
Timing of Deep and REM Sleep Based on Fitbit Sleep Staging in Young Healthy Adults under Real-Life Conditions


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics, Study Design and Sample
2.2. Data Analyses
- The midpoints of sleep in minutes after sleep onset and clock time, reflecting the phase of sleep regarding the homeostatic and the circadian component, respectively.
- The sleep stage percentiles, indicating at what times 25%, 50%, 75% and 100% of the respective sleep stage was reached. The sleep stage percentiles were also determined in relation to minutes after sleep onset and clock time, reflecting the phases regarding the homeostatic and circadian component, respectively.
- To investigate the strength of intraindividual phase relationships between total sleep and the sleep stages, bivariate correlations between the midpoint of sleep and the sleep stage percentiles were computed for Spearman’s rank coefficients with a 95% confidence interval. The resulting intraindividual correlation coefficients (ICC) were used as a measure of the strengths of the phase relationship. The larger the ICC, the stronger the phase relationship.
- Sleep duration.
- Proportion of REM sleep in percentage of total sleep.
- Fragmentation index of REM sleep (RFI) as the number of transitions from REM sleep to any other sleep stage per hour of REM sleep.
3. Results
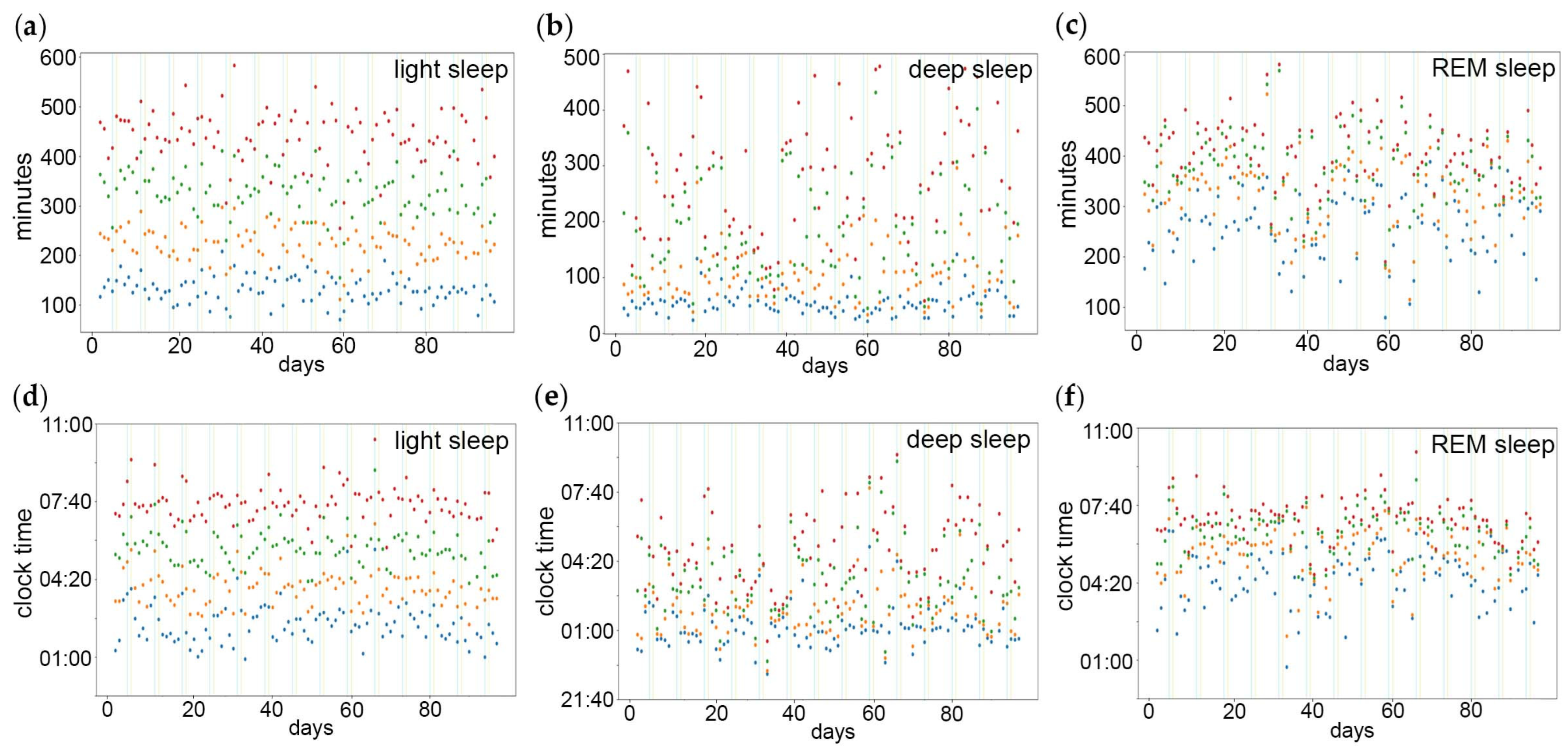
3.1. Within-Subject Sleep Data Analyses
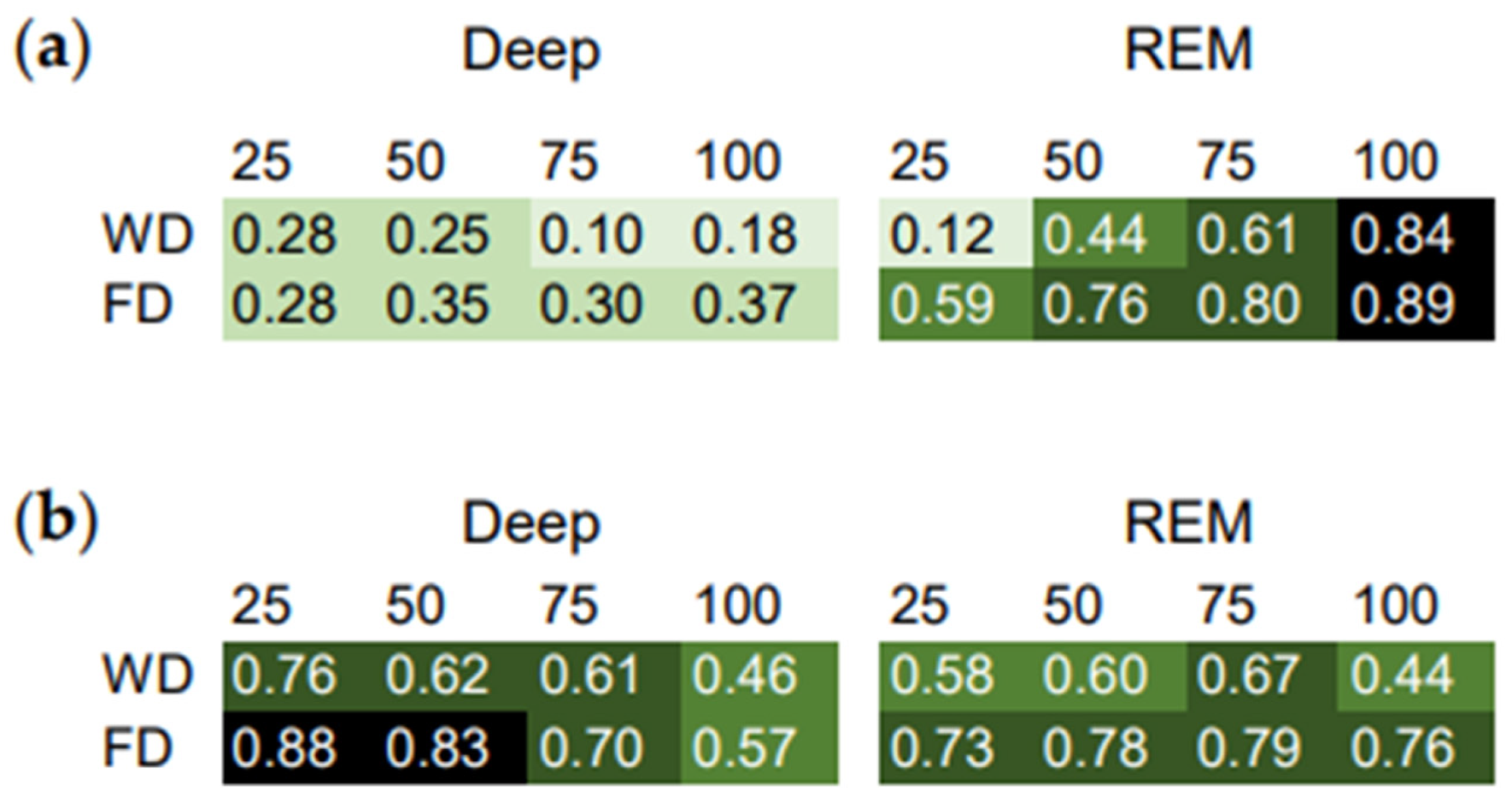
3.1.1. Sleep Stage Percentiles
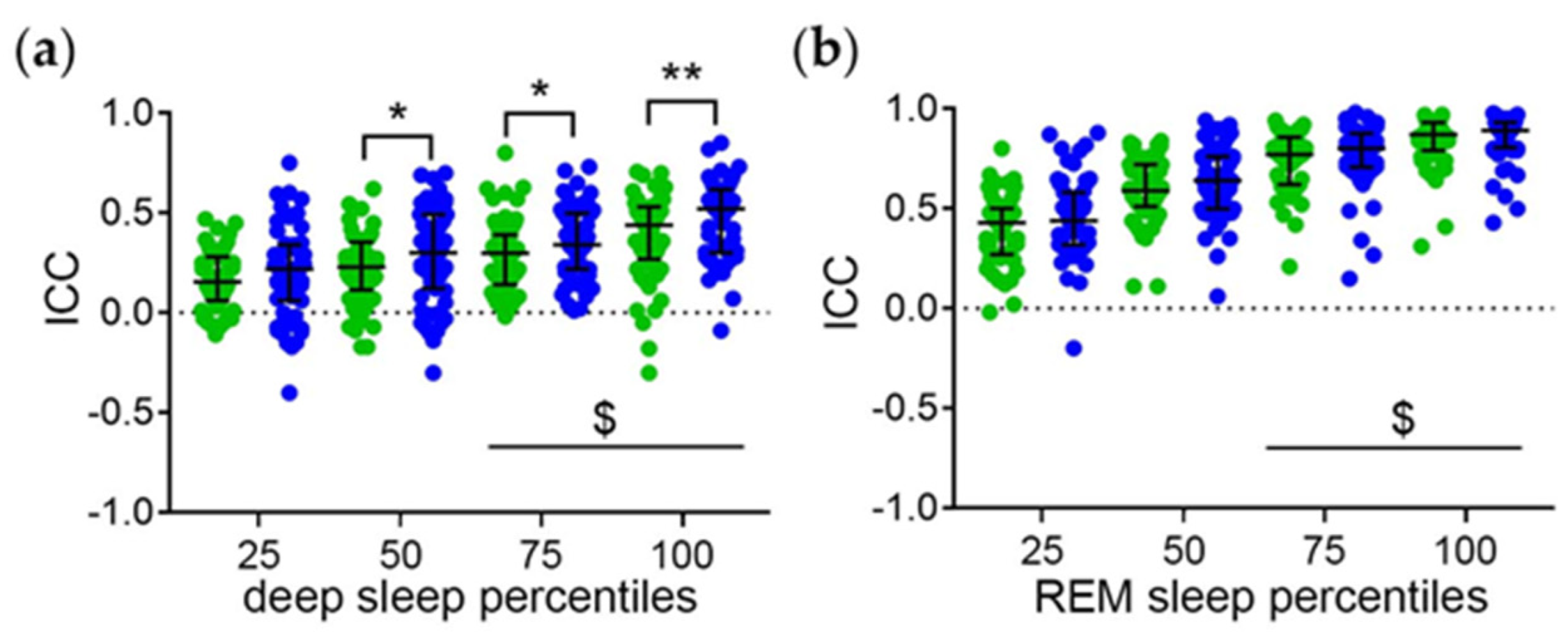
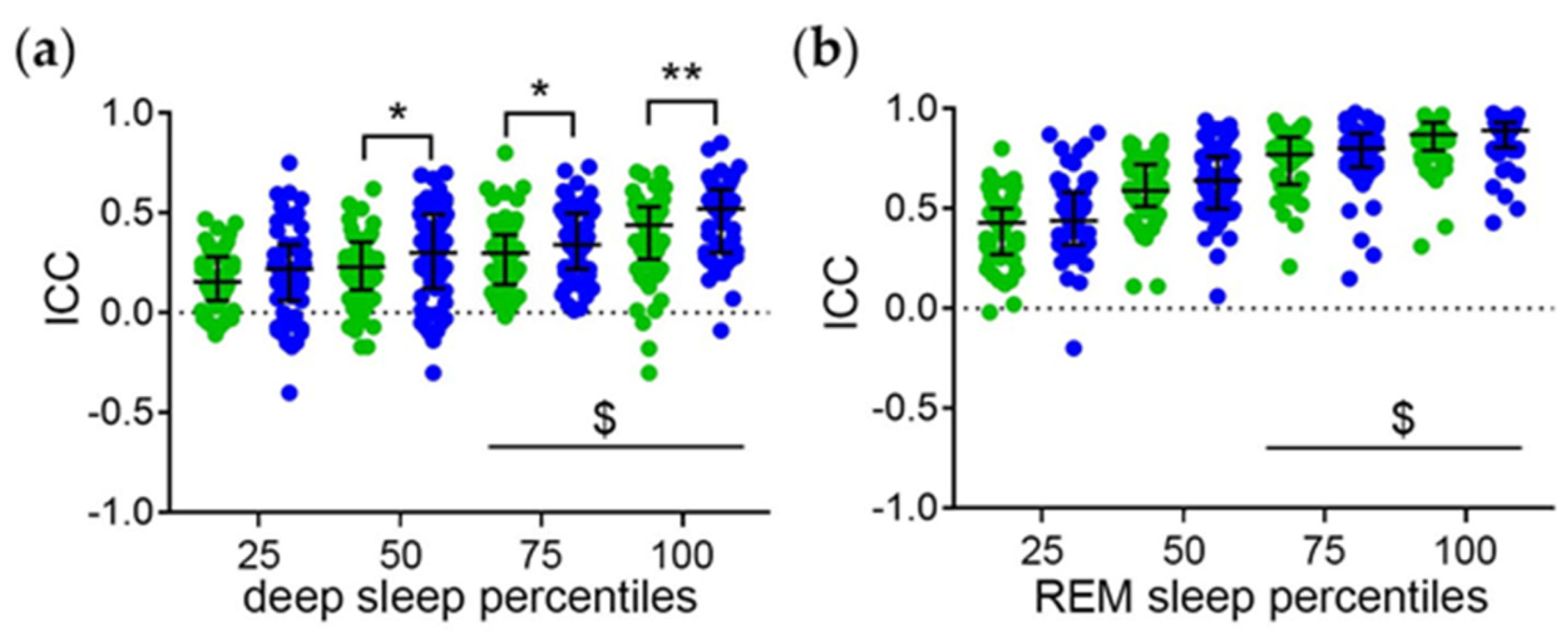
3.1.2. Intraindividual Correlation Coefficients (ICC) of the Sleep Phase Percentiles with the Midpoint of Sleep as a Measure of the Strength of the Phase Relationships
3.2. Between-Subject Sleep Data Analyses
3.2.1. Distribution of the ICC
3.2.2. Relationship between the ICC and Measures of Sleep Quality
4. Discussion
5. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Consensus Conference Panel; Watson, N.F.; Badr, M.S.; Belenky, G.; Bliwise, D.L.; Buxton, O.M.; Buysse, D.; Dinges, D.F.; Gangwisch, J.; Grandner, M.A.; et al. Joint Consensus Statement of the American Academy of Sleep Medicine and Sleep Research Society on the Recommended Amount of Sleep for a Healthy Adult: Methodology and Discussion. Sleep 2015, 38, 1161–1183. [Google Scholar] [CrossRef]
