

Biological Factors Underpinning Suicidal Behaviour: An Update
 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
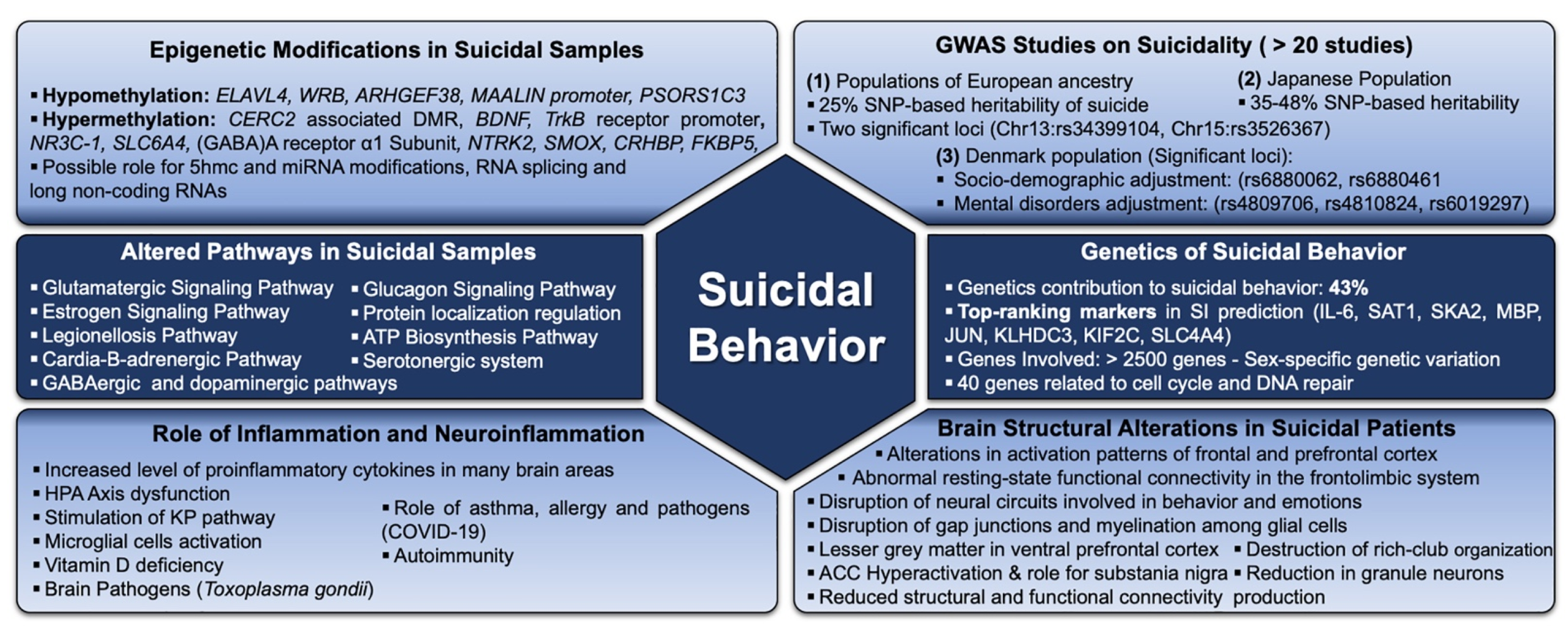
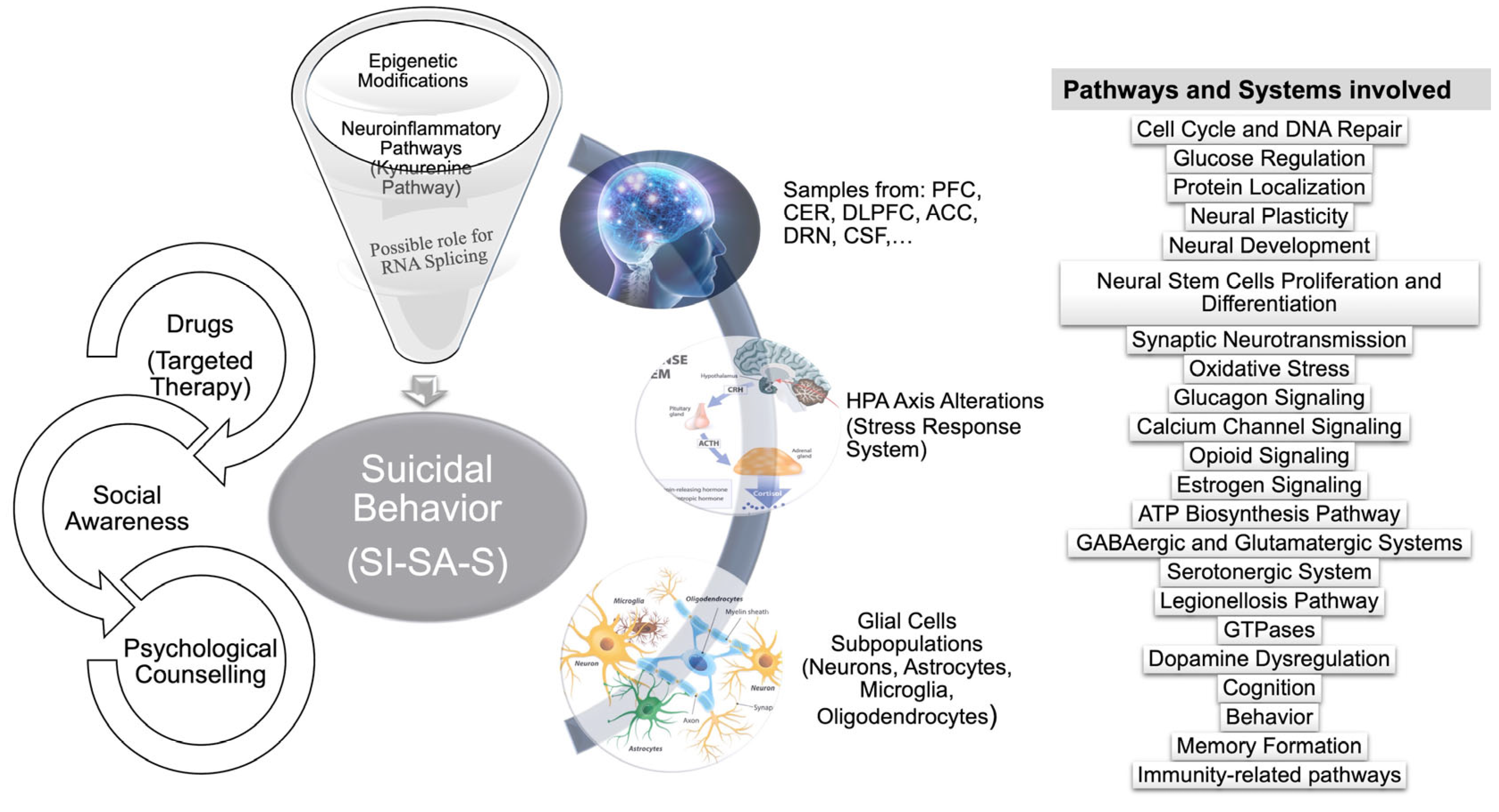
2. Biological Alterations Underlying Suicidal Behaviour
2.1. Genetic/Molecular Factors
2.1.1. Genome-Wide Association Studies
2.1.2. Gene Expression Alteration and SB
2.1.3. Methylation Status of Some Genes Implicated in SB
2.1.4. Other Genetic Endophenotypes Underlying SB
2.2. Neurobiological Underpinning of Suicidal Behaviour
2.2.1. Structural Brain Changes in SB Patients
2.2.2. Other Neurological Alterations
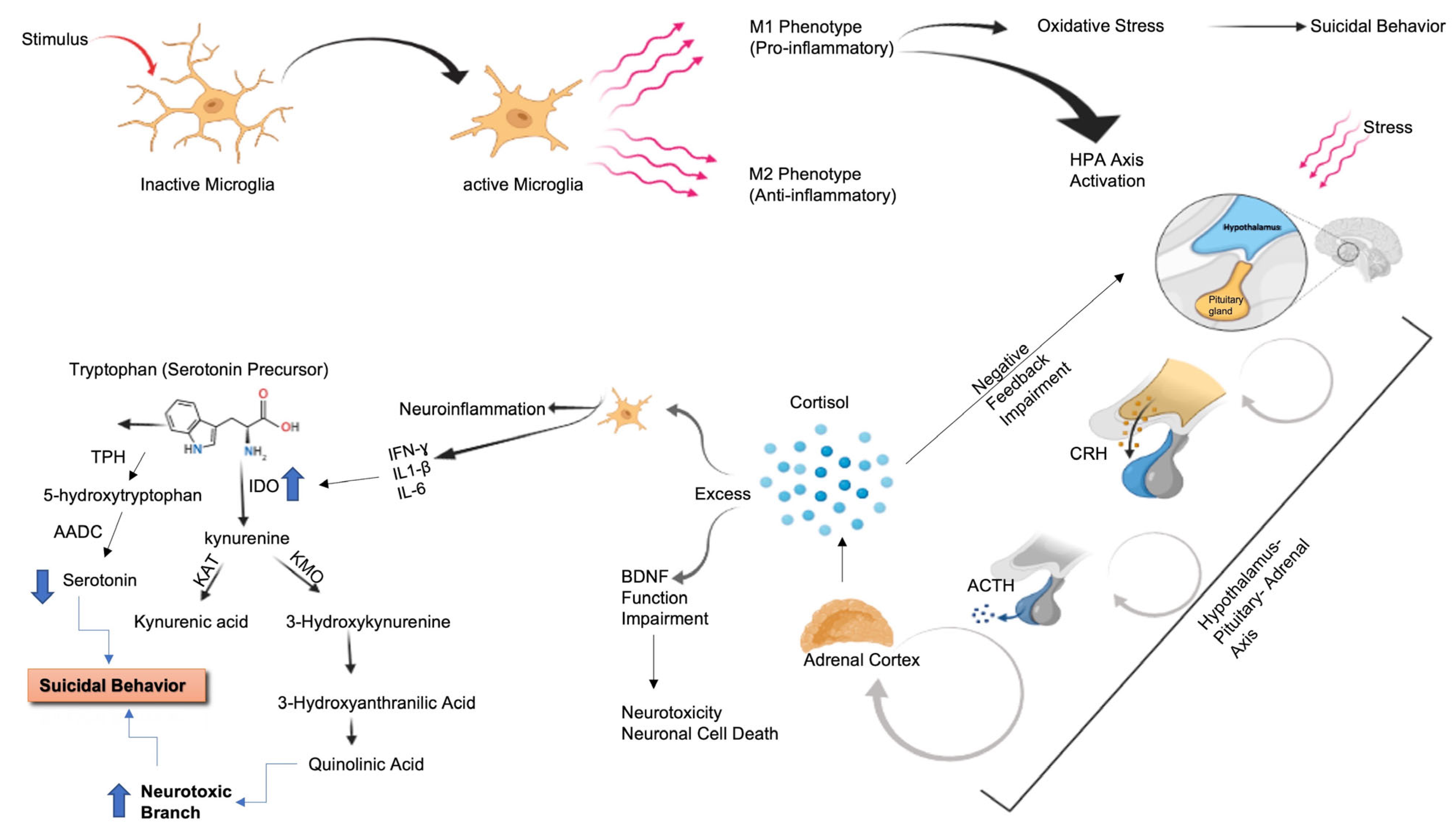
2.3. Inflammation and SB
2.3.1. Inflammatory and Immune Pathways
2.3.2. COVID-19 Pandemic and SB
2.3.3. Neuroinflammation and SB
2.4. Metabolism
3. Pharmacotherapy
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wendt, F.R.; Pathak, G.A.; Levey, D.F.; Nuñez, Y.Z.; Overstreet, C.; Tyrrell, C.; Adhikari, K.; De Angelis, F.; Tylee, D.S.; Goswami, A.; et al. Sex-stratified gene-by-environment genome-wide interaction study of trauma, posttraumatic-stress, and suicidality. Neurobiol. Stress 2021, 14, 100309. [Google Scholar] [CrossRef] [PubMed]
- Turecki, G.; Brent, D.A.; Gunnell, D.; O’Connor, R.C.; Oquendo, M.A.; Pirkis, J.; Stanley, B.H. Suicide and suicide risk. Nat. Rev. Dis. Primer 2019, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- Chikritzhs, T.; Livingston, M. Alcohol and the Risk of Injury. Nutrients 2021, 13, 2777. [Google Scholar] [CrossRef] [PubMed]
- Isometsä, E. Suicidal Behaviour in Mood Disorders—Who, When, and Why? Can. J. Psychiatry Rev. Can. Psychiatr. 2014, 59, 120–130. [Google Scholar] [CrossRef]
- Borentain, S.; Nash, A.I.; Dayal, R.; DiBernardo, A. Patient-reported outcomes in major depressive disorder with suicidal ideation: A real-world data analysis using PatientsLikeMe platform. BMC Psychiatry 2020, 20, 384. [Google Scholar] [CrossRef]
- Lopes, F.L.; Zhu, K.; Purves, K.L.; Song, C.; Ahn, K.; Hou, L.; Akula, N.; Kassem, L.; Bergen, S.E.; Landen, M.; et al. Polygenic risk for anxiety influences anxiety comorbidity and suicidal behavior in bipolar disorder. Transl. Psychiatry 2020, 10, 298. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Kong, X.; Wang, Z.; Zhu, M.; Ren, Y.; Dong, H.; Fang, Y.; Wang, J. Subsequent risk of suicide among 9,300,812 cancer survivors in US: A population-based cohort study covering 40 years of data. EClinicalMedicine 2022, 44, 101295. [Google Scholar] [CrossRef]
- Daray, F.M.; Mann, J.J.; Sublette, M.E. How lipids may affect risk for suicidal behavior. J. Psychiatr. Res. 2018, 104, 16–23. [Google Scholar] [CrossRef]
- Doran, C.M.; Kinchin, I. Economic and epidemiological impact of youth suicide in countries with the highest human development index. PLoS ONE 2020, 15, e0232940. [Google Scholar] [CrossRef]
- DiBlasi, E.; Kang, J.; Docherty, A.R. Genetic contributions to suicidal thoughts and behaviors. Psychol. Med. 2021, 51, 2148–2155. [Google Scholar] [CrossRef]
