Abstract
Background: Cognitive control (CC) underpins our ability to maintain task focus, update goals, and flexibly shift between strategies, and it is closely tied to prefrontal cortex (PFC) activity. Electronic gaming (e-gaming), such as the first-person shooter (FPS) genre, is a specialized domain that demands refined CC skills developed over years of practice. Although previous research has demonstrated that PFC hemodynamic activity can effectively evaluate CC in several skilled domains, the impact of prolonged FPS experience on CC and its underlying neural correlates remains unclear. Objectives: In this study, we examined differences in both behavioral performance and PFC hemodynamic responses between 70 novices and 50 experienced FPS gamers (n=120). Methods: We targeted three core CC subdomains—updating, shifting, and inhibition—by employing the Digit–Symbol Substitution Task, Dual Visual Search Task, and Stroop Task, respectively. Functional near-infrared spectroscopy (fNIRS)-based PFC activity was recorded as participants engaged in each task. Results: Experienced gamers showed higher levels of prefrontal neural efficiency for updating and shifting, but minimal differences for inhibitory control. Conclusions: These findings inform neuroergonomic approaches to performance evaluation and may be generalized to other complex, real-world environments that rely on extensive training for skill acquisition.
1. Introduction
Cognitive control (CC) encompasses a suite of executive functions essential for goal-directed behavior, flexible attention, and adaptive problem-solving. These functions critically depend on the prefrontal cortex (PFC) and its associated networks [1,2], which regulate the balance between deliberate and controlled, versus more habitual, automated processing. The PFC, in particular, is responsible for facilitating adaptive behavior with rapid decision making and response control in dynamic environments by orchestrating cognitive control functions, including attentional focus, strategic flexibility, and inhibitory control [2,3].
Within the broader CC framework, three subdomains have been extensively examined in cognitive neuroscience: (i) information updating (“updating”), involving the continuous monitoring and manipulation of relevant information in working memory; (ii) mental set-shifting (“shifting”), enabling rapid adaptation to changing rules or task demands; and (iii) inhibition of prepotent responses (“inhibition”), necessary for suppressing dominant yet inappropriate actions [4]. Each subdomain is typically probed by distinct cognitive tasks that elicit characteristic PFC activation patterns. For example, the Stroop task reliably recruits the dorsolateral PFC when participants override the habitual inclination to read words rather than name colors [5,6]. Shifting is commonly assessed with paradigms that require switching between task sets [7], while both the N-back and the Digit–Symbol Substitution Task (DSST) are known to engage lateral anterior PFC regions that subserve working memory and attentional control [8,9,10].
First-person shooter (FPS) games represent a unique segment of the electronic gaming (e-gaming) space, demanding complex and rapid decision making that appears to draw heavily on all three CC subdomains [11,12,13,14]. From a first-person vantage, players must rapidly identify moving targets, detect threats, and continually shift attention among multiple objectives and environmental cues. This interactive environment necessitates continual updating of one’s internal representation of the game state, integrating new information as it emerges. At the same time, impulsive actions may carry fewer immediate consequences, potentially diminishing the need for strong inhibitory control. Indeed, previous work has suggested that prolonged exposure to FPS gaming does not show improvement in inhibitory responses [11,13,14], possibly explaining why improvements in task probing inhibition (e.g., the Stroop task) are not consistently observed among experienced FPS players.
Despite growing interest in how gameplay experience shapes CC, neuroimaging work has primarily focused on brain changes during or directly following gameplay [15,16]. Although numerous studies have addressed how FPS experience can influence behavioral performance on CC tasks [7,11,12,13], few have examined its long-term neural correlates [17,18]. This leaves important questions open about whether (and how) prolonged FPS exposure might reshape the neurophysiological underpinnings of CC, especially within the PFC—the primary locus of executive function [19,20].
To bridge this gap, the present study combined three tasks known to regulate the CC subdomains of updating (DSST), shifting (DVST), and inhibition (Stroop) with functional near-infrared spectroscopy (fNIRS) to capture PFC hemodynamic responses in novice versus experienced FPS gamers. By isolating each subcomponent of CC, our design aimed to clarify the extent to which years of FPS gaming alter both performance and PFC engagement across the three CC domains. The results of this investigation will not only shed light on the neurocognitive mechanisms of FPS expertise but also inform wearable neuroimaging applications in other complex real-world contexts where specialized training is pivotal.
2. Materials and Methods
2.1. Participants
A total of 120 participants (30 females, mean age = 21.7 years, SD = ±3.8 years) volunteered for the study and received monetary compensation for their time. All confirmed that they met the eligibility requirements of being right-handed with vision correctable to 20/20, did not have a history of brain injury or psychological disorder, and were not on medication affecting brain activity. Prior to the study, all participants signed consent forms approved by the Institutional Review board of Drexel University (Philadelphia, PA, USA). All participants had previous e-game experience and were divided into novice and experienced groups based on a questionnaire given prior to the experiment to assess e-gaming background.
Participants were asked to report their gameplay frequency as well as their length of experience with FPS gaming. If participants reported FPS genre gameplay experience as part of their profile (more than 12 months), participants were also asked to report the average hours per week that they played FPS games. If they reported FPS gameplay experience equal to or greater than 10 h per week, they were admitted to the study and classified as an experienced FPS gamer. If they reported less than 10 h per week of gameplay they were classified as novice FPS gamers. If no FPS gameplay was reported, participants were admitted to the novice group. For all participants admitted to the study, the median value of reported weekly gameplay was 10 h. This value corresponds to the average reported gameplay of US adolescents [21] and aligns with the demarcating value specified by previous FPS gaming behavioral research [7], showing that 10 or more hours of weekly action videogame play improved performance in a behavioral task. Qualifying FPS games included titles such as Overwatch, Call of Duty, Counter Strike, Halo, Half-Life, Medal of Honor, Fortnite, and Rainbow Six.
2.2. Experimental Procedure
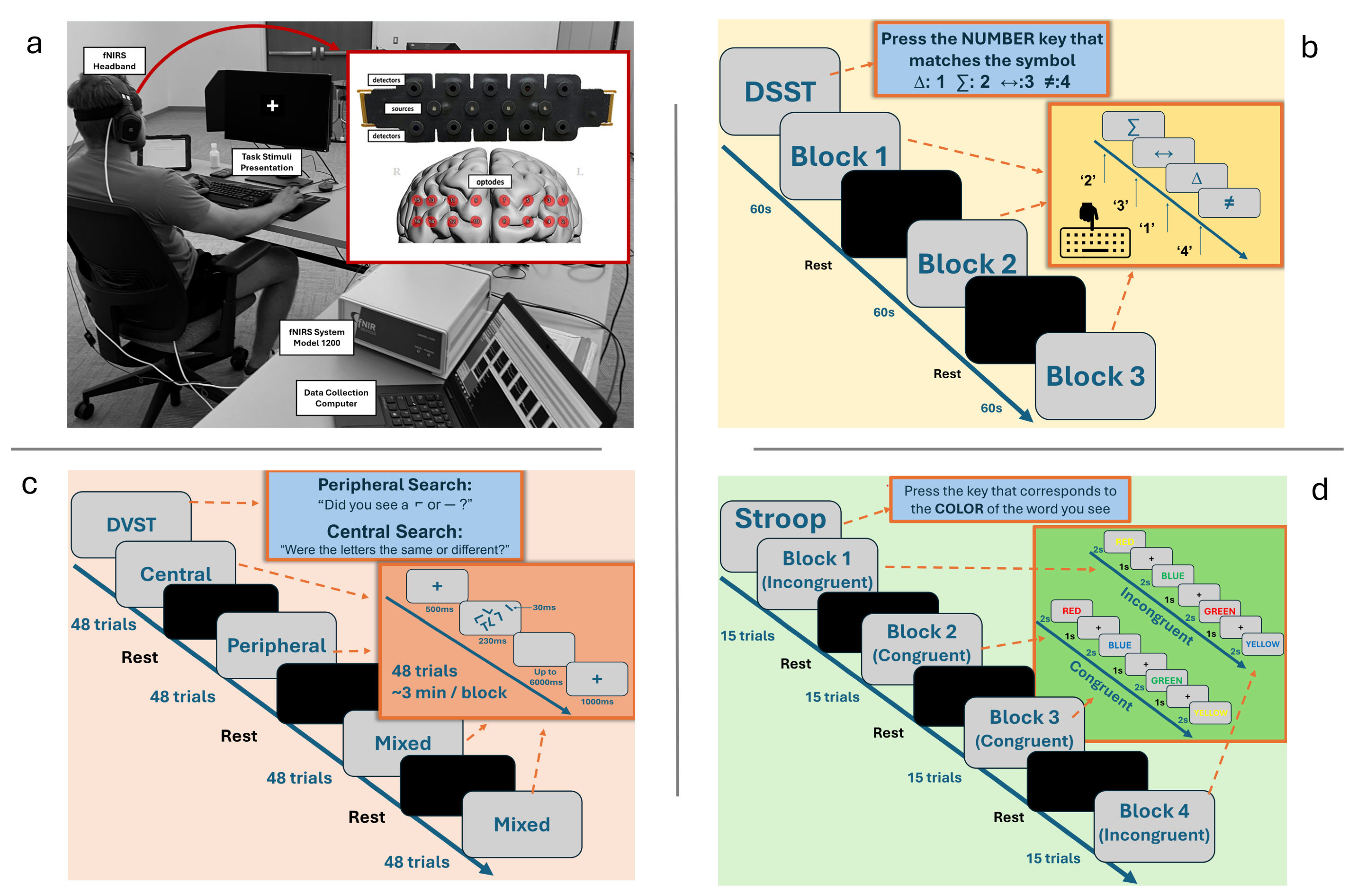
We designed a discrete laboratory cognitive task battery to assess the CC subdomains of shifting, updating, and inhibition using discrete laboratory tasks that were previously demonstrated to evoke responses in the PFC [3]. Participants performed this CC task battery over a thirty-minute session while seated 18 inches from a 24-in LED monitor. All participants were fitted with a 16-optode continuous wave fNIRS system Model 1200 (fNIR Devices, LLC., Potomac, MD, USA) positioned on the forehead to measure prefrontal cortex hemodynamic activity. After device set-up, participants completed a Digit–Symbol Substitution Task (DSST), a Dual Visual Search Task (DVST), and Stroop Task (Figure 1).
