
Emergence of Treadmill Running Ability and Quantitative Assessment of Gait Dynamics in Young Ts65Dn Mice: A Mouse Model for Down Syndrome

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Gait Dynamics Recording
2.3. Treadmill Longitudinal Training Procedure
2.4. Paw Classification Analysis
2.5. Statistical Analyses
3. Results
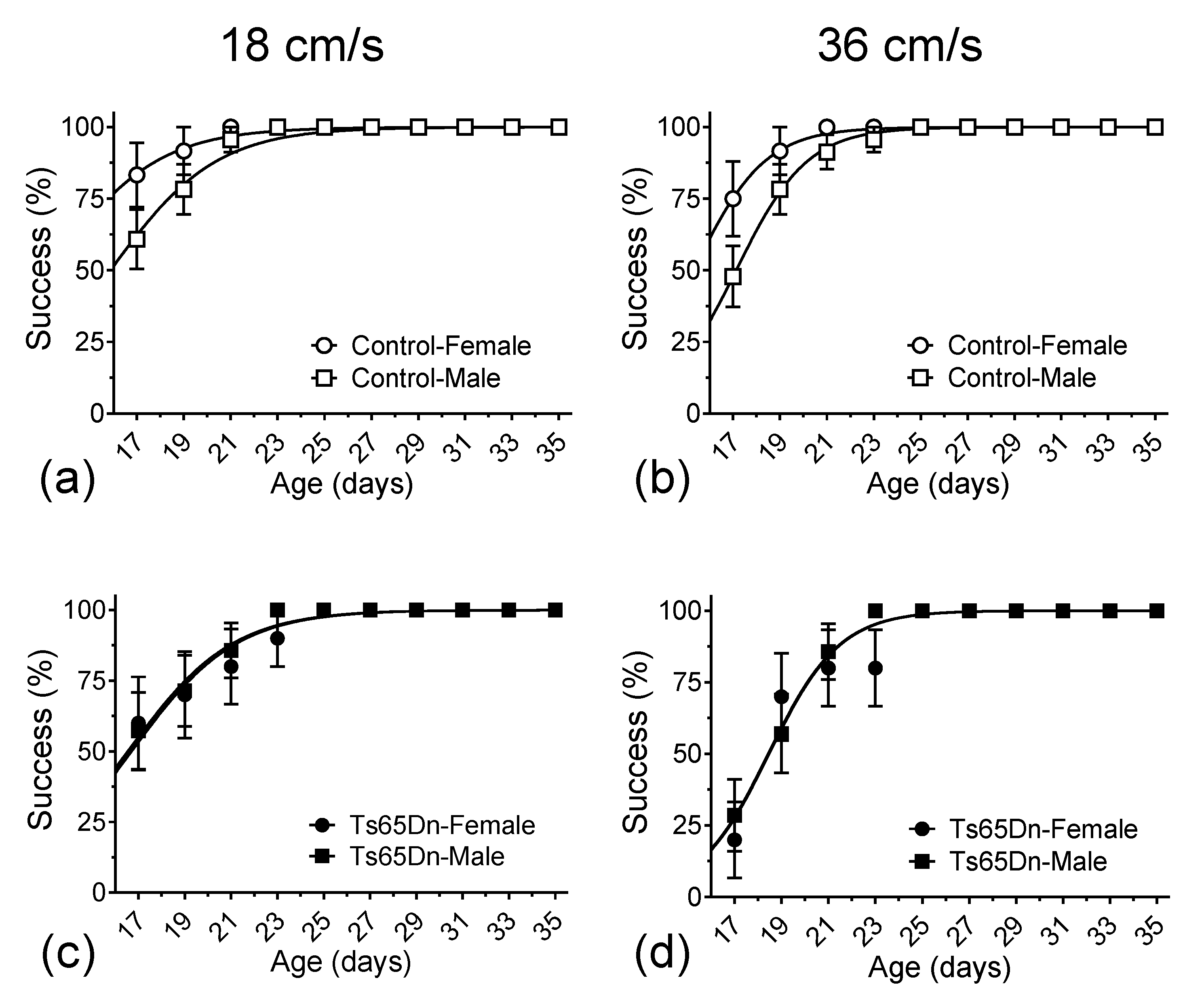
3.1. Delay in Motor Development
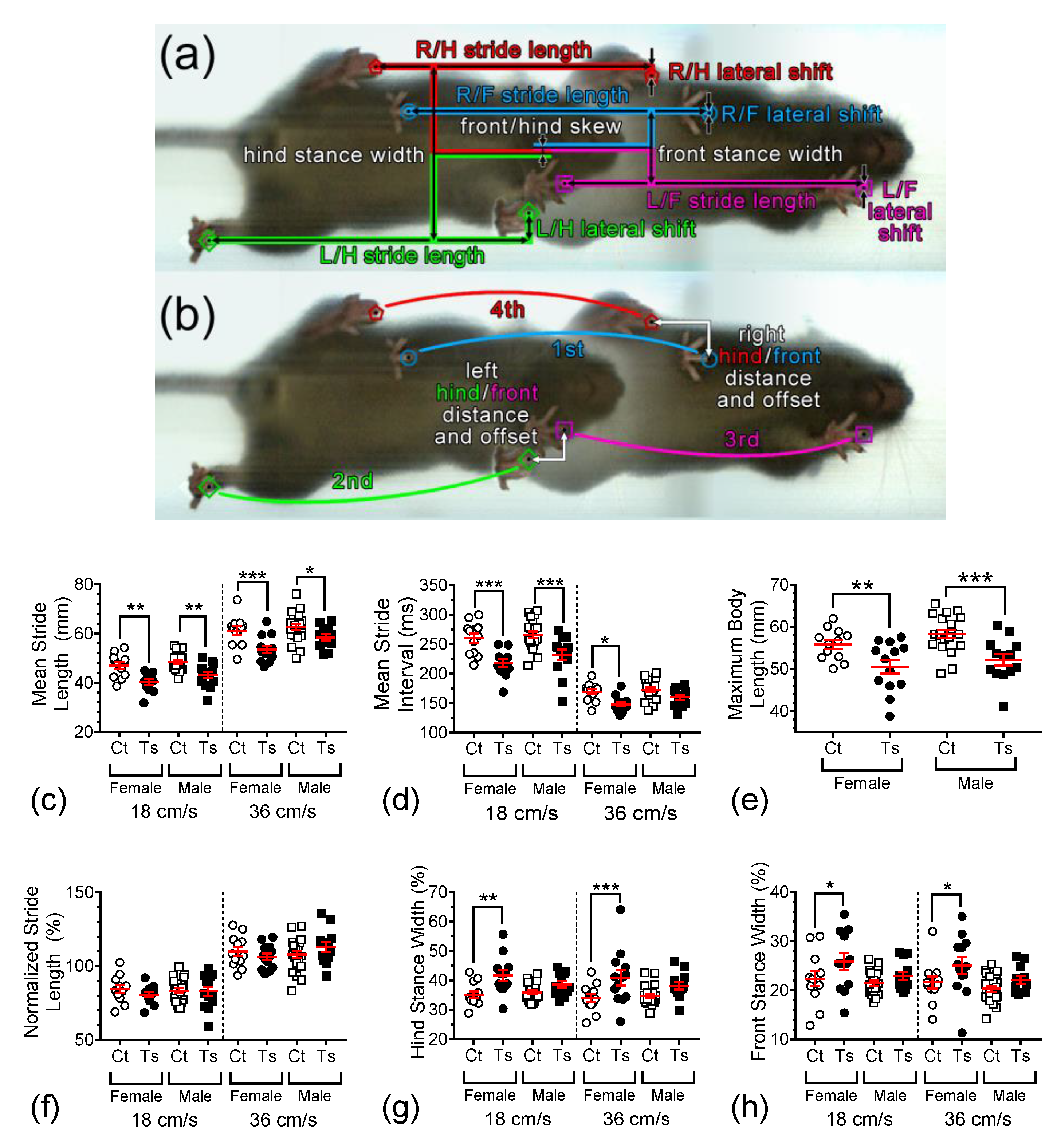
3.2. Gait Dynamics
3.3. Variability in Gait Dynamics Measures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Normalized Gait Dynamics Measure from Figure A2 (Panel) | ANOVA Results for Each Dependent Variable F1,108; p = | Post Hoc Pairwise Comparisons p = @18 cm/s; p = @36 cm/s | |||||
|---|---|---|---|---|---|---|---|
| Speed | Genotype | Sex | Ts65Dn Female vs. Control Female | Ts65Dn Male vs. Control Male | Ts65Dn Female vs. Ts65Dn Male | Control Female vs. Control Male | |
| Swing Percentage (a) | 273.61; <0.0001 | 0.85; 0.3574 | 6.15; 0.0147 | N/A | N/A | N/A | N/A |
| Lateral Shift (b) | 0.0745; 0.7853 | 0.0913; 0.7631 | 0.0000; 0.9947 | N/A | N/A | N/A | N/A |
| Front/Hind Distance (c) | 88.203; <0.0001 | 8.4596; 0.0044 | 1.0436; 0.3093 | 0.3668; 0.0840 | 0.2550; 0.0412 | 0.6618; 0.1936 | 0.6960; 0.1161 |
| Hind/Front Offset (d) | 0.028; 0.8674 | 9.589; 0.0025 | 1.702; 0.1948 | 0.0431; 0.0519 | 0.4052; 0.2206 | 0.9063; 0.7952 | 0.1756; 0.2212 |
| Front/Hind Skew (e) | 0.8042; 0.37185 | 1.10640; 0.2952 | 0.0120;0.9130 | N/A | N/A | N/A | N/A |
References
- Patterson, D.; Costa, A.C. Down syndrome and genetics—A case of linked histories. Nat. Rev. Genet. 2005, 6, 137–147. [Google Scholar] [CrossRef]
- Mai, C.T.; Isenburg, J.L.; Canfield, M.A.; Meyer, R.E.; Correa, A.; Alverson, C.J.; Lupo, P.J.; Riehle-Colarusso, T.; Cho, S.J.; Aggarwal, D.; et al. National population-based estimates for major birth defects, 2010–2014. Birth Defects Res. 2019, 111, 1420–1435. [Google Scholar] [CrossRef]
- Turner, S.; Alborz, A. Academic attainments of children with Down’s syndrome: A longitudinal study. Br. J. Educ. Psychol. 2003, 73, 563–583. [Google Scholar] [CrossRef]
