

Metabolic Dysfunction and Dietary Interventions in Migraine Management: The Role of Insulin Resistance and Neuroinflammation—A Narrative and Scoping Review


Abstract

1. Introduction
The Scope of This Paper
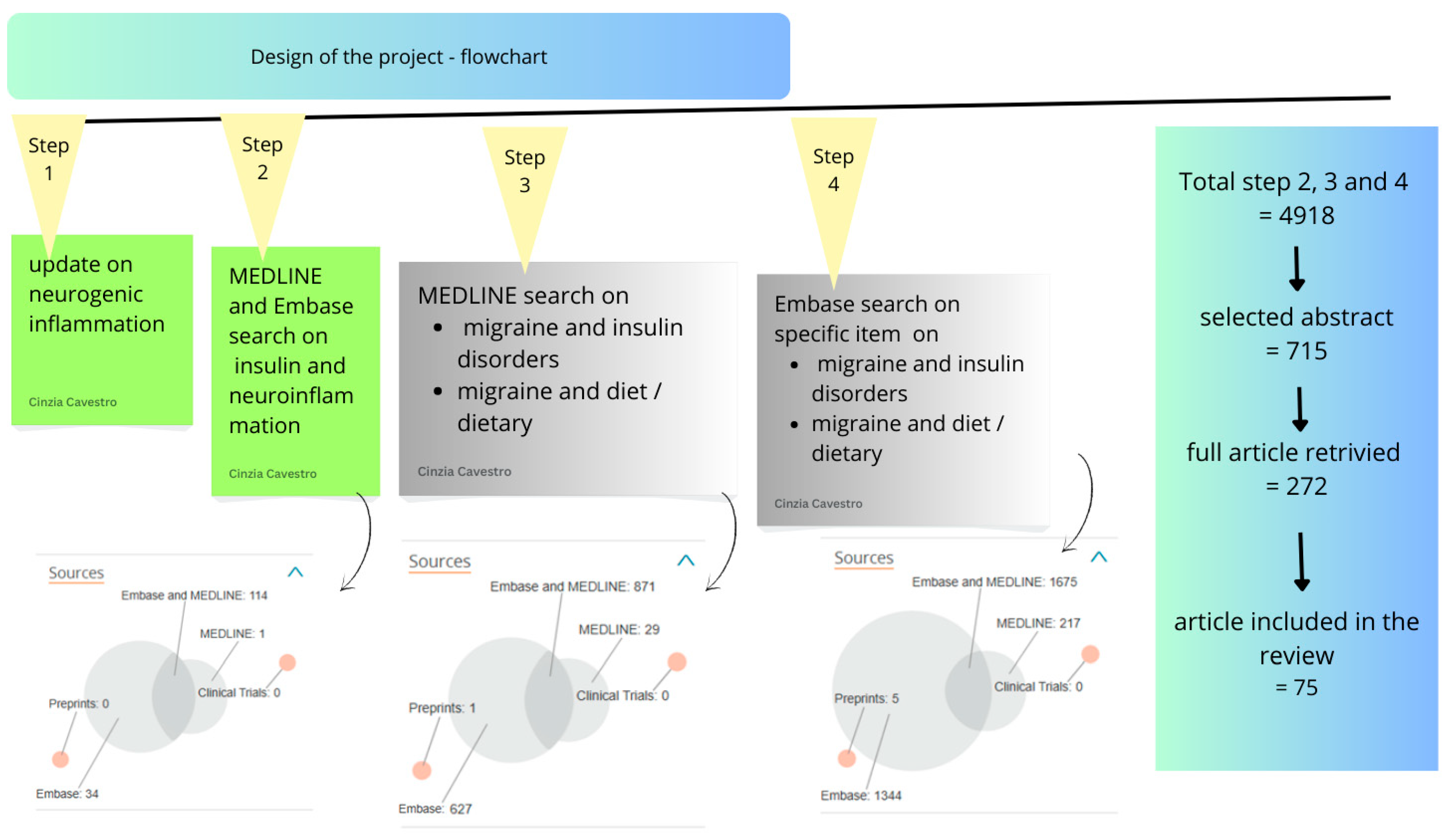
2. Methods
3. Results
3.1. Migraine and Neurogenic Inflammation
3.2. Mast Cells and Pro-Inflammatory Molecules
3.3. Migraine Pathophysiology and Immune Involvement in Cortical Spreading Depression (CSD)
3.4. Insulin Resistance and Neurogenic Inflammation
3.5. Migraine and Insulin Resistance: Clinical Evidence and Observations
3.6. Mechanistic Pathways Linking Hyperinsulinism and Migraine
3.7. Dietary and Nutritional Interventions for Migraine Management
3.8. Weight Loss
3.9. Diets
3.10. Anti-Inflammatory, Mediterranean, and DASH Diets
3.11. Ketogenic Diet/Low-Carb Diet and Migraine
3.12. The Mediterranean Diet May Offer an Alternative Approach
3.13. Do Not Skip Meals
4. Conclusions
Take-Home Messages
- ○
- Migraine involves complex interactions among genetic, neurological, inflammatory, and metabolic factors.
- ○
- Insulin resistance and hyperinsulinism significantly contribute to migraine pathogenesis.
- ○
- Metabolic dysfunction and systemic inflammation intensify migraine severity and frequency.
- ○
- Targeted dietary interventions, such as ketogenic and Mediterranean diets, can effectively reduce migraine episodes.
- ○
- Personalized nutritional and lifestyle approaches are crucial for optimal migraine management and improving patient quality of life.