- The Lancet. Waking up to the importance of sleep. Lancet 2022, 400, 973. [Google Scholar] [CrossRef]
- Williams, R.L.; Agnew, H.W., Jr.; Webb, W.B. Sleep Patterns in Young Adults: An EEG Study. Electroencephalogr. Clin. Neurophysiol. 1964, 17, 376–381. [Google Scholar] [CrossRef]
- Williams, R.L.; Agnew, H.W., Jr.; Webb, W.B. Sleep patterns in the young adult female: An EEG study. Electroencephalogr. Clin. Neurophysiol. 1966, 20, 264–266. [Google Scholar] [CrossRef]
- Patel, A.K.; Reddy, V.; Shumway, K.R.; Araujo, J.F. Physiology, Sleep Stages; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Eugene, A.R.; Masiak, J. The Neuroprotective Aspects of Sleep. MEDtube Sci. 2015, 3, 35–40. [Google Scholar]
- Dijk, D.J.; Landolt, H.P. Sleep Physiology, Circadian Rhythms, Waking Performance and the Development of Sleep-Wake Therapeutics. Handb. Exp. Pharmacol. 2019, 253, 441–481. [Google Scholar] [CrossRef]
- Aime, M. To “feel” better, sleep on it! Science 2023, 382, 528. [Google Scholar] [CrossRef]
- Saper, C.B.; Fuller, P.M. Wake-sleep circuitry: An overview. Curr. Opin. Neurobiol. 2017, 44, 186–192. [Google Scholar] [CrossRef]
