
Selection for Reduced Fear of Humans Changes Brain and Cerebellum Size in Red Junglefowl in Line with Effects of Chicken Domestication


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Note
2.2. Animals and Housing
2.3. Brain Dissection and Assessment of Neurons
2.4. Variables
2.5. Statistics
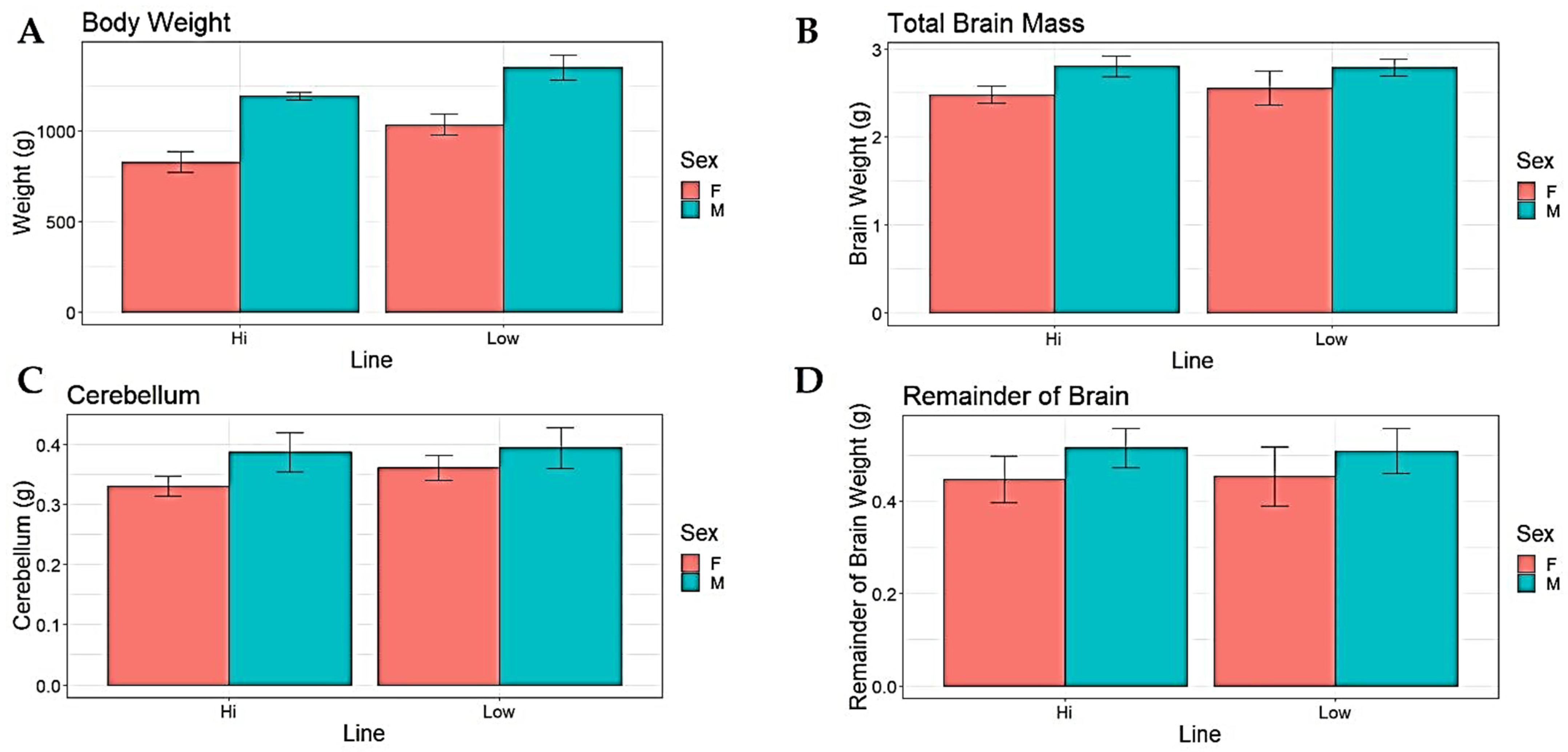
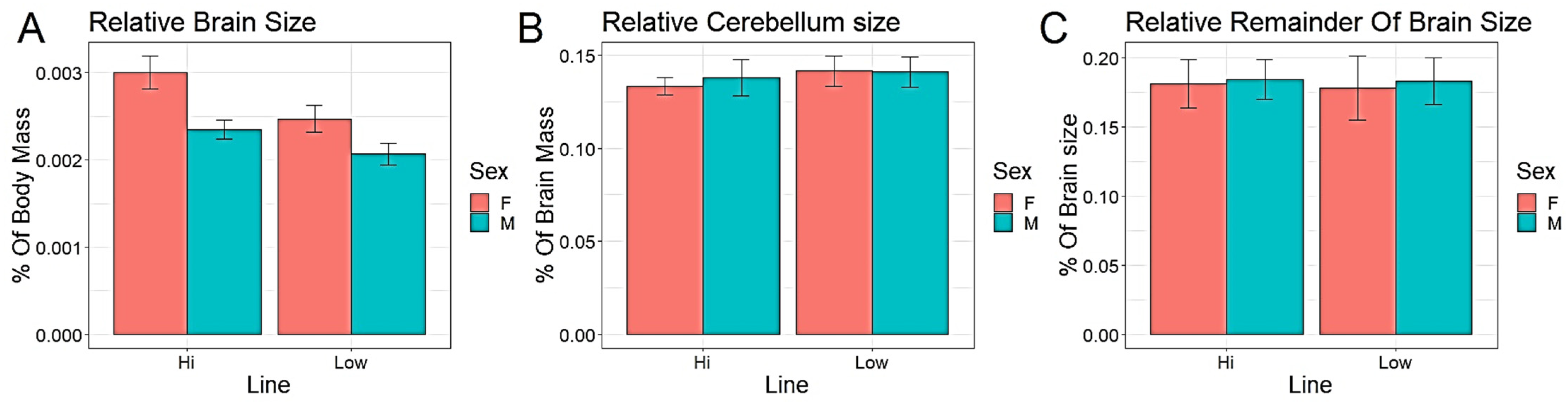
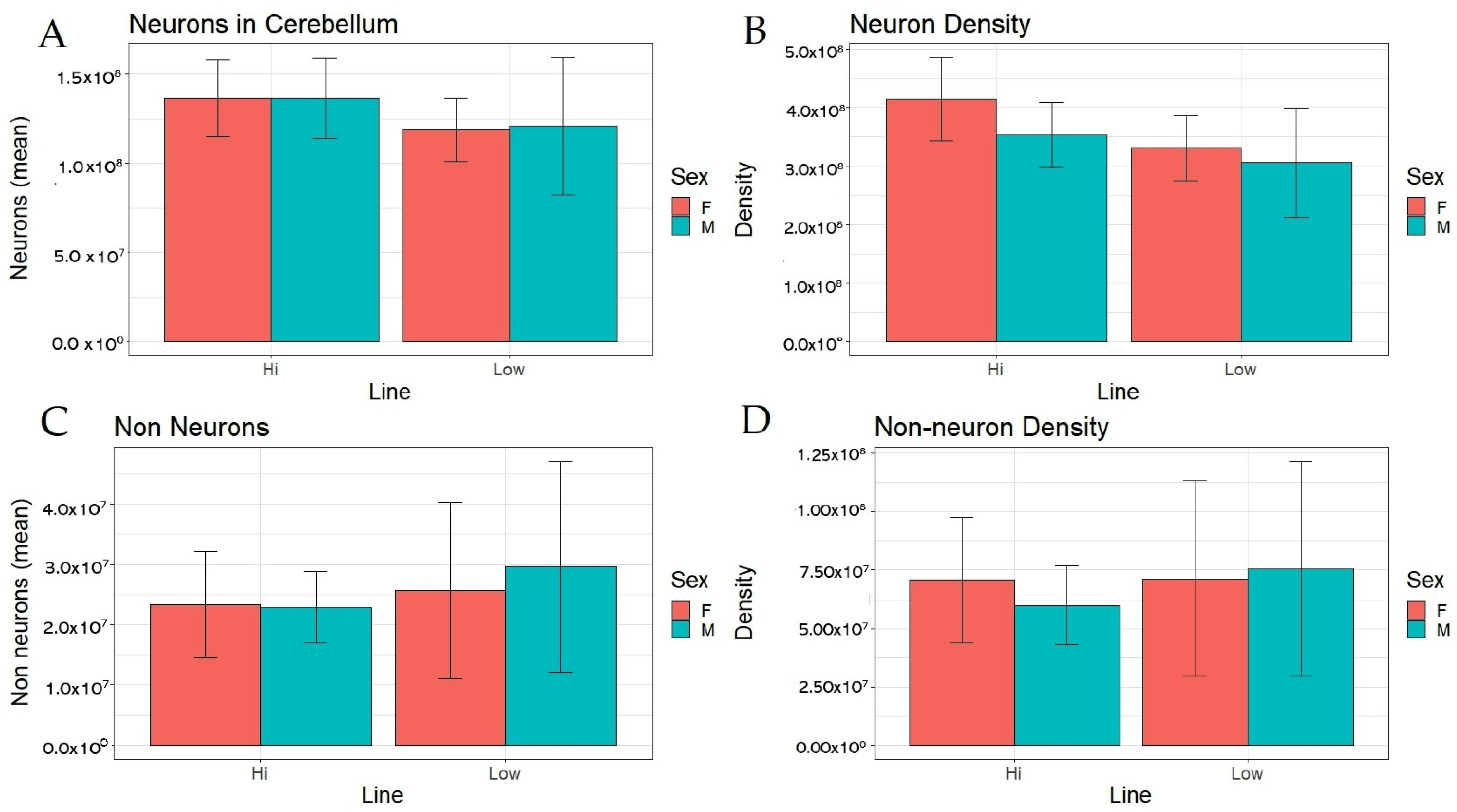
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, P.; Wright, D. Behavioral genetics and animal domestication. In Genetics and the Behavior of Domestic Animals; Elsevier: Amsterdam, The Netherlands, 2022; pp. 49–93. [Google Scholar]
- Price, E.O. Behavioral Aspects of Animal Domestication. Q. Rev. Biol. 1984, 59, 1–32. [Google Scholar] [CrossRef]
- Belyaev, D.K. Destabilizing selection as a factor in domestication. J. Hered. 1979, 70, 301–308. [Google Scholar] [CrossRef]
- Price, E.O. Animal Domestication and Behavior; Cabi: Wallingford, UK, 2002. [Google Scholar]
- Wright, D.; Henriksen, R.; Johnsson, M. Defining the Domestication Syndrome: Comment on Lord et al. 2020. Trends Ecol. Evol. 2020, 35, 1059–1060. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and Genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Rehkämper, G.; Frahm, H.D.; Cnotka, J. Mosaic Evolution and Adaptive Brain Component Alteration under Domestication Seen on the Background of Evolutionary Theory. Brain Behav. Evol. 2008, 71, 115–126. [Google Scholar] [CrossRef]
- Torres-Pérez, J.V.; Anagianni, S.; Mech, A.M.; Havelange, W.; García-González, J.; Fraser, S.E.; Vallortigara, G.; Brennan, C.H. baz1b loss-of-function in zebrafish produces phenotypic alterations consistent with the domestication syndrome. iScience 2023, 26, 105704. [Google Scholar] [CrossRef] [PubMed]
- Tixier-Boichard, M.; Bed’hom, B.; Rognon, X. Chicken domestication: From archeology to genomics. Comptes Rendus Biol. 2011, 334, 197–204. [Google Scholar] [CrossRef]
