
Cortical Structure Differences in Relation to Age, Sexual Attractions, and Gender Dysphoria in Adolescents: An Examination of Mean Diffusivity and T1 Relaxation Time
 , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.3. Measures
2.3.1. Age
2.3.2. Gender Dysphoria
2.3.3. Sexual Orientation
2.4. MRI Methods
2.4.1. Image Acquisition
2.4.2. Image Processing
2.5. Statistical Analyses
2.5.1. Group Differences in Demographic and Psychosexual Variables
2.5.2. Multivariate Correlation of Regional MD and T1 Relaxation Time
2.5.3. Multivariate Associations of Regional MD and T1 Relaxation Time with Age, Sexual Orientation, and Gender
3. Results
3.1. Differences in Demographic and Psychosexual Variables
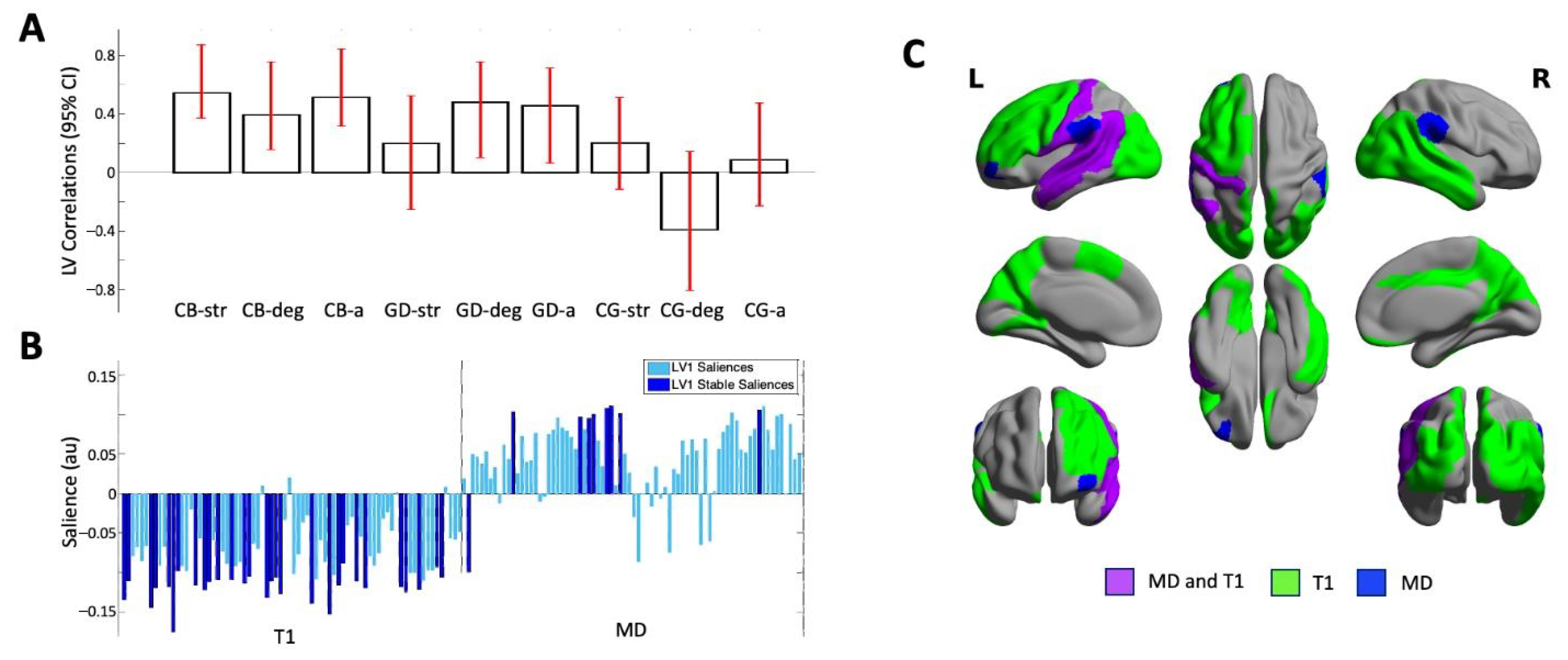
3.2. Multivariate Correlation of Regional MD and T1 Relaxation Time
3.3. Multivariate Associations of Regional MD and T1 Relaxation Time with Age and Sexual Attractions by Group
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frigerio, A.; Ballerini, L.; Valdes Hernandez, M. Structural, functional, and metabolic brain differences as a function of gender identity or sexual orientation: A systematic review of the human neuroimaging literature. Arch. Sex. Behav. 2021, 50, 3329–3352. [Google Scholar] [CrossRef] [PubMed]
- Guillamon, A.; Junque, C.; Gomez-Gil, E. A review of the status of brain structure research in transsexualism. Arch. Sex. Behav. 2016, 45, 1615–1648. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.M.; Manzouri, A.H.; Dhejne, C.; Bergstrom, K.; Arver, S.; Feusner, J.D.; Savic-Berglund, I. Testosterone effects on the brain in transgender men. Cereb. Cortex 2018, 28, 1582–1596. [Google Scholar] [CrossRef] [PubMed]
- Hoekzema, E.; Schagen, S.E.E.; Kreukels, B.P.C.; Veltman, D.J.; Cohen-Kettenis, P.T.; Delemarre-van de Waal, H.; Bakker, J. Regional volumes and spatial volumetric distribution of gray matter in the gender dysphoric brain. Psychoneuroendocrinology 2015, 55, 59–71. [Google Scholar] [CrossRef]
- Luders, E.; Sanchez, F.J.; Tosun, D.; Shattuck, D.W.; Gaser, C.; Vilain, E.; Toga, A.W. Increased cortical thickness in male-to-female transsexualism. J. Behav. Brain Sci. 2012, 2, 357–362. [Google Scholar] [CrossRef]
- Manzouri, A.; Savic, I. Possible neurobiological underpinnings of homosexuality and gender dysphoria. Cereb. Cortex 2019, 29, 2084–2101. [Google Scholar] [CrossRef]
- Mueller, S.C.; Guillamon, A.; Zubiaurre-Elorza, L.; Junque, C.; Gomez-Gil, E.; Uribe, C.; Khorashad, B.S.; Khazai, B.; Talaei, A.; Habel, U.; et al. The neuroanatomy of transgender identity: Mega-analytic findings from the ENIGMA transgender persons working group. J. Sex. Med. 2021, 18, 1122–1129. [Google Scholar] [CrossRef]
- Skorska, M.N.; Chavez, S.; Devenyi, G.A.; Patel, R.; Thurston, L.; Lai, M.-C.; Zucker, K.J.; Chakravarty, M.M.; Lobaugh, N.J.; VanderLaan, D. A multi-modal MRI analysis of cortical structure in relation to gender dysphoria, sexual orientation, and age in adolescents. J. Clin. Med. 2021, 10, 345. [Google Scholar] [CrossRef]