- Dijk, D.J.; von Schantz, M. Timing and consolidation of human sleep, wakefulness, and performance by a symphony of oscillators. J. Biol. Rhythm. 2005, 20, 279–290. [Google Scholar] [CrossRef]
- Deboer, T. Sleep homeostasis and the circadian clock: Do the circadian pacemaker and the sleep homeostat influence each other’s functioning? Neurobiol. Sleep Circadian Rhythm. 2018, 5, 68–77. [Google Scholar] [CrossRef]
- Borbely, A.A.; Daan, S.; Wirz-Justice, A.; Deboer, T. The two-process model of sleep regulation: A reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kuehnle, T.; Juda, M.; Kantermann, T.; Allebrandt, K.; Gordijn, M.; Merrow, M. Epidemiology of the human circadian clock. Sleep Med. Rev. 2007, 11, 429–438. [Google Scholar] [CrossRef]
- Wittmann, M.; Dinich, J.; Merrow, M.; Roenneberg, T. Social jetlag: Misalignment of biological and social time. Chronobiol. Int. 2006, 23, 497–509. [Google Scholar] [CrossRef]
- Van den Berg, J.F.; Kivela, L.; Antypa, N. Chronotype and depressive symptoms in students: An investigation of possible mechanisms. Chronobiol. Int. 2018, 35, 1248–1261. [Google Scholar] [CrossRef]
- Sudy, A.R.; Ella, K.; Bodizs, R.; Kaldi, K. Association of Social Jetlag with Sleep Quality and Autonomic Cardiac Control During Sleep in Young Healthy Men. Front. Neurosci. 2019, 13, 950. [Google Scholar] [CrossRef]