![Brainsci 15 00474 i001]()
![Brainsci 15 00474 i002]()


Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2019 Diseases and Injuries Collaborators. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef] [PubMed]
- Vetvik, K.G.; MacGregor, E.A. Sex differences in the epidemiology, clinical features, and pathophysiology of migraine. Lancet Neurology 2017, 16, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Burch, R.C.; Buse, D.C.; Lipton, R.B. Migraine: Epidemiology, Burden, and Comorbidity. Neurol. Clin. 2019, 37, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Headache Classification Committee of the International Headache Society. The international classification of headache disorders, 3rd edition. Cephalalgia 2018, 38, 1–211. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, M.D.; Goadsby, P.J.; Burstein, R.; Kurth, T.; Ayata, C.; Charles, A.; Ashina, M.; van den Maagdenberg, A.M.J.M.; Dodick, D.W. Migraine. Nat. Rev. Dis. Primers. 2022, 8, 2. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Asoom, L.I.A.; Sunni, A.A.; Rafique, N.; Latif, R.; Saif, S.A.; Almandil NBAlmohazey, D.; AbdulAzeez, S.; Borgio, J.F. Genetics, pathophysiology, diagnosis, treatment, management, and prevention of migraine. Biomed. Pharmacother. 2021, 139, 111557. [Google Scholar] [CrossRef] [PubMed]
- Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. Pathophysiology of migraine: A disorder of sensory processing. Physiol. Rev. 2017, 97, 553–622. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Grangeon, L.; Lange, K.S.; Waliszewska-Prosół, M.; Onan, D.; Marschollek, K.; Wiels, W.; Mikulenka, P.; Farham, F.; Gollion, C.; Ducros, A.; et al. Genetics of migraine: Where are we now? J. Headache Pain 2023, 24, 12. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lipton, R.B.; Buse, D.C.; Nahas, S.J.; Tietjen, G.E.; Martin, V.T.; Löf, E.; Brevig, T.; Cady, R.; Diener, H.C. Risk factors for migraine disease progression: A narrative review for a patient-centered approach. J. Neurol. 2023, 270, 5692–5710. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- García-Azorín, D. The complex relationship between alcohol and migraine. Headache 2022, 62, 1245–1246. [Google Scholar] [CrossRef] [PubMed]
- Panconesi, A.; Bartolozzi, M.L.; Guidi, L. Alcohol and migraine: What should we tell patients? Curr. Pain Headache Rep. 2011, 15, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Daghlas, I.; Larsson, S.C. Alcohol, coffee consumption, and smoking in relation to migraine: A bidirectional Mendelian randomization study. Pain 2022, 163, e342–e348. [Google Scholar] [CrossRef] [PubMed]
- Viudez-Martínez, A.; Torregrosa, A.B.; Navarrete, F.; García-Gutiérrez, M.S. Understanding the biological relationship between migraine and depression. Biomol. Ther. 2024, 14, 163. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wachowska, K.; Bliźniewska-Kowalska, K.; Sławek, J.; Adamczyk-Sowa, M.; Szulc, A.; Maes, M.; Kuan-Pin, S.; Gałecki, P. Common pathomechanism of migraine and depression. Psychiatr. Pol. 2023, 57, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Furman, J.M.; Balaban, C.D.; Jacob, R.G.; Marcus, D.A. Migraine-anxiety related dizziness (MARD): A new disorder? J. Neurol. Neurosurg. Psychiatry 2005, 76, 1–8. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Burrowes, S.A.B.; Goloubeva, O.; Stafford, K.; McArdle, P.F.; Goyal, M.; Peterlin, B.L.; Haythornthwaite, J.A.; Seminowicz, D.A. Enhanced mindfulness-based stress reduction in episodic migraine-effects on sleep quality, anxiety, stress, and depression: A secondary analysis of a randomized clinical trial. Pain 2022, 163, 436–444. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jahromi, S.R.; Martami, F.; Morad Soltani, K.; Togha, M. Migraine and obesity: What is the real direction of their association? Expert. Rev. Neurother. 2023, 23, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.C.; Parapuram, S.K.; Tripathi, B.J.; Zhong, Y.; Chalam, K.V. Corticosteroids and glaucoma risk. Drugs Aging 1999, 15, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.J.; Stark, C.D. Migraine prophylaxis. Med. J. Aust. 2008, 189, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Spekker, E.; Tanaka, M.; Szabó, Á.; Vécsei, L. Neurogenic inflammation: The participant in migraine and recent advancements in translational research. Biomedicines 2021, 10, 76. [Google Scholar] [CrossRef] [PubMed Central]
- Acabelos, R.; Torrellas, C.; Fernández-Novoa, L.; López-Muñoz, F. Histamine and Immune Biomarkers in CNS Disorders. Mediat. Inflamm. 2016, 2016, 1924603. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bruno, P.P.; Carpino, F.; Carpino, G.; Zicari, A. An overview on immune system and migraine. Eur. Rev. Med. Pharmacol. Sci. 2007, 11, 245–248. [Google Scholar] [PubMed]
- Cavestro, C.; Ferrero, M.; Mandrino, S.; di Tavi, M.; Rota, E. Novelty in inflammation and immunomodulation in migraine. Curr. Pharm. Des. 2019, 25, 2919–2936. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P. Migraine and treatment-resistant depression. Prog. Brain Res. 2023, 281, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Terhart, M.; Overeem, L.H.; Hong, J.B.; Reuter, U.; Raffaelli, B. Comorbidities as risk factors for migraine onset: A systematic review and three-level meta-analysis. Eur. J. Neurol. 2025, 32, e16590. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Smitherman, T.A.; Burch, R.; Sheikh, H.; Loder, E. The prevalence, impact, and treatment of migraine and severe headaches in the United States: A review of statistics from national surveillance studies. Headache J. Head Face Pain 2013, 53, 427–436. [Google Scholar] [CrossRef]
- Breslau, N.; Lipton, R.B.; Stewart, W.F.; Schultz, L.R.; Welch, K.M. Comorbidity of migraine and depression: Investigating potential etiology and prognosis. Neurology 2003, 60, 1308–1312. [Google Scholar] [CrossRef] [PubMed]
- Tfelt-Hansen, P.; De Vries, P.; Saxena, P.R. Triptans in migraine: A comparative review of pharmacology, pharmacokinetics and efficacy. Drugs 2000, 60, 1259–1287. [Google Scholar] [CrossRef]
- Robblee, J.; Starling, A.J. SEEDS for success: Lifestyle management in migraine. Clevel. Clin. J. Med. 2019, 86, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Ashina, M.; Katsarava, Z.; Do, T.P.; Buse, D.C.; Pozo-Rosich, P.; Özge, A.; Krymchantowski, A.V.; Lebedeva, E.R.; Ravishankar, K.; Yu, S.; et al. Migraine: Epidemiology and systems of care. Lancet 2021, 397, 1485–1495. [Google Scholar] [CrossRef] [PubMed]
- Rainero, I.; Govone, F.; Gai, A.; Vacca, A.; Rubino, E. Is Migraine Primarily a Metaboloendocrine Disorder? Curr. Pain Headache Rep. 2018, 22, 36. [Google Scholar] [CrossRef] [PubMed]
- Bruijn, N.; van Lohuizen, R.; Boron, M.; Fitzek, M.; Gabriele, F.; Giuliani, G.; Melgarejo, L.; Řehulka, P.; Sebastianelli, G.; Triller, P.; et al. Influence of metabolic state and body composition on the action of pharmacological treatment of migraine. J. Headache Pain 2024, 25, 20. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bigal, M.E.; Lipton, R.B.; Holland, P.R.; Goadsby, P.J. Obesity, migraine, and chronic migraine: Possible mechanisms of interaction. Neurology 2007, 68, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Rist, P.M.; Buring, J.E.; Kurth, T. Dietary patterns according to headache and migraine status: A cross-sectional study. BMJ Open 2015, 5, e006070. [Google Scholar] [CrossRef]