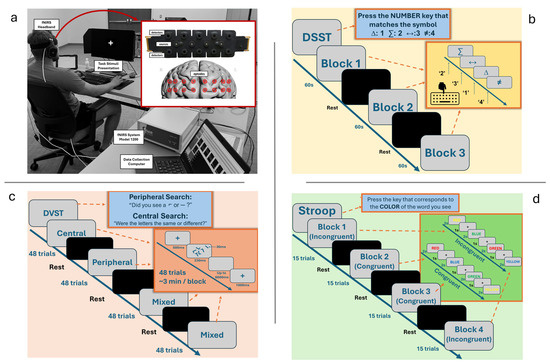
Figure 1.
Experiment set-up and schematic description of tasks. (a) Participant sitting in front of task stimuli presentation with fNIRS flat sensor pad on forehead and respective measurement areas represented on the prefrontal cortex. (b) DSST representative instructions and flow. (c) DVST representative instructions and flow. (d) The Stroop Task outline with examples of both congruent and incongruent trials included.
2.2.1. Digit–Symbol Substitution Task
Visual monitoring and speed of processing are fundamental characteristics of the FPS skillset as gameplay requires not only precise sensory–motor coordination but also a constant updating of goals and targets across the player’s visual field [22,23]. In three separate 1 min blocks of probe presentation (trials lasting a maximum duration of 2 s each), a symbol probe was presented visually at the center of the screen. Participants were required to press the number on their keyboard that corresponded to the correct symbol–digit pairing on a key at the top of their screen as quickly and as accurately as possible. Probe presentation and digit–symbol pairings were pseudorandomized across trials. Accuracy and reaction times were recorded, and participants received scores based on their number of correct responses per block.
2.2.2. Dual Visual Search Task
FPS players often detect and identify threats in the periphery, while being simultaneously engaged in central search tasks. These demands require the ability to flexibly shift between cognitive strategies. In our laboratory analog, participants performed two simultaneous search tasks that required discrimination and identification in central and peripheral areas of the visual field [24]. When performing both searches simultaneously, a cost to attentional resources is incurred due to switching from one search strategy to the other. For the central search condition, participants were required to report whether five randomly rotated letters were identical. During the peripheral search condition, participants had to locate and identify a briefly presented target (either “¬” or “-”) that could appear anywhere and with any rotation in a circular locus in the periphery (peripheral search condition). In the dual-search condition, participants performed the peripheral and central searches and were randomly asked to respond to either the central or peripheral search queries. By presenting both search conditions separately as well as within a dual-task context, both baseline performance and task-switching costs could be evaluated. Accuracy and RTs were recorded.
In four separate task blocks, the participants completed three task conditions. These included one block for each single-search condition: peripheral search only, where participants were asked which symbol they saw (¬ or ), and central search only, where participants were asked if they saw identical or different letters. Additionally, two blocks of dual-search tasks were performed, where both central search and peripheral search question trials were evenly distributed in each block. Each block consisted of 48 trials. The order of the individual trials was pseudorandomized, and the order of the four task blocks was counterbalanced across participants. Each of the tasks was carried out as follows.
Peripheral Search Alone: Participants were required to fixate at the center of the screen and to locate and identify the peripheral stimulus while ignoring the stimuli at the center. Because the position of the peripheral stimulus was chosen pseudo-randomly, participants could not improve their performance by fixating on a location other than the center. They did, however, know that the target would appear somewhere on the circumference of an imaginary circle in the periphery. Participants responded as quickly as possible while minimizing errors, pressing “7” if they identified a “¬” or “9” if they identified a “-”, depending on the symbol presented.
Central Search Alone: Participants were required to fixate at the center of the screen and to identify whether the group of five letters that appeared were all identical or different while ignoring the peripheral stimulus. The participants pressed the “7” key if the five central letters were identical, and the “9” if any letter was different.
Dual-Search (Mixed): Both search tasks were performed simultaneously. After the stimulus disappeared, a cue was presented that instructed participants to produce a response for either the central search or the peripheral search (equal probabilities across trials). Participants were expected to maintain performance in both conditions without prioritizing one task condition over another.
2.2.3. Stroop Task
The Stroop paradigm examines an individual’s ability to shift their cognitive set in the presence of distraction and to suppress irrelevant information and maintain focus on a given task as a measure of cognitive inhibition and control [25]. Participants were asked to look at words (i.e., red, blue, yellow, green) in two different conditions: congruent, where the word was displayed in the color font that its name denoted (e.g., “yellow” displayed in yellow font), and incongruent, where the word was displayed in a different color font than its name (e.g., “yellow” displayed in red font). The task consisted of four blocks—2 congruent (15 trials per block) and 2 incongruent (15 trials per block)—with presentation pseudorandomized across participants to prevent an order effect. Stimuli presentation trials were two seconds in duration with one-second interstimulus rest intervals and thirty-second rest intervals between blocks. Accuracy and reaction times were recorded.
2.3. fNIRS Neuroimaging
2.3.1. Data Collection
Hemodynamic response in the prefrontal cortex was measured using a continuous wave fNIRS system (fNIR Imager Model 1200; fNIR Devices, LLC., Potomac, MD, USA) first described by [26] and developed at Drexel University [27,28]. Light intensity at two near-infrared wavelengths of 730 and 850 nm and one additional ambient light channel from 16 optodes each were recorded at 2 Hz using COBI studio software (version 1.5.0.55) [28]. The fNIRS headband contains 4 light-emitting diodes (LEDs) and 10 photodetectors for a total of 16 optodes (measurement areas). The sensor was positioned based on the anatomical landmarks, as previously described [28], by aligning the center of the sensor to the midline of the forehead, with the bottom of the sensor touching the participants eyebrow so that the centroid point of the sensor was approximately at the prefrontal center (Fpz, where Fp refers to the frontopolar area of the prefrontal region, and Z as the central region), according to the 10–20 international coordinate system. Approximate anatomical mapping between fNIRS headband optodes and cortical regions is described in [29]. Baseline light intensity measurements were taken immediately preceding the start of the experiment. Time synchronization markers were sent from the stimulus presentation computer to the fNIRS acquisition computer for the registration of task responses, blocks, and trials for the temporal alignment of recorded data for all subjects.