- Pilz, L.K.; Keller, L.K.; Lenssen, D.; Roenneberg, T. Time to rethink sleep quality: PSQI scores reflect sleep quality on workdays. Sleep 2018, 41, zsy029. [Google Scholar] [CrossRef]
- Wulterkens, B.M.; Fonseca, P.; Hermans, L.W.A.; Ross, M.; Cerny, A.; Anderer, P.; Long, X.; van Dijk, J.P.; Vandenbussche, N.; Pillen, S.; et al. It is All in the Wrist: Wearable Sleep Staging in a Clinical Population versus Reference Polysomnography. Nat. Sci. Sleep 2021, 13, 885–897. [Google Scholar] [CrossRef]
- Cook, J.D.; Eftekari, S.C.; Dallmann, E.; Sippy, M.; Plante, D.T. Ability of the Fitbit Alta HR to quantify and classify sleep in patients with suspected central disorders of hypersomnolence: A comparison against polysomnography. J. Sleep Res. 2018, e12789. [Google Scholar] [CrossRef]
- Enomoto, M.; Kitamura, S.; Nakazaki, K. Validity of an algorithm for determining sleep/wake states using FS-760 in school-aged children. J. Physiol. Anthropol. 2022, 41, 29. [Google Scholar] [CrossRef]
- Fitbit. Available online: https://help.fitbit.com/articles/en_US/Help_article/2163.htm (accessed on 5 December 2023).