- Carballo, J.J.; Llorente, C.; Kehrmann, L.; Flamarique, I.; Zuddas, A.; Purper-Ouakil, D.; Hoekstra, P.J.; Coghill, D.; Schulze, U.M.E.; Dittmann, R.W.; et al. Psychosocial risk factors for suicidality in children and adolescents. Eur. Child Adolesc. Psychiatry 2020, 29, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Zulic-Agramunt, C.M.; Guzmán-Guzmán, I.P.; Delgado-Floody, P.; Saavedra, M.B.C.; De La Fuente, P.G.; Solano, M.M.; Berrios, C.S.; Testor, C.P. Psychosocial Factors and Sociodemographic Characteristics Associated with Suicidality Risk in Chilean Adolescents. Children 2022, 9, 1185. [Google Scholar] [CrossRef] [PubMed]
- Batty, G.D.; Kivimäki, M.; Bell, S.; Gale, C.R.; Shipley, M.; Whitley, E.; Gunnell, D. Psychosocial characteristics as potential predictors of suicide in adults: An overview of the evidence with new results from prospective cohort studies. Transl. Psychiatry 2018, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Sha, Q.; Madaj, Z.; Keaton, S.; Galvis, M.E.; Smart, L.; Krzyzanowski, S.; Fazleabas, A.T.; Leach, R.; Postolache, T.T.; Achtyes, E.D.; et al. Cytokines and tryptophan metabolites can predict depressive symptoms in pregnancy. Transl. Psychiatry 2022, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Pompili, M.; Serafini, G.; Innamorati, M.; Dominici, G.; Ferracuti, S.; Kotzalidis, G.D.; Serra, G.; Girardi, P.; Janiri, L.; Tatarelli, R.; et al. Suicidal Behavior and Alcohol Abuse. Int. J. Environ. Res. Public Health 2010, 7, 1392–1431. [Google Scholar] [CrossRef]
- Du, J.; Zhu, M.; Bao, H.; Li, B.; Dong, Y.; Xiao, C.; Zhang, G.Y.; Henter, I.; Rudorfer, M.; Vitielo, B. The Role of Nutrients in Protecting Mitochondrial Function and Neurotransmitter Signaling: Implications for the Treatment of Depression, PTSD, and Suicidal Behaviors. Crit. Rev. Food Sci. Nutr. 2016, 56, 2560–2578. [Google Scholar] [CrossRef]
- Brisch, R.; Wojtylak, S.; Saniotis, A.; Steiner, J.; Gos, T.; Kumaratilake, J.; Henneberg, M.; Wolf, R. The role of microglia in neuropsychiatric disorders and suicide. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 929–945. [Google Scholar] [CrossRef]
- Policicchio, S.; Washer, S.; Viana, J.; Iatrou, A.; Burrage, J.; Hannon, E.; Turecki, G.; Kaminsky, Z.; Mill, J.; Dempster, E.L.; et al. Genome-wide DNA methylation meta-analysis in the brains of suicide completers. Transl. Psychiatry 2020, 10, 69. [Google Scholar] [CrossRef]
- Cai, Z.; Junus, A.; Chang, Q.; Yip, P.S.F. The lethality of suicide methods: A systematic review and meta-analysis. J. Affect. Disord. 2022, 300, 121–129. [Google Scholar] [CrossRef]
- Elnour, A.A.; Harrison, J. Lethality of suicide methods. Inj. Prev. J. Int. Soc. Child Adolesc. Inj. Prev. 2008, 14, 39–45. [Google Scholar] [CrossRef]
- Miranda-Mendizabal, A.; Castellví, P.; Parés-Badell, O.; Alayo, I.; Almenara, J.; Alonso, I.; Blasco, M.J.; Cebrià, A.; Gabilondo, A.; Gili, M.; et al. Gender differences in suicidal behavior in adolescents and young adults: Systematic review and meta-analysis of longitudinal studies. Int. J. Public Health 2019, 64, 265–283. [Google Scholar] [CrossRef]