- Peterlin, B.L.; Rosso, A.L.; Rapoport, A.M.; Scher, A.I. Obesity and migraine: The effect of age, gender and adipose tissue distribution. Headache J. Head Face Pain 2010, 50, 52–62. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Coppola, G.; Sirianni, G.; Di Lorenzo, G.; Bracaglia, M.; Di Lenola, D.; Siracusano, A.; Rossi, P.; Pierelli, F. Migraine improvement during short lasting ketogenesis: A proof-of-concept study. Eur. J. Neurol. 2015, 22, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Kristoffersen, E.S.; Børte, S.; Hagen, K.; Zwart, J.A.; Winsvold, B.S. Migraine, obesity and body fat distribution—A population-based study. J. Headache Pain. 2020, 21, 97. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ramírez-Salazar, S.A.; Herren, C.; McCartney, J.; Ortiz García, J.G. Dietary Insights in Neurological Diseases. Curr. Neurol. Neurosci Rep. 2021, 21, 55. [Google Scholar] [CrossRef] [PubMed]
- Weiller, C.; May, A.; Limmroth, V.; Jüptner, M.; Kaube, H.; Schayck, R.V.; Coenen, H.H.; Diener, H.C. Brain stem activation in spontaneous human migraine attacks. Nat. Med. 1995, 1, 658–660. [Google Scholar] [CrossRef] [PubMed]
- Bahra, A.; Matharu, M.; Buche, C.; Frackowiak, R.; Goadsby, P. Brainstem activation specific to migraine headache. Lancet 2001, 357, 1016–1017. [Google Scholar] [CrossRef]
- Goadsby, P.; Charbit, A.; Andreou, A.; Akerman, S.; Holland, P. Neurobiology of migraine. Neuroscience 2009, 161, 327–341. [Google Scholar] [CrossRef]
- Olesen, J.; Burstein, R.; Ashina, M.; Tfelt-Hansen, P. Origin of pain in migraine: Evidence for peripheral sensitisation. Lancet Neurol. 2009, 8, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.; Edvinsson, L.; Goadsby, P. CGRP and its receptors provide new insights into migraine pathophysiology. Nat. Rev. Neurol. 2010, 6, 573–582. [Google Scholar] [CrossRef]
- Strassman, A.; Levy, D. Response Properties of Dural Nociceptors in Relation to Headache. J. Neurophysiol. 2006, 95, 1298–1306. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.; Wolff, H. Mechanism of migraine headache and action of ergotamine tartrate. Arch. Neurol. 1938, 39, 737–763. [Google Scholar] [CrossRef]
- Welch, K. Stroke and migraine—The spectrum of cause and effect. Funct. Neurol. 2003, 18, 121–126. [Google Scholar]
- Moskowitz, M. The neurobiology of vascular head pain. Ann. Neurol. 1984, 16, 157–168. [Google Scholar] [CrossRef]
- Williamson, D.; Hargreaves, R. Neurogenic Inflammation in the Context of Migraine. Microsc. Res. Tech. 2001, 53, 167–178. [Google Scholar] [CrossRef]
- Moskowitz, M.; Reinhard, J.; Romero, J.; Melamed, E.; Pettibone, D.J. Neurotrasmitters and the fifth cranial nerve: Is there a relation to the headache fase of migraine ? Lancet 1979, 2, 883–885. [Google Scholar] [CrossRef]
- Moskowitz, M. Neurogenic inflammation in the pathohysiology and treatment of migraine. Neurology 1993, 43 (Suppl. S3), S16–S20. [Google Scholar]
- Akerman, S.; Holland, P.; Hoffmann, J. Pearls and pitfalls in experimental in vivo models of migraine: Dural trigeminovascular nociception. Cephalalgia 2013, 33, 577–592. [Google Scholar] [CrossRef] [PubMed]
- Lembeck, H. Substance P as neurogenic mediator of antidromic vasodilation and neurogenic lasma extravasation. Naunyn Schmiedeberg’s Arch. Pharmacol. 1979, 310, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Jansen, I.; Alafaci, C.; McCulloch, J.; Uddman, R.; Edvinsson, L. Tachykinins (substance P, neurokinin A, neuropeptide K, and neurokinin B) in the cerebral circulation: Vasomotor responses in vitro and in situ. J. Cereb. Blood Flow. Metab. 1991, 11, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Brain, S.; Williams, T.J.; Tippins, J.R.; Morris, H.R.; MacIntyre, I. Calcitonin gene-related peptide is a potent vasodilator. Nature 1985, 313, 54–56. [Google Scholar] [CrossRef]
- Markowitz, S.; Saito, K.; Moskowitz, M. Neurogenicallymediated leakage of plasma protein occurs from blood vessels in dura mater but not brain. J. Neurosci. 1987, 7, 4129–4136. [Google Scholar] [CrossRef]
- Dimitriadou, V.; Buzzi, M.; Moskowitz, M.; Theoharides, T.C. Trigeminal sensory fiber stimulation induces morphological changesreflecting secretion in rat dura mater mast cells. Neuroscience 1991, 44, 97–112. [Google Scholar] [CrossRef]
- Matsubara, T.; Moskowitz, M.; Huang, Z. UK-14,304, R(−)-α-methyl-histamine and SMS 201-995 block plasma protein leakage within dura mater by prejunctional mechanisms. Eur. J. Pharmacol. 1992, 224, 145–150. [Google Scholar]
- Buzzi, M.; Sakas, D.; Moskowitz, M. Indomethacin and acetylsalicylic acid block neurogenic plasma protein extravasation in rat dura mater. Eur. J. Pharmacol. 1989, 165, 251–258. [Google Scholar] [CrossRef]
- Moskowitz, M.; Macfarlane, R. Neurovascular and molecular mechanisms in migraine headaches. Cerebrovasc. Brain Metab. Rev. 1993, 5, 159–177. [Google Scholar]
- Lee, W.; Limmroth, V.; Ayata, C.; Cutrer, F.M.; Waeber, C.; Yu, X.; Moskowitz, M.A. Peripheral GABAA receptor-mediated effects of sodium valproate on dural plasma protein extravasation to substance P and trigeminal stimulation. Br. J. Pharmacol. 1995, 116, 1661–1667. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Buzzi, M.; Dimitriadou, V.; Theoharides, T.; Moskowitz, M. 5-Hydroxytryptamine receptor agonists for the abortive treatment of vascular headaches block mast cell, endothelial and platelet activation within the rat dura mater after trigeminal stimulation. Brain Res. 1992, 583, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Shepheard, S.; Williamson, D.; Hill, R.; Hargreaves, R. The non-peptide neurokinin1 receptor antagonist, RP 67580, blocks neurogenic plasma extravasation in the dura mater of rats. Br. J. Pharmacol. 1993, 108, 11–12. [Google Scholar] [CrossRef]
- Jansen, I.; Uddman, R.; Ekman, R.; Olesen, J.; Ottosson, A.; Edvinsson, I. Distribution and effects of neuropeptide Y, vasoactive intestinal peptide, sustance P, and calcitonin gene-related peptide in uman middle meningeal arteries: Comparison with cerebral and temporal arteries. Peptides 1992, 13, 527–536. [Google Scholar] [CrossRef]
- Edvinson, L. Sensory nerves in man and their role in primary headache. Cephalalgia 2001, 21, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Uddman, R. Adrenergic, cholinergic and peptidergic nerve fibres in dura mater—Involvement in headache? Cephalalgia 1981, 1, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Sicuteri, F. Mast cells and their active substances: Their role in the pathogenesis of migraine. Headache. 1963, 3, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R. Neurogenic inflammation and its role in migraine. Semin. Immunopathol. 2018, 40, 301–314. [Google Scholar] [CrossRef]
- Hendriksen, E.; van Bergeijk, D.; Oosting, R.; Redegeld, F. Mast cells in neuroinflammation and brain disorders. Neurosci. Biobehav. Rev. 2017, 79, 119–133. [Google Scholar] [CrossRef]
- Eftekhari, S.; Warfvinge, K.; Blixt, F.; Edvinsson, L. Differentiation of nerve fibers storing CGRP and CGRP receptors in the peripheral trigeminovascular system. J. Pain 2013, 14, 1289–1303. [Google Scholar] [CrossRef]
