
Neurophysiological Markers of Reward Processing Can Inform Preclinical Neurorehabilitation Approaches for Cognitive Impairments Following Brain Injury

{kind=link}
{kind=link}
Abstract
1. Introduction
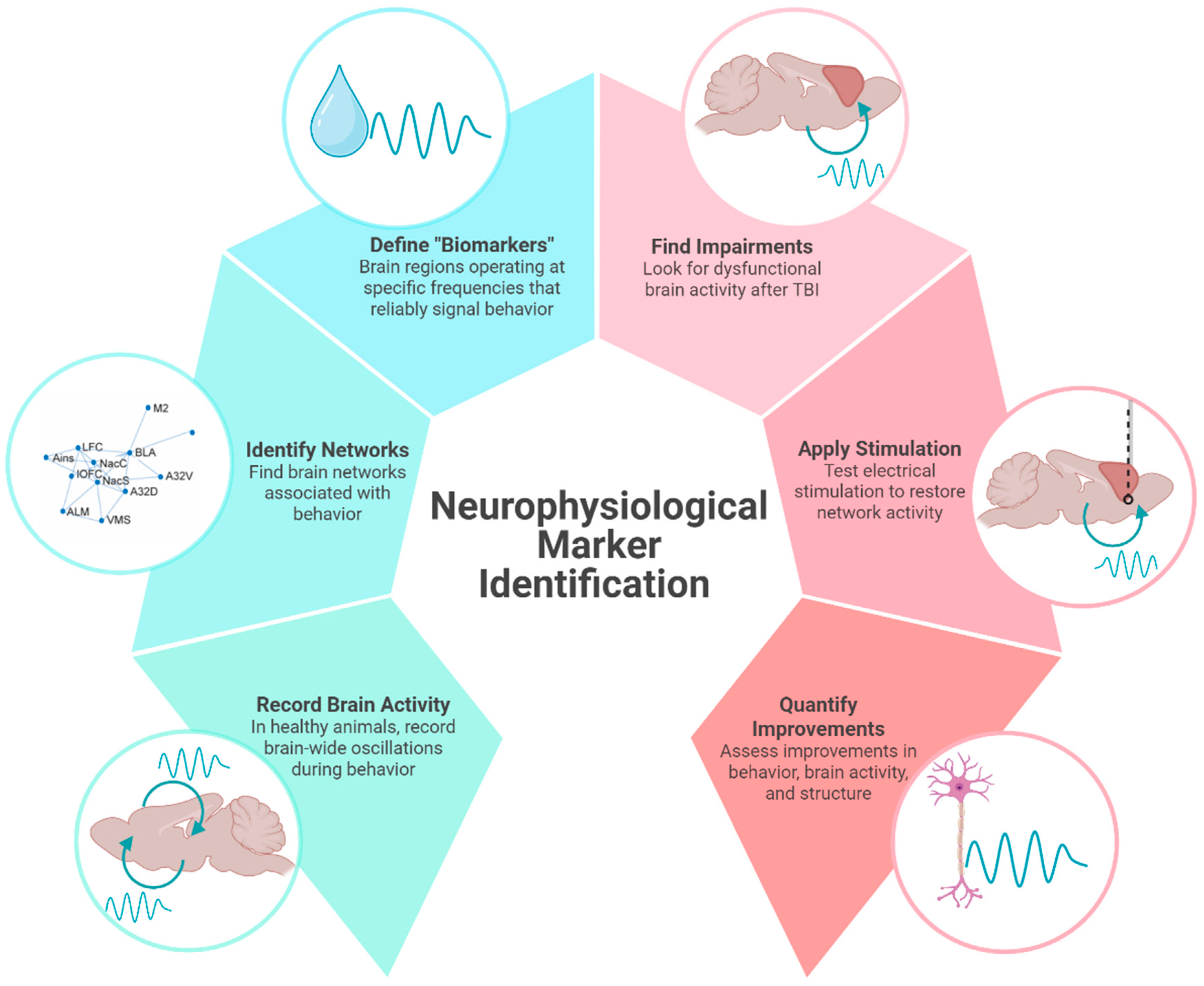
2. Neurophysiological Markers Can Identify Potential Therapeutic Targets
2.1. Brain Oscillations and Their Disruption After Injury
2.2. Neuromodulation to Restore Brain Activity
2.3. How Does Stimulation Restore Brain Function Following TBI?
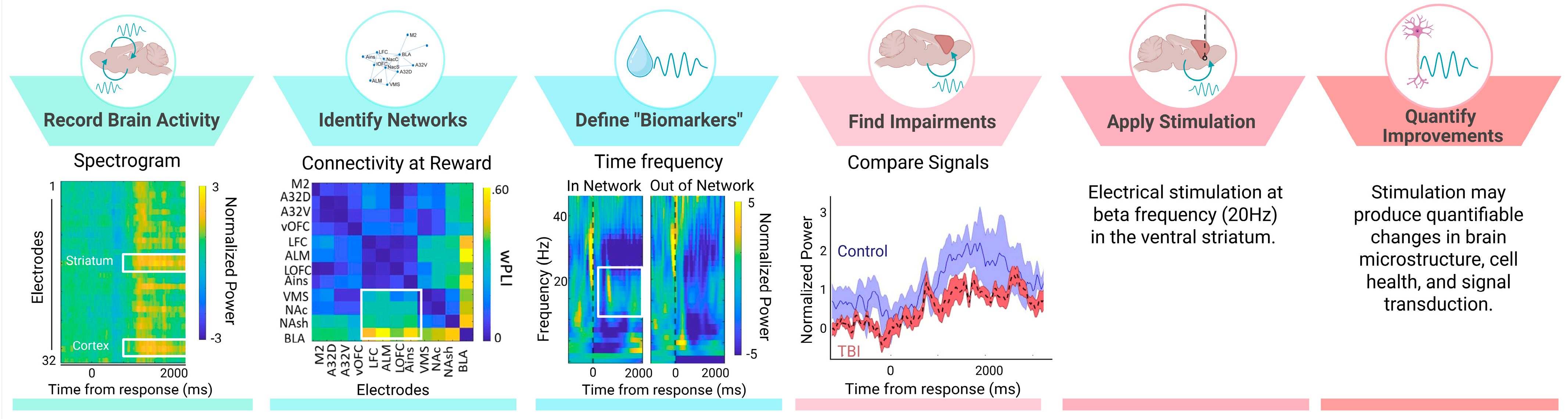
3. A Potential Biomarker of Reward-Processing Deficits in TBI
3.1. Preclinical Models of Frontal TBI
3.2. Deficient Cortical Beta Oscillations Indicate Reward-Processing Issues After TBI
3.3. Other Potential Neurophysiological Biomarkers
4. Limitations and Challenges (In Translation and Beyond)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wilde, E.A.; Wanner, I.-B.; Kenney, K.; Gill, J.; Stone, J.R.; Disner, S.; Schnakers, C.; Meyer, R.; Prager, E.M.; Haas, M.; et al. A Framework to Advance Biomarker Development in the Diagnosis, Outcome Prediction, and Treatment of Traumatic Brain Injury. J. Neurotrauma 2022, 39, 436–457. [Google Scholar] [CrossRef] [PubMed]
- Ghaith, H.S.; Nawar, A.A.; Gabra, M.D.; Abdelrahman, M.E.; Nafady, M.H.; Bahbah, E.I.; Ebada, M.A.; Ashraf, G.M.; Negida, A.; Barreto, G.E. A Literature Review of Traumatic Brain Injury Biomarkers. Mol. Neurobiol. 2022, 59, 4141–4158. [Google Scholar] [CrossRef] [PubMed]
- Gutierre, M.U.; Telles, J.P.M.; Welling, L.C.; Rabelo, N.N.; Teixeira, M.J.; Figueiredo, E.G. Biomarkers for Traumatic Brain Injury: A Short Review. Neurosurg. Rev. 2021, 44, 2091–2097. [Google Scholar] [CrossRef]
- Bigler, E.D. Neuroimaging Biomarkers in Mild Traumatic Brain Injury (mTBI). Neuropsychol. Rev. 2013, 23, 169–209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Safar, K.; Emami, Z.; Ibrahim, G.M.; Scratch, S.E.; Da Costa, L.; Dunkley, B.T. Local and Large-Scale Beta Oscillatory Dysfunction in Males with Mild Traumatic Brain Injury. J. Neurophysiol. 2020, 124, 1948–1958. [Google Scholar] [CrossRef]
- Freire, M.A.M.; Rocha, G.S.; Bittencourt, L.O.; Falcao, D.; Lima, R.R.; Cavalcanti, J.R.L.P. Cellular and Molecular Pathophysiology of Traumatic Brain Injury: What Have We Learned So Far? Biology 2023, 12, 1139. [Google Scholar] [CrossRef]
- Ladak, A.A.; Enam, S.A.; Ibrahim, M.T. A Review of the Molecular Mechanisms of Traumatic Brain Injury. World Neurosurg. 2019, 131, 126–132. [Google Scholar] [CrossRef]
- Risbrough, V.B.; Vaughn, M.N.; Friend, S.F. Role of Inflammation in Traumatic Brain Injury–Associated Risk for Neuropsychiatric Disorders: State of the Evidence and Where Do We Go From Here. Biol. Psychiatry 2022, 91, 438–448. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Huang, E.Y.-K.; Kuo, T.-T.; Ma, H.-I.; Hoffer, B.J.; Tsui, P.-F.; Tsai, J.-J.; Chou, Y.-C.; Chiang, Y.-H. Dopamine Release Impairment in Striatum after Different Levels of Cerebral Cortical Fluid Percussion Injury. Cell Transplant. 2015, 24, 2113–2128. [Google Scholar] [CrossRef]
- Chou, A.; Morganti, J.M.; Rosi, S. Frontal Lobe Contusion in Mice Chronically Impairs Prefrontal-Dependent Behavior. PLoS ONE 2016, 11, e0151418. [Google Scholar] [CrossRef]