- Baharikhoob, P.; Kolla, N.J. Microglial Dysregulation and Suicidality: A Stress-Diathesis Perspective. Front. Psychiatry 2020, 11, 781. [Google Scholar] [CrossRef]
- Berardelli, I.; Serafini, G.; Cortese, N.; Fiaschè, F.; O’Connor, R.C.; Pompili, M. The Involvement of Hypothalamus-Pituitary-Adrenal (HPA) Axis in Suicide Risk. Brain Sci. 2020, 10, E653. [Google Scholar] [CrossRef]
- Comai, S.; Gobbi, G. Translational Research in Suicide: Is It Possible to Study Suicide in Animal Models? In Understanding Suicide: From Diagnosis to Personalized Treatment; Courtet, P., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 177–188. [Google Scholar] [CrossRef]
- Gould, T.D.; Georgiou, P.; Brenner, L.A.; Brundin, L.; Can, A.; Courtet, P.; Donaldson, Z.R.; Dwivedi, Y.; Guillaume, S.; Gottesman, I.I.; et al. Animal models to improve our understanding and treatment of suicidal behavior. Transl. Psychiatry 2017, 7, e1092. [Google Scholar] [CrossRef]
- American Psychiatric Association (Ed.) Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- McGuffin, P.; Perroud, N.; Uher, R.; Butlera, A.; Aitchison, K.; Craig, I.; Lewis, C.; Farmer, A. The genetics of affective disorder and suicide. Eur. Psychiatry 2010, 25, 275–277. [Google Scholar] [CrossRef]
- Ludwig, B.; Kienesberger, K.; Carlberg, L.; Swoboda, P.; Bernegger, A.; Koller, R.; Wang, Q.; Inaner, M.; Zotter, M.; Kapusta, N.D.; et al. Influence of CRHR1 Polymorphisms and Childhood Abuse on Suicide Attempts in Affective Disorders: A GxE Approach. Front. Psychiatry 2018, 9, 165. [Google Scholar] [CrossRef]
- Sokolowski, M.; Wasserman, D. Genetic origins of suicidality? A synopsis of genes in suicidal behaviours, with regard to evidence diversity, disorder specificity and neurodevelopmental brain transcriptomics. Eur. Neuropsychopharmacol. 2020, 37, 1–11. [Google Scholar] [CrossRef]
- Docherty, A.R.; Shabalin, A.A.; DiBlasi, E.; Monson, E.; Mullins, N.; Adkins, D.E.; Bacanu, S.-A.; Bakian, A.V.; Crowell, S.; Chen, D.; et al. Genome-Wide Association Study of Suicide Death and Polygenic Prediction of Clinical Antecedents. Am. J. Psychiatry 2020, 177, 917–927. [Google Scholar] [CrossRef]
- Strawbridge, R.J.; Ward, J.; Ferguson, A.; Graham, N.; Shaw, R.J.; Cullen, B.; Pearsall, R.; Lyall, L.M.; Johnston, K.J.; Niedzwiedz, C.L.; et al. Identification of novel genome-wide associations for suicidality in UK Biobank, genetic correlation with psychiatric disorders and polygenic association with completed suicide. EBioMedicine 2019, 41, 517–525. [Google Scholar] [CrossRef]
- Erlangsen, A.; Appadurai, V.; Wang, Y.; Turecki, G.; Mors, O.; Werge, T.; Mortensen, P.B.; Starnawska, A.; Børglum, A.D.; Schork, A.; et al. Genetics of suicide attempts in individuals with and without mental disorders: A population-based genome-wide association study. Mol. Psychiatry 2020, 25, 2410–2421. [Google Scholar] [CrossRef]