- Galli, S. New concepts about the mast cell. N. Engl. J. Med. 1993, 328, 257–265. [Google Scholar] [CrossRef]
- Artico, M.; Cavallotti, C. Catecholaminergic and acetylcholine esterase containing nerves of cranial and spinal dura mater in humans and rodents. Microsc. Res. Tech. 2001, 53, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Strassman, A.; Weissner, W.; Williams, M.; Ali, S.; Levy, D. Axon diameters and intradural trajectories of the dural innervation in the rat. J. Comp. Neurol. 2004, 473, 364–376. [Google Scholar] [CrossRef]
- Orr, E.; Pace, K. The significance of mast cells as a source of histamine in mouse brain. J. Neurochem. 1984, 42, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Strassman, A.; Burstein, R.; Levy, D. Sensitization and activation of intracranial meningeal nociceptors by mast cell mediators. J. Pharmacol. Exp. Ther. 2007, 322, 806–912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Levy, D. Modulation of meningeal nociceptors mechanosensitivity by peripheral proteinase-activated receptor-2: The role of mast cells. Cephalalgia 2008, 28, 276–284. [Google Scholar] [CrossRef]
- Levy, D.; Burstein, R.; Kainz, V.; Jakubowski, M.; Strassman, A. Mast cell degranulation activates a pain pathway underlying migraine headache. Pain 2007, 130, 166–176. [Google Scholar] [CrossRef]
- Reuter, U.; Bolay, H.; Jansen-Olesen, I.; Chiarugi, A.; del Rio, M.S.; Letourneau, R.; Theoharides, T.C.; Waeber, C.; Moskowitz, M.A. Delayed inflammation in rat meninges: Implications for migraine pathophysiology. Brain 2001, 124, 2490–2502. [Google Scholar] [CrossRef]
- Cuesta, M.; Quintero, L.; Pons, H.; Suarez-Roca, H. Substance P and calcitonin gene-related peptide increase IL-1 beta, IL-6 and TNF alpha secretion from human peripheral blood mononuclear cells. Neurochem. Int. 2002, 40, 301–306. [Google Scholar] [CrossRef]
- Levite, M. Neuropeptides, by direct interaction with T cells, induce cytokine secretion and break the commitment to a distinct T helper phenotype. Proc. Natl. Acad. Sci. USA 1998, 95, 12544–12549. [Google Scholar] [CrossRef]
- Benveniste, E. Inflammatory cytokines within the central nervous system: Sources, function, and mechanism of action. Am. J. Physiol. 1992, 263, C1–C16. [Google Scholar] [CrossRef]
- Benveniste, E. Cytokine actions in the central nervous system. Cytokine Growth Factor. Rev. 1998, 9, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Kemper, R.; Meijler, W.; Korf, J.; Ter Horst, G. Migraine and function of the immune system: A meta-analysis of clinical literature published between 1966 and 1999. Cephalalgia 2001, 21, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Perini, F.; D’Andrea, G.; Galloni, E.; Pignatelli, F.; Billo, G.; Alba, S.; Bussone, G.; Toso, V. Plasma cytokine levels in migraineurs and controls. Headache 2005, 45, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Sarchielli, P.; Alberti, A.; Baldi, A.; Coppola, F.; Rossi, C.; Pierguidi, L.; Floridi, A.; Calabresi, P. Proinflammatory cytokines, adhesion molecules, and lymphocyte integrin expression in the internal jugular blood of migraine patients without aura assessed ictally. Headache 2006, 46, 200–207. [Google Scholar] [CrossRef]
- Levite, M.; Cahalon, L.; Hershkoviz, R.; Steinman, L.; Lider, O. Neuropeptides, via specific receptors, regulate T cell adhesion to fibronectin. J. Immunol. 1998, 15, 993–1000. [Google Scholar] [CrossRef]
- Gerring, Z.; Powell, J.; Montgomery, G.; Nyholt, D. Genome-wide analysis of blood gene expression in migraine implicates immune-inflammatory pathways. Cephalalgia 2018, 38, 292–303. [Google Scholar] [CrossRef]
- Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and associated neurological symptoms, CSD, sensitization and modulation of pain. Pain 2013, 154 (Suppl. S1), 1. [Google Scholar] [CrossRef]
- Close, L.; Eftekhari, S.; Wang, M.; Charles, A.; Russo, A. Cortical spreading depression as a site of origin for migraine: Role of CGRP. Cephalalgia 2019, 39, 428–434. [Google Scholar] [CrossRef]
- Schain, A.; Melo-Carrillo, A.; Borsook, D.; Grutzendler, J.; Strassman, A.; Burstein, R. Activation of pial and dural macrophages and dendritic cells by cortical spreading depression. Ann. Neurol. 2018, 83, 508–521. [Google Scholar] [CrossRef]
- Jessen, N.; Munk, A.; Lundgaard, I.; Nedergaard, M. The Glymphatic System: A Beginner’s Guide. Neurochem. Res. 2015, 40, 2583–2599. [Google Scholar] [CrossRef]
- Schain, A.; Melo-Carrillo, A.; Strassman, A.; Burstein, R. Cortical Spreading Depression Closes Paravascular Space and Impairs Glymphatic Flow: Implications for Migraine Headache. J. Neurosci. 2017, 15, 2904–2915. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, D.S.; Mainero, C.; Ichijo, E.; Ward, N.; Granziera, C.; Zürcher, N.R.; Akeju, O.; Bonnier, G.; Price, J.; Hooker, J.M.; et al. Imaging of neuroinflammation in migraine with aura: A [11C]PBR28 PET/MRI study. Neurology 2019, 92, e2038–e2050. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Loggia, M.; Chonde, D.; Akeju, O.; Arabasz, G.; Catana, C.; Edwards, R.R.; Hill, E.; Hsu, S.; Izquierdo-Garcia, D.; Ji, R.R.; et al. Evidence for brain glial activation in chronic pain patients. Brain 2015, 138, 604–615. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mainero, C.; Boshyan, J.; Hadjikhani, N. Altered functional magnetic resonance imaging resting-state connectivity in periaqueductal gray networks in migraine. Ann. Neurol. 2011, 70, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Salahi, M.; Parsa, S.; Nourmohammadi, D.; Razmkhah, Z.; Salimi, O.; Rahmani, M.; Zivary, S.; Askarzadeh, M.; Tapak, M.A.; Vaezi, A.; et al. Immunologic aspects of migraine: A review of literature. Front. Neurol. 2022, 13, 944791. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, R.; Luo, M.; Gong, H.; Li, K.; Zhao, Q. Inflammo-immune perspective on the association of eight migraine risk factors with migraine: A multi-omics Mendelian randomization study. Front. Neurol. 2024, 15, 1440995. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlying molecular mechanisms. J. Cell Physiol. 2019, 234, 8152–8161. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular Mechanisms Linking Oxidative Stress and Diabetes Mellitus. Oxid. Med. Cell Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Barghchi, H.; Milkarizi, N.; Belyani, S.; Norouzian Ostad, A.; Askari, V.R.; Rajabzadeh, F.; Goshayeshi, L.; Ghelichi Kheyrabadi, S.Y.; Razavidarmian, M.; Dehnavi, Z.; et al. Pomegranate (Punica granatum L.) peel extract ameliorates metabolic syndrome risk factors in patients with non-alcoholic fatty liver disease: A randomized double-blind clinical trial. Nutr. J. 2023, 22, 40. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fu, Z.; Gilbert, E.R.; Liu, D. Regulation of insulin synthesis and secretion and pancreatic Beta-cell dysfunction in diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Paseban, M.; Butler, A.E.; Sahebkar, A. Mechanisms of statin-induced new-onset diabetes. J. Cell Physiol. 2019, 234, 12551–12561. [Google Scholar] [CrossRef] [PubMed]
- Shayan, M.; Kamalian, S.; Sahebkar, A.; Tayarani-Najaran, Z. Flaxseed for Health and Disease: Review of Clinical Trials. Comb. Chem. High. Throughput Screen. 2020, 23, 699–722. [Google Scholar] [CrossRef] [PubMed]