- Peters, J.; Lebrasseur, O.; Irving-Pease, E.K.; Paxinos, P.D.; Best, J.; Smallman, R.; Callou, C.; Gardeisen, A.; Trixl, S.; Frantz, L.; et al. The biocultural origins and dispersal of domestic chickens. Proc. Natl. Acad. Sci. USA 2022, 119, e2121978119. [Google Scholar] [CrossRef]
- Henriksen, R.; Johnsson, M.; Andersson, L.; Jensen, P.; Wright, D. The domesticated brain: Genetics of brain mass and brain structure in an avian species. Sci. Rep. 2016, 6, 34031. [Google Scholar] [CrossRef]
- Eklund, B.; Jensen, P. Domestication effects on behavioural synchronization and individual distances in chickens (Gallus gallus). Behav. Process. 2011, 86, 250–256. [Google Scholar] [CrossRef]
- Schuètz, K.E.; Jensen, P. Effects of resource allocation on behavioural strategies: A comparison of red junglefowl (Gallus gallus) and two domesticated breeds of poultry. Ethology 2001, 107, 753–765. [Google Scholar] [CrossRef]
- Bastian, A.J. Learning to predict the future: The cerebellum adapts feedforward movement control. Curr. Opin. Neurobiol. 2006, 16, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Adamaszek, M.; D’agata, F.; Ferrucci, R.; Habas, C.; Keulen, S.; Kirkby, K.C.; Leggio, M.; Mariën, P.; Molinari, M.; Moulton, E.; et al. Consensus Paper: Cerebellum and Emotion. Cerebellum 2017, 16, 552–576. [Google Scholar] [CrossRef] [PubMed]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’Arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus Paper: The Cerebellum’s Role in Movement and Cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef]
- Singer, T.; Seymour, B.; O’Doherty, J.; Kaube, H.; Dolan, R.J.; Frith, C.D. Empathy for Pain Involves the Affective but not Sensory Components of Pain. Science 2004, 303, 1157–1162. [Google Scholar] [CrossRef]
- Van Overwalle, F.; Manto, M.; Cattaneo, Z.; Clausi, S.; Ferrari, C.; Gabrieli, J.D.E.; Guell, X.; Heleven, E.; Lupo, M.; Ma, Q.; et al. Consensus Paper: Cerebellum and Social Cognition. Cerebellum 2020, 19, 833–868. [Google Scholar] [CrossRef] [PubMed]