- Abbeduto, L.; Warren, S.F.; Conners, F.A. Language development in Down syndrome: From the prelinguistic period to the acquisition of literacy. Ment. Retard Dev. Disabil Res. Rev. 2007, 13, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.S. Language learning in Down syndrome: The speech and language profile compared to adolescents with cognitive impairMent. of unknown origin. Downs Syndr. Res. Pract. 2006, 10, 61–66. [Google Scholar] [CrossRef]
- Chapman, R.S.; Hesketh, L.J. Behavioral phenotype of individuals with Down syndrome. Ment. Retard Dev. Disabil Res. Rev. 2000, 6, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Pennington, B.F.; Moon, J.; Edgin, J.; Stedron, J.; Nadel, L. The neuropsychology of Down syndrome: Evidence for hippocampal dysfunction. Child Dev. 2003, 74, 75–93. [Google Scholar] [CrossRef]
- Leverenz, J.B.; Raskind, M.A. Early amyloid deposition in the medial temporal lobe of young Down syndrome patients: A regional quantitative analysis. Exp. Neurol. 1998, 150, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Zigman, W.; Schupf, N.; Haveman, M.; Silverman, W. The epidemiology of Alzheimer disease in intellectual disability: Results and recommendations from an international conference. J. Intellect Disabil. Res. 1997, 41 Pt 1, 76–80. [Google Scholar] [CrossRef]
- Carter, G.; Jancar, J. Mortality in the mentally handicapped: A 50 year survey at the Stoke Park group of hospitals (1930–1980). J. Ment. Defic. Res. 1983, 27 Pt 2, 143–156. [Google Scholar] [CrossRef]
- Puri, B.K.; Lekh, S.K.; Langa, A.; Zaman, R.; Singh, I. Mortality in a hospitalized mentally handicapped population: A 10-year survey. J. Intellect Disabil. Res. 1995, 39 Pt 5, 442–446. [Google Scholar] [CrossRef]
- Hampton, T.G.; Stasko, M.R.; Kale, A.; Amende, I.; Costa, A.C. Gait dynamics in trisomic mice: Quantitative neurological traits of Down syndrome. Physiol. Behav. 2004, 82, 381–389. [Google Scholar] [CrossRef]