- Polinder, S.; Cnossen, M.C.; Real, R.G.L.; Covic, A.; Gorbunova, A.; Voormolen, D.C.; Master, C.L.; Haagsma, J.A.; Diaz-Arrastia, R.; Von Steinbuechel, N. A Multidimensional Approach to Post-Concussion Symptoms in Mild Traumatic Brain Injury. Front. Neurol. 2018, 9, 1113. [Google Scholar] [CrossRef]
- Gosselin, N.; Bottari, C.; Chen, J.-K.; Huntgeburth, S.C.; De Beaumont, L.; Petrides, M.; Cheung, B.; Ptito, A. Evaluating the Cognitive Consequences of Mild Traumatic Brain Injury and Concussion by Using Electrophysiology. Neurosurg. Focus 2012, 33, E7. [Google Scholar] [CrossRef] [PubMed]
- Shaver, T.K.; Ozga, J.E.; Zhu, B.; Anderson, K.G.; Martens, K.M.; Vonder Haar, C. Long-Term Deficits in Risky Decision-Making after Traumatic Brain Injury on a Rat Analog of the Iowa Gambling Task. Brain Res. 2019, 1704, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, A.R.; Levin, H.S. Cognitive Sequelae of Traumatic Brain Injury. Psychiatr. Clin. N. Am. 2014, 37, 1–11. [Google Scholar] [CrossRef]
- Rutherford, W.H.; Merrett, J.D.; McDonald, J.R. Symptoms at One Year Following Concussion from Minor Head Injuries. Injury 1979, 10, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Kundu, B.; Brock, A.A.; Englot, D.J.; Butson, C.R.; Rolston, J.D. Deep Brain Stimulation for the Treatment of Disorders of Consciousness and Cognition in Traumatic Brain Injury Patients: A Review. Neurosurg. Focus 2018, 45, E14. [Google Scholar] [CrossRef]
- Kraus, M.F.; Susmaras, T.; Caughlin, B.P.; Walker, C.J.; Sweeney, J.A.; Little, D.M. White Matter Integrity and Cognition in Chronic Traumatic Brain Injury: A Diffusion Tensor Imaging Study. Brain 2007, 130, 2508–2519. [Google Scholar] [CrossRef]
- Smith, D.H.; Meaney, D.F. Axonal Damage in Traumatic Brain Injury. Neuroscientist 2000, 6, 483–495. [Google Scholar] [CrossRef]
- Stephens, J.A.; Williamson, K.-N.C.; Berryhill, M.E. Cognitive Rehabilitation After Traumatic Brain Injury: A Reference for Occupational Therapists. OTJR Occup. Ther. J. Res. 2015, 35, 5–22. [Google Scholar] [CrossRef]
- Yin, B.; Li, D.-D.; Huang, H.; Gu, C.-H.; Bai, G.-H.; Hu, L.-X.; Zhuang, J.-F.; Zhang, M. Longitudinal Changes in Diffusion Tensor Imaging Following Mild Traumatic Brain Injury and Correlation With Outcome. Front. Neural Circuits 2019, 13, 28. [Google Scholar] [CrossRef]
- Dockree, P.M.; Robertson, I.H. Electrophysiological Markers of Cognitive Deficits in Traumatic Brain Injury: A Review. Int. J. Psychophysiol. 2011, 82, 53–60. [Google Scholar] [CrossRef] [PubMed]
- McNerney, M.W.; Gurkoff, G.G.; Beard, C.; Berryhill, M.E. The Rehabilitation Potential of Neurostimulation for Mild Traumatic Brain Injury in Animal and Human Studies. Brain Sci. 2023, 13, 1402. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, F.; Wee, I.C.; Collins-Praino, L.E. Chronic Motor Performance Following Different Traumatic Brain Injury Severity-A Systematic Review. Front. Neurol. 2023, 14, 1180353. [Google Scholar] [CrossRef]