- Otsuka, I.; Akiyama, M.; Shirakawa, O.; Okazaki, S.; Momozawa, Y.; Kamatani, Y.; Izumi, T.; Numata, S.; Takahashi, M.; Boku, S.; et al. Genome-wide association studies identify polygenic effects for completed suicide in the Japanese population. Neuropsychopharmacology 2019, 44, 2119–2124. [Google Scholar] [CrossRef] [PubMed]
- Kimbrel, N.A.; Ashley-Koch, A.E.; Qin, X.J.; Lindquist, J.H.; Garrett, M.E.; Dennis, M.F.; Hair, L.P.; Huffman, J.E.; Jacobson, D.A.; Madduri, R.K.; et al. A genome-wide association study of suicide attempts in the million veterans program identifies evidence of pan-ancestry and ancestry-specific risk loci. Mol. Psychiatry 2022, 27, 2264–2272. [Google Scholar] [CrossRef] [PubMed]
- Mullins, N.; Kang, J.; Campos, A.I.; Coleman, J.R.; Edwards, A.C.; Galfalvy, H.; Levey, D.F.; Lori, A.; Shabalin, A.; Starnawska, A.; et al. Dissecting the Shared Genetic Architecture of Suicide Attempt, Psychiatric Disorders, and Known Risk Factors. Biol. Psychiatry 2022, 91, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Kimbrel, N.A.; Ashley-Koch, A.E.; Qin, X.J.; Lindquist, J.H.; Garrett, M.E.; Dennis, M.F.; Hair, L.P.; Huffman, J.E.; Jacobson, D.A.; Madduri, R.K.; et al. Identification of Novel, Replicable Genetic Risk Loci for Suicidal Thoughts and Behaviors Among US Military Veterans. JAMA Psychiatry 2023, 80, 135–145. [Google Scholar] [CrossRef]
- González-Castro, T.B.; Tovilla-Zárate, C.A.; Genis-Mendoza, A.D.; Juárez-Rojop, I.E.; Nicolini, H.; López-Narváez, M.L.; Martínez-Magaña, J.J. Identification of gene ontology and pathways implicated in suicide behavior: Systematic review and enrichment analysis of GWAS studies. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2019, 180, 320–329. [Google Scholar] [CrossRef]
- Reichman, R.D.; Gaynor, S.C.; Monson, E.T.; Gaine, M.E.; Parsons, M.G.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Targeted sequencing of the LRRTM gene family in suicide attempters with bipolar disorder. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2020, 183, 128–139. [Google Scholar] [CrossRef]
- Gaine, M.E.; Seifuddin, F.; Sabunciyan, S.; Lee, R.S.; Benke, K.S.; Monson, E.T.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Differentially methylated regions in bipolar disorder and suicide. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2019, 180, 496–507. [Google Scholar] [CrossRef]
- Gaynor, S.C.; Monson, E.T.; Gaine, M.E.; Chimenti, M.S.; Reichman, R.D.; Parsons, M.; Oonthonpan, L.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Male-specific association of the 2p25 region with suicide attempt in bipolar disorder. J. Psychiatr. Res. 2020, 121, 151–158. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, L.; Wu, Y.; Lu, Z.; Zhang, S. Genome-wide study of key genes and scoring system as potential noninvasive biomarkers for detection of suicide behavior in major depression disorder. Bioengineered 2020, 11, 1189–1196. [Google Scholar] [CrossRef]