- Vinuesa, A.; Pomilio, C.; Gregosa, A.; Bentivegna, M.; Presa, J.; Bellotto, M.; Saravia, F.; Beauquis, J. Inflammation and Insulin Resistance as Risk Factors and Potential Therapeutic Targets for Alzheimer’s Disease. Front. Neurosci. 2021, 15, 653651. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gruber, H.J.; Bernecker, C.; Pailer, S.; Fauler, G.; Horejsi, R.; Möller, R.; Lechner, A.; Fazekas, F.; Truschnig-Wilders, M. Hyperinsulinaemia in migraineurs is associated with nitric oxide stress. Cephalalgia 2010, 30, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Cai, W.; Hoover, B.; Kahn, C.R. Insulin action in the brain: Cell types, circuits, and diseases. Trends Neurosci. 2022, 45, 384–400. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rainero, I.; Limone, P.; Ferrero, M.; Valfrè, W.; Pelissetto, C.; Rubino, E.; Gentile, S.; Lo Giudice, R.; Pinessi, L. Insulin sensitivity is impaired in patients with migraine. Cephalalgia. 2005, 25, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Guldiken, B.; Guldiken, S.; Demir, M.; Turgut, N.; Kabayel, L.; Ozkan, H.; Ozcelik, E.; Tugrul, A. Insulin resistance and high sensitivity C-reactive protein in migraine. Can. J. Neurol. Sci. 2008, 35, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Bernecker, C.; Ragginer, C.; Fauler, G.; Horejsi, R.; Möller, R.; Zelzer, S.; Lechner, A.; Wallner-Blazek, M.; Weiss, S.; Fazekas, F.; et al. Oxidative stress is associated with migraine and migraine-related metabolic risk in females. Eur. J. Neurol. 2011, 18, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Bernecker, C.; Pailer, S.; Kieslinger, P.; Horejsi, R.; Möller, R.; Lechner, A.; Wallner-Blazek, M.; Weiss, S.; Fazekas, F.; Truschnig-Wilders, M.; et al. Increased matrix metalloproteinase activity is associated with migraine and migraine-related metabolic dysfunctions. Eur. J. Neurol. 2011, 18, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Bernecker, C.; Pailer, S.; Kieslinger, P.; Horejsi, R.; Möller, R.; Lechner, A.; Wallner-Blazek, M.; Weiss, S.; Fazekas, F.; Truschnig-Wilders, M.; et al. GLP-2 and leptin are associated with hyperinsulinemia in non-obese female migraineurs. Cephalalgia 2010, 30, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Sacco, S.; Altobelli, E.; Ornello, R.; Ripa, P.; Pistoia, F.; Carolei, A. Insulin resistance in migraineurs: Results from a case-control study. Cephalalgia. 2014, 34, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Cavestro, C.; Rosatello, A.; Micca, G.; Ravotto, M.; Marino, M.P.; Asteggiano, G.; Beghi, E. Insulin metabolism is altered in migraineurs: A new pathogenic mechanism for migraine? Headache. 2007, 47, 1436–1442. [Google Scholar] [CrossRef] [PubMed]
- Bhoi, S.K.; Kalita, J.; Misra, U.K. Metabolic syndrome and insulin resistance in migraine. J. Headache Pain 2012, 13, 321–326. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, X.; Li, X.; Diao, Y.; Meng, S.; Xing, Y.; Zhou, H.; Yang, D.; Sun, J.; Chen, H.; Zhao, Y. Are Glucose and Insulin Metabolism and Diabetes Associated with Migraine? A Community-Based, Case-Control Study. J. Oral Facial Pain Headache 2017, 31, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Hosseinpour, M.; Maleki, F.; Khoramdad, M.; Sullman, M.J.M.; Nejadghaderi, S.A.; Kolahi, A.A.; Safiri, S. A systematic literature review of observational studies of the bilateral association between diabetes and migraine. Diabetes Metab. Syndr. 2021, 15, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Hussein, M.; Magdy, R.; Khamis, A.; Al-Azayem, S.A.; Othman, A.M.; Ahmed, A.; Osama, W. The potential impact of insulin resistance and metabolic syndrome on migraine headache characteristics. BMC Neurol. 2022, 22, 422. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rosta, J.; Tóth, M.; Friedrich, N.; Sántha, P.; Jancsó, G.; Dux, M. Insulin sensitizes neural and vascular TRPV1 receptors in the trigeminovascular system. J. Headache Pain 2022, 23, 7. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fava, A.; Pirritano, D.; Consoli, D.; Plastino, M.; Casalinuovo, F.; Cristofaro, S.; Colica, C.; Ermio, C.; De Bartolo, M.; Opipari, C.; et al. Chronic migraine in women is associated with insulin resistance: A cross-sectional study. Eur. J. Neurol. 2014, 21, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Gur-Ozmen, S.; Karahan-Ozcan, R. Factors Associated with Insulin Resistance in Women with Migraine: A Cross-Sectional Study. Pain Med. 2019, 20, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Grinberg, Y.Y.; Zitzow, L.A.; Kraig, R.P. Intranasally administered IGF-1 inhibits spreading depression in vivo. Brain Res. 2017, 1677, 47–57. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cavestro, C.; Bedogni, G.; Molinari, F.; Mandrino, S.; Rota, E.; Frigeri, M.C. Alpha-Lipoic Acid Shows Promise to Improve Migraine in Patients with Insulin Resistance: A 6-Month Exploratory Study. J. Med. Food 2018, 21, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Olivito, I.; Simona, F.; Tarsitano, A.; Pagliuso, M.; Tarantino, C.; De Lorenzo, A.; Alò, R.; Avolio, E.; Facciolo, R.M. Mediterranean ketogenic diet accounts for reduced pain frequency and intensity in patients with chronic migraine: A pilot study. Clin. Nutr. 2024, 43, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Halloum, W.; Dughem, Y.A.; Beier, D.; Pellesi, L. Glucagon-like peptide-1 (GLP-1) receptor agonists for headache and pain disorders: A systematic review. J. Headache Pain 2024, 25, 112. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, C.; He, Y.; Liu, L. Identifying therapeutic target genes for migraine by systematic druggable genome-wide Mendelian randomization. J. Headache Pain 2024, 25, 100. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- May, A.; Schulte, L.H. Chronic migraine: Risk factors, mechanisms and treatment. Nat. Rev. Neurol. 2016, 12, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Di Vincenzo, A.; Beghetto, M.; Vettor, R.; Tana, C.; Rossato, M.; Bond, D.S.; Pagano, C. Effects of Surgical and Non-surgical Weight Loss on Migraine Headache: A Systematic Review and Meta-Analysis. Obes. Surg. 2020, 30, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Gazerani, P. A Bidirectional View of Migraine and Diet Relationship. Neuropsychiatr. Dis. Treat. 2021, 17, 435–451. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gazerani, P. Diet and migraine: What is proven? Curr. Opin. Neurol. 2023, 36, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Evers, S.; Grube, H.C.; Gendolla, A.; Gaul, C.; Ewald, K.; Witt, O.; Voggel, J.; Nitschke, M.; Sina, C.; Thaci, D.; et al. Efficacy of Digital Therapeutic sinCephalea for Personalised Nutrition Versus Control for Migraine Prevention: A 12-Week Open-Label Randomised Clinical Trial. SSRN 2025. [Google Scholar] [CrossRef]