- Hornby, T.G.; Reisman, D.S.; Ward, I.G.; Scheets, P.L.; Miller, A.; Haddad, D.; Fox, E.J.; Fritz, N.E.; Hawkins, K.; Henderson, C.E.; et al. Clinical Practice Guideline to Improve Locomotor Function Following Chronic Stroke, Incomplete Spinal Cord Injury, and Brain Injury. J. Neurol. Phys. Ther. JNPT 2020, 44, 49–100. [Google Scholar] [CrossRef] [PubMed]
- Tani, J.; Wen, Y.-T.; Hu, C.-J.; Sung, J.-Y. Current and Potential Pharmacologic Therapies for Traumatic Brain Injury. Pharmaceuticals 2022, 15, 838. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.B.; Jain, S.; Giacino, J.T.; Levin, H.; Dikmen, S.; Nelson, L.D.; Vassar, M.J.; Okonkwo, D.O.; Diaz-Arrastia, R.; Robertson, C.S.; et al. Risk of Posttraumatic Stress Disorder and Major Depression in Civilian Patients After Mild Traumatic Brain Injury: A TRACK-TBI Study. JAMA Psychiatry 2019, 76, 249–258. [Google Scholar] [CrossRef]
- Barone, V.; De Koning, M.E.; Van Der Horn, H.J.; Van Der Naalt, J.; Eertman-Meyer, C.J.; Van Putten, M.J.A.M. Neurophysiological Signatures of Mild Traumatic Brain Injury in the Acute and Subacute Phase. Neurol. Sci. 2024, 45, 3313–3323. [Google Scholar] [CrossRef]
- Buzsáki, G.; Anastassiou, C.A.; Koch, C. The Origin of Extracellular Fields and Currents--EEG, ECoG, LFP and Spikes. Nat. Rev. Neurosci. 2012, 13, 407–420. [Google Scholar] [CrossRef]
- Masimore, B.; Kakalios, J.; Redish, A.D. Measuring Fundamental Frequencies in Local Field Potentials. J. Neurosci. Methods 2004, 138, 97–105. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Rieger, R.E.; Wilson, J.K.; Gill, D.; Fullerton, L.; Brandt, E.; Mayer, A.R. Joint Analysis of Frontal Theta Synchrony and White Matter Following Mild Traumatic Brain Injury. Brain Imaging Behav. 2020, 14, 2210–2223. [Google Scholar] [CrossRef]
- Dong, X.; Ye, W.; Tang, Y.; Wang, J.; Zhong, L.; Xiong, J.; Liu, H.; Lu, G.; Feng, Z. Wakefulness-Promoting Effects of Lateral Hypothalamic Area-Deep Brain Stimulation in Traumatic Brain Injury-Induced Comatose Rats: Upregulation of A1-Adrenoceptor Subtypes and Downregulation of Gamma-Aminobutyric Acid β Receptor Expression Via the Orexins Pathway. World Neurosurg. 2021, 152, e321–e331. [Google Scholar] [CrossRef] [PubMed]
- Antón Álvarez, X.; Sampedro, C.; Pérez, P.; Laredo, M.; Couceiro, V.; Hernández, Á.; Figueroa, J.; Varela, M.; Arias, D.; Corzo, L.; et al. Positive Effects of Cerebrolysin on Electroencephalogram Slowing, Cognition and Clinical Outcome in Patients with Postacute Traumatic Brain Injury: An Exploratory Study. Int. Clin. Psychopharmacol. 2003, 18, 271–278. [Google Scholar] [CrossRef]
- Sherman, M.T.; Kanai, R.; Seth, A.K.; VanRullen, R. Rhythmic Influence of Top–Down Perceptual Priors in the Phase of Prestimulus Occipital Alpha Oscillations. J. Cogn. Neurosci. 2016, 28, 1318–1330. [Google Scholar] [CrossRef]
- Xiong, Y.; Fries, P.; Bastos, A.M. Which Rhythms Reflect Bottom-Up and Top-Down Processing? In Intracranial EEG; Axmacher, N., Ed.; Studies in Neuroscience, Psychology and Behavioral Economics; Springer International Publishing: Cham, Switzerland, 2023; pp. 389–414. ISBN 978-3-031-20909-3. [Google Scholar]