- Winders, P.; Wolter-Warmerdam, K.; Hickey, F. A schedule of gross motor developMent. for children with Down syndrome. J. Intellect Disabil. Res. 2019, 63, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Anson, J.G. Chapter 13 Neuromotor Control and Down Syndrome. In Advances in Psychology; Summers, J.J., Ed.; North-Holland: Amsterdam, The Netherlands, 1992; Volume 84, pp. 387–412. [Google Scholar]
- Aruin, A.S.; Almeida, G.L.; Latash, M.L. Organization of a simple two-joint synergy in individuals with Down syndrome. Am. J. Ment. Retard. 1996, 101, 256–268. [Google Scholar] [PubMed]
- Latash, M.L. Motor control in Down syndrome: The role of adaptation and practice. J. Dev. Phys. Disabil. 1992, 4, 227–261. [Google Scholar] [CrossRef]
- Shumway-Cook, A.; Woollacott, M.H. Dynamics of postural control in the child with Down syndrome. Phys. Ther. 1985, 65, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.C. An assessMent. of optokinetic nystagmus (OKN) in persons with Down syndrome. Exp. Brain Res. 2011, 214, 381–391. [Google Scholar] [CrossRef]
- Costa, A.C. An assessMent. of the vestibulo-ocular reflex (VOR) in persons with Down syndrome. Exp. Brain Res. 2011, 214, 199–213. [Google Scholar] [CrossRef]
- Alsakhawi, R.S.; Elshafey, M.A. Effect of Core Stability Exercises and Treadmill Training on Balance in Children with Down Syndrome: Randomized Controlled Trial. Adv Ther. 2019, 36, 2364–2373. [Google Scholar] [CrossRef]
- Baxter, L.L.; Moran, T.H.; Richtsmeier, J.T.; Troncoso, J.; Reeves, R.H. Discovery and genetic localization of Down syndrome cerebellar phenotypes using the Ts65Dn mouse. Hum. Mol. Genet. 2000, 9, 195–202. [Google Scholar] [CrossRef]
- Bodensteiner, J.B.; Smith, S.D.; Schaefer, G.B. Hypotonia, congenital hearing loss, and hypoactive labyrinths. J. Child Neurol. 2003, 18, 171–173. [Google Scholar] [CrossRef]
- Koo, B.K.; Blaser, S.; Harwood-Nash, D.; Becker, L.E.; Murphy, E.G. Magnetic resonance imaging evaluation of delayed myelination in Down syndrome: A case report and review of the literature. J. Child Neurol. 1992, 7, 417–421. [Google Scholar] [CrossRef]
- Molnar, G.E. Analysis of motor disorder in retarded infants and young children. Am. J. Ment. Defic. 1978, 83, 213–222. [Google Scholar] [PubMed]