2.3.2. Preprocessing
Raw light intensity data were pre-processed using a low-pass finite impulse response (FIR) filter, with a filter order of 20 and a cut-off frequency of 0.1 Hz [30]. The fluctuations from motion artifacts in the filtered raw signal and hemoglobin concentration changes were inspected both visually and also by using the automated sliding motion artifact rejection (SMAR) algorithm [31], which uses a coefficient-of-variation-based statistical filtering to assess signal quality and reject problematic channels with poor contact or saturated raw light intensity. Time-synchronized blocks of fNIRS pre-processed light intensity signals for all task periods were processed using the modified Beer–Lambert law [32] and baseline-corrected to calculate oxygenated hemoglobin (HbO) changes.
2.3.3. Analysis
Linear mixed-effects models (LMMs) were used for statistical analysis to account for intersubject variability and repeated measures [33]. Separate models were constructed to identify the impact of the fixed effects of interest on dependent measures of task response variables and respective biomarkers. LMMs incorporate random effects for individual participants while capturing variability between groups of participants and are flexible in handling missing data [34]. They do not require the same number of measurements across participants, which applies to fNIRS studies in which data may be missing due to technical issues or participant non-compliance. Finally, LMMs have been frequently applied in fNIRS studies, including those assessing cognitive processing [35,36].
For each task, model functionality was specified as follows: . Where ‘Outcome’ represents the dependent variable (e.g., response time, accuracy, HbO, neural efficiency), ‘Group’ represents the experience level (novice vs. experienced), and ‘Condition’ represents the task-specific conditions. Random intercepts were included to account for individual differences.
Here, the statistical significance in model fixed effects was assessed using the Satterthwaite approximation for degrees of freedom. Post hoc tests were conducted using Tukey contrasts adjusted for family-wise error rates using the Bonferroni correction. A criterion of α = 0.05 was designated as a threshold for statistical significance. For fNIRS analysis, false discovery rate (FDR) correction was applied to LMM results to correct for multiple family-wise testing across the entire list of optodes (with q = 0.05) [37]. Bonferroni correction was applied during the post hoc analysis of individual contrast comparisons within an optode that survived the previous FDR correction [38].
The process of skill acquisition and training can be viewed as the acquisition of knowledge and the formulation of strategies that attempt to reduce relative cognitive demands and increase overall mental efficiency, thereby allowing for better performance. The demands of metabolically expensive neuronal activity involved in the maintenance of active cognitive resources imply that changes in mental efficiency are coupled with changes in neural efficiency (NE) [39]. In this study, neural efficiency analysis was conducted as a way of providing a consolidated metric of behavioral performance which relates mental effort to outcome [39,40], and it was calculated using the following formula: , where z represents standardized scores. Here, the outcome was described by variables such as response time, accuracy, number of correct responses, and attentional flexibility while mental effort was assessed based on fNIRS-measured HbO. Effort and outcome values were converted into Z-scored measurements and then efficiency was computed using the distance of the point from the zero-efficiency line (i.e., where unit performance score is equal to the effort score) and assessed as dependent measurements using statistical tests [39]; this approach was utilized in diverse neuroergonomics studies by da Silva Soares, Ramirez-Chavez [36,41,42,43].
DSST models for behavioral performance utilized the independent variables of participant mean response time, and the number of correct responses per block were compared for each segment of the task (Task Block) as well as for novice and experienced skill level (Group). Additionally, independent variables for HbO and reaction time-based neural efficiency (NeurEffRxTime) at every optode location (optodes 1–16) of the prefrontal cortex were compared for each segment of the task (Task Block) as well as for novice and experienced skill levels (Group). All fixed terms were held as both between and within fixed factors for comparisons.
For the single-search component of the DVST, independent variables of participant mean response time and accuracy were compared between the central search condition and peripheral search condition of the task (Condition) as well as for novice and experienced skill level (Group). For neural-based measures, independent variables for HbO and reaction time-based neural efficiency (NeurEffRxTime) at every optode location (optode 1–16) of the prefrontal cortex were compared between Conditions as well as for Group.
In terms of dual-search condition DVST measures, independent variables of participant mean response time, accuracy, and attentional delay (mean response times for peripheral search trials subtracted from mean response times for central search trials per task block) were compared between novice and experienced skill level (Group). Independent variables for HbO, reaction time-based neural efficiency (NeurEffRxTime), and attentional flexibility-based neural efficiency (NeurEffFlexibilty) were compared at every optode location (1–16) of the prefrontal cortex for Group.
The Stroop task’s independent variables of participant mean response time and accuracy were compared between the congruent and incongruent conditions of the task (Condition) as well as for novice and experienced skill level (Group). Independent variables for HbO and reaction time-based neural efficiency (NeurEffRxTime) at every optode location (optodes 1–16) of the prefrontal cortex were compared between the congruent and incongruent conditions of the task as well as for novice and experienced skill level (Group).
3. Results
3.1. DSST Results
3.1.1. Response Time
A main effect was observed for Task Block, where the mean response times for all participants became significantly faster (F2,347 = 4.03, p < 0.05) as the task progressed (Block 1 = 0.9196 s, SEM = 0.0202; Block 2 = 0.8649 s, SEM = 0.0203; Block 3 = 0.8404 s, SEM = 0.0203). Additionally, there was a main effect present for Group (F1,347 = 24.73, p < 0.001), with experienced players demonstrating an overall faster mean response time (0.8168 s, SEM = 0.0177) compared to novice players (0.9331 s, SEM 0.0152).
3.1.2. Number of Correct Responses
Over the duration of the DSST, there was a significant increase in participant scores (number of correct responses) with each repetition of the task block (F2,347 = 34.45, p < 0.05), with task scores improving from the first block to the last (Block 1 = 58.7103, SEM = 1.0069; Block 2 = 61.7575, SEM = 1.0132; Block 3 = 62.8110, SEM = 0.0132). Furthermore, there was a significant main effect for Group (F1,347 = 25.49, p < 0.001), with experienced players showing heightened response efficiency (64.04 correct, SEM = 0.89) over novices (58.15 correct, SEM = 0.76).
3.1.3. fNIRS
While no main effects for Group were present at any optode, significant main effects for Task Block were identified for HbO signals at optode 5 (F2,322 = 6.33, p < 0.01), optode 7 (F2,316 = 5.57, p < 0.005), optode 8 (F2,304 = 4.18, p < 0.05), optode 9 (F2,313 = 7.00, p = 0.001), optode 10 (F2,311 = 4.12, p < 0.05), and optode 11 (F2,300 = 3.55, p < 0.05). For all optodes, hemodynamic activity increased monotonically from the start of the task to the end of the task.
