.; Licht, T.; et al. Vascular Endothelial Growth Factor Augments the Tolerance Towards Cerebral Stroke by Enhancing Neurovascular Repair Mechanism. Transl. Stroke Res. 2022, 13, 774–791. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Liu, H.; Zhao, J.; Chen, L.Y.; Chen, J.; Lu, Z.; Hu, X. Pericytes in Brain Injury and Repair After Ischemic Stroke. Transl. Stroke Res. 2017, 8, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Payne, L.B.; Zhao, H.; James, C.C.; Darden, J.; McGuire, D.; Taylor, S.; Smyth, J.W.; Chappell, J.C. The pericyte microenvironment during vascular development. Microcirculation 2019, 26, e12554. [Google Scholar] [CrossRef] [PubMed]
- Dabravolski, S.A.; Markin, A.M.; Andreeva, E.R.; Eremin, I.I.; Orekhov, A.N.; Melnichenko, A.A. Emerging role of pericytes in therapy of cardiovascular diseases. Biomed. Pharmacother. 2022, 156, 113928. [Google Scholar] [CrossRef]
- Di, Z.; Wu, X.; Xie, W.; Lin, X. Effect of Pericytes on Cerebral Microvasculature at Different Time Points of Stroke. Biomed. Res. Int. 2021, 2021, 5281182. [Google Scholar] [CrossRef]
- Yang, Y.; Torbey, M.T. Angiogenesis and Blood-Brain Barrier Permeability in Vascular Remodeling after Stroke. Curr. Neuropharmacol. 2020, 18, 1250–1265. [Google Scholar] [CrossRef]
- Yang, Y.; Thompson, J.F.; Taheri, S.; Salayandia, V.M.; McAvoy, T.A.; Hill, J.W.; Yang, Y.; Estrada, E.Y.; Rosenberg, G.A. Early inhibition of MMP activity in ischemic rat brain promotes expression of tight junction proteins and angiogenesis during recovery. J. Cereb. Blood Flow Metab. 2013, 33, 1104–1114. [Google Scholar] [CrossRef]
- Stafford, P.; Mitra, S.; Debot, M.; Lutz, P.; Stem, A.; Hadley, J.; Hom, P.; Schaid, T.R.; Cohen, M.J. Astrocytes and pericytes attenuate severely injured patient plasma mediated expression of tight junction proteins in endothelial cells. PLoS ONE. 2022, 17, e0270817. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, Y.; Fan, X. Microvascular pericytes in brain-associated vascular disease. Biomed. Pharmacother. 2020, 121, 109633. [Google Scholar] [CrossRef]
- Singh, S.A.; Vellapandian, C. Structure of the Blood Brain Barrier and its Role in the Transporters for the Movement of Substrates across the Barriers. Curr. Drug Metab. 2023, 24, 250–269. [Google Scholar] [PubMed]
- Wang, L.; Xiong, X.; Zhang, L.; Shen, J. Neurovascular unit: A critical role in ischemic stroke. CNS Neurosci. Ther. 2021, 27, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Fan, H. Pericyte Loss in Diseases. Cells 2023, 12, 1931. [Google Scholar] [CrossRef] [PubMed]
- Enström, A.; Carlsson, R.; Özen, I.; Paul, G. RGS5: A novel role as a hypoxia-responsive protein that suppresses chemokinetic and chemotactic migration in brain pericytes. Biol. Open 2022, 11, bio059371. [Google Scholar] [CrossRef]
- Roth, M.; Carlsson, R.; Buizza, C.; Enström, A.; Paul, G. Pericyte response to ischemic stroke precedes endothelial cell death and blood-brain barrier breakdown. J. Cereb. Blood Flow Metab. 2025, 45, 617–629. [Google Scholar] [CrossRef]