- Davisson, M.T.; Schmidt, C.; Akeson, E.C. Segmental trisomy of murine chromosome 16: A new model system for studying Down syndrome. Prog. Clin. Biol. Res. 1990, 360, 263–280. [Google Scholar] [PubMed]
- Duchon, A.; Raveau, M.; Chevalier, C.; Nalesso, V.; Sharp, A.J.; Herault, Y. Identification of the translocation breakpoints in the Ts65Dn and Ts1Cje mouse lines: Relevance for modeling Down syndrome. Mamm. Genome. 2011, 22, 674–684. [Google Scholar] [CrossRef]
- Costa, A.C.; Scott-McKean, J.J. Prospects for improving brain function in individuals with Down syndrome. CNS Drugs 2013, 27, 679–702. [Google Scholar] [CrossRef]
- Costa, A.C.; Walsh, K.; Davisson, M.T. Motor dysfunction in a mouse model for Down syndrome. Physiol. Behav. 1999, 68, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Holtzman, D.M.; Santucci, D.; Kilbridge, J.; Chua-Couzens, J.; Fontana, D.J.; Daniels, S.E.; Johnson, R.M.; Chen, K.; Sun, Y.; Carlson, E.; et al. Developmental abnormalities and age-related neurodegeneration in a mouse model of Down syndrome. Proc. Natl. Acad. Sci. USA 1996, 93, 13333–13338. [Google Scholar] [CrossRef] [PubMed]
- Olmos-Serrano, J.L.; Tyler, W.A.; Cabral, H.J.; Haydar, T.F. Longitudinal measuRes. of cognition in the Ts65Dn mouse: Refining windows and defining modalities for therapeutic intervention in Down syndrome. Exp. Neurol. 2016, 279, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.C.; Stasko, M.R.; Schmidt, C.; Davisson, M.T. Behavioral validation of the Ts65Dn mouse model for Down syndrome of a genetic background free of the retinal degeneration mutation Pde6b(rd1). Behav. Brain Res. 2010, 206, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Akula, S.K.; McCullough, K.B.; Weichselbaum, C.; Dougherty, J.D.; Maloney, S.E. The trajectory of gait developMent. in mice. Brain Behav. 2020, 10, e01636. [Google Scholar] [CrossRef]
- Ahmed, M.M.; Block, A.; Busquet, N.; Gardiner, K.J. Context Fear Conditioning in Down Syndrome Mouse Models: Effects of Trisomic Gene Content, Age, Sex and Genetic Background. Genes 2021, 12, 1528. [Google Scholar] [CrossRef]