- Surendrakumar, S.; Rabelo, T.K.; Campos, A.C.P.; Mollica, A.; Abrahao, A.; Lipsman, N.; Burke, M.J.; Hamani, C. Neuromodulation Therapies in Pre-Clinical Models of Traumatic Brain Injury: Systematic Review and Translational Applications. J. Neurotrauma 2023, 40, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Pevzner, A.; Izadi, A.; Lee, D.J.; Shahlaie, K.; Gurkoff, G.G. Making Waves in the Brain: What Are Oscillations, and Why Modulating Them Makes Sense for Brain Injury. Front. Syst. Neurosci. 2016, 10, 30. [Google Scholar] [CrossRef]
- Slobounov, S.; Sebastianelli, W.; Hallett, M. Residual Brain Dysfunction Observed One Year Post-Mild Traumatic Brain Injury: Combined EEG and Balance Study. Clin. Neurophysiol. 2012, 123, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, R.W.; Walker, R.A.; Gerson, I.; Geisler, F.H. EEG Discriminant Analyses of Mild Head Trauma. Electroencephalogr. Clin. Neurophysiol. 1989, 73, 94–106. [Google Scholar] [CrossRef]
- Demirtas-Tatlidede, A.; Vahabzadeh-Hagh, A.M.; Bernabeu, M.; Tormos, J.M.; Pascual-Leone, A. Noninvasive Brain Stimulation in Traumatic Brain Injury. J. Head Trauma Rehabil. 2012, 27, 274–292. [Google Scholar] [CrossRef]
- Tanaka, Y.; Matsuwaki, T.; Yamanouchi, K.; Nishihara, M. Exacerbated Inflammatory Responses Related to Activated Microglia after Traumatic Brain Injury in Progranulin-Deficient Mice. Neuroscience 2013, 231, 49–60. [Google Scholar] [CrossRef]
- Bundy, D.T.; Nudo, R.J. Preclinical Studies of Neuroplasticity Following Experimental Brain Injury: An Update. Stroke 2019, 50, 2626–2633. [Google Scholar] [CrossRef]
- Ghasemi, P.; Sahraee, T.; Mohammadi, A. Closed- and Open-Loop Deep Brain Stimulation: Methods, Challenges, Current and Future Aspects. J. Biomed. Phys. Eng. 2018, 8, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Calderone, A.; Cardile, D.; Gangemi, A.; De Luca, R.; Quartarone, A.; Corallo, F.; Calabrò, R.S. Traumatic Brain Injury and Neuromodulation Techniques in Rehabilitation: A Scoping Review. Biomedicines 2024, 12, 438. [Google Scholar] [CrossRef] [PubMed]
- Ziesel, D.; Nowakowska, M.; Scheruebel, S.; Kornmueller, K.; Schäfer, U.; Schindl, R.; Baumgartner, C.; Üçal, M.; Rienmüller, T. Electrical Stimulation Methods and Protocols for the Treatment of Traumatic Brain Injury: A Critical Review of Preclinical Research. J. Neuroeng. Rehabil. 2023, 20, 51. [Google Scholar] [CrossRef] [PubMed]
- Perera, T.; George, M.S.; Grammer, G.; Janicak, P.G.; Pascual-Leone, A.; Wirecki, T.S. The Clinical TMS Society Consensus Review and Treatment Recommendations for TMS Therapy for Major Depressive Disorder. Brain Stimul. 2016, 9, 336–346. [Google Scholar] [CrossRef]
- Pink, A.E.; Williams, C.; Alderman, N.; Stoffels, M. The Use of Repetitive Transcranial Magnetic Stimulation (rTMS) Following Traumatic Brain Injury (TBI): A Scoping Review. Neuropsychol. Rehabil. 2021, 31, 479–505. [Google Scholar] [CrossRef]
- Klomjai, W.; Katz, R.; Lackmy-Vallée, A. Basic Principles of Transcranial Magnetic Stimulation (TMS) and Repetitive TMS (rTMS). Ann. Phys. Rehabil. Med. 2015, 58, 208–213. [Google Scholar] [CrossRef]
- Vahabzadeh-Hagh, A.M.; Muller, P.A.; Gersner, R.; Zangen, A.; Rotenberg, A. Translational Neuromodulation: Approximating Human Transcranial Magnetic Stimulation Protocols in Rats. Neuromodulation Technol. Neural Interface 2012, 15, 296–305. [Google Scholar] [CrossRef]