3.1.4. Neural Efficiency
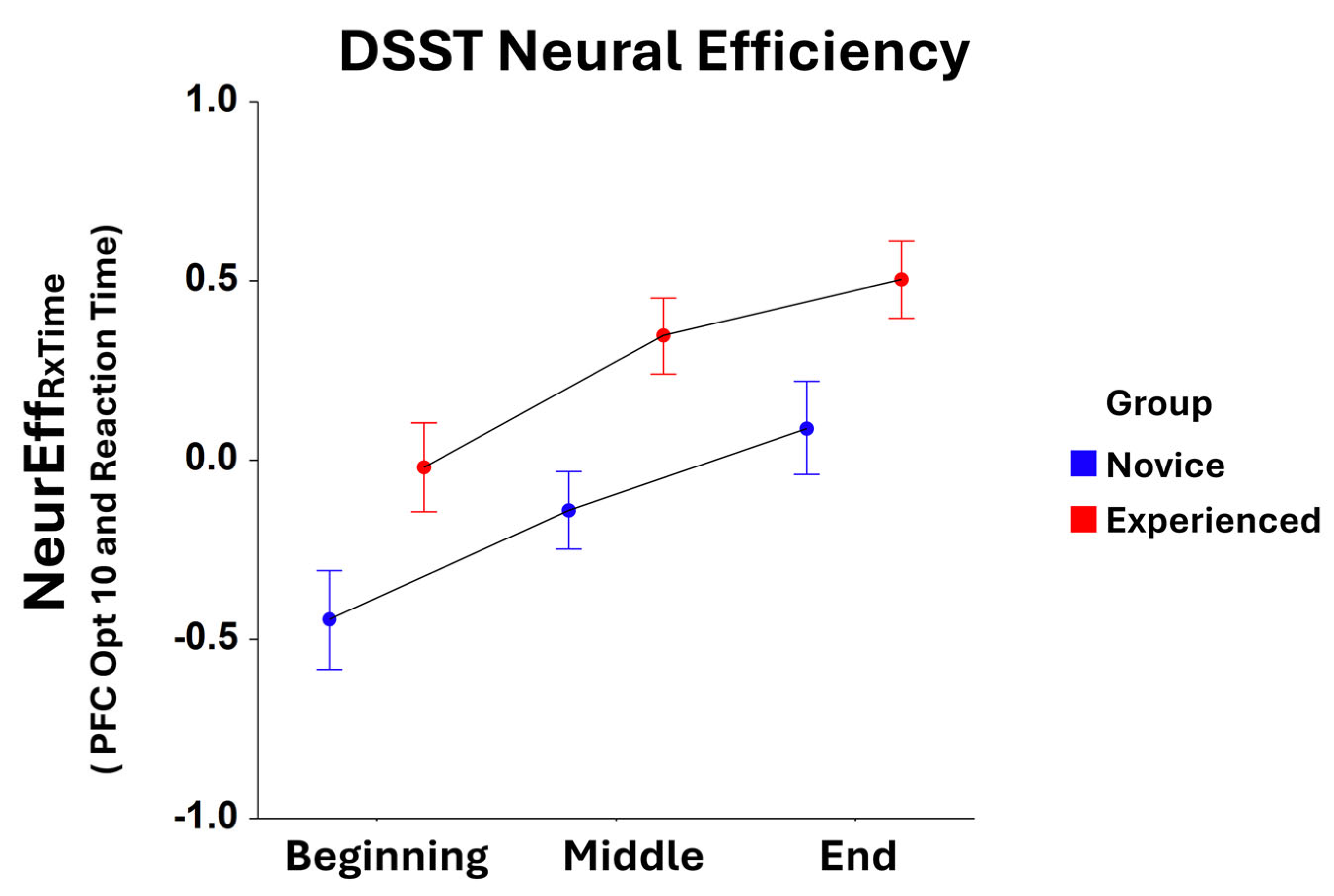
Significant main effects for the Task Block were found for NeurEffRxTime at optode 1 (F2,305 = 3.65, p < 0.05), optode 3 (F2,321 = 7.40, p < 0.001), optode 4 (F2,309 = 6.42, p < 0.005), optode 5 (F2,322 = 11.58, p < 0.0001), optode 6 (F2,308 = 7.35, p < 0.001), optode 7 (F2,316 = 11.07, p < 0.001), optode 8 (F2,304 = 10.53, p < 0.001), optode 9 (F2,313 = 13.22, p < 0.001), optode 10 (F2,311 = 9.27, p < 0.001), optode 11 (F2,300 = 10.27, p < 0.001), optode 12 (F2,304 = 7.85, p < 0.001), optode 13 (F2,309 = 4.78, p < 0.01), optode 14 (F2,310 = 3.89, p < 0.05), and optode 15 (F2,282 = 4.03, p < 0.05). For all optodes, there were increases in NeurEffRxTime from the initial task block to the final task block. Moreover, main effects for Group were observed for NeurEffRxTime at optode 1 (F1,305 = 7.74, p < 0.01), optode 2 (F1,300 = 13.52, p < 0.001), optode 3 (F1,321 = 7.24, p < 0.01), optode 4 (F1,309 = 15.24, p < 0.001), optode 5 (F1,322 = 5.03, p < 0.05), optode 6 (F1,308 = 16.31, p < 0.001), optode 7 (F1,316 = 5.28, p < 0.05), optode 8 (F1,304 = 9.81, p < 0.005), optode 9 (F1,313 = 8.39, p < 0.005), optode 10 (F1,311 = 19.07, p < 0.001), optode 12 (F1,304 = 13.23, p < 0.001), and optode 14 (F1,310 = 6.24, p < 0.05). Here, neural efficiency was higher for experienced players compared to novices at every optode location, with optode 10 being most significant (Figure 2).
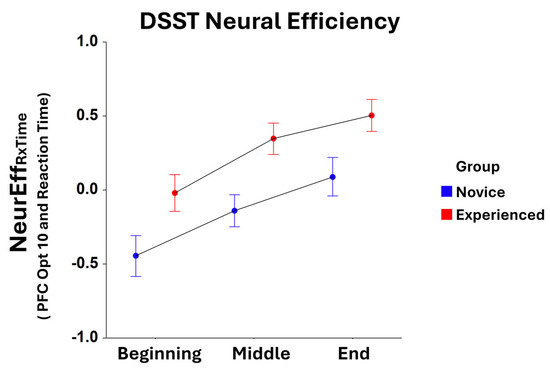
Figure 2.
Comparison of reaction time-based NeurEffRxTime at optode 10 between novice and experienced FPS players for each block of the DSST. There is a significant main effect (F1,311 = 19.07, p < 0.001) in the novice and experienced Group as well as a significant main effect (F2,311 = 9.27, p < 0.001) for Task Block. Whiskers are the standard error of the mean (SEM).
3.2. Single-Search DVST Results
3.2.1. Response Time
Response times for all participants were first compared for the single-search conditions (central search condition and peripheral search condition) of the DVST. Here, there was a significant main effect for Condition as mean central search response times (0.8525 s, SEM = 0.0235) were significantly faster (F1,234 = 11.14, p < 0.001) than the mean peripheral search response times (0.9634 s, SEM = 0.0235). No main effect from Group was observed.
3.2.2. Accuracy
Participants demonstrated significantly higher accuracy (F1,234 = 195.92, p < 0.001) in responses during the peripheral search condition (0.8792, SEM < 0.01) as compared to the central search condition (0.6884, SEM = 0.0096) of the DVST, with no main effect for Group present.
3.2.3. fNIRS
Changes in HbO for participants during the central search condition and the peripheral search condition were compared. While no main effects for Group were present at any optode, significant main effects for Condition were identified for HbO signals at optode 6 (F1,212 = 4.58, p < 0.05), 14 (F1,203 = 6.04, p < 0.05) and 16 (F1,186 = 4.48, p < 0.05). For all optodes, the central search condition resulted in higher activity compared to the peripheral search condition. Next, significant Group main effects were observed for HbO signals in the dual-search condition alone at optode 2 (F1,202 = 5.44, p < 0.05), 3 (F1,214 = 5.82, p < 0.05), 4 (F1,210 = 6.54, p < 0.05), 5 (F1,219 = 4.28, p < 0.05), 6 (F1,209 = 5.76, p < 0.05), 7 (F1,209 = 4.60, p < 0.05), 8 (F1,199 = 5.03, p < 0.05), and 9 (F1,211 = 6.94, p < 0.01). For all optode locations, experienced players displayed higher HbO activity compared to novices in the dual-search condition.
3.2.4. Neural Efficiency
There were no significant main effects found for NeurEffRxTime for Condition or Group factors.
3.3. Dual-Search DVST Results
3.3.1. Response Time
When comparing mean response times within the dual-search condition of the task alone, no main effect for Group was observed.
3.3.2. Accuracy
When accuracy measures are compared for participants within the dual-search condition of the task alone, there is no main effect for Group.
3.3.3. Attentional Delay
Within the dual-search condition of the DVST, the mean response times for peripheral search trials were subtracted from the mean response times for central search trials to obtain a metric for attentional delay between cognitive strategies for each participant. Here, there was a significant difference between Groups (F1,117 = 4.36, p < 0.05), with novices displaying increased delay (0.1093 s, SEM = 0.0501) compared to experienced players (0.0484 s, SEM = 0.0220).
3.3.4. fNIRS
Significant Group main effects were observed for HbO signals in the dual-search condition alone at optode 2 (F1,202 = 5.44, p < 0.05), 3 (F1,214 = 5.82, p < 0.05), 4 (F1,210 = 6.54, p < 0.05), 5 (F1,219 = 4.28, p < 0.05), 6 (F1,209 = 5.76, p < 0.05), 7 (F1,209 = 4.60, p < 0.05), 8 (F1,199 = 5.03, p < 0.05), and 9 (F1,211 = 6.94, p < 0.01). For all optode locations, experienced players displayed higher HbO activity compared to novices.