- Ghoreishy, S.M.; Askari, G.; Mohammadi, H.; Campbell, M.S.; Khorvash, F.; Arab, A. Associations between potential inflammatory properties of the diet and frequency, duration, and severity of migraine headaches: A cross-sectional study. Sci. Rep. 2022, 12, 2878. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hajishizari, S.; Mirzababaei, A.; Abaj, F.; Bahrampour, N.; Moradi, S.; Clark, C.C.T.; Mirzaei, K. The association between dietary antioxidant quality score and intensity and frequency of migraine headaches among women: A cross-sectional study. BMC Women’s Health 2024, 24, 497. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Arab, A.; Khorvash, F.; Karimi, E.; Hadi, A.; Askari, G. Associations between adherence to Mediterranean dietary pattern and frequency, duration, and severity of migraine headache: A cross-sectional study. Nutr. Neurosci. 2023, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, A.V.; Danilov, A.B.; Clayton, P.; Moskalev, A.A.; Karasev, A.V.; Tarasevich, A.F.; Vorobyeva, Y.D.; Novikov, V.N. Perspectives on Neuronutrition in Prevention and Treatment of Neurological Disorders. Nutrients 2023, 15, 2505. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Khalili, P.; Mirzababaei, A.; Abaj, F.; Ansari, S.; Harsini, A.R.; Hajmir, M.M.; Clark, C.C.T.; Mirzaei, K. The association between adherence to alternative healthy Diet Index (AHEI) and severity, disability, duration, and frequency of migraine headache among women: A cross-sectional study. Nutr. J. 2023, 22, 41. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Arab, A.; Khorvash, F.; Karimi, E.; Heidari, Z.; Askari, G. The effects of the dietary approaches to stop hypertension (DASH) diet on oxidative stress and clinical indices of migraine patients: A randomized controlled trial. Nutr. Neurosci. 2022, 25, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Hajjarzadeh, S.; Bakhshimoghaddam, F.; Behrouz, M.; Nikniaz, Z.; Mahdavi, R.; Shalilahmadi, D.; Karandish, M. The relation of adherence to the DASH diet with migraine attack frequency and pain intensity in Iranian women: A cross-sectional study. Nutr. Neurosci. 2024, 27, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Bakhshimoghaddam, F.; Shalilahmadi, D.; Mahdavi, R.; Nikniaz, Z.; Karandish, M.; Hajjarzadeh, S. Association of dietary and lifestyle inflammation score (DLIS) with chronic migraine in women: A cross-sectional study. Sci. Rep. 2024, 14, 16406, Erratum in Sci Rep. 2024, 14, 18954. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tirani, S.A.; Lotfi, K.; Shahdadian, F.; Hajhashemy, Z.; Rouhani, P.; Saneei, P. Dietary Phytochemical Index in Relation to Metabolic Health Status, Serum Adropin, and Brain-Derived Neurotrophic Factor Levels in Adults. Curr. Dev. Nutr. 2024, 8, 102103. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Razeghi Jahromi, S.; Togha, M.; Ghorbani, Z.; Hekmatdoost, A.; Khorsha, F.; Rafiee, P.; Shirani, P.; Nourmohammadi, M.; Ansari, H. The association between dietary tryptophan intake and migraine. Neurol. Sci. 2019, 40, 2349–2355. [Google Scholar] [CrossRef] [PubMed]
- Khorsha, F.; Mirzababaei, A.; Ghodoosi, N.; Togha, M.; Yekaninejad, M.S.; Askarpour, M.; Mirzaei, K. Association between diet and migraine characteristics: The role of dietary inflammatory index. Curr. J. Neurol. 2020, 19, 67–75. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Martins, L.B.; Braga Tibães, J.R.; Dos Santos Rodrigues, A.M.; Hassanzadeh Keshteli, A.; Karam Vono, C.; Borges, E.; Borges, J.; Horta, P.M.; Teixeira, A.L.; Matos Ferreira, A.V. The quality and inflammatory index of the diet of patients with migraine. Nutr. Neurosci. 2022, 25, 2092–2099. [Google Scholar] [CrossRef] [PubMed]
- Khorsha, F.; Mirzababaei, A.; Togha, M.; Mirzaei, K. Association of dietary diversity score (DDS) and migraine headache severity among women. Neurol. Sci. 2021, 42, 3403–3410. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, D.; Wu, F.; Dong, Z.; Yu, S. Association between inflammatory potential of diet and self-reported severe headache or migraine: A cross-sectional study of the National Health and Nutrition Examination Survey. Nutrition 2023, 113, 112098. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Q.; Dong, Z.; Yu, S. Dietary zinc intake and migraine in adults: A cross-sectional analysis of the National Health and Nutrition Examination Survey 1999–2004. Headache 2023, 63, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Magdy, R.; Eid, R.A.; Hassan, M.; Abdelghaffar, M.; El Sayed, A.F.; Mohammed, Z.; Hussein, M. The potential impact of nutritional intake on symptoms severity in patients with comorbid migraine and irritable bowel syndrome. BMC Neurol. 2022, 22, 199. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mansouri, M.; Sharifi, F.; Varmaghani, M.; Yaghubi, H.; Shokri, A.; Moghadas-Tabrizi, Y.; Keshtkar, A.; Sadeghi, O. Dairy consumption in relation to primary headaches among a large population of university students: The MEPHASOUS study. Complement. Ther. Med. 2020, 48, 102269. [Google Scholar] [CrossRef] [PubMed]
- Martami, F.; Togha, M.; Qorbani, M.; Shahamati, D.; Salami, Z.; Shab-Bidar, S. Association of dietary patterns with migraine: A matched case-control study. Curr. J. Neurol. 2023, 22, 87–95. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Menon, S.; Lea, R.A.; Ingle, S.; Sutherland, M.; Wee, S.; Haupt, L.M.; Palmer, M.; Griffiths, L.R. Effects of dietary folate intake on migraine disability and frequency. Headache 2015, 55, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Nazari, F.; Eghbali, M. Migraine and its relationship with dietary habits in women. Iran. J. Nurs. Midwifery Res. 2012, 17 (Suppl. S1), S65–S71. [Google Scholar] [PubMed] [PubMed Central]
- Parohan, M.; Sarraf, P.; Javanbakht, M.H.; Foroushani, A.R.; Ranji-Burachaloo, S.; Djalali, M. The synergistic effects of nano-curcumin and coenzyme Q10 supplementation in migraine prophylaxis: A randomized, placebo-controlled, double-blind trial. Nutr. Neurosci. 2021, 24, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Parohan, M.; Sarraf, P.; Javanbakht, M.H.; Ranji-Burachaloo, S.; Djalali, M. Effect of coenzyme Q10 supplementation on clinical features of migraine: A systematic review and dose-response meta-analysis of randomized controlled trials. Nutr. Neurosci. 2020, 23, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Slavin, M.; Ailani, J. A Clinical Approach to Addressing Diet with Migraine Patients. Curr. Neurol. Neurosci. Rep. 2017, 17, 17. [Google Scholar] [CrossRef] [PubMed]
- Abdolahi, M.; Jafarieh, A.; Sarraf, P.; Sedighiyan, M.; Yousefi, A.; Tafakhori, A.; Abdollahi, H.; Salehinia, F.; Djalali, M. The Neuromodulatory Effects of ω-3 Fatty Acids and Nano-Curcumin on the COX-2/iNOS Network in Migraines: A Clinical Trial Study from Gene Expression to Clinical Symptoms. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Abdolahi, M.; Tafakhori, A.; Togha, M.; Okhovat, A.A.; Siassi, F.; Eshraghian, M.R.; Sedighiyan, M.; Djalali, M.; Mohammadzadeh Honarvar, N.; Djalali, M. The synergistic effects of ω-3 fatty acids and nano-curcumin supplementation on tumor necrosis factor (TNF)-α gene expression and serum level in migraine patients. Immunogenetics. 2017, 69, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Long, X.; Wang, J. Association between dietary vitamin C intake and migraine in adults: A cross-sectional study of the National Health and Nutrition Examination Survey. J. Hum. Nutr. Diet. 2024, 37, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Guo, Y.; Xia, M.; Zhang, J.; Zang, W. Dietary intake of thiamine and riboflavin in relation to severe headache or migraine: A cross-sectional survey. Headache 2022, 62, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez-de-Sevilla, G.; González-de-la-Flor, Á. Impact of Fatty Acid Supplementation on Migraine Outcomes: A Systematic Review and Meta-analysis. Nutr. Rev. 2025, nuae219. [Google Scholar] [CrossRef] [PubMed]