- Zhang, L.; Verwer, R.W.H.; Zhao, J.; Huitinga, I.; Lucassen, P.J.; Swaab, D.F. Changes in glial gene expression in the prefrontal cortex in relation to major depressive disorder, suicide and psychotic features. J. Affect. Disord. 2021, 295, 893–903. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Navarrete, F.; Navarro, G.; Reyes-Resina, I.; Franco, R.; Lanciego, J.L.; Giner, S.; Manzanares, J. Alterations in Gene and Protein Expression of Cannabinoid CB2 and GPR55 Receptors in the Dorsolateral Prefrontal Cortex of Suicide Victims. Neurother. J. Am. Soc. Exp. Neurother. 2018, 15, 796–806. [Google Scholar] [CrossRef]
- McLaughlin, R.J.; Hill, M.N.; Gorzalka, B.B. A critical role for prefrontocortical endocannabinoid signaling in the regulation of stress and emotional behavior. Neurosci. Biobehav. Rev. 2014, 42, 116–131. [Google Scholar] [CrossRef]
- Pantazatos, S.P.; Huang, Y.-Y.; Rosoklija, G.B.; Dwork, A.J.; Arango, V.; Mann, J.J. Whole-transcriptome brain expression and exon-usage profiling in major depression and suicide: Evidence for altered glial, endothelial and ATPase activity. Mol. Psychiatry 2017, 22, 760–773. [Google Scholar] [CrossRef]
- Bokor, J.; Sutori, S.; Torok, D.; Gal, Z.; Eszlari, N.; Gyorik, D.; Baksa, D.; Petschner, P.; Serafini, G.; Pompili, M.; et al. Inflamed Mind: Multiple Genetic Variants of IL6 Influence Suicide Risk Phenotypes in Interaction with Early and Recent Adversities in a Linkage Disequilibrium-Based Clumping Analysis. Front. Psychiatry 2021, 12, 746206. [Google Scholar] [CrossRef]
- Kouter, K.; Zupanc, T.; Videtič Paska, A. Genome-Wide DNA Methylation Patterns in Suicide Victims: Identification of New Candidate Genes. Psychiatr. Danub. 2019, 31, 392–396. [Google Scholar] [CrossRef]
- Furczyk, K.; Schutová, B.; Michel, T.M.; Thome, J.; Büttner, A. The neurobiology of suicide—A Review of post-mortem studies. J. Mol. Psychiatry 2013, 1, 2. [Google Scholar] [CrossRef]
- Krzyżanowska, M.; Steiner, J.; Karnecki, K.; Kaliszan, M.; Brisch, R.; Wiergowski, M.; Braun, K.; Jankowski, Z.; Gos, T. Decreased ribosomal DNA transcription in dorsal raphe nucleus neurons differentiates between suicidal and non-suicidal death. Eur. Arch. Psychiatry Clin. Neurosci. 2016, 266, 217–224. [Google Scholar] [CrossRef]
- Keller, S.; Sarchiapone, M.; Zarrilli, F.; Videtič, A.; Ferraro, A.; Carli, V.; Sacchetti, S.; Lembo, F.; Angiolillo, A.; Jovanovic, N.; et al. Increased BDNF promoter methylation in the Wernicke area of suicide subjects. Arch. Gen. Psychiatry 2010, 67, 258–267. [Google Scholar] [CrossRef]
- Kang, H.-J.; Lee, H.; Kim, W.; Kim, W.; Shin, S.; Kim, T.; Park, S.; Cho, H.; Han, S.; Lyoo, I.K.; et al. Association of SLC6A4 methylation with long-term outcomes after stroke: Focus on the interaction with suicidal ideation. Sci. Rep. 2021, 11, 2710. [Google Scholar] [CrossRef]