- Boonzaier, J.; Petrov, P.I.; Otte, W.M.; Smirnov, N.; Neggers, S.F.W.; Dijkhuizen, R.M. Design and Evaluation of a Rodent-Specific Transcranial Magnetic Stimulation Coil: An In Silico and In Vivo Validation Study. Neuromodulation 2020, 23, 324–334. [Google Scholar] [CrossRef]
- Moshe, H.; Gal, R.; Barnea-Ygael, N.; Gulevsky, T.; Alyagon, U.; Zangen, A. Prelimbic Stimulation Ameliorates Depressive-Like Behaviors and Increases Regional BDNF Expression in a Novel Drug-Resistant Animal Model of Depression. Brain Stimul. 2016, 9, 243–250. [Google Scholar] [CrossRef]
- Collins-Praino, L.E. Traumatic Axonal Injury as a Key Driver of the Relationship between Traumatic Brain Injury, Cognitive Dysfunction, and Dementia. In Cellular, Molecular, Physiological, and Behavioral Aspects of Traumatic Brain Injury; Elsevier: Amsterdam, The Netherlands, 2022; pp. 475–486. ISBN 978-0-12-823036-7. [Google Scholar]
- Smith, D.H.; Kochanek, P.M.; Rosi, S.; Meyer, R.; Ferland-Beckham, C.; Prager, E.M.; Ahlers, S.T.; Crawford, F. Roadmap for Advancing Pre-Clinical Science in Traumatic Brain Injury. J. Neurotrauma 2021, 38, 3204–3221. [Google Scholar] [CrossRef]
- Xu, H.; Wang, Z.; Li, J.; Wu, H.; Peng, Y.; Fan, L.; Chen, J.; Gu, C.; Yan, F.; Wang, L.; et al. The Polarization States of Microglia in TBI: A New Paradigm for Pharmacological Intervention. Neural Plast. 2017, 2017, 5405104. [Google Scholar] [CrossRef] [PubMed]
- Johnson, V.E.; Stewart, J.E.; Begbie, F.D.; Trojanowski, J.Q.; Smith, D.H.; Stewart, W. Inflammation and White Matter Degeneration Persist for Years after a Single Traumatic Brain Injury. Brain 2013, 136, 28–42. [Google Scholar] [CrossRef]
- Maas, A.I.R.; Menon, D.K.; Manley, G.T.; Abrams, M.; Åkerlund, C.; Andelic, N.; Aries, M.; Bashford, T.; Bell, M.J.; Bodien, Y.G.; et al. Traumatic Brain Injury: Progress and Challenges in Prevention, Clinical Care, and Research. Lancet Neurol. 2022, 21, 1004–1060. [Google Scholar] [CrossRef] [PubMed]
- Carballosa Gonzalez, M.M.; Blaya, M.O.; Alonso, O.F.; Bramlett, H.M.; Hentall, I.D. Midbrain Raphe Stimulation Improves Behavioral and Anatomical Recovery from Fluid-Percussion Brain Injury. J. Neurotrauma 2013, 30, 119–130. [Google Scholar] [CrossRef]
- Yoon, Y.-S.; Cho, K.H.; Kim, E.-S.; Lee, M.-S.; Lee, K.J. Effect of Epidural Electrical Stimulation and Repetitive Transcranial Magnetic Stimulation in Rats With Diffuse Traumatic Brain Injury. Ann. Rehabil. Med. 2015, 39, 416–424. [Google Scholar] [CrossRef]
- Yu, K.P.; Yoon, Y.-S.; Lee, J.G.; Oh, J.S.; Lee, J.-S.; Seog, T.; Lee, H.-Y. Effects of Electric Cortical Stimulation (ECS) and Transcranial Direct Current Stimulation (tDCS) on Rats With a Traumatic Brain Injury. Ann. Rehabil. Med. 2018, 42, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Zhang, Y.; Miranzadeh Mahabadi, H.; Parvizi, A.; Taghibiglou, C. Low-Field Magnetic Stimulation Restores Cognitive and Motor Functions in the Mouse Model of Repeated Traumatic Brain Injury: Role of Cellular Prion Protein. J. Neurotrauma 2019, 36, 3103–3114. [Google Scholar] [CrossRef]