- Mäe, M.A.; He, L.; Nordling, S.; Vazquez-Liebanas, E.; Nahar, K.; Jung, B.; Li, X.; Tan, B.C.; Chin Foo, J.; Cazenave-Gassiot, A.; et al. Single-Cell Analysis of Blood-Brain Barrier Response to Pericyte Loss. Circ. Res. 2021, 128, e123. [Google Scholar] [CrossRef]
- Baumann, J.; Tsao, C.C.; Patkar, S.; Huang, S.F.; Francia, S.; Magnussen, S.N.; Gassmann, M.; Vogel, J.; Köster-Hegmann, C.; Ogunshola, O.O. Pericyte, but not astrocyte, hypoxia inducible factor-1 (HIF-1) drives hypoxia-induced vascular permeability in vivo. Fluids Barriers CNS 2022, 19, 6. [Google Scholar] [CrossRef]
- Tsao, C.C.; Baumann, J.; Huang, S.F.; Kindler, D.; Schroeter, A.; Kachappilly, N.; Gassmann, M.; Rudin, M.; Ogunshola, O.O. Pericyte hypoxia-inducible factor-1 (HIF-1) drives blood-brain barrier disruption and impacts acute ischemic stroke outcome. Angiogenesis 2021, 24, 823–842. [Google Scholar] [CrossRef]
- Ogunshola, O.O.; Tsao, C.C. Harnessing the power of pericytes and hypoxia-inducible factor-1 to modulate stroke outcome. Neural Regen. Res. 2024, 19, 473–474. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Cao, C.; Ding, H.; Shi, X.; Zhang, J.; Li, H. Critical role of Slc22a8 in maintaining blood-brain barrier integrity after experimental cerebral ischemia-reperfusion. J. Cereb. Blood Flow Metab. 2025, 45, 85–101. [Google Scholar] [CrossRef]
- Bohannon, D.G.; Wellman, L.L.; Kaul, M.; Galkina, E.V.; Guo, M.L.; Datta, P.K.; Kim, W.K. Type-1-to-type-2 transition of brain microvascular pericytes induced by cytokines and disease-associated proteins: Role in neuroinflammation and blood-brain barrier disruption. J. Cereb. Blood Flow Metab. 2025, 45, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.M.; Abdelazim, H.; Jenkins-Houk, C.; Wright, D.; Patel, B.M.; Chappell, J.C. The cerebral microvasculature: Basic and clinical perspectives on stroke and glioma. Microcirculation 2021, 28, e12671. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhang, Y.; Zhong, Y.; Ye, Y.; Hu, X.; Gu, L.; Xiong, X. Inflammation-Mediated Angiogenesis in Ischemic Stroke. Front. Cell. Neurosci. 2021, 15, 652647. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Lindner, M.; Fleck, A.K.; Liebmann, M.; Eschborn, M.; Zondler, L.; Diéguez-Hurtado, R.; Adams, R.H.; Meyer Zu Hörste, G.; Zarbock, A.; et al. CNS Pericytes Modulate Local T Cell Infiltration in EAE. Int. J. Mol. Sci. 2022, 23, 13081. [Google Scholar] [CrossRef]
- Pieper, C.; Marek, J.J.; Unterberg, M.; Schwerdtle, T.; Galla, H.J. Brain capillary pericytes contribute to the immune defense in response to cytokines or LPS in vitro. Brain Res. 2014, 1550, 1–8. [Google Scholar] [CrossRef]
- Gautam, J.; Yao, Y. Roles of Pericytes in Stroke Pathogenesis. Cell Transplant. 2018, 27, 1798–1808. [Google Scholar] [CrossRef]
- Zapata-Acevedo, J.F.; Mantilla-Galindo, A.; Vargas-Sánchez, K.; González-Reyes, R.E. Blood-brain barrier biomarkers. Adv. Clin. Chem. 2024, 121, 1–88. [Google Scholar]