- Deoni, S.C.L. Quantitative relaxometry of the brain. Top. Magn. Reson. Imaging 2010, 21, 101–113. [Google Scholar] [CrossRef]
- Burke, S.M.; Manzouri, A.H.; Savic, I. Structural connections in the brain in relation to gender identity and sexual orientation. Sci. Rep. 2017, 7, 17954. [Google Scholar] [CrossRef]
- Kranz, G.S.; Hahn, A.; Kaufmann, U.; Kublbock, M.; Hummer, A.; Ganger, S.; Seiger, R.; Winkler, D.; Swaab, D.F.; Windischberger, C.; et al. White matter microstructure in transsexuals and controls investigated by diffusion tensor imaging. J. Neurosci. 2014, 34, 15466–15475. [Google Scholar] [CrossRef] [PubMed]
- Kranz, G.S.; Seiger, R.; Kaufmann, U.; Hummer, A.; Hahn, A.; Ganger, S.; Tik, M.; Windischberger, C.; Kasper, S.; Lanzenberger, R. Effects of sex hormone treatment on white matter microstructure in individuals with gender dysphoria. NeuroImage 2017, 150, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Kranz, G.S.; Zhang, B.B.B.; Handschuh, P.; Ritter, V.; Lanzenberger, R. Gender-affirming hormone treatment—A unique approach to study the effects of sex hormones on brain structure and function. Cortex 2020, 129, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Rametti, G.; Carrillo, B.; Gomez-Gil, E.; Junque, C.; Segovia, S.; Gomez, A.; Guillamon, A. White matter microstructure in female to male transsexuals before cross-sex hormonal treatment. A diffusion tensor imaging study. J. Psychiatr. Res. 2011, 45, 199–204. [Google Scholar] [CrossRef]
- Rametti, G.; Carrillo, B.; Gomez-Gil, E.; Junque, C.; Zubiarre-Elorza, L.; Segovia, S.; Gomez, A.; Guillamon, A. The microstructure of white matter in male to female transsexuals before cross-sex hormonal treatment. A DTI study. J. Psychiatr. Res. 2011, 45, 949–954. [Google Scholar] [CrossRef]
- Rametti, G.; Carrillo, B.; Gomez-Gil, E.; Junque, C.; Zubiarre-Elorza, L.; Segovia, S.; Gomez, A.; Karadi, K.; Guillamon, A. Effects of androgenization on the white matter microstructure of female-to-male transsexuals. A diffusion tensor imaging study. Psychoneuroendocrinology 2012, 37, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.M.; Marques, P.; Alves, V.; Sousa, N. A hitchhiker’s guide to diffusion tensor imaging. Front. Neurosci. 2013, 7, 31. [Google Scholar] [CrossRef]
- Tamnes, C.K.; Roalf, D.R.; Goddings, A.L.; Lebel, C. Diffusion MRI of white matter microstructure development in childhood and adolescence: Methods, challenges and progress. Dev. Cogn. Neurosci. 2018, 33, 161–175. [Google Scholar] [CrossRef]
- Elman, J.A.; Panizzon, M.S.; Hagler, D.J., Jr.; Fennema-Notestine, C.; Eyler, L.T.; Gillespie, N.A.; Neale, M.C.; Lyons, M.J.; Franz, C.E.; McEvoy, L.K.; et al. Genetic and environmental influences on cortical mean diffusivity. NeuroImage 2017, 146, 90–99. [Google Scholar] [CrossRef]