- Chiken, S.; Nambu, A. Mechanism of Deep Brain Stimulation: Inhibition, Excitation, or Disruption? Neuroscientist 2016, 22, 313–322. [Google Scholar] [CrossRef]
- Xu, N.; LaGrow, T.J.; Anumba, N.; Lee, A.; Zhang, X.; Yousefi, B.; Bassil, Y.; Clavijo, G.P.; Khalilzad Sharghi, V.; Maltbie, E.; et al. Functional Connectivity of the Brain Across Rodents and Humans. Front. Neurosci. 2022, 16, 816331. [Google Scholar] [CrossRef]
- Ma, X.; Aravind, A.; Pfister, B.J.; Chandra, N.; Haorah, J. Animal Models of Traumatic Brain Injury and Assessment of Injury Severity. Mol. Neurobiol. 2019, 56, 5332–5345. [Google Scholar] [CrossRef]
- Vonder Haar, C.; Lam, F.C.W.; Adams, W.K.; Riparip, L.-K.; Kaur, S.; Muthukrishna, M.; Rosi, S.; Winstanley, C.A. Frontal Traumatic Brain Injury in Rats Causes Long-Lasting Impairments in Impulse Control That Are Differentially Sensitive to Pharmacotherapeutics and Associated with Chronic Neuroinflammation. ACS Chem. Neurosci. 2016, 7, 1531–1542. [Google Scholar] [CrossRef] [PubMed]
- Francoeur, M.J.; Tang, T.; Fakhraei, L.; Wu, X.; Hulyalkar, S.; Cramer, J.; Buscher, N.; Ramanathan, D.R. Chronic, Multi-Site Recordings Supported by Two Low-Cost, Stationary Probe Designs Optimized to Capture Either Single Unit or Local Field Potential Activity in Behaving Rats. Front. Psychiatry 2021, 12, 678103. [Google Scholar] [CrossRef]
- Fakhraei, L.; Francoeur, M.; Balasubramani, P.; Tang, T.; Hulyalkar, S.; Buscher, N.; Claros, C.; Terry, A.; Gupta, A.; Xiong, H.; et al. Mapping Large-Scale Networks Associated with Action, Behavioral Inhibition and Impulsivity. eNeuro 2021, 8, ENEURO.0406-20.2021. [Google Scholar] [CrossRef] [PubMed]
- Fakhraei, L.; Francoeur, M.; Balasubramani, P.P.; Tang, T.; Hulyalkar, S.; Buscher, N.; Mishra, J.; Ramanathan, D.S. Electrophysiological Correlates of Rodent Default-Mode Network Suppression Revealed by Large-Scale Local Field Potential Recordings. Cereb. Cortex Commun. 2021, 2, tgab034. [Google Scholar] [CrossRef] [PubMed]
- Paterno, R.; Folweiler, K.A.; Cohen, A.S. Pathophysiology and Treatment of Memory Dysfunction After Traumatic Brain Injury. Curr. Neurol. Neurosci. Rep. 2017, 17, 52. [Google Scholar] [CrossRef]
- VanSolkema, M.; McCann, C.; Barker-Collo, S.; Foster, A. Attention and Communication Following TBI: Making the Connection through a Meta-Narrative Systematic Review. Neuropsychol. Rev. 2020, 30, 345–361. [Google Scholar] [CrossRef]
- Kim, N.; Jamison, K.; Jaywant, A.; Garetti, J.; Blunt, E.; RoyChoudhury, A.; Butler, T.; Dams-O’Connor, K.; Khedr, S.; Chen, C.-C.; et al. Comparisons of Electrophysiological Markers of Impaired Executive Attention after Traumatic Brain Injury and in Healthy Aging. NeuroImage 2023, 274, 120126. [Google Scholar] [CrossRef]
- De Simoni, S.; Jenkins, P.O.; Bourke, N.J.; Fleminger, J.J.; Hellyer, P.J.; Jolly, A.E.; Patel, M.C.; Cole, J.H.; Leech, R.; Sharp, D.J. Altered Caudate Connectivity Is Associated with Executive Dysfunction after Traumatic Brain Injury. Brain 2018, 141, 148–164. [Google Scholar] [CrossRef]