- Musafargani, S.; Mishra, S.; Gulyás, M.; Mahalakshmi, P.; Archunan, G.; Padmanabhan, P.; Gulyás, B. Blood brain barrier: A tissue engineered microfluidic chip. J. Neurosci. Methods 2020, 331, 108525. [Google Scholar] [CrossRef]
- Kempuraj, D.; Ahmed, M.E.; Selvakumar, G.P.; Thangavel, R.; Dhaliwal, A.S.; Dubova, I.; Mentor, S.; Premkumar, K.; Saeed, D.; Zahoor, H.; et al. Brain Injury-Mediated Neuroinflammatory Response and Alzheimer’s Disease. Neuroscientist 2020, 26, 134–155. [Google Scholar] [CrossRef]
- Buizza, C.; Enström, A.; Carlsson, R.; Paul, G. The Transcriptional Landscape of Pericytes in Acute Ischemic Stroke. Transl. Stroke Res. 2024, 15, 714–728. [Google Scholar] [CrossRef]
- Rehnström, M.; Frederiksen, S.D.; Ansar, S.; Edvinsson, L. Transcriptome profiling revealed early vascular smooth muscle cell gene activation following focal ischemic stroke in female rats—Comparisons with males. BMC Genom. 2020, 21, 883. [Google Scholar] [CrossRef] [PubMed]
- Lemarchant, S.; Dunghana, H.; Pomeshchik, Y.; Leinonen, H.; Kolosowska, N.; Korhonen, P.; Kanninen, K.M.; García-Berrocoso, T.; Montaner, J.; Malm, T.; et al. Anti-inflammatory effects of ADAMTS-4 in a mouse model of ischemic stroke. Glia 2016, 64, 1492–1507. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Qiu, W.; Yang, Y.; Huang, H.; Dai, Z.M.; Yang, A.; Tang, T.; Zhao, X.; Qiu, M. ADAMTS4 Enhances Oligodendrocyte Differentiation and Remyelination by Cleaving NG2 Proteoglycan and Attenuating PDGFRα Signaling. J. Neurosci. 2023, 43, 4405–4417. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Zhou, Y.; Chen, M.; Li, L.; Zhang, W. Pericytes for Therapeutic Approaches to Ischemic Stroke. Front. Neurosci. 2021, 15, 629297. [Google Scholar] [CrossRef]
- Kempuraj, D.; Dourvetakis, K.D.; Cohen, J.; Valladares, D.S.; Joshi, R.S.; Kothuru, S.P.; Anderson, T.; Chinnappan, B.; Cheema, A.K.; Klimas, N.G.; et al. Neurovascular unit, neuroinflammation and neurodegeneration markers in brain disorders. Front. Cell. Neurosci. 2024, 18, 1491952. [Google Scholar] [CrossRef]
- Nakagomi, T.; Narita, A.; Nishie, H.; Nakano-Doi, A.; Sawano, T.; Fukuda, Y.; Matsuyama, T. L-PGDS-PGD2-DP1 Axis Regulates Phagocytosis by CD36+ MGs/MΦs That Are Exclusively Present Within Ischemic Areas After Stroke. Cells 2024, 13, 1737. [Google Scholar] [CrossRef]
- Shibahara, T.; Ago, T.; Tachibana, M.; Nakamura, K.; Yamanaka, K.; Kuroda, J.; Wakisaka, Y.; Kitazono, T. Reciprocal Interaction Between Pericytes and Macrophage in Poststroke Tissue Repair and Functional Recovery. Stroke 2020, 51, 3095–3106. [Google Scholar] [CrossRef]
- Wu, B.; Zhou, D.; Mei, Z. Targeting the neurovascular unit: Therapeutic potential of traditional Chinese medicine for the treatment of stroke. Heliyon 2024, 10, e38200. [Google Scholar] [CrossRef]
- Gaceb, A.; Roupé, L.; Enström, A.; Almasoudi, W.; Carlsson, R.; Lindgren, A.G.; Paul, G. Pericyte Microvesicles as Plasma Biomarkers Reflecting Brain Microvascular Signaling in Patients with Acute Ischemic Stroke. Stroke 2024, 55, 558–568. [Google Scholar] [CrossRef]