- Finelli, F.; Catalano, A.; De Lisa, M.; Ferraro, G.A.; Genovese, S.; Giuzio, F.; Salvia, R.; Scieuzo, C.; Sinicropi, M.S.; Svolacchia, F.; et al. CGRP Antagonism and Ketogenic Diet in the Treatment of Migraine. Medicina 2024, 60, 163. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Di Lorenzo, C.; Currà, A.; Sirianni, G.; Coppola, G.; Bracaglia, M.; Cardillo, A.; De Nardis, L.; Pierelli, F. Diet transiently improves migraine in two twin sisters: Possible role of ketogenesis? Funct. Neurol. 2013, 28, 305–358. [Google Scholar] [PubMed] [PubMed Central]
- Caminha, M.C.; Moreira, A.B.; Matheus, F.C.; Rieger, D.K.; Moreira, J.D.; Dalmarco, E.M.; Demarchi, I.G.; Lin, K. Efficacy and tolerability of the ketogenic diet and its variations for preventing migraine in adolescents and adults: A systematic review. Nutr. Rev. 2022, 80, 1634–1647. [Google Scholar] [CrossRef] [PubMed]
- Barbanti, P.; Fofi, L.; Aurilia, C.; Egeo, G.; Caprio, M. Ketogenic diet in migraine: Rationale, findings and perspectives. Neurol. Sci. 2017, 38 (Suppl. S1), 111–115. [Google Scholar] [CrossRef] [PubMed]
- Tereshko, Y.; Dal Bello, S.; Di Lorenzo, C.; Pittino, A.; Filippi, F.; Belgrado, E.; Lettieri, C.; Merlino, G.; Gigli, G.L.; Valente, M. The Effect of Three Different Ketogenic Diet Protocols on Migraine and Fatigue in Chronic and High-Frequency Episodic Migraine: A Pilot Study. Nutrients 2023, 15, 4334. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Di Lorenzo, C.; Coppola, G.; Bracaglia, M.; Di Lenola, D.; Sirianni, G.; Rossi, P.; Di Lorenzo, G.; Parisi, V.; Serrao, M.; Cervenka, M.C.; et al. A ketogenic diet normalizes interictal cortical but not subcortical responsivity in migraineurs. BMC Neurol. 2019, 19, 136. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lovati, C.; d’Alessandro, C.M.; Ventura, S.D.; Muzio, F.; Pantoni, L. Ketogenic diet in refractory migraine: Possible efficacy and role of ketone bodies-a pilot experience. Neurol. Sci. 2022, 43, 6479–6485. [Google Scholar] [CrossRef] [PubMed]
- Maggioni, F.; Margoni, M.; Zanchin, G. Ketogenic diet in migraine treatment: A brief but ancient history. Cephalalgia 2011, 31, 1150–1151. [Google Scholar] [CrossRef] [PubMed]
- Caprio, M.; Moriconi, E.; Camajani, E.; Feraco, A.; Marzolla, V.; Vitiello, L.; Proietti, S.; Armani, A.; Gorini, S.; Mammi, C.; et al. Very-low-calorie ketogenic diet vs hypocaloric balanced diet in the prevention of high-frequency episodic migraine: The EMIKETO randomized, controlled trial. J. Transl. Med. 2023, 21, 692. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Evcili, G.; Utku, U.; Öğün, M.N.; Özdemir, G. Early and long period follow-up results of low glycemic index diet for migraine prophylaxis. Agri 2018, 30, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Cavestro, C.; Rosatello, A.; Micca, G.M.; Asteggiano, G. Alterations in glucose-insulin metabolism in migraineurs: Prospective study on the clinical impact of diet on migraine severity and meta-bolic data in a migraineur population—Preliminary results. ORAL COMMUNICATIONS WITH POSTERS. J. Headache Pain 2006, 7, 287. [Google Scholar] [CrossRef]
- Legesse, S.M.; Addila, A.E.; Jena, B.H.; Jikamo, B.; Abdissa, Z.D.; Hailemarim, T. Irregular meal and migraine headache: A scoping review. BMC Nutr. 2025, 11, 60. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sacks, F.M.; Carey, V.J.; Anderson, C.A.; Miller, E.R., 3rd; Copeland, T.; Charleston, J.; Harshfield, B.J.; Laranjo, N.; McCarron, P.; Swain, J.; et al. Effects of high vs low glycemic index of dietary carbohydrate on cardiovascular disease risk factors and insulin sensitivity: The OmniCarb randomized clinical trial. JAMA 2014, 312, 2531–2541. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance/Peptide | Vasodilation | Plasma Extravasation | Mechanism of Action | Site of Action | Modulating Agents (Agonists/Antagonists) |
|---|---|---|---|---|---|
| CGRP | YES | NO | Acts on CGRP receptors and induces prolonged vasodilation | Cerebral and meningeal vessels | Inhibited by CGRP antagonists (gepants) and triptans |
| Substance P (SP) | YES | YES | Acts on endothelial NK1 receptors and induces extravasation and vasodilation | Vascular endothelium of dura mater | Inhibited by NK1 receptor antagonists |
| Neurokinin A (NKA) | YES | YES | Acts on endothelial receptors and induces extravasation and vasodilation | Vascular endothelium of dura mater | Inhibited by NK2 receptor antagonists |
| PACAP | YES | NO | Activates PAC1 receptors and induces cerebral vasodilation | Cerebral arteries (PAC1) and ganglia | Stimulated by PACAP; inhibited by PAC1 antagonists (under investigation) |
| Histamine | YES | YES | Inflammatory mediator and increases vascular permeability and dilation | Vascular endothelium and inflamed tissues | Inhibited by antihistamines (H1 blockers) |
| Serotonin (5-HT) | YES | YES | Acts on 5-HT receptors and involved in inflammation and vasodilation | C-fiber receptors and trigeminal system | Stimulated by serotonin; inhibited by triptans (5-HT1B/1D agonists) |
| Bradykinin | YES | YES | Inflammatory mediator and increases vascular permeability | Vascular endothelium and peripheral nociceptors | Inhibited by bradykinin B2 receptor antagonists |
| Prostaglandins | YES | YES | Inflammatory mediators and stimulate permeability and vasodilation | Blood vessels and inflamed tissues | Inhibited by NSAIDs (cyclooxygenase inhibitors) |
| Leukotrienes | YES | YES | Like prostaglandins, involved in inflammation and extravasation | Blood vessels and inflamed tissues | Inhibited by leukotriene receptor antagonists |
| VIP | YES | NO | Acts on VPAC receptors and serves as cerebral vasodilator without inducing migraine | Cerebral vessels (VPAC receptors) | Stimulated by VIP; not affected by standard antimigraine agents |
| NPY | NO | NO | Vasoconstrictor and enhances noradrenaline effect | Sympathetic system and cerebral vessels | No specific inhibitor used clinically |
| Ketorolac | NO | YES | Inhibits cyclooxygenase, blocks extravasation, and does not inhibit CGRP | Dura mater and brainstem | NSAIDs (e.g., ketorolac) inhibit extravasation |
| Aspirin | NO | YES | Inhibits cyclooxygenase and blocks neurogenic extravasation | Dura mater and brainstem | NSAIDs (e.g., aspirin) inhibit extravasation |
| Valproate | NO | YES | Acts on GABA A receptors and inhibits dural extravasation and transmission | Dura mater and trigeminal ganglia | Inhibits neurogenic inflammation; no upstream stimulator |
| Opioids | NO | YES | Acts on μ-opioid receptors, inhibiting neurogenic plasma extravasation and vasodilation | Central and peripheral µ-opioid receptors | Inhibit µ-opioid receptors (e.g., morphine) |
| Sumatriptan | NO | YES | 5-HT1D agonist and blocks extravasation and mast cell degranulation | 5-HT1D receptors on C fibers and trigeminal ganglion | 5-HT1D agonist (e.g., sumatriptan) inhibits extravasation |
| PHM-29 | YES | NO | Potent vasodilator peptide in cerebral vessels | Cerebral vessels | No clinical modulators known |
| Acetylcholine | YES | NO | Neurotransmitter and induces vasodilation in cerebral vessels | Cerebral vessels | No clinical modulators known |