- Schmidt, R.; Herrojo Ruiz, M.; Kilavik, B.E.; Lundqvist, M.; Starr, P.A.; Aron, A.R. Beta Oscillations in Working Memory, Executive Control of Movement and Thought, and Sensorimotor Function. J. Neurosci. 2019, 39, 8231–8238. [Google Scholar] [CrossRef]
- Spitzer, B.; Haegens, S. Beyond the Status Quo: A Role for Beta Oscillations in Endogenous Content (Re)Activation. eNeuro 2017, 4, ENEURO.0170-17.2017. [Google Scholar] [CrossRef]
- Dunkley, B.T.; Freeman, T.C.A.; Muthukumaraswamy, S.D.; Singh, K.D. Cortical Oscillatory Changes in Human Middle Temporal Cortex Underlying Smooth Pursuit Eye Movements. Hum. Brain Mapp. 2013, 34, 837–851. [Google Scholar] [CrossRef] [PubMed]
- Little, S.; Bonaiuto, J.; Barnes, G.; Bestmann, S. Human Motor Cortical Beta Bursts Relate to Movement Planning and Response Errors. PLOS Biol. 2019, 17, e3000479. [Google Scholar] [CrossRef]
- Hoy, C.W.; De Hemptinne, C.; Wang, S.S.; Harmer, C.J.; Apps, M.A.J.; Husain, M.; Starr, P.A.; Little, S. Beta and Theta Oscillations Track Effort and Previous Reward in Human Basal Ganglia and Prefrontal Cortex during Decision Making. Proc. Natl. Acad. Sci. USA 2024, 121, e2322869121. [Google Scholar] [CrossRef]
- Hunt, A.W.; Mah, K.; Reed, N.; Engel, L.; Keightley, M. Oculomotor-Based Vision Assessment in Mild Traumatic Brain Injury: A Systematic Review. J. Head Trauma Rehabil. 2016, 31, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Koloski, M.F.; Hulyalkar, S.; Barnes, S.A.; Mishra, J.; Ramanathan, D.S. Cortico-Striatal Beta Oscillations as a Reward-Related Signal. Cogn. Affect. Behav. Neurosci. 2024, 24, 839–859. [Google Scholar] [CrossRef] [PubMed]
- HajiHosseini, A.; Holroyd, C.B. Sensitivity of Frontal Beta Oscillations to Reward Valence but Not Probability. Neurosci. Lett. 2015, 602, 99–103. [Google Scholar] [CrossRef]
- Marco-Pallarés, J.; Münte, T.F.; Rodríguez-Fornells, A. The Role of High-Frequency Oscillatory Activity in Reward Processing and Learning. Neurosci. Biobehav. Rev. 2015, 49, 1–7. [Google Scholar] [CrossRef]
- Mas-Herrero, E.; Ripollés, P.; HajiHosseini, A.; Rodríguez-Fornells, A.; Marco-Pallarés, J. Beta Oscillations and Reward Processing: Coupling Oscillatory Activity and Hemodynamic Responses. NeuroImage 2015, 119, 13–19. [Google Scholar] [CrossRef]
- D’Cruz, A.-M.; Ragozzino, M.E.; Mosconi, M.W.; Pavuluri, M.N.; Sweeney, J.A. Human Reversal Learning under Conditions of Certain versus Uncertain Outcomes. NeuroImage 2011, 56, 315–322. [Google Scholar] [CrossRef]
- Koloski, M.F.; O’Hearn, C.M.; Frankot, M.; Giesler, L.P.; Ramanathan, D.S.; Vonder Haar, C. Behavioral Interventions Can Improve Brain Injury-Induced Deficits in Behavioral Flexibility and Impulsivity Linked to Impaired Reward-Feedback Beta Oscillations. J. Neurotrauma 2024, 41, e1721–e1737. [Google Scholar] [CrossRef]
- Lee, D.J.; Gurkoff, G.G.; Izadi, A.; Seidl, S.E.; Echeverri, A.; Melnik, M.; Berman, R.F.; Ekstrom, A.D.; Muizelaar, J.P.; Lyeth, B.G.; et al. Septohippocampal Neuromodulation Improves Cognition after Traumatic Brain Injury. J. Neurotrauma 2015, 32, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Yaple, Z.; Martinez-Saito, M.; Novikov, N.; Altukhov, D.; Shestakova, A.; Klucharev, V. Power of Feedback-Induced Beta Oscillations Reflect Omission of Rewards: Evidence From an EEG Gambling Study. Front. Neurosci. 2018, 12, 776. [Google Scholar] [CrossRef]