- Bonnier, G.; Marechal, B.; Fartaria, M.J.; Falkowskiy, P.; Marques, J.P.; Simioni, S.; Schluep, M.; Du Pasquier, R.; Thiran, J.P.; Krueger, G.; et al. The combined quantification and interpretation of multiple quantitative magnetic resonance imaging metrics enlightens longitudinal changes compatible with brain repair in relapsing-remitting multiple sclerosis patients. Front. Neurol. 2017, 8, 506. [Google Scholar] [CrossRef]
- Goubran, M.; Hammond, R.R.; de Ribaupierre, S.; Burneo, J.G.; Mirsattari, S.; Steven, D.A.; Parrent, A.G.; Peters, T.M.; Khan, A.R. Magnetic resonance imaging and histology correlation in the neocortex in temporal lobe epilepsy. Ann. Neurol. 2015, 77, 237–250. [Google Scholar] [CrossRef]
- Beaulieu, C. The biological basis of diffusion anisotropy. In Diffusion MRI, 2nd ed.; Johansen-Berg, H., Behrens, T.E.J., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 155–183. [Google Scholar] [CrossRef]
- Buyanova, I.S.; Arsalidou, M. Cerebral white matter myelination and relations to age, gender, and cognition: A selective review. Front. Hum. Neurosci. 2021, 15, 662031. [Google Scholar] [CrossRef]
- Hasan, K.M.; Moeller, F.G.; Narayana, P.A. DTI-based segmentation and quantification of human brain lateral ventricular CSF volumetry and mean diffusivity: Validation, age, gender effects and biophysical implications. Magn. Reson. Imaging 2014, 32, 405–412. [Google Scholar] [CrossRef]
- O’Donnell, L.J.; Westin, C.F. An introduction to diffusion tensor image analysis. Neurosurg. Clin. N. Am. 2011, 22, 185–196. [Google Scholar] [CrossRef]
- Laguna, P.A.L.; Combes, A.J.E.; Streffer, J.; Einstein, S.; Timmers, M.; Williams, S.C.R.; Dell’Acqua, F. Reproducibility, reliability and variability of FA and MD in the older healthy population: A test-retest multiparametric analysis. NeuroImage Clin. 2020, 26, 102168. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Press: Arlington, VA, USA, 2013. [Google Scholar]
- Skorska, M.N.; Lobaugh, N.J.; Lombardo, M.V.; van Bruggen, N.; Chavez, S.; Thurston, L.T.; Aitken, M.; Zucker, K.J.; Chakravarty, M.M.; Lai, M.-C.; et al. Inter-network brain functional connectivity in adolescents assigned female at birth who experience gender dysphoria. Front. Endocrinol. 2022, 13, 903058. [Google Scholar] [CrossRef]
- Skorska, M.N.; Thurston, L.T.; Biasin, J.M.; Devenyi, G.A.; Zucker, K.J.; Chakravarty, M.M.; Lai, M.-C.; VanderLaan, D.P. Data from: Cortical structure differences in relation to age, sexual attractions, and gender dysphoria in adolescents: An examination of mean diffusivity and T1 relaxation time. Borealis 2023. [Google Scholar] [CrossRef]
- Deogracias, J.J.; Johnson, L.L.; Meyer-Bahlburg, H.F.L.; Kessler, S.J.; Schober, J.M.; Zucker, K.J. The Gender Identity/Gender Dysphoria Questionnaire for Adolescents and Adults. J. Sex Res. 2007, 44, 370–379. [Google Scholar] [CrossRef]
- Singh, D.; Deogracias, J.J.; Johnson, L.J.; Bradley, S.J.; Kibblewhite, S.J.; Owen-Anderson, A.; Badali-Peterson, M.; Meyer-Bahlburg, H.F.L.; Zucker, K.J. The Gender Identity/Gender Dysphoria Questionnaire for Adolescents and Adults: Further validity evidence. J. Sex Res. 2010, 47, 49–58. [Google Scholar] [CrossRef]