3.3.5. Neural Efficiency
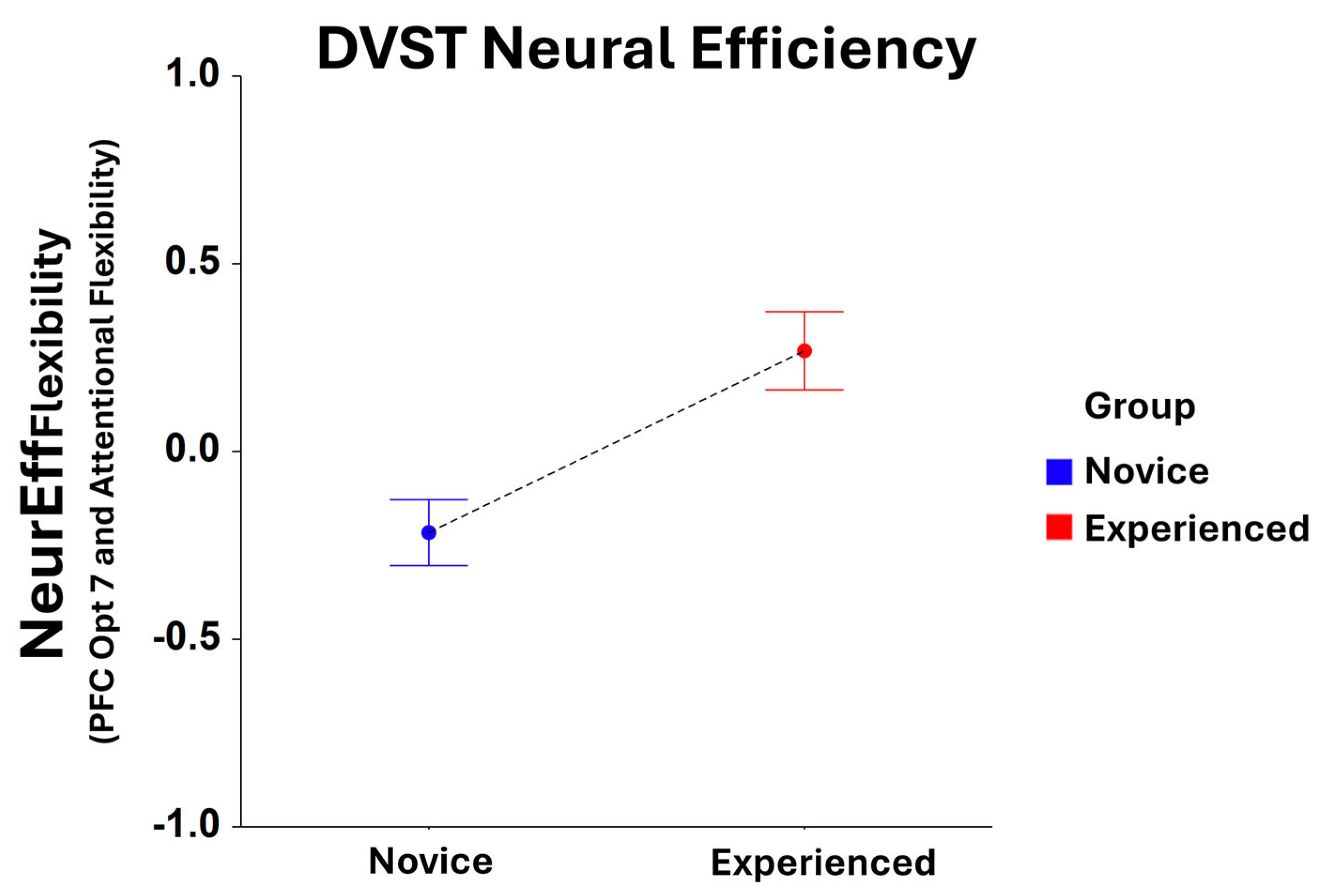
While no significant main effects were found for NeurEffRxTime for Group in the dual-search condition, significant Group main effects were identified for NeurEffFlexibilty at optode 1 (F1,208 = 6.39, p < 0.05), 2 (F1,202 = 8.43, p < 0.005), 3 (F1,214 = 10.91, p = 0.001), 4 (F1,210 = 10.94, p = 0.01), 5 (F1,219 = 9.22, p < 0.005), 6 (F1,209 = 10.14, p < 0.005), 7 (F1,209 = 12.34, p < 0.001), 8 (F1,199 = 9.15, p < 0.005), 9 (F1,211 = 11.90, p < 0.001), 10 (F1,204 = 6.78, p < 0.01), 12 (F1,200 = 4.92, p < 0.05), and 16 (F1,184 = 4.01, p < 0.05). Here, NeurEffFlexibilty was optimized for experienced players compared to novices at every optode location, with optode 7 being the most significant location (Figure 3).
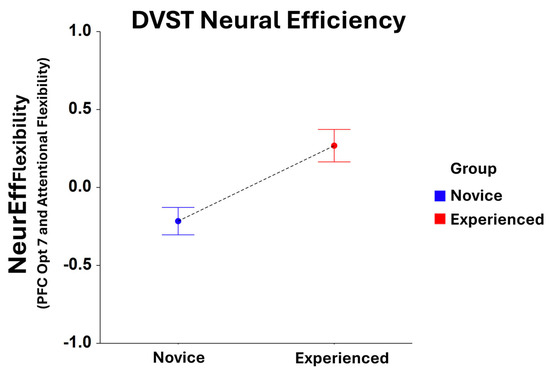
Figure 3.
Comparison of NeurEffFlexibility at optode 7 for novice and experienced FPS players performing the dual-search condition of the DVST. There is a significant main effect (F1,209 = 12.34, p = 0.001) in the novice and experienced Groups. Whiskers are SEM.
3.4. Stroop Task
3.4.1. Response Time
A main effect was observed for Condition, where the mean response times for all participants were significantly faster (F1,468 = 37.54, p < 0.001) in the congruent condition of the task (0.8372 s, SEM = 0.01) compared to the incongruent condition of the task (0.9274 s, SEM = 0.01). Additionally, there is a main effect present for Group (F1,468 = 6.51, p = 0.01), with experienced players demonstrating overall faster response times (0.8635 s, SEM = 0.0111) compared to novice players (0.9011 s, SEM = 0.0097). However, post hoc comparison tests show that these significant Group effects are present due to response times in the congruent condition for each respective Group as both experienced players (F1,468 = 18.93, p < 0.001) and novices (F1,468 = 18.78, p < 0.001) logged faster reaction times in the congruent condition compared to the incongruent condition within their respective groups with experienced players maintaining faster responses than novices in the congruent condition.
3.4.2. Accuracy
There is a main effect for present for Condition (F1,468 = 16.45, p < 0.001) where a significant difference in correct response accuracy is observed (F1,468 = 16.45, p < 0.001) between participant mean correct response rate in the congruent condition of the task (0.9845, SEM = 0.0038) and the incongruent condition of the task (0.9628, SEM < 0.005). No main effect is observed for Group in the model.
3.4.3. fNIRS
Changes in HbO for participants during the congruent condition and the incongruent condition of the Stroop Task were compared. Significant main effects for Condition were identified for HbO signals at optodes 1 (F1,401 = 4.19, p < 0.05), 3 (F1,396 = 13.80, p < 0.001), 4 (F1,392 = 9.80, p < 0.005), 6 (F1,375 = 4.15, p < 0.05), and 13 (F1,387 = 5.62, p < 0.05). For all optodes, the incongruent condition resulted in higher activity compared to the congruent condition. Next, significant Group main effects were observed for HbO signals at optode 7 (F1,398 = 4.67, p < 0.05), 8 (F1,368 = 3.88, p < 0.05), and 10 (F1,366 = 9.44, p < 0.005). Here, experienced players displayed higher activity compared to novices at these optodes. More importantly, the interaction effect of Group * Condition was found to be significant at optode 10 (F1,366 = 4.25, p < 0.005) and 14 (F1,380 = 6.04, p = 0.01). Post hoc comparison tests for optode 10 with Bonferroni correction showed that within the incongruent condition of the task, experienced players demonstrated higher HbO activity than novices (F1,366 = 13.24, p < 0.001).