- Weiss, C.; Woods, K.; Filipowicz, A.; Ingram, K.K. Sleep Quality, Sleep Structure, and PER3 Genotype Mediate Chronotype Effects on Depressive Symptoms in Young Adults. Front. Psychol. 2020, 11, 2028. [Google Scholar] [CrossRef]
- Von Gall, C.; Holub, L.; Pfeffer, M.; Eickhoff, S. Chronotype-Dependent Sleep Loss Is Associated with a Lower Amplitude in Circadian Rhythm and a Higher Fragmentation of REM Sleep in Young Healthy Adults. Brain Sci. 2023, 13, 1482. [Google Scholar] [CrossRef]
- Von Gall, C.; Muth, T.; Angerer, P. Sleep Duration on Workdays Is Correlated with Subjective Workload and Subjective Impact of High Workload on Sleep in Young Healthy Adults. Brain Sci. 2023, 13, 818. [Google Scholar] [CrossRef]
- Vivaldi, E.A.; Ocampo, A.; Wyneken, U.; Roncagliolo, M.; Zapata, A.M. Short-term homeostasis of active sleep and the architecture of sleep in the rat. J. Neurophysiol. 1994, 72, 1745–1755. [Google Scholar] [CrossRef]
- Ocampo-Garces, A.; Bassi, A.; Brunetti, E.; Estrada, J.; Vivaldi, E.A. REM sleep-dependent short-term and long-term hourglass processes in the ultradian organization and recovery of REM sleep in the rat. Sleep 2020, 43, zsaa023. [Google Scholar] [CrossRef]
- Ocampo-Garces, A.; Vivaldi, E.A. Short-term homeostasis of REM sleep assessed in an intermittent REM sleep deprivation protocol in the rat. J. Sleep Res. 2002, 11, 81–89. [Google Scholar] [CrossRef]
- Paech, G.M.; Ferguson, S.A.; Sargent, C.; Kennaway, D.J.; Roach, G.D. The relative contributions of the homeostatic and circadian processes to sleep regulation under conditions of severe sleep restriction. Sleep 2012, 35, 941–948. [Google Scholar] [CrossRef]
- Sargent, C.; Darwent, D.; Ferguson, S.A.; Kennaway, D.J.; Roach, G.D. Sleep restriction masks the influence of the circadian process on sleep propensity. Chronobiol. Int. 2012, 29, 565–571. [Google Scholar] [CrossRef]
- Dijk, D.J.; Archer, S.N. Circadian and Homeostatic Regulation of Human Sleep and Cognitive Performance and Its Modulation by PERIOD3. Sleep Med. Clin. 2009, 4, 111–125. [Google Scholar] [CrossRef]
- Nollet, M.; Wisden, W.; Franks, N.P. Sleep deprivation and stress: A reciprocal relationship. Interface Focus 2020, 10, 20190092. [Google Scholar] [CrossRef]
- Lee, M.L.; Swanson, B.E.; de la Iglesia, H.O. Circadian timing of REM sleep is coupled to an oscillator within the dorsomedial suprachiasmatic nucleus. Curr. Biol. 2009, 19, 848–852. [Google Scholar] [CrossRef]
- Barbato, G. REM Sleep: An Unknown Indicator of Sleep Quality. Int. J. Environ. Res. Public Health 2021, 18, 12976. [Google Scholar] [CrossRef]
- Wahlstrom, K.L.; Owens, J.A. School start time effects on adolescent learning and academic performance, emotional health and behaviour. Curr. Opin. Psychiatry 2017, 30, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, K.S.R.; Prado, I.M.; Abreu, L.G.; Serra-Negra, J.M.C.; Auad, S.M. Association among chronotype, dietary behaviours, and caries experience in Brazilian adolescents: Is there a behavioural pattern? Int. J. Paediatr. Dent. 2018, 28, 608–615. [Google Scholar] [CrossRef]