- Labonté, B.; Abdallah, K.; Maussion, G.; Yerko, V.; Yang, J.; Bittar, T.; Quessy, F.; Golden, S.A.; Navarro, L.; Checknita, D.; et al. Regulation of impulsive and aggressive behaviours by a novel lncRNA. Mol. Psychiatry 2021, 26, 3751–3764. [Google Scholar] [CrossRef]
- Gross, J.A.; Pacis, A.; Chen, G.G.; Drupals, M.; Lutz, P.-E.; Barreiro, L.B.; Turecki, G. Gene-body 5-hydroxymethylation is associated with gene expression changes in the prefrontal cortex of depressed individuals. Transl. Psychiatry 2017, 7, e1119. [Google Scholar] [CrossRef] [PubMed]
- Bach, H.; Arango, V. Neuroanatomy of Serotonergic Abnormalities in Suicide. In The Neurobiological Basis of Suicide; Dwivedi, Y., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2012. Available online: http://www.ncbi.nlm.nih.gov/books/NBK107204/ (accessed on 18 September 2012).
- Salas-Magaña, M.; Tovilla-Zárate, C.A.; González-Castro, T.B.; Juárez-Rojop, I.E.; López-Narváez, M.L.; Rodríguez-Pérez, J.M.; Bello, J.R. Decrease in brain-derived neurotrophic factor at plasma level but not in serum concentrations in suicide behavior: A systematic review and meta-analysis. Brain Behav. 2017, 7, e00706. [Google Scholar] [CrossRef] [PubMed]
- Orsolini, L.; Latini, R.; Pompili, M.; Serafini, G.; Volpe, U.; Vellante, F.; Fornaro, M.; Valchera, A.; Tomasetti, C.; Fraticelli, S.; et al. Understanding the Complex of Suicide in Depression: From Research to Clinics. Psychiatry Investig. 2020, 17, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Abu-Akel, A.; Baxendale, L.; Mohr, C.; Sullivan, S. The association between schizotypal traits and social functioning in adolescents from the general population. Psych. Res. 2018, 270, 895–900. [Google Scholar] [CrossRef]
- Stanley, B.; Itzhaky, I.; Oquendo, M.A. Identifying Neurobiological Underpinnings of Two Suicidal Subtypes. J. Psychiatry Brain Sci. 2021, 6, e210016. [Google Scholar]
- Oquendo, M.A.; Mann, J.J. Neuroimaging findings in major depression, suicidal behavior and aggression. Clin. Neurosci. Res. 2001, 1, 377–380. [Google Scholar] [CrossRef]
- Johnston, J.A.; Wang, F.; Liu, J.; Blond, B.N.; Wallace, A.; Liu, J.; Spencer, L.; Cox Lippard, E.T.; Purves, K.L.; Landeros-Weisenberger, A.; et al. Multimodal Neuroimaging of Frontolimbic Structure and Function Associated with Suicide Attempts in Adolescents and Young Adults with Bipolar Disorder. Am. J. Psychiatry 2017, 174, 667–675. [Google Scholar] [CrossRef]
- Kim, D.J.; Bartlett, E.A.; DeLorenzo, C.; Parsey, R.V.; Kilts, C.; Cáceda, R. Examination of structural brain changes in recent suicidal behavior. Psychiatry Res. Neuroimaging 2021, 307, 111216. [Google Scholar] [CrossRef]
- Zhu, R.; Tian, S.; Wang, H.; Jiang, H.; Wang, X.; Shao, J.; Wang, Q.; Yan, R.; Tao, S.; Liu, H.; et al. Discriminating Suicide Attempters and Predicting Suicide Risk Using Altered Frontolimbic Resting-State Functional Connectivity in Patients With Bipolar II Disorder. Front. Psychiatry 2020, 11, 597770. Available online: https://www.frontiersin.org/articles/10.3389/fpsyt.2020.597770 (accessed on 18 September 2022).