3.4.4. Neural Efficiency
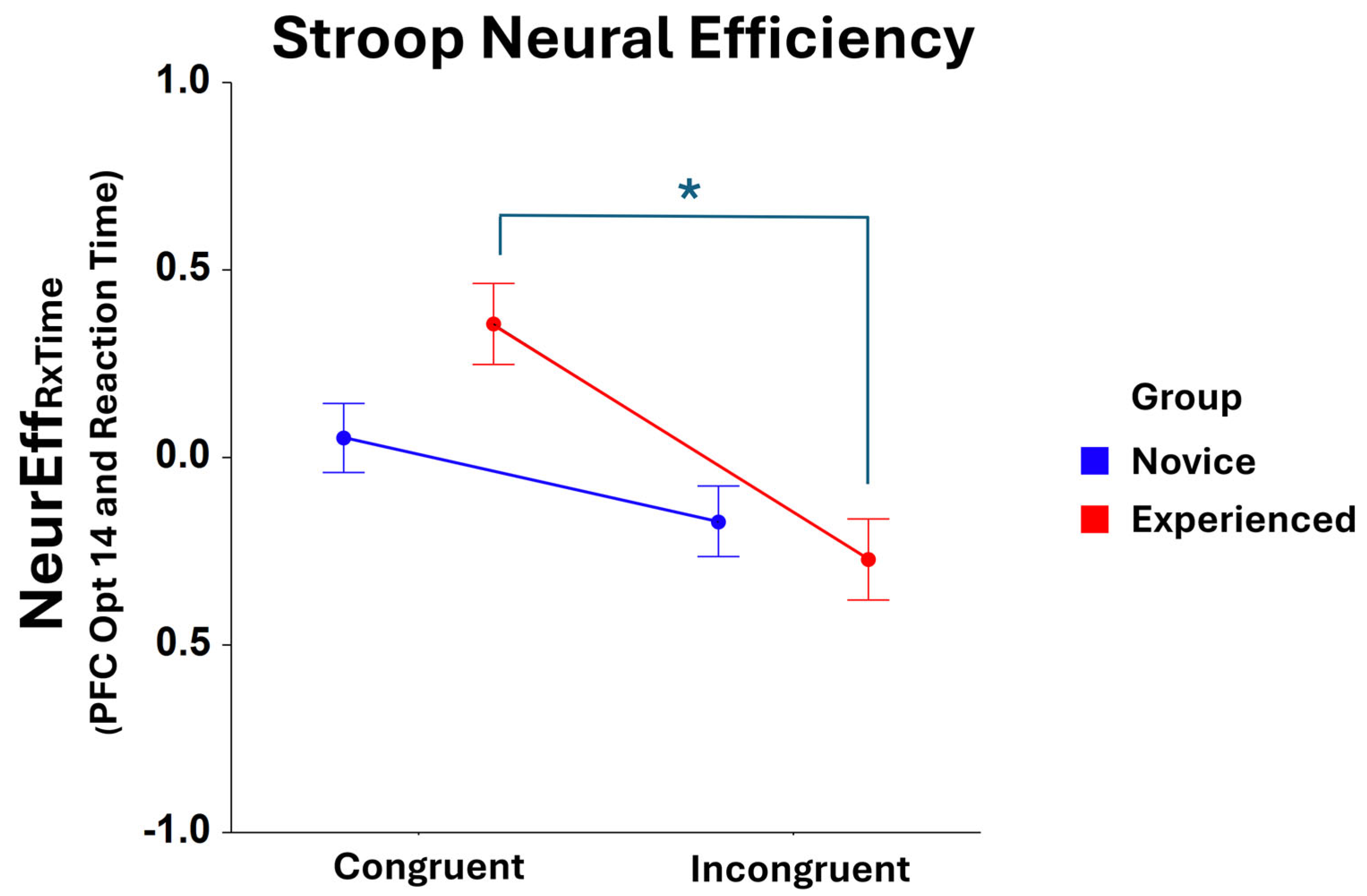
Significant Condition main effects were found for NeurEffRxTime at optode 1 (F1,401 = 25.64, p < 0.001), optode 2 (F1,374 = 22.34, p < 0.001), optode 3 (F1,396 = 37.18, p < 0.001), optode 4 (F1,392 = 35.37, p < 0.001), optode 5 (F1,405 = 21.23, p < 0.001), optode 6 (F1,375 = 25.50, p < 0.001), optode 7 (F1,398 = 19.87, p < 0.001), optode 8 (F1,368 = 17.55, p < 0.001), optode 9 (F1,395 = 16.58, p < 0.001), optode 10 (F1,366 = 14.29, p < 0.001), optode 11 (F1,376 = 20.28, p < 0.001), optode 12 (F1,371 = 20.75, p < 0.001), optode 13 (F1,387 = 24.00, p < 0.001), optode 14 (F1,380 = 17.78, p < 0.001), optode 15 (F1,352 = 18.52, p < 0.001), and optode 16 (F1,336 = 21.18, p < 0.001). For all optodes, NeurEffRxTime was higher in the congruent condition compared to the incongruent condition of the Stroop Task. An interaction effect of Group * Condition was also found to be significant at optode 14 (F1,380 = 4.06, p < 0.05), with post hoc comparison tests showing that experienced players exhibit compromised neural efficiency in the incongruent condition compared to the congruent condition of the task (F1,380 = 17.51, p < 0.001), as shown in Figure 4. No other optode produced a significant NeurEffRxTime result for Group * Condition.
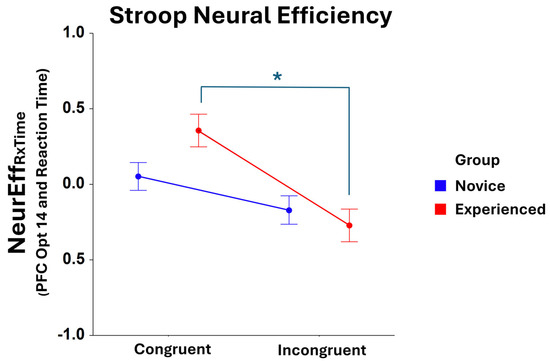
Figure 4.
Comparison of NeurEffRxTime at optode 14 for novice and experienced FPS players in the Stroop task. There is a significant main effect (F1,209 = 12.34, p < 0.001) between the congruent and incongruent conditions of the task as well as * a significant interaction for novice and experienced Group and Condition (F1,380 = 4.06, p < 0.05). A significant post hoc contrast is observed for the experienced group: congruent vs. incongruent (F1,380 = 17.51, p < 0.001). Whiskers are SEM.
4. Discussion
The present study aimed to explicate how prolonged FPS gaming experience influences the behavioral performance and neural efficiency of cognitive control (CC) processes. By employing three established cognitive tasks, DSST for updating, DVST for attentional flexibility or shifting, and the Stroop Task for inhibition, we examined how novice and experienced FPS gamers differ in behavioral outcomes and PFC hemodynamic activity, as measured by fNIRS wearable neuroimaging.
4.1. Updating and the DSST
Our DSST findings demonstrate that experienced FPS players responded both faster and more accurately than their novice counterparts. These data echo prior observations [23], indicating that well-practiced cognitive routines—such as rapid symbol–digit matching—may be reinforced by the fast-paced demands of FPS gameplay. Notably, while both groups exhibited a significant task-load effect in prefrontal oxyhemoglobin (HbO) levels over the duration of the DSST, no clear between-group difference in HbO emerged. This suggests that although both novices and experienced gamers increasingly recruited PFC resources as the task demands accumulated, the relative volume of PFC activation remained comparable across groups.
However, NeurEffRxTime metrics reveal that experienced players employed these prefrontal resources more efficiently for each correct response. In other words, despite similar overall levels of HbO, experienced players achieved higher accuracy and faster responses, indicating that their FPS-based training may sharpen the ability to allocate cognitive resources selectively. These results add nuance to previous reports showing that trained individuals can achieve performance advantages without necessarily demonstrating a greater magnitude of PFC activation [3]. Instead, the critical distinction often lies in how these neural resources are utilized, a hallmark of “neural efficiency” in expert populations [4].
4.2. Attentional Flexibility and the DVST
The DVST was designed to parse out how gaming experience might influence shifting and search strategies under more complex conditions. While no significant group differences were found in single or dual-search accuracy, we did observe that experienced gamers switched more fluidly between central and peripheral targets during the dual-search component. This faster switching aligns with a body of work suggesting that intensive FPS gameplay trains players to handle multiple attention-demanding stimuli more rapidly [5,6].
From a neurophysiological standpoint, our fNIRS results support these behavioral observations and align with existing research [7], indicating that heightened CC performance under escalating cognitive demands is accompanied by increased PFC engagement. Critically, we observed not only an upregulation of HbO in the experienced group but also improved NeurEffFlexibility. This suggests that when faced with higher complexity, experienced gamers may recruit PFC networks both more robustly and more effectively to support rapid attentional shifting and the integration of competing visual information. These findings bolster the notion that repeated exposure to FPS gameplay refines the neural circuitry underlying flexible visual search and the timely updating of mental sets.