- Chakrabarti, L.; Scafidi, J.; Gallo, V.; Haydar, T.F. Environmental enrichMent. rescues postnatal neurogenesis defect in the male and female Ts65Dn mouse model of Down syndrome. Dev. Neurosci. 2011, 33, 428–441. [Google Scholar] [CrossRef]
- Martinez-Cue, C.; Baamonde, C.; Lumbreras, M.; Paz, J.; Davisson, M.T.; Schmidt, C.; Dierssen, M.; Florez, J. Differential effects of environmental enrichMent. on behavior and learning of male and female Ts65Dn mice, a model for Down syndrome. Behav. Brain Res. 2002, 134, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Cue, C.; Rueda, N.; Garcia, E.; Florez, J. Anxiety and panic responses to a predator in male and female Ts65Dn mice, a model for Down syndrome. Genes Brain Behav. 2006, 5, 413–422. [Google Scholar] [CrossRef]
- Thomas, J.R.; Sloan, K.; Cave, K.; Wallace, J.M.; Roper, R.J. Skeletal Deficits in Male and Female down Syndrome Model Mice Arise Independent of Normalized Dyrk1a Expression in Osteoblasts. Genes 2021, 12, 1729. [Google Scholar] [CrossRef] [PubMed]
- Duchon, A.; Del Mar Muniz Moreno, M.; Chevalier, C.; Nalesso, V.; Andre, P.; Fructuoso-Castellar, M.; Mondino, M.; Po, C.; Noblet, V.; Birling, M.C.; et al. Ts66Yah, a mouse model of Down syndrome with improved construct and face validity. Dis. Model. Mech. 2022, 15, dmm049721. [Google Scholar] [CrossRef]
- Kazuki, Y.; Gao, F.J.; Li, Y.; Moyer, A.J.; Devenney, B.; Hiramatsu, K.; Miyagawa-Tomita, S.; Abe, S.; Kazuki, K.; Kajitani, N.; et al. A non-mosaic transchromosomic mouse model of down syndrome carrying the long arm of human chromosome 21. Elife 2020, 9, e56223. [Google Scholar] [CrossRef] [PubMed]




Measure Variability Figure 4 (Panel) | ANOVA Results for Each Dependent Variable F1,108; p = | Post Hoc Pairwise Comparisons p = @18cm/s; p = @36cm/s | |||||
|---|---|---|---|---|---|---|---|
| Speed | Genotype | Sex | Ts65Dn Female vs. Control Female | Ts65Dn Male vs Control Male | Ts65Dn Female vs. Ts65Dn Male | Control Female vs. Control Male | |
| Swing Percentage (c) | 20.193; <0.0001 | 31.408; <0.0001 | 1.412; 0.2373 | 0.1334; <0.0001 | 0.0003; 0.0577 | 0.5106; 0.0349 | 0.2429; 0.7638 |
| Hind Stance Width (d) | 1.777; 0.1853 | 32.840; <0.0001 | 11.826; 0.0008 | 0.0760; <0.0001 | 0.1138; 0.0148 | 0.0533; 0.0022 | 0.0931; 0.9370 |