- Qiu, Y.M.; Zhang, C.L.; Chen, A.Q.; Wang, H.L.; Zhou, Y.F.; Li, Y.N.; Hu, B. Immune Cells in the BBB Disruption After Acute Ischemic Stroke: Targets for Immune Therapy? Front. Immunol. 2021, 12, 678744. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Li, Y.N.; Jin, H.J.; Wu, J.H.; He, Q.W.; Wang, X.X.; Lei, H.; Hu, B. Sema4d/PlexinB1 Inhibition Ameliorates Blood-Brain Barrier Damage and Improves Outcome After Stroke in Rats. FASEB J. 2018, 32, 2181–2196. [Google Scholar] [CrossRef] [PubMed]
- Nirwane, A.; Johnson, J.; Nguyen, B.; Miner, J.H.; Yao, Y. Mural Cell-Derived Laminin-α5 Plays a Detrimental Role in Ischemic Stroke. Acta Neuropathol. Commun. 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yu, Z.; Tian, J.; Chen, T.; Wei, A.; Mei, C.; Chen, S.; Li, Y. m6A RNA modification pathway: Orchestrating fibrotic mechanisms across multiple organs. Brief. Funct. Genomics 2025, 24, elae051. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.; Liu, Y.; Peng, S.; Cai, Z.; Tang, G.; Zhou, Z.; Hu, K.; Zhong, Y. Cerebral [18F]AIF-FAPI-42-Based PET Imaging of Fibroblast Activation Protein for Non-invasive Quantification of Fibrosis After Ischemic Stroke. Transl. Stroke Res. 2024, 16, 848–858. [Google Scholar] [CrossRef]
- Dorrier, C.E.; Aran, D.; Haenelt, E.A.; Sheehy, R.N.; Hoi, K.K.; Pintarić, L.; Chen, Y.; Lizama, C.O.; Cautivo, K.M.; Weiner, G.A.; et al. CNS fibroblasts form a fibrotic scar in response to immune cell infiltration. Nat. Neurosci. 2021, 24, 234–244. [Google Scholar] [CrossRef]
- Shao, Z.Q.; Dou, S.S.; Zhu, J.G.; Wang, H.Q.; Wang, C.M.; Cheng, B.H.; Bai, B. Apelin-13 inhibits apoptosis and excessive autophagy in cerebral ischemia/reperfusion injury. Neural Regen. Res. 2021, 16, 1044–1051. [Google Scholar]
- Makihara, N.; Arimura, K.; Ago, T.; Tachibana, M.; Nishimura, A.; Nakamura, K.; Matsuo, R.; Wakisaka, Y.; Kuroda, J.; Sugimori, H.; et al. Involvement of platelet-derived growth factor receptor β in fibrosis through extracellular matrix protein production after ischemic stroke. Exp. Neurol. 2015, 264, 127–134. [Google Scholar] [CrossRef]
- Shibahara, T.; Ago, T.; Nakamura, K.; Tachibana, M.; Yoshikawa, Y.; Komori, M.; Yamanaka, K.; Wakisaka, Y.; Kitazono, T. Pericyte-Mediated Tissue Repair through PDGFRβ Promotes Peri-Infarct Astrogliosis, Oligodendrogenesis, and Functional Recovery after Acute Ischemic Stroke. eNeuro 2020, 7. [Google Scholar] [CrossRef]
- Shibahara, T.; Nakamura, K.; Wakisaka, Y.; Shijo, M.; Yamanaka, K.; Takashima, M.; Takaki, H.; Hidaka, M.; Kitazono, T.; Ago, T. PDGFRβ-positive cell-mediated post-stroke remodeling of fibronectin and laminin α2 for tissue repair and functional recovery. J. Cereb. Blood Flow Metab. 2023, 43, 518–530. [Google Scholar] [CrossRef]
- Huang, J.G.; Ren, J.X.; Chen, Y.; Tian, M.F.; Zhou, L.; Wen, J.; Song, X.S.; Wu, Y.L.; Yang, Q.H.; Jiang, P.R.; et al. M2 macrophages mediate fibrotic scar formation in the early stages after cerebral ischemia in rats. Neural Regen. Res. 2023, 18, 2208–2218. [Google Scholar]