- Drummond, K.D.; Bradley, S.J.; Badali-Peterson, M.; Zucker, K.J. A follow-up study of girls with gender identity disorder. Dev. Psychol. 2008, 44, 34–45. [Google Scholar] [CrossRef]
- Zucker, K.J.; Bradley, S.J.; Owen-Anderson, A.; Kibblewhite, S.J.; Wood, H.; Singh, D.; Choi, K. Demographics, behavior problems, and psychosexual characteristics of adolescents with gender identity disorder or transvestic fetishism. J. Sex Marital Ther. 2012, 38, 151–189. [Google Scholar] [CrossRef]
- Storms, M.D. Theories of sexual orientation. J. Pers. Soc. Psychol. 1980, 38, 783–792. [Google Scholar] [CrossRef]
- Zucker, K.J.; Bradley, S.J.; Oliver, G.; Blake, J.; Fleming, S.; Hood, J. Psychosexual development of women with congenital adrenal hyperplasia. Horm. Behav. 1996, 30, 300–318. [Google Scholar] [CrossRef]
- Deoni, S.C.L. High-resolution T1 mapping of the brain at 3T with driven equilibrium single pulse observation of T1 with high-speed incorporation of RF field inhomogeneities (DESPOT1-HIFI). J. Magn. Reson. Imaging 2007, 26, 1106–1111. [Google Scholar] [CrossRef]
- Wang, J.; Qiu, M.; Kim, H.; Constable, R.T. T1 Measurements incorporating flip angle calibration and correction in vivo. J. Mag. Reson. 2006, 182, 283–292. [Google Scholar] [CrossRef]
- Chavez, S.; Stanisz, G.J. A novel method for simultaneous 3D B1 and T1 mapping: The method of slopes (MoS). NMR Biomed. 2012, 25, 1043–1055. [Google Scholar] [CrossRef]
- Loken, C.; Gruner, D.; Groer, L.; Peltier, R.; Bunn, N.; Craig, M.; Henriques, T.; Dempsey, J.; Yu, C.-H.; Chen, J.; et al. SciNet: Lessons learned from building a power-efficient top-20 system and data centre. J. Phys. Conf. Ser. 2010, 256, 012026. [Google Scholar] [CrossRef]
- Ponce, M.; van Zon, R.; Northrup, S.; Gruner, D.; Chen, J.; Ertinaz, F.; Fedoseev, A.; Groer, L.; Mao, F.; Mundim, B.C.; et al. Deploying a top-100 supercomputer for large parallel workloads: The Niagara Supercomputer. In Proceedings of the Practice and Experience in Advanced Research Computing (PEARC 2019), Chicago, IL, USA, 28 July–1 August 2019. [Google Scholar] [CrossRef]
- Tustison, N.J.; Avants, B.B.; Cook, P.A.; Zheng, Y.; Egan, A.; Yushkevich, P.A.; Gee, J.C. N4ITK: Improved N3 bias correction. IEEE Trans. Med. Imaging 2010, 29, 1310–1320. [Google Scholar] [CrossRef]
- Eskildsen, S.F.; Coupe, P.; Fonov, V.; Manjon, J.V.; Leung, K.K.; Guizard, N.; Wassef, S.H.; Ostergaard, L.R.; Collins, D.L.; The Alzheimer’s Neuroimaging Initiative. BEaST: Brain extraction based on nonlocal segmentation technique. NeuroImage 2012, 59, 2362–2373. [Google Scholar] [CrossRef]
- Chavez, S. Calibrating variable flip angle (VFA)-based T1 maps: When and why a simple scaling factor is justified. In Proceedings of the International Society of Magnetic Resonance in Medicine (ISMRM), Paris, France, 16–21 June 2018. [Google Scholar]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Avants, B.B.; Tustison, N.J.; Song, G.; Cook, P.A.; Klein, A.; Gee, J.C. A reproducible evaluation of ANTs similarity metric performance in brain image registration. NeuroImage 2011, 54, 2033–2044. [Google Scholar] [CrossRef] [PubMed]