| Front Stance Width (e) | 0.3965; 0.5302 | 28.1412; <0.0001 | 1.5217; 0.2200 | 0.0918; <0.0001 | 0.1705; 0.0141 | 0.4371; 0.0502 | 0.7727; 0.4601; |
| Lateral Shift (f) | 0.6177; 0.4336 | 36.9112; <0.0001 | 2.1496; 0.1455 | 0.0391; <0.0001 | 0.1753; 0.0004 | 0.1739; 0.2009 | 0.6265; 0.7391 |
| Front/Hind Distance (g) | 15.275; 0.0002 | 23.6809; <0.0001 | 11.559; 0.0009 | 0.1896; 0.0001 | 0.0547; 0.0167 | 0.0428; 0.0575 | 0.0087; 0.9154 |
| Hind/Front Offset (h) | 0.8000; 0.3731 | 31.7336; <0.0001 | 6.3541; 0.0132 | 0.0172; <0.0001 | 0.0145; 0.1366 | 0.2161; 0.0035 | 0.2871; 0.6681 |
| Front/Hind Skew (i) | 0.2981; 0.5862 | 28.7693; <0.0001 | 1.9632; 0.1640 | 0.0573; 0.0001 | 0.0543; 0.0061 | 0.3222; 0.2297 | 0.4087; 0.7563 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scott-McKean, J.J.; Jones, R.; Johnson, M.W.; Mier, J.; Basten, I.A.; Stasko, M.R.; Costa, A.C.S. Emergence of Treadmill Running Ability and Quantitative Assessment of Gait Dynamics in Young Ts65Dn Mice: A Mouse Model for Down Syndrome. Brain Sci. 2023, 13, 743. https://doi.org/10.3390/brainsci13050743
Scott-McKean JJ, Jones R, Johnson MW, Mier J, Basten IA, Stasko MR, Costa ACS. Emergence of Treadmill Running Ability and Quantitative Assessment of Gait Dynamics in Young Ts65Dn Mice: A Mouse Model for Down Syndrome. Brain Sciences. 2023; 13(5):743. https://doi.org/10.3390/brainsci13050743
Chicago/Turabian StyleScott-McKean, Jonah J., Ryan Jones, Mark W. Johnson, Joyce Mier, Ines A. Basten, Melissa R. Stasko, and Alberto C. S. Costa. 2023. "Emergence of Treadmill Running Ability and Quantitative Assessment of Gait Dynamics in Young Ts65Dn Mice: A Mouse Model for Down Syndrome" Brain Sciences 13, no. 5: 743. https://doi.org/10.3390/brainsci13050743
APA StyleScott-McKean, J. J., Jones, R., Johnson, M. W., Mier, J., Basten, I. A., Stasko, M. R., & Costa, A. C. S. (2023). Emergence of Treadmill Running Ability and Quantitative Assessment of Gait Dynamics in Young Ts65Dn Mice: A Mouse Model for Down Syndrome. Brain Sciences, 13(5), 743. https://doi.org/10.3390/brainsci13050743