- Dias, D.O.; Kim, H.; Holl, D.; Werne Solnestam, B.; Lundeberg, J.; Carlén, M.; Göritz, C.; Frisén, J. Reducing pericyte-derived scarring promotes recovery after spinal cord injury. Cell 2018, 173, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Cregg, J.M.; DePaul, M.A.; Filous, A.R.; Lang, B.T.; Tran, A.; Silver, J. Functional regeneration beyond the glial scar. Exp. Neurol. 2014, 253, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Leaf, I.A.; Nakagawa, S.; Johnson, B.G.; Cha, J.J.; Mittelsteadt, K.; Guckian, K.M.; Gomez, I.G.; Altemeier, W.A.; Duffield, J.S. Pericyte MyD88 and IRAK4 control inflammatory and fibrotic responses to tissue injury. J. Clin. Investig. 2017, 127, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Sancho, M.; Klug, N.R.; Mughal, A.; Koide, M.; Huerta de la Cruz, S.; Heppner, T.J.; Bonev, A.D.; Hill-Eubanks, D.; Nelson, M.T. Adenosine signaling activates ATP-sensitive K+ channels in endothelial cells and pericytes in CNS capillaries. Sci. Signal. 2022, 15, eabl5405. [Google Scholar] [CrossRef]
- Guo, R.B.; Dong, Y.F.; Yin, Z.; Cai, Z.Y.; Yang, J.; Ji, J.; Sun, Y.Q.; Huang, X.X.; Xue, T.F.; Cheng, H.; et al. Iptakalim improves cerebral microcirculation in mice after ischemic stroke by inhibiting pericyte contraction. Acta Pharmacol. Sin. 2022, 43, 1349–1359. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, Y.H.; Wang, X.M.; Chen, H.S. PI3Kδ inhibition alleviates the brain injury during cerebral ischemia reperfusion via suppressing pericyte contraction in a TNF-α dependent manner. Exp. Neurol. 2024, 375, 114728. [Google Scholar] [CrossRef]
- Beard, D.J.; Brown, L.S.; Morris, G.P.; Couch, Y.; Adriaanse, B.A.; Karali, C.S.; Schneider, A.M.; Howells, D.W.; Redzic, Z.B.; Sutherland, B.A.; et al. Rapamycin Treatment Reduces Brain Pericyte Constriction in Ischemic Stroke. Transl. Stroke Res. 2024. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, W.; Yang, J.; Xu, H.; Cao, Y.; Guo, L.; Sun, J.; Liang, B.; Du, X.; Chai, L.; et al. Components of Salvia miltiorrhiza and Panax notoginseng Protect Pericytes Against OGD/R-Induced Injury via Regulating the PI3K/AKT/mTOR and JNK/ERK/P38 Signaling Pathways. J. Mol. Neurosci. 2022, 72, 2377–2388. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, M.; Huang, M.; Chen, M.; Zhang, D.; Luo, L.; Ye, G.; Deng, L.; Peng, Y.; Wu, X.; et al. Ginsenoside F1 promotes angiogenesis by activating the IGF-1/IGF1R pathway. Pharmacol. Res. 2019, 144, 292–305. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Cui, L.; Chen, R.; Zhang, Y.; Zhang, C.; Zhu, X.; He, T.; Shen, Z.; Dong, L.; et al. Salvianolic acids enhance cerebral angiogenesis and neurological recovery by activating JAK2/STAT3 signaling pathway after ischemic stroke in mice. J. Neurochem. 2017, 143, 87–99. [Google Scholar] [CrossRef]