- Hyman, J.M.; Wyble, B.P.; Goyal, V.; Rossi, C.A.; Hasselmo, M.E. Stimulation in Hippocampal Region CA1 in Behaving Rats Yields Long-Term Potentiation When Delivered to the Peak of Theta and Long-Term Depression When Delivered to the Trough. J. Neurosci. 2003, 23, 11725–11731. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Gurkoff, G.G.; Izadi, A.; Berman, R.F.; Ekstrom, A.D.; Muizelaar, J.P.; Lyeth, B.G.; Shahlaie, K. Medial Septal Nucleus Theta Frequency Deep Brain Stimulation Improves Spatial Working Memory after Traumatic Brain Injury. J. Neurotrauma 2013, 30, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Izadi, A.; Melnik, M.; Seidl, S.; Echeverri, A.; Shahlaie, K.; Gurkoff, G.G. Stimulation of the Medial Septum Improves Performance in Spatial Learning Following Pilocarpine-Induced Status Epilepticus. Epilepsy Res. 2017, 130, 53–63. [Google Scholar] [CrossRef]
- Yang, X.; Li, X.; Yuan, Y.; Sun, T.; Yang, J.; Deng, B.; Yu, H.; Gao, A.; Guan, J. 40 Hz Blue LED Relieves the Gamma Oscillations Changes Caused by Traumatic Brain Injury in Rat. Front. Neurol. 2022, 13, 882991. [Google Scholar] [CrossRef]
- Lundqvist, M.; Miller, E.K.; Nordmark, J.; Liljefors, J.; Herman, P. Beta: Bursts of Cognition. Trends Cogn. Sci. 2024, 28, 662–676. [Google Scholar] [CrossRef]
- Zaninotto, A.L.; El-Hagrassy, M.M.; Green, J.R.; Babo, M.; Paglioni, V.M.; Benute, G.G.; Paiva, W.S. Transcranial Direct Current Stimulation (tDCS) Effects on Traumatic Brain Injury (TBI) Recovery: A Systematic Review. Dement. Neuropsychol. 2019, 13, 172–179. [Google Scholar] [CrossRef]
- Leśniak, M.; Polanowska, K.; Seniów, J.; Członkowska, A. Effects of Repeated Anodal tDCS Coupled With Cognitive Training for Patients With Severe Traumatic Brain Injury: A Pilot Randomized Controlled Trial. J. Head Trauma Rehabil. 2014, 29, E20–E29. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koloski, M.F.; Menon, R.; Krasnyanskiy, V. Neurophysiological Markers of Reward Processing Can Inform Preclinical Neurorehabilitation Approaches for Cognitive Impairments Following Brain Injury. Brain Sci. 2025, 15, 471. https://doi.org/10.3390/brainsci15050471
Koloski MF, Menon R, Krasnyanskiy V. Neurophysiological Markers of Reward Processing Can Inform Preclinical Neurorehabilitation Approaches for Cognitive Impairments Following Brain Injury. Brain Sciences. 2025; 15(5):471. https://doi.org/10.3390/brainsci15050471
Chicago/Turabian StyleKoloski, Miranda Francoeur, Reyana Menon, and Victoria Krasnyanskiy. 2025. "Neurophysiological Markers of Reward Processing Can Inform Preclinical Neurorehabilitation Approaches for Cognitive Impairments Following Brain Injury" Brain Sciences 15, no. 5: 471. https://doi.org/10.3390/brainsci15050471
APA StyleKoloski, M. F., Menon, R., & Krasnyanskiy, V. (2025). Neurophysiological Markers of Reward Processing Can Inform Preclinical Neurorehabilitation Approaches for Cognitive Impairments Following Brain Injury. Brain Sciences, 15(5), 471. https://doi.org/10.3390/brainsci15050471