- Tustison, N.J.; Avants, B.B. Explicit B-spline regularization in diffeomorphic image registration. Front. Neuroinform. 2013, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.L.R.; Sotiropoulous, S.N. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. NeuroImage 2016, 125, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Muthen, L.K.; Muthen, B.O. Mplus User’s Guide, 8th ed.; Muthen & Muthen: Los Angeles, CA, USA, 1998–2017. [Google Scholar]
- Bell, A.; Fairbrother, M.; Jones, K. Fixed and random effects models: Making an informed choice. Qual. Quant. 2019, 53, 1051–1074. [Google Scholar] [CrossRef]
- McIntosh, A.R.; Bookstein, F.L.; Haxby, V.; Grady, C.L. Spatial pattern analysis of functional brain images using partial least squares. NeuroImage 1996, 3, 143–157. [Google Scholar] [CrossRef]
- McIntosh, A.R.; Lobaugh, N.J. Partial least squares analysis of neuroimaging data: Applications and advances. NeuroImage 2004, 23, S250–S263. [Google Scholar] [CrossRef]
- Geladi, P.; Kowalski, B.R. Partial Least-Squares Regression: A Tutorial. Analytica Chimica Acta 1986, 185, 1–17. [Google Scholar] [CrossRef]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef]
- Rapuano, K.M.; Laurent, J.S.; Hagler, D.J., Jr.; Hatton, S.N.; Thompson, W.K.; Jernigan, T.L.; Dale, A.M.; Casey, B.J.; Watts, R. Nucleus accumbens cytoarchitecture predicts weight gain in children. Proc. Natl. Acad. Sci. USA 2020, 117, 26977–26984. [Google Scholar] [CrossRef]
- White, N.S.; Leergaard, T.B.; D’Arceuil, H.; Bjaalie, J.G.; Dale, A.M. Probing tissue microstructure with restriction spectrum imaging: Histological and theoretical validation. Hum. Brain Mapp. 2013, 34, 327–346. [Google Scholar] [CrossRef]
- Zhang, H.; Schneider, T.; Wheeler-Kingshott, C.A.; Alexander, D.C. NODDI: Practical in vivo neurite orientation dispersion and density imaging of the human brain. NeuroImage 2012, 61, 1000–1016. [Google Scholar] [CrossRef]
- Herting, M.M.; Sowell, E.R. Puberty and structural brain development in humans. Front Neuroendocrinol. 2017, 44, 122–137. [Google Scholar] [CrossRef]
- Hines, M. Prenatal endocrine influences on sexual orientation and on sexually differentiated childhood behavior. Front. Neuroendocrinol. 2011, 32, 170–182. [Google Scholar] [CrossRef]
- Roselli, C.E. Neurobiology of gender identity and sexual orientation. J. Neuroendocrinol. 2018, 30, e12562. [Google Scholar] [CrossRef]
- Schulz, K.M.; Molenda-Figueira, H.A.; Sisk, C.L. Back to the future: The organizational–activational hypothesis adapted to puberty and adolescence. Horm. Behav. 2009, 55, 597–604. [Google Scholar] [CrossRef]
- Balthazart, J. Sexual partner preference in animals and humans. Neurosci. Biobehav. Rev. 2020, 115, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Bogaert, A.F.; Skorska, M.N. A short review of biological research on the development of sexual orientation. Horm. Behav. 2020, 119, 104659. [Google Scholar] [CrossRef] [PubMed]