- Sun, Z.; Zhao, H.; Yang, S.; Liu, R.; Yi, L.; Gao, J.; Liu, S.; Chen, Y.; Zhang, Z. Edaravone Dexborneol protects against blood-brain barrier disruption following cerebral ischemia/reperfusion by upregulating pericyte coverage via vitronectin-integrin and PDGFB/PDGFR-β signaling. Free Radic. Biol. Med. 2024, 225, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Omote, Y.; Deguchi, K.; Kono, S.; Liu, N.; Liu, W.; Kurata, T.; Yamashita, T.; Ikeda, Y.; Abe, K. Neurovascular protection of cilostazol in stroke-prone spontaneous hypertensive rats associated with angiogenesis and pericyte proliferation. J. Neurosci. Res. 2014, 92, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Wei, Q.; Leng, S.; Li, C.; Han, B.; Bai, Y.; Zhang, H.; Yao, H. Activation of Sigma-1 Receptor Enhanced Pericyte Survival via the Interplay Between Apoptosis and Autophagy: Implications for Blood-Brain Barrier Integrity in Stroke. Transl. Stroke Res. 2020, 11, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Ikeuchi, T.; Nara, K.; Rhodes, C.S.; Zhang, P.; Chiba, Y.; Kazuno, S.; Miura, Y.; Ago, T.; Arikawa-Hirasawa, E.; et al. Perlecan regulates pericyte dynamics in the maintenance and repair of the blood-brain barrier. J. Cell Biol. 2019, 218, 3506–3525. [Google Scholar] [CrossRef]
- Kurmann, L.; Okoniewski, M.; Dubey, R.K. Estradiol Inhibits Human Brain Vascular Pericyte Migration Activity: A Functional and Transcriptomic Analysis. Cells 2021, 10, 2314. [Google Scholar] [CrossRef]
- Kurmann, L.; Azzarito, G.; Leeners, B.; Rosselli, M.; Dubey, R.K. 17β-Estradiol Abrogates TNF-α-Induced Human Brain Vascular Pericyte Migration by Downregulating miR-638 via ER-β. Int. J. Mol. Sci. 2024, 25, 11416. [Google Scholar] [CrossRef]
- Zhang, H.; Lyv, H.; Feng, Y.; Peng, Y. Electroacupuncture Serum Protects against Blood-brain Barrier Damage after Ischemic Stroke by Regulating Pericytes in vitro. Curr. Neurovasc. Res. 2025, 21, 491–502. [Google Scholar] [CrossRef]
- Wang, Y.; Kisler, K.; Nikolakopoulou, A.M.; Fernandez, J.A.; Griffin, J.H.; Zlokovic, B.V. 3K3A-Activated Protein C Protects the Blood-Brain Barrier and Neurons From Accelerated Ischemic Injury Caused by Pericyte Deficiency in Mice. Front. Neurosci. 2022, 16, 841916. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, D.; Lu, Z.; Man, J.; Zhang, Z.; Fu, X.; Cui, K.; Wang, J. Metformin protects against pericyte apoptosis and promotes neurogenesis through suppressing JNK p38 MAPK signalling activation in ischemia/reperfusion injury. Neurosci. Lett. 2022, 783, 136708. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, L.Y.; Salayandia, V.M.; Thompson, J.F.; Torbey, M.; Yang, Y. Treatment with Atorvastatin During Vascular Remodeling Promotes Pericyte-Mediated Blood-Brain Barrier Maturation Following Ischemic Stroke. Transl. Stroke Res. 2021, 12, 905–922. [Google Scholar] [CrossRef]
- Shen, J.; Xu, G.; Zhu, R.; Yuan, J.; Ishii, Y.; Hamashima, T.; Matsushima, T.; Yamamoto, S.; Takatsuru, Y.; Nabekura, J.; et al. PDGFR-β restores blood-brain barrier functions in a mouse model of focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2019, 39, 1501–1515. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Xu, W.; Ge, Y.; Ge, Y.; Zhang, L.; Zhu, Y.; Zhang, Z.; Fan, J.; Gao, R.; Jiang, L.; et al. Protection of Oxygen Glucose Deprivation-Induced Human Brain Vascular Pericyte Injury: Beneficial Effects of Bellidifolin in Cellular Pyroptosis. Neurochem. Res. 2023, 48, 2794–2807. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yang, L.; Li, Q.; Yang, X.; Chen, Z.; Zhou, Y.; Jia, Y.; Gong, Z. Long-Term Alcohol Exposure Aggravates Ischemic Stroke-Induced Damage by Promoting Pericyte NLRP3 Inflammasome Activation via Pre-Activating the TLR4/NF-κB Pathway in Rats. J. Inflamm. Res. 2024, 17, 4791–4810. [Google Scholar] [CrossRef] [PubMed]
- Almaghrabi, T.S.; McDonald, M.M.; Cai, C.; Rahbar, M.H.; Choi, H.A.; Lee, K.; Naval, N.S.; Grotta, J.C.; Chang, T.R. Cocaine Use is Associated with More Rapid Clot Formation and Weaker Clot Strength in Acute Stroke Patients. Int. J. Cerebrovasc. Dis. Stroke 2019, 2, 110. [Google Scholar]
- Niu, F.; Liao, K.; Hu, G.; Sil, S.; Callen, S.; Guo, M.L.; Yang, L.; Buch, S. Cocaine-induced release of CXCL10 from pericytes regulates monocyte transmigration into the CNS. J. Cell Biol. 2019, 218, 700–721. [Google Scholar] [CrossRef]
- Lu, G.; Du, R.; Liu, Y.; Zhang, S.; Li, J.; Pei, J. RGS5 as a Biomarker of Pericytes, Involvement in Vascular Remodeling and Pulmonary Arterial Hypertension. Vasc. Health Risk Manag. 2023, 19, 673–688. [Google Scholar] [CrossRef]
- Roth, M.; Gaceb, A.; Enström, A.; Padel, T.; Genové, G.; Özen, I.; Paul, G. Regulator of G-protein signaling 5 regulates the shift from perivascular to parenchymal pericytes in the chronic phase after stroke. FASEB J. 2019, 33, 8990–8998. [Google Scholar] [CrossRef]
- Liu, S.L.; Li, Y.H.; Shi, G.Y.; Jiang, M.J.; Chang, J.H.; Wu, H.L. The effect of statin on the aortic gene expression profiling. Int. J. Cardiol. 2007, 114, 71–77. [Google Scholar] [CrossRef]
- Kytö, V.; Åivo, J.; Ruuskanen, J.O. Intensity of statin therapy after ischaemic stroke and long-term outcomes: A nationwide cohort study. Stroke Vasc. Neurol. 2025, 10, 142–145. [Google Scholar] [CrossRef]
- Bernard, M.; Menet, R.; Lecordier, S.; ElAli, A. Endothelial PDGF-D contributes to neurovascular protection after ischemic stroke by rescuing pericyte functions. Cell. Mol. Life Sci. 2024, 81, 225. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, Z.; Liu, C.; Man, J.; Li, X.; Cui, K.; Lu, H.; Wang, J. Protective effects of Dimethyl malonate on neuroinflammation and blood-brain barrier after ischemic stroke. Neuroreport 2021, 32, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Zhao, W. Bidirectional Role of Pericytes in Ischemic Stroke. Brain Sci. 2025, 15, 605. https://doi.org/10.3390/brainsci15060605
Xu J, Zhao W. Bidirectional Role of Pericytes in Ischemic Stroke. Brain Sciences. 2025; 15(6):605. https://doi.org/10.3390/brainsci15060605
Chicago/Turabian StyleXu, Jingya, and Weiming Zhao. 2025. "Bidirectional Role of Pericytes in Ischemic Stroke" Brain Sciences 15, no. 6: 605. https://doi.org/10.3390/brainsci15060605
APA StyleXu, J., & Zhao, W. (2025). Bidirectional Role of Pericytes in Ischemic Stroke. Brain Sciences, 15(6), 605. https://doi.org/10.3390/brainsci15060605