- Costanza, A.; Radomska, M.; Bondolfi, G.; Zenga, F.; Amerio, A.; Aguglia, A.; Serafini, G.; Amore, M.; Berardelli, I.; Pompili, M.; et al. Suicidality Associated with Deep Brain Stimulation in Extrapyramidal Diseases: A Critical Review and Hypotheses on Neuroanatomical and Neuroimmune Mechanisms. Front. Integr. Neurosci. 2021, 15, 632249. [Google Scholar] [CrossRef]
- Liu, X.; He, C.; Fan, D.; Zang, F.; Zhu, Y.; Zhang, H.; Zhang, Z.; Zhang, H.; Xie, C. Alterations of core structural network connectome associated with suicidal ideation in major depressive disorder patients. Transl. Psychiatry 2021, 11, 243. [Google Scholar] [CrossRef]
- Jia, Z.; Wang, Y.; Huang, X.; Kuang, W.; Wu, Q.; Lui, S.; Sweeney, J.A.; Gong, Q. Impaired frontothalamic circuitry in suicidal patients with depression revealed by diffusion tensor imaging at 3.0 T. J. Psychiatry Neurosci. JPN 2014, 39, 170–177. [Google Scholar] [CrossRef]
- Lan, M.J.; Rizk, M.M.; Pantazatos, S.P.; Rubin-Falcone, H.; Miller, J.M.; Sublette, M.E.; Oquendo, M.A.; Keilp, J.G.; Mann, J.J. Resting-state amplitude of low-frequency fluctuation is associated with suicidal ideation. Depress. Anxiety 2019, 36, 433–441. [Google Scholar] [CrossRef]
- Tanti, A.; Lutz, P.-E.; Kim, J.; O’Leary, L.; Théroux, J.-F.; Turecki, G.; Mechawar, N. Evidence of decreased gap junction coupling between astrocytes and oligodendrocytes in the anterior cingulate cortex of depressed suicides. Neuropsychopharmacology 2019, 44, 2099–2111. [Google Scholar] [CrossRef]
- Mahar, I.; Labonte, B.; Yogendran, S.; Isingrini, E.; Perret, L.; Davoli, M.A.; Rachalski, A.; Giros, B.; Turecki, G.; Mechawar, N. Disrupted hippocampal neuregulin-1/ErbB3 signaling and dentate gyrus granule cell alterations in suicide. Transl. Psychiatry 2017, 7, e1161. [Google Scholar] [CrossRef]
- Vegeto, E.; Belcredito, S.; Etteri, S.; Ghisletti, S.; Brusadelli, A.; Meda, C.; Krust, A.; Dupont, S.; Ciana, P.; Chambon, P.; et al. Estrogen receptor-α mediates the brain antiinflammatory activity of estradiol. Proc. Natl. Acad. Sci. USA 2003, 100, 9614–9619. [Google Scholar] [CrossRef]
- Piel, R.B.; Shiferaw, M.T.; Vashisht, A.A.; Marcero, J.R.; Praissman, J.L.; Phillips, J.D.; Wohlschlegel, J.A.; Medlock, A.E. A Novel Role for Progesterone Receptor Membrane Component 1 (PGRMC1): A Partner and Regulator of Ferrochelatase. Biochemistry 2016, 55, 5204–5217. [Google Scholar] [CrossRef]
- Nässberger, L.; Träskman-Bendz, L. Increased soluble interleukin-2 receptor concentrations in suicide attempters. Acta Psychiatr. Scand. 1993, 88, 48–52. [Google Scholar] [CrossRef]
- Engler, H.; Benson, S.; Wegner, A.; Spreitzer, I.; Schedlowski, M.; Elsenbruch, S. Men and women differ in inflammatory and neuroendocrine responses to endotoxin but not in the severity of sickness symptoms. Brain. Behav. Immun. 2016, 52, 18–26. [Google Scholar] [CrossRef]
- Garcia-Oscos, F.; Salgado, H.; Hall, S.; Thomas, F.; Farmer, G.E.; Bermeo, J.; Galindo, L.C.; Ramirez, R.D.; D’Mello, S.; Rose-John, S.; et al. The stress-induced cytokine interleukin-6 decreases the inhibition/excitation ratio in the rat temporal cortex via trans-signaling. Biol. Psychiatry 2012, 71, 574–582. [Google Scholar] [CrossRef]
- Pandey, G.N.; Rizavi, H.S.; Bhaumik, R.; Zhang, H. Chemokines gene expression in the prefrontal cortex of depressed suicide victims and normal control subjects. Brain. Behav. Immun. 2021, 94, 266–273. [Google Scholar] [CrossRef]
- Tonelli, L.H.; Stiller, J.; Rujescu, D.; Giegling, I.; Schneider, B.; Maurer, K.; Schnabel, A.; Möller, H.-J.; Chen, H.H.; Postolache, T.T. Elevated cytokine expression in the orbitofrontal cortex of victims of suicide. Acta Psychiatr. Scand. 2008, 117, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Gadad, B.S.; Vargas-Medrano, J.; Ramos, E.I.; Najera, K.; Fagan, M.; Forero, A. Altered levels of interleukins and neurotrophic growth factors in mood disorders and suicidality: An analysis from periphery to central nervous system. Transl. Psychiatry 2021, 11, 341. [Google Scholar] [CrossRef] [PubMed]