- Gennatas, E.D.; Avants, B.B.; Wolf, D.H.; Satterthwaite, T.D.; Ruparel, K.; Ciric, R.; Hakonarson, H.; Gur, R.E.; Gur, R.C. Age-related effects and sex differences in gray matter density, volume, mass, and cortical thickness from childhood to young adulthood. J. Neurosci. 2017, 37, 5065–5073. [Google Scholar] [CrossRef] [PubMed]
- Breger, R.K.; Yetkin, Z.; Fischer, M.E.; Papke, R.A.; Haughton, V.M.; Rimm, A.A. T1 and T2 in the cerebrum: Correlation with age, gender, and demographic factors. Radiology 1991, 181, 545–547. [Google Scholar] [CrossRef]
- Colby, J.B.; O’Hare, E.D.; Bramen, J.E.; Sowell, E.R. Structural brain development: Birth through adolescence. In Neural Circuit Development and Function in the Brain; Rubenstein, J.L.R., Rakic, P., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 207–230. [Google Scholar] [CrossRef]
- Abi Ghanem, C.; Degerny, C.; Hussain, R.; Liere, P.; Pianos, A.; Tourpin, S.; Habert, R.; Macklin, W.B.; Schumacher, M.; Ghoumari, A.M. Long-lasting masculinizing effects of postnatal androgens on myelin governed by the brain androgen receptor. PLoS Genet. 2017, 13, e1007049. [Google Scholar] [CrossRef]
- Juraska, J.M.; Markham, J.A. The cellular basis for volume changes in the rat cortex during puberty: White and gray matter. Ann. N. Y. Acad. Sci. 2004, 1021, 431–435. [Google Scholar] [CrossRef]
- Marin-Husstege, M.; Muggironi, M.; Raban, D.; Skoff, R.P.; Casaccia-Bonnefil, P. Oligodendrocyte progenitor proliferation and maturation is differentially regulated by male and female sex steroid hormones. Dev. Neurosci. 2004, 26, 245–254. [Google Scholar] [CrossRef]
- Prayer, D.; Roberts, T.; Barkovich, A.J.; Prayer, L.; Kucharcyzk, J.; Moseley, M.; Arieff, A. Diffusion-weighted MRI of myelination in the rat brain following treatment with gonadal hormones. Neuroradiology 1997, 39, 320–325. [Google Scholar] [CrossRef]
- Stuber, C.; Morawski, M.; Schafer, A.; Labadie, C.; Wahnert, M.; Leuze, C.; Streigher, M.; Barapatre, N.; Reimann, K.; Geyer, S.; et al. Myelin and iron concentration in the human brain: A quantitative study of MRI contrast. NeuroImage 2014, 93, 95–106. [Google Scholar] [CrossRef]


{kind=link}
| Cisgender Boys | GD AFAB | Cisgender Girls | F (df) | p | |
|---|---|---|---|---|---|
| n | 14 | 15 | 17 | ||
| Age (months) a | |||||
| M | 184.93 | 193.07 | 191.71 | 0.68 (2, 43) | 0.510 |
| SD | 25.61 | 14.65 | 19.00 | ||
| Range | 147–216 | 162–216 | 152–214 | ||
| GIDYQ-AA | |||||
| M | 4.91 | 2.17 | 4.90 | 778.90 (2, 43) d | <0.001 |
| SD | 0.12 | 0.33 | 0.15 | ||
| Range (1–5) b | 4.63–5.00 | 1.74–3.04 | 4.48–5.00 | ||
| Strength of attractions | |||||
| M | 3.25 | 2.98 | 2.83 | 0.53 (2, 43) e | 0.591 |
| SD | 1.28 | 1.34 | 0.76 | ||
| Range (1.41–7.07) c | 1.70–5.37 | 1.41–6.05 | 1.41–4.44 | ||
| Degree of androphilia–gynephilia | |||||
| M | 64.60 | 46.99 | 30.94 | 25.67 (2, 43) | <0.001 |
| SD | 9.29 | 15.56 | 13.18 | ||