- Dahhaoui, M.; Caston, J.; Auvray, N.; Reber, A. Role of the cerebellum in an avoidance conditioning task in the rat. Physiol. Behav. 1990, 47, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Katajamaa, R.; Wright, D.; Henriksen, R.; Jensen, P. Cerebellum size is related to fear memory and domestication of chickens. Biol. Lett. 2021, 17, 20200790. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R.; Botez, M. The cerebellum and learning processes in animals. Brain Res. Rev. 1990, 15, 325–332. [Google Scholar] [CrossRef]
- Sultan, F.; Glickstein, M. The cerebellum: Comparative and animal studies. Cerebellum 2008, 6, 168–176. [Google Scholar] [CrossRef]
- Nomura, T.; Izawa, E.-I. Avian brains: Insights from development, behaviors and evolution. Dev. Growth Differ. 2017, 59, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Voogd, J.; Glickstein, M. The anatomy of the cerebellum. Trends Cogn. Sci. 1998, 2, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Agnvall, B.; Bélteky, J.; Katajamaa, R.; Jensen, P. Is evolution of domestication driven by tameness? A selective review with focus on chickens. Appl. Anim. Behav. Sci. 2018, 205, 227–233. [Google Scholar] [CrossRef]
- Trut, L.; Oskina, I.; Kharlamova, A. Animal evolution during domestication: The domesticated fox as a model. Bioessays 2009, 31, 349–360. [Google Scholar] [CrossRef]
- Gjøen, J.; Jensen, P. Selection for Reduced Fear of Humans Changes Intra-Specific Social Behavior in Red Jungle-fowl—Implications for Chicken Domestication. Genes 2021, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Agnvall, B.; Jöngren, M.; Strandberg, E.; Jensen, P. Heritability and Genetic Correlations of Fear-Related Behaviour in Red Junglefowl–Possible Implications for Early Domestication. PLoS ONE 2012, 7, e35162. [Google Scholar] [CrossRef]
- Herculano-Houzel, S.; Lent, R. Isotropic Fractionator: A Simple, Rapid Method for the Quantification of Total Cell and Neuron Numbers in the Brain. J. Neurosci. 2005, 25, 2518–2521. [Google Scholar] [CrossRef]
- Mullen, R.J.; Buck, C.R.; Smith, A.M. NeuN, a neuronal specific nuclear protein in vertebrates. Development 1992, 116, 201–211. [Google Scholar] [CrossRef]