4.3. Inhibition and the Stroop Task
Consistent with expectations, participants’ performance declined in the incongruent Stroop condition relative to the congruent one, underscoring the classic interference effect associated with response inhibition demands [8]. Contrary to what might be seen in domains where strong inhibitory control is explicitly trained (e.g., certain sports or specialized tasks requiring high impulse control, like golf, archery, surgery, and piloting), our results revealed no significant behavioral advantage for experienced FPS gamers on incongruent trials. This aligns with earlier propositions that FPS environments may not actively encourage the same inhibitory skill development as they do for other CC subdomains like updating or shifting [9,10]. For instance, most causal first-person shooter games often lack penalties for “friendly fire”.
Interestingly, however, the experienced group did exhibit faster reaction times in the congruent Stroop condition. This faster baseline performance likely reflects the generalized quick-reflex advantage frequently reported among experienced FPS players [11]. From a neural standpoint, experienced players manifested significantly elevated medial prefrontal cortex (mPFC; optodes 7, 8, and 10) activity during incongruent trials compared to novices, suggesting a greater recruitment of executive control mechanisms to achieve comparable behavioral outcomes. While experienced gamers displayed superior or equal performance on incongruent trials in absolute terms, NeurEffRxTime data revealed a significant interaction effect in the right lateral PFC (middle frontal gyrus) (optode 14). Specifically, experienced gamers showed substantially higher neural efficiency during congruent trials, compared to incongruent trials, showing a more pronounced Stroop effect at a neural-efficiency level. This significant contrast in efficiency between conditions was not observed to the same degree in novices. However, the neural efficiency of novices and experienced participants for incongruent trials were not significantly different. Thus, gaming experience did not appear to confer advantages for inhibition-depending trials (incongruent condition). Experienced players increased neural recruitment during inhibition-demanding trials, while achieving performance comparable to novices in the incongruent condition despite significant advantages during the congruent condition. This pattern suggests differential optimization for inhibitory control processes in experienced FPS players, potentially reflecting an adaptation to gaming environments where rapid responses may be rewarded over inhibitory control.
4.4. Implications
Taken together, these findings illustrate that prolonged FPS gameplay may bolster certain CC domains—particularly those related to rapid updating and attentional shifting—while providing less support for response inhibition. Although experienced gamers were able to mobilize the PFC more effectively in the DSST and DVST, they demonstrated a pattern of increased neural recruitment for the incongruent Stroop condition, but without a strong corresponding behavioral benefit. This pattern resonates with theories of cognitive plasticity that propose skill-specific adaptations in experts: repeated practice in certain tasks can yield considerable performance gains for those task-relevant processes, but such gains do not necessarily transfer to tasks that require a fundamentally different cognitive approach [12].
4.5. Limitations and Future Directions
Several avenues for future work as well as protocol limitations are worth highlighting. First, the current study did not differentiate between various FPS game types (e.g., tactical vs. run-and-gun) that may impose diverse inhibitory or attentional demands or attempt to compare the influence of other video game genres (e.g., real-time strategy, racing, simulators, puzzle games). Future research should include multi-genre gaming comparisons to isolate FPS-specific effects from the general effects of gaming experience.
Second, although video game experience is increasingly common amongst the population at large, the study population was primarily young-adult and experienced FPS gamers were predominantly male. In addition, binary classification of experienced gamers may overlook dose-dependent relationships between gaming and cognition, potentially revealing more nuanced relationships between practice intensity and cognitive control development. Other potential confounders may include participant fatigue and time-of-day variability in testing, as well as other individual differences which may affect both behavioral performance and neural measurements.
Third, while fNIRS offers high ecological validity and moderate spatial resolution in measuring cortical oxygenation, combining it with complementary methods (e.g., electroencephalography, EEG, for temporal resolution or functional magnetic resonance imaging, fMRI, for deeper subcortical insights) would provide a more comprehensive mapping of neural processes.
Lastly, although we noted that the experienced group performed better in tasks emphasizing updating and shifting, a longitudinal study design would be ideal to confirm a direct causal link between FPS training and changes in CC and PFC activation patterns. However, even with longitudinal examination, continued gaming practice which helps develop eventual expertise may also involve a form of performance-based self-selection in which individuals who show early advantages in a particular activity are also more likely to continue practice, dramatizing “natural” differences between experience and novices [44].
5. Conclusions
Overall, our findings make a strong case that the intense cognitive demands of FPS games foster improvements in updating and shifting processes, reflected by both faster behavioral performance and more efficient PFC resource allocation. In contrast, the inhibitory control domain appears less influenced—or even detrimentally shaped—by prolonged FPS experience, as evidenced by the increased mPFC activation required for incongruent Stroop trials in experienced gamers. These discoveries not only refine our understanding of how specific dimensions of CC respond to domain-specific training but also open new perspectives on leveraging fNIRS in skill acquisition research. By mapping the neural signatures of CC development in real-world contexts, this work contributes to a broader neuroergonomic framework that can be extended to other high-performance domains requiring rapid decision making and attentional agility.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/brainsci15060568/s1, Table S1: Demographic characteristics and gaming experience of participants (N = 120); Table S2: Cognitive performance measures comparing novice and experienced participants.
Author Contributions
Conceptualization, H.A., J.W., and R.S.; Methodology, H.A., R.S., J.W., and A.C.; Software, A.C. and J.W.; Resources, R.S. and H.A.; Data Collection, A.C., J.W., and Y.T.; Formal Analysis, J.W., A.C., and H.A.; Writing—Original Draft Preparation, J.W.; Writing—Review and Editing, H.A., A.C., Y.T., and R.S.; Visualization, J.W. and H.A.; Supervision, H.A.; Funding Acquisition, H.A. and R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by Drexel Solutions Institute.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of Drexel University (protocol number 2102008356, 27 May 2021).
Informed Consent Statement
Informed consent, which was approved by Drexel University Institutional Review Board, was obtained from all subjects involved in the study.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material; further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to thank the Drexel Solutions Institute staff for their help with study.