| Endothelin | NO | YES | Vasoconstrictive system involved in extravasation and lacks therapeutic efficacy | Cerebral vessels and dura mater | Inhibited by endothelin receptor antagonists (ineffective clinically) |
| Octreotide | NO | YES | Somatostatin analog that inhibits extravasation and neuronal activation | Brainstem and somatostatin receptors | Somatostatin analog (e.g., octreotide) |
| Somatostatin | NO | YES | Inhibits neurogenic inflammation and neuropeptide release | Central and peripheral nervous system | Somatostatin analogs; inhibitory G-protein coupled mechanisms |
| Category | Key Mechanisms/Concepts | Implications |
|---|---|---|
| Insulin Resistance (IR) | Impaired PI3K/AKT signaling Chronic inflammation (TNF-α, IL-6, CRP) Oxidative stress, mitochondrial dysfunction | Hyperglycemia, dyslipidemia Risk of T2DM and metabolic syndrome |
| β-Cell Dysfunction | ER stress → unfolded protein response (UPR) Cytokine inhibition (IL-1β, TNF-α via NF-κB) Mitochondrial impairment, ↓ GSIS | Reduced insulin synthesis/secretion Progression of diabetes |
| Adipose Tissue and Adipokines | CTRP3: ↑ insulin sensitivity (via AMPK) WISP1: ↓ insulin signaling Macrophage-driven inflammation | Worsening of IR and systemic inflammation |
| Neuroinflammation in IR | Brain insulin resistance affects microglia Cytokine-induced IR (IL-6, TNF-α) ↓ Amyloid-β clearance | Alzheimer’s disease risk cognitive decline, depression, and schizophrenia |
| CGRP and Neurogenic Inflammation | CGRP promotes vasodilation, immune cell activation, and mast cell degranulation | Target in migraine; diabetic complications (neuropathy, retinopathy) |
| Brain Insulin Signaling | InsR on neurons, astrocytes, microglia, and endothelium Modulates GABA, NMDA, and AMPA Affects dopamine (VTA), cognition, and BBB transport | Mood, motivation, memory, and reward regulation |
| Therapeutic Strategies | Natural agents: curcumin and pomegranate peel (↓ NF-κB and ↑ Nrf2) CR and exercise: AMPK/SIRT1, ↑ mitochondrial function Intranasal insulin | Improve systemic and brain insulin sensitivity; ↓ neuroinflammation; ↑ cognitive function |
| Intervention | BMI Reduction | Effect on Migraine | Metabolic Impact |
|---|---|---|---|
| Bariatric Surgery | −10.2 kg/m2 | Moderate to large improvement | Marked reduction in systemic inflammation, insulin resistance, and leptin levels |
| Behavioral Weight Loss (diet + exercise) | −4.1 kg/m2 | Moderate improvement | Improved energy balance and reduced inflammatory cytokines |
| Ketogenic Diet | −3.0 kg/m2 | Strongest migraine improvement | Increased ketone metabolism and mitochondrial efficiency |
| Low-Fat Vegan Diet | −1.3 kg/m2 | Weakest effect | Minimal metabolic impact on neuroinflammation and mitochondrial function |
| Aspect | Gazerani (2021) [127] | Gazerani (2023) [128] | Evers (2025) [129] |
|---|---|---|---|
| Diet as a Trigger | Identifies food triggers but emphasizes bidirectionality. | Re-evaluates traditional triggers, suggesting some may be prodromal symptoms. | Focuses on food as a modifiable factor via CGM-personalized guidance through DTx. |
| Migraine as a Metabolic Disorder | Explores mitochondrial dysfunction and energy metabolism. | Supports the ketogenic diet as a metabolic intervention. | Supports the role of glucose metabolism in migraine susceptibility and treatment response. |
| Dietary Interventions | Focuses on weight loss, the ketogenic diet, and omega-3. | Provides strong evidence for KD and DASH diets and personalized low-glycemic diets. | Demonstrates the significant efficacy of a personalized low-glycemic diet using DTx. |
| Gut Microbiota | Introduces the gut–brain axis as a key factor. | Reports mixed evidence on probiotics for migraine relief. | Not directly addressed, but suggests that dietary personalization may indirectly support microbiota balance. |
| Technological Advancements | Highlights the need for personalized interventions. | Advocates AI-based tracking and digital health tools. | Highlights the clinical utility of digital therapeutics and CGM-based tools for individualized dietary interventions. |
| Meal Frequency | Clinical Indications | Mechanism/Metabolic Effects |
|---|---|---|
| 3 main meals (breakfast, lunch, dinner) | General population, normal weight, good glycemic regulation | Maintains energy balance, physiologically stimulates insulin, prevents caloric excess |
| 3 main meals + 2 snacks | Diabetes, insulin resistance, fasting-induced headache, reactive hypoglycemia | Prevents glycemic spikes, improves satiety, reduces trigeminovascular activation |
| 5–6 small meals/day | Metabolic syndrome, insulin resistance, migraine with metabolic component | Reduces insulin stress, enhances insulin sensitivity, prevents cortical spreading depression (CSD) |
| 2 meals/day or intermittent fasting | Obesity, patients with high metabolic flexibility, but not suitable for migraine patients | Activates autophagy, improves insulin sensitivity, but may cause glycemic drops and migraines |
| Macronutrient | General Recommendations (EFSA, WHO, USDA) | For Insulin Resistance/Hyperinsulinemia (ADA; Evers (2025) [129]; García-Pérez-de-Sevilla (2025) [157]) | Metabolic Mechanism/Rationale |
|---|---|---|---|
| Carbohydrates | 45–60% Whole grains, fruits, vegetables, and legumes | 30–40% Low-GI vegetables, legumes, oats, and limited whole grains | Lowering postprandial glycemia and insulinemia and improving insulin sensitivity (Sacks et al., 2014 [170]; ADA 2023) |
| Fats (Total) | 20–35% Olive oil, nuts, seeds, and fatty fish | 35–40% Rich in MUFAs and PUFAs (e.g., olive oil, fish); limit saturated and trans fats | Improves lipid profile and enhances insulin action; omega-3 modulates TRPV1-CGRP axis [157] |
| Proteins | 10–20% Fish, legumes, dairy, lean meats, and eggs | 20–25% Lean proteins, legumes, eggs, and fish | Enhances satiety and thermogenesis, and reduces postprandial glucose excursions [129] |
| Fiber | 25–30 g/day Fruits, vegetables, legumes, and whole grains | 30–35 g/day Legumes, leafy greens, oats, and fruits with skin | Slows carbohydrate absorption, reduces glycemic load, and improves gut microbiota [130] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavestro, C. Metabolic Dysfunction and Dietary Interventions in Migraine Management: The Role of Insulin Resistance and Neuroinflammation—A Narrative and Scoping Review. Brain Sci. 2025, 15, 474. https://doi.org/10.3390/brainsci15050474
Cavestro C. Metabolic Dysfunction and Dietary Interventions in Migraine Management: The Role of Insulin Resistance and Neuroinflammation—A Narrative and Scoping Review. Brain Sciences. 2025; 15(5):474. https://doi.org/10.3390/brainsci15050474
Chicago/Turabian StyleCavestro, Cinzia. 2025. "Metabolic Dysfunction and Dietary Interventions in Migraine Management: The Role of Insulin Resistance and Neuroinflammation—A Narrative and Scoping Review" Brain Sciences 15, no. 5: 474. https://doi.org/10.3390/brainsci15050474
APA StyleCavestro, C. (2025). Metabolic Dysfunction and Dietary Interventions in Migraine Management: The Role of Insulin Resistance and Neuroinflammation—A Narrative and Scoping Review. Brain Sciences, 15(5), 474. https://doi.org/10.3390/brainsci15050474