- Stingo-Hirmas, D.; Cunha, F.; Cardoso, R.F.; Carra, L.G.; Rönnegård, L.; Wright, D.; Henriksen, R. Proportional Cerebellum Size Predicts Fear Habituation in Chickens. Front. Physiol. 2022, 13, 124. [Google Scholar] [CrossRef]
- Ströckens, F.; Neves, K.; Kirchem, S.; Schwab, C.; Herculano-Houzel, S.; Güntürkün, O. High associative neuron numbers could drive cognitive performance in corvid species. J. Comp. Neurol. 2022, 530, 1588–1605. [Google Scholar] [CrossRef]
- Collias, N.E.; Collias, E.C. A Field Study of the Red Jungle Fowl in North-Central India. Condor 1967, 69, 360–386. [Google Scholar] [CrossRef]
- Hecht, E.E.; Barton, S.A.; Flattery, C.N.R.; Meza, A.M. The evolutionary neuroscience of domestication. Trends Cogn. Sci. 2023, 27, 553–567. [Google Scholar] [CrossRef]
- Agnvall, B.; Katajamaa, R.; Altimiras, J.; Jensen, P. Is domestication driven by reduced fear of humans? Boldness, metabolism and serotonin levels in divergently selected red junglefowl (Gallus gallus). Biol. Lett. 2015, 11, 20150509. [Google Scholar] [CrossRef] [PubMed]
- Racicot, K.J.; Popic, C.; Cunha, F.; Wright, D.; Henriksen, R.; Iwaniuk, A.N. The cerebellar anatomy of red junglefowl and white leghorn chickens: Insights into the effects of domesti-cation on the cerebellum. R. Soc. Open Sci. 2021, 8, 211002. [Google Scholar] [CrossRef] [PubMed]
- Rondi-Reig, L.; Dubreuil, Y.L.; Martinou, J.-C.; Delhaye-Bouchaud, N.; Caston, J.; Mariani, J. Fear decrease in transgenic mice over-expressing bcl-2 in neurons. Neuroreport 1997, 8, 2429–2432. [Google Scholar] [CrossRef]
- Byers, J.A.; Walker, C. Refining the Motor Training Hypothesis for the Evolution of Play. Am. Nat. 1995, 146, 25–40. [Google Scholar] [CrossRef]
- Myers, K.M.; Ressler, K.J.; Davis, M. Different mechanisms of fear extinction dependent on length of time since fear ac-quisition. Learn Mem. 2006, 13, 216–223. [Google Scholar] [CrossRef]
- Urrutia Desmaison, J.D.; Sala, R.W.; Ayyaz, A.; Nondhalee, P.; Popa, D.; Léna, C. Cerebellar control of fear learning via the cerebellar nuclei–Multiple pathways, multiple mechanisms? Front. Syst. Neurosci. 2023, 17, 1176668. [Google Scholar] [CrossRef]