- Murkar, A.L.A.; De Koninck, J. Consolidative mechanisms of emotional processing in REM sleep and PTSD. Sleep Med. Rev. 2018, 41, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Pace-Schott, E.F.; Germain, A.; Milad, M.R. Sleep and REM sleep disturbance in the pathophysiology of PTSD: The role of extinction memory. Biol. Mood Anxiety Disord. 2015, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Germain, A.; Buysse, D.J.; Nofzinger, E. Sleep-specific mechanisms underlying posttraumatic stress disorder: Integrative review and neurobiological hypotheses. Sleep Med. Rev. 2008, 12, 185–195. [Google Scholar] [CrossRef]
- Meyer, N.; Lok, R.; Schmidt, C.; Kyle, S.D.; McClung, C.A.; Cajochen, C.; Scheer, F.; Jones, M.W.; Chellappa, S.L. The sleep-circadian interface: A window into mental disorders. Proc. Natl. Acad. Sci. USA 2024, 121, e2214756121. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Aeschbach, D. Sleep and anxiety: From mechanisms to interventions. Sleep Med. Rev. 2022, 61, 101583. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ICC50-H Deep | ICC50-H REM | |||||||
|---|---|---|---|---|---|---|---|---|
| Workdays | Work-Free Days | Workdays | Work-Free Days | |||||
| R | p | R | p | R | p | R | p | |
| Sleep loss | −0.1 | 0.45 | −0.25 | 0.056 | 0.03 | 0.82 | −0.18 | 0.16 |
| Tiredness | 0.15 | 0.25 | −0.15 | 0.42 | −0.04 | 0.78 | 0.05 | 0.73 |
| REM sleep | ||||||||
| Proportion | 0.09 | 0.52 | 0.1 | 0.44 | 0.29 | 0.025 * | 0.21 | 0.11 |
| Fragmentation | −0.12 | 0.35 | −0.12 | 0.38 | −0.25 | 0.053 | −0.16 | 0.22 |
| ICC50-C Deep | ICC50-C REM | |||||||
|---|---|---|---|---|---|---|---|---|
| Workdays | Work-Free Days | Workdays | Work-Free Days | |||||
| R | p | R | p | R | p | R | p | |
| Sleep loss | 0.13 | 0.34 | −0.49 | 0.00009 **** | 0.24 | 0.07 | −0.21 | 0.11 |
| Tiredness | 0.01 | 0.93 | −0.26 | 0.048 * | 0.04 | 0.78 | −0.22 | 0.09 |
| REM sleep | ||||||||
| Proportion | −0.08 | 0.53 | 0.24 | 0.06 | 0.27 | 0.03 * | 0.43 | 0.0007 *** |
| Fragmentation | 0.08 | 0.53 | −0.26 | 0.045 * | −0.28 | 0.03 * | −0.39 | 0.002 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
von Gall, C.; Holub, L.; Ali, A.A.H.; Eickhoff, S. Timing of Deep and REM Sleep Based on Fitbit Sleep Staging in Young Healthy Adults under Real-Life Conditions. Brain Sci. 2024, 14, 260. https://doi.org/10.3390/brainsci14030260
von Gall C, Holub L, Ali AAH, Eickhoff S. Timing of Deep and REM Sleep Based on Fitbit Sleep Staging in Young Healthy Adults under Real-Life Conditions. Brain Sciences. 2024; 14(3):260. https://doi.org/10.3390/brainsci14030260
Chicago/Turabian Stylevon Gall, Charlotte, Leon Holub, Amira A. H. Ali, and Simon Eickhoff. 2024. "Timing of Deep and REM Sleep Based on Fitbit Sleep Staging in Young Healthy Adults under Real-Life Conditions" Brain Sciences 14, no. 3: 260. https://doi.org/10.3390/brainsci14030260
APA Stylevon Gall, C., Holub, L., Ali, A. A. H., & Eickhoff, S. (2024). Timing of Deep and REM Sleep Based on Fitbit Sleep Staging in Young Healthy Adults under Real-Life Conditions. Brain Sciences, 14(3), 260. https://doi.org/10.3390/brainsci14030260