| Range (11–79) b | 43.96–75.55 | 17.35–75.55 | 14.04–68.20 | ||
| Hem. | ROI | ||
|---|---|---|---|
| Frontal Lobe | T1 | MD | |
| L | Inferior Frontal Gyrus: Opercular Part | x | - |
| L | Inferior Frontal Gyrus: Triangular Part | x | - |
| L | Middle Frontal Gyrus | x | - |
| L | Middle Frontal Gyrus: Orbital Part | - | x |
| L | Precentral Gyrus | x | - |
| L | Rolandic Operculum | x | x |
| L | Superior Frontal Gyrus: Dorsolateral | x | - |
| L | Supplementary Motor Area | x | - |
| Parietal Lobe | |||
| L | Angular Gyrus | x | x |
| R | Angular Gyrus | x | - |
| L | Postcentral Gyrus | x | x |
| L | Precuneus | x | - |
| R | Precuneus | x | - |
| L | Supramarginal Gyrus | - | x |
| R | Supramarginal Gyrus | - | x |
| Temporal Lobe | |||
| L | Heschl Gyrus | x | x |
| R | Heschl Gyrus | x | - |
| R | Inferior Temporal Gyrus | x | - |
| L | Middle Temporal Gyrus | x | x |
| R | Middle Temporal Gyrus | x | - |
| L | Superior Temporal Gyrus | x | x |
| R | Superior Temporal Gyrus | x | - |
| Occipital Lobe | |||
| L | Cuneus | x | - |
| R | Cuneus | x | - |
| L | Inferior Occipital Gyrus | x | - |
| R | Inferior Occipital Gyrus | x | - |
| L | Lingual Gyrus | x | - |
| L | Middle Occipital Gyrus | x | - |
| R | Middle Occipital Gyrus | x | - |
| L | Superior Occipital Gyrus | x | - |
| R | Superior Occipital Gyrus | x | - |
| Insula and Cingulate Gyri | |||
| R | Median Cingulate and Paracingulate Gyri | x | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skorska, M.N.; Thurston, L.T.; Biasin, J.M.; Devenyi, G.A.; Zucker, K.J.; Chakravarty, M.M.; Lai, M.-C.; VanderLaan, D.P. Cortical Structure Differences in Relation to Age, Sexual Attractions, and Gender Dysphoria in Adolescents: An Examination of Mean Diffusivity and T1 Relaxation Time. Brain Sci. 2023, 13, 963. https://doi.org/10.3390/brainsci13060963
Skorska MN, Thurston LT, Biasin JM, Devenyi GA, Zucker KJ, Chakravarty MM, Lai M-C, VanderLaan DP. Cortical Structure Differences in Relation to Age, Sexual Attractions, and Gender Dysphoria in Adolescents: An Examination of Mean Diffusivity and T1 Relaxation Time. Brain Sciences. 2023; 13(6):963. https://doi.org/10.3390/brainsci13060963
Chicago/Turabian StyleSkorska, Malvina N., Lindsey T. Thurston, Jessica M. Biasin, Gabriel A. Devenyi, Kenneth J. Zucker, M. Mallar Chakravarty, Meng-Chuan Lai, and Doug P. VanderLaan. 2023. "Cortical Structure Differences in Relation to Age, Sexual Attractions, and Gender Dysphoria in Adolescents: An Examination of Mean Diffusivity and T1 Relaxation Time" Brain Sciences 13, no. 6: 963. https://doi.org/10.3390/brainsci13060963
APA StyleSkorska, M. N., Thurston, L. T., Biasin, J. M., Devenyi, G. A., Zucker, K. J., Chakravarty, M. M., Lai, M.-C., & VanderLaan, D. P. (2023). Cortical Structure Differences in Relation to Age, Sexual Attractions, and Gender Dysphoria in Adolescents: An Examination of Mean Diffusivity and T1 Relaxation Time. Brain Sciences, 13(6), 963. https://doi.org/10.3390/brainsci13060963