Conflicts of Interest
fNIR Devices, LLC manufactures the optical brain imaging instrument and licensed IP and know-how from Drexel University. HA was involved in the technology development and thus offered a minor share in the startup firm fNIR Devices, LLC. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Miller, E.K.; Cohen, J.D. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.P.; Robbins, T.W. The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacology 2022, 47, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Fedorenko, E.; Duncan, J.; Kanwisher, N. Broad domain generality in focal regions of frontal and parietal cortex. Proc. Natl. Acad. Sci. USA 2013, 110, 16616–16621. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cogn. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef]
- Carter, C.S.; Braver, T.S.; Barch, D.M.; Botvinick, M.M.; Noll, D.; Cohen, J.D. Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 1998, 280, 747–749. [Google Scholar] [CrossRef]
- Kerns, J.G.; Cohen, J.D.; MacDonald, A.W., III; Cho, R.Y.; Stenger, V.A.; Carter, C.S. Anterior cingulate conflict monitoring and adjustments in control. Science 2004, 303, 1023–1026. [Google Scholar] [CrossRef]
- Wu, S.; Spence, I. Playing shooter and driving videogames improves top-down guidance in visual search. Atten. Percept. Psychophys. 2013, 75, 673–686. [Google Scholar] [CrossRef]
- Derrfuss, J.; Brass, M.; Von Cramon, D.Y. Cognitive control in the posterior frontolateral cortex: Evidence from common activations in task coordination, interference control, and working memory. Neuroimage 2004, 23, 604–612. [Google Scholar] [CrossRef]
- Cole, M.W.; Yarkoni, T.; Repovš, G.; Anticevic, A.; Braver, T.S. Global connectivity of prefrontal cortex predicts cognitive control and intelligence. J. Neurosci. 2012, 32, 8988–8999. [Google Scholar] [CrossRef]
- Schad, D.J.; Jünger, E.; Sebold, M.; Garbusow, M.; Bernhardt, N.; Javadi, A.-H.; Zimmermann, U.S.; Smolka, M.N.; Heinz, A.; Rapp, M.A. Processing speed enhances model-based over model-free reinforcement learning in the presence of high working memory functioning. Front. Psychol. 2014, 5, 1450. [Google Scholar] [CrossRef]
- Colzato, L.S.; van den Wildenberg, W.P.; Zmigrod, S.; Hommel, B. Action video gaming and cognitive control: Playing first person shooter games is associated with improvement in working memory but not action inhibition. Psychol. Res. 2013, 77, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Colzato, L.S.; Van Leeuwen, P.J.; Van Den Wildenberg, W.; Hommel, B. DOOM’d to switch: Superior cognitive flexibility in players of first person shooter games. Front. Psychol. 2010, 1, 1515. [Google Scholar] [CrossRef]
- Steenbergen, L.; Sellaro, R.; Stock, A.-K.; Beste, C.; Colzato, L.S. Action video gaming and cognitive control: Playing first person shooter games is associated with improved action cascading but not inhibition. PLoS ONE 2015, 10, e0144364. [Google Scholar] [CrossRef]
- Deleuze, J.; Christiaens, M.; Nuyens, F.; Billieux, J. Shoot at first sight! First person shooter players display reduced reaction time and compromised inhibitory control in comparison to other video game players. Comput. Hum. Behav. 2017, 72, 570–576. [Google Scholar] [CrossRef]
- Izzetoglu, K.; Bunce, S.; Onaral, B.; Pourrezaei, K.; Chance, B. Functional optical brain imaging using near-infrared during cognitive tasks. Int. J. Hum. -Comput. Interact. 2004, 17, 211–227. [Google Scholar] [CrossRef]
- Matsuda, G.; Hiraki, K. Prefrontal cortex deactivation during video game play. In Gaming, Simulations, and Society: Research Scope and Perspective; Springer: Tokyo, Japan, 2005; pp. 101–109. [Google Scholar]
- Basak, C.; Voss, M.W.; Erickson, K.I.; Boot, W.R.; Kramer, A.F. Regional differences in brain volume predict the acquisition of skill in a complex real-time strategy videogame. Brain Cogn. 2011, 76, 407–414. [Google Scholar] [CrossRef]
- Hyun, G.J.; Shin, Y.W.; Kim, B.-N.; Cheong, J.H.; Jin, S.N.; Han, D.H. Increased cortical thickness in professional on-line gamers. Psychiatry Investig. 2013, 10, 388. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Emory, E. Executive function and the frontal lobes: A meta-analytic review. Neuropsychol. Rev. 2006, 16, 17–42. [Google Scholar] [CrossRef]
- Smith, E.E.; Jonides, J. Storage and executive processes in the frontal lobes. Science 1999, 283, 1657–1661. [Google Scholar] [CrossRef]
- Alanko, D. The health effects of video games in children and adolescents. Pediatr. Rev. 2023, 44, 23–32. [Google Scholar] [CrossRef]
- Rypma, B.; Prabhakaran, V. When less is more and when more is more: The mediating roles of capacity and speed in brain-behavior efficiency. Intelligence 2009, 37, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Hubert-Wallander, B.; Green, C.S.; Sugarman, M.; Bavelier, D. Changes in search rate but not in the dynamics of exogenous attention in action videogame players. Atten. Percept. Psychophys. 2011, 73, 2399–2412. [Google Scholar] [CrossRef]
- VanRullen, R.; Reddy, L.; Koch, C. Visual search and dual tasks reveal two distinct attentional resources. J. Cogn. Neurosci. 2004, 16, 4–14. [Google Scholar] [CrossRef]
- Strauss, E.; Sherman, E.M.; Spreen, O. A Compendium of Neuropsychological Tests: Administration, Norms, and Commentary; American Chemical Society: Washington, DC, USA, 2006. [Google Scholar]
- Chance, B.; Anday, E.; Nioka, S.; Zhou, S.; Hong, L.; Worden, K.; Li, C.; Murray, T.; Ovetsky, Y.; Pidikiti, D. A novel method for fast imaging of brain function, non-invasively, with light. Opt. Express 1998, 2, 411–423. [Google Scholar] [CrossRef]
- Izzetoglu, M.; Izzetoglu, K.; Bunce, S.; Ayaz, H.; Devaraj, A.; Onaral, B.; Pourrezaei, K. Functional near-infrared neuroimaging. IEEE Trans. Neural Syst. Rehabil. Eng. 2005, 13, 153–159. [Google Scholar] [CrossRef]
- Ayaz, H.; Shewokis, P.A.; Curtin, A.; Izzetoglu, M.; Izzetoglu, K.; Onaral, B. Using MazeSuite and functional near infrared spectroscopy to study learning in spatial navigation. JoVE (J. Vis. Exp.) 2011, 56, e3443. [Google Scholar]
- Liu, Y.; Piazza, E.A.; Simony, E.; Shewokis, P.A.; Onaral, B.; Hasson, U.; Ayaz, H. Measuring speaker–listener neural coupling with functional near infrared spectroscopy. Sci. Rep. 2017, 7, 43293. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, H.; Izzetoglu, M.; Izzetoglu, K.; Onaral, B. The use of functional near-infrared spectroscopy in neuroergonomics. In Neuroergonomics; Elsevier: Amsterdam, The Netherlands, 2019; pp. 17–25. [Google Scholar]
- Ayaz, H.; Izzetoglu, M.; Shewokis, P.A.; Onaral, B. Sliding-window motion artifact rejection for functional near-infrared spectroscopy. In Proceedings of the 2010 Annual International Conference of the IEEE Engineering in Medicine and Biology, Buenos Aires, Argentina, 31 August–4 September 2010; IEEE: Piscataway, NJ, USA, 2010. [Google Scholar]
- Cope, M.; Delpy, D.; Reynolds, E.; Wray, S.; Wyatt, J.; Van der Zee, P. Methods of quantitating cerebral near infrared spectroscopy data. In Oxygen Transport to Tissue X; Springer: New York, NY, USA, 1988; pp. 183–189. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Krueger, C.; Tian, L. A comparison of the general linear mixed model and repeated measures ANOVA using a dataset with multiple missing data points. Biol. Res. Nurs. 2004, 6, 151–157. [Google Scholar] [CrossRef]
- Mark, J.A.; Curtin, A.; Kraft, A.E.; Ziegler, M.D.; Ayaz, H. Mental workload assessment by monitoring brain, heart, and eye with six biomedical modalities during six cognitive tasks. Front. Neuroergon. 2024, 5, 1345507. [Google Scholar] [CrossRef]
- da Silva Soares, R.; Ramirez-Chavez, K.L.; Tufanoglu, A.; Barreto, C.; Sato, J.R.; Ayaz, H. Cognitive Effort during Visuospatial Problem Solving in Physical Real World, on Computer Screen, and in Virtual Reality. Sensors 2024, 24, 977. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Dan, I. Exploring the false discovery rate in multichannel NIRS. Neuroimage 2006, 33, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Tak, S.; Ye, J.C. Statistical analysis of fNIRS data: A comprehensive review. Neuroimage 2014, 85, 72–91. [Google Scholar] [CrossRef]
- Curtin, A.; Ayaz, H. Neural efficiency metrics in neuroergonomics: Theory and applications. In Neuroergonomics; Elsevier: Amsterdam, The Netherlands, 2019; pp. 133–140. [Google Scholar]
- Tuovinen, J.E.; Paas, F. Exploring multidimensional approaches to the efficiency of instructional conditions. Instr. Sci. 2004, 32, 133–152. [Google Scholar] [CrossRef]
- Joshi, S.; Weedon, B.D.; Esser, P.; Liu, Y.-C.; Springett, D.N.; Meaney, A.; Inacio, M.; Delextrat, A.; Kemp, S.; Ward, T.; et al. Neuroergonomic assessment of developmental coordination disorder. Sci. Rep. 2022, 12, 10239. [Google Scholar] [CrossRef]
- Thomas, N.; Miller, A.J.; Ayaz, H.; Brown, J.D. Haptic shared control improves neural efficiency during myoelectric prosthesis use. Sci. Rep. 2023, 13, 484. [Google Scholar] [CrossRef]
- Curtin, A.; Ayaz, H.; Tang, Y.; Sun, J.; Wang, J.; Tong, S. Enhancing neural efficiency of cognitive processing speed via training and neurostimulation: An fNIRS and TMS study. NeuroImage 2019, 198, 73–82. [Google Scholar] [CrossRef]
- Bavelier, D.; Green, C.S. Enhancing attentional control: Lessons from action video games. Neuron 2019, 104, 147–163. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).