- Frontera, J.L.; Sala, R.W.; Georgescu, I.A.; Aissa, H.B.; D’almeida, M.N.; Popa, D.; Léna, C. The cerebellum regulates fear extinction through thalamo-prefrontal cortex interactions in male mice. Nat. Commun. 2023, 14, 1508. [Google Scholar] [CrossRef]
- Palanza, P. Animal models of anxiety and depression: How are females different? Neurosci. Biobehav. Rev. 2001, 25, 219–233. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Dependent Variable | Line (Wald χ2) | Sex (Wald χ2) |
|---|---|---|
| Body Weight | 103.921 *** | 360.274 *** |
| Absolute Brain Size | 0.485 | 38.179 *** |
| Absolute Cerebellum Size | 4.250 * | 23.838 *** |
| Remainder of Brain Absolute Size | 0.001 | 12.220 *** |
| Number of Neurons | 3.687 | 0.013 |
| Number of Non-neurons | 1.185 | 0.190 |
| Neuron Density | 8.010 ** | 3.366 |
| Non-neuron Density | 0.499 | 0.080 |
| Relative Brain Size | 70.613 *** | 118.531 *** |
| Relative Cerebellum Size | 4.363 * | 0.556 |
| Relative Remainder of Brain Size | 0.977 | 0.972 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gjøen, J.; Cunha, F.; Jensen, P. Selection for Reduced Fear of Humans Changes Brain and Cerebellum Size in Red Junglefowl in Line with Effects of Chicken Domestication. Brain Sci. 2023, 13, 988. https://doi.org/10.3390/brainsci13070988
Gjøen J, Cunha F, Jensen P. Selection for Reduced Fear of Humans Changes Brain and Cerebellum Size in Red Junglefowl in Line with Effects of Chicken Domestication. Brain Sciences. 2023; 13(7):988. https://doi.org/10.3390/brainsci13070988
Chicago/Turabian StyleGjøen, Johanna, Felipe Cunha, and Per Jensen. 2023. "Selection for Reduced Fear of Humans Changes Brain and Cerebellum Size in Red Junglefowl in Line with Effects of Chicken Domestication" Brain Sciences 13, no. 7: 988. https://doi.org/10.3390/brainsci13070988
APA StyleGjøen, J., Cunha, F., & Jensen, P. (2023). Selection for Reduced Fear of Humans Changes Brain and Cerebellum Size in Red Junglefowl in Line with Effects of Chicken Domestication. Brain Sciences, 13(7), 988. https://doi.org/10.3390/brainsci13070988