- Ganança, L.; Galfalvy, H.C.; Cisneros-Trujillo, S.; Basseda, Z.; Cooper, T.B.; Ren, X.; Figueira, M.L.; Oquendo, M.A.; Mann, J.J.; Sublette, M.E. Relationships between inflammatory markers and suicide risk status in major depression. J. Psychiatr. Res. 2021, 134, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Nowak, W.; Grendas, L.N.; Sanmarco, L.M.; Estecho, I.G.; Arena, Á.R.; Eberhardt, N.; Rodante, D.E.; Aoki, M.P.; Daray, F.M.; Silva, E.A.C.; et al. Pro-inflammatory monocyte profile in patients with major depressive disorder and suicide behaviour and how ketamine induces anti-inflammatory M2 macrophages by NMDAR and mTOR. EBioMedicine 2019, 50, 290–305. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Jeon, S.-W.; Lim, W.-J.; Oh, K.-S.; Shin, D.-W.; Cho, S.J.; Park, J.-H.; Kim, Y.H.; Shin, Y.-C. Vitamin D deficiency and suicidal ideation: A cross-sectional study of 157,211 healthy adults. J. Psychosom. Res. 2020, 134, 110125. [Google Scholar] [CrossRef]
- Brundin, L.; Bryleva, E.Y.; Thirtamara Rajamani, K. Role of Inflammation in Suicide: From Mechanisms to Treatment. Neuropsychopharmacology 2017, 42, 271–283. [Google Scholar] [CrossRef]
- Vargas, P.A.; Robles, E. Asthma and allergy as risk factors for suicidal behavior among young adults. J. Am. Coll. Health 2019, 67, 97–112. [Google Scholar] [CrossRef]
- Conejero, I.; Nobile, B.; Olié, E.; Courtet, P. How Does COVID-19 Affect the Neurobiology of Suicide? Curr. Psychiatry Rep. 2021, 23, 16. [Google Scholar] [CrossRef]
- Dubé, J.P.; Smith, M.M.; Sherry, S.B.; Hewitt, P.L.; Stewart, S.H. Suicide behaviors during the COVID-19 pandemic: A meta-analysis of 54 studies. Psychiatry Res. 2021, 301, 113998. [Google Scholar] [CrossRef]
- Sher, L. The impact of the COVID-19 pandemic on suicide rates. QJM Mon. J. Assoc. Physicians 2020, 113, 707–712. [Google Scholar] [CrossRef]
- Eyles, E.; Moran, P.; Okolie, C.; Dekel, D.; Macleod-Hall, C.; Webb, R.T.; Schmidt, L.; Knipe, D.; Sinyor, M.; McGuinness, L.A.; et al. Systematic review of the impact of the COVID-19 pandemic on suicidal behaviour amongst health and social care workers across the world. J. Affect. Disord. Rep. 2021, 6, 100271. [Google Scholar] [CrossRef]
- Pirkis, J.; John, A.; Shin, S.; DelPozo-Banos, M.; MRes, V.A.; Analuisa-Aguilar, P.; FRCPsych, L.A.; Arensman, E.; Bantjes, J.; Baran, A.; et al. Suicide trends in the early months of the COVID-19 pandemic: An interrupted time-series analysis of preliminary data from 21 countries. Lancet Psychiatry 2021, 8, 579–588. [Google Scholar] [CrossRef]
- Czeisler, M.É.; Wiley, J.F.; Facer-Childs, E.R.; Robbins, R.; Weaver, M.D.; Barger, L.K.; Czeisler, C.A.; Howard, M.E.; Rajaratnam, S.M.W. Mental health, substance use, and suicidal ideation during a prolonged COVID-19-related lockdown in a region with low SARS-CoV-2 prevalence. J. Psychiatr. Res. 2021, 140, 533–544. [Google Scholar] [CrossRef]
- Tanaka, T.; Okamoto, S. Increase in suicide following an initial decline during the COVID-19 pandemic in Japan. Nat. Hum. Behav. 2021, 5, 229–238. [Google Scholar] [CrossRef]
- Cimino, I.; Kim, H.; Tung, Y.C.L.; Pedersen, K.; Rimmington, D.; Tadross, J.A.; Kohnke, S.N.; Neves-Costa, A.; Barros, A.; Joaquim, S.; et al. Activation of the hypothalamic-pituitary-adrenal axis by exogenous and endogenous GDF15. Proc. Natl. Acad. Sci. USA 2021, 118, e2106868118. [Google Scholar] [CrossRef]
- Trifunovic, S.; Stevanovic, I.; Milosevic, A.; Ristic, N.; Janjic, M.; Bjelobaba, I.; Savic, D.; Bozic, I.; Jakovljevic, M.; Tesovic, K.; et al. The Function of the Hypothalamic-Pituitary-Adrenal Axis during Experimental Autoimmune Encephalomyelitis: Involvement of Oxidative Stress Mediators. Front. Neurosci. 2021, 15, 649485. [Google Scholar] [CrossRef]
- Timmermans, S.; Souffriau, J.; Libert, C. A General Introduction to Glucocorticoid Biology. Front. Immunol. 2019, 10, 1545. [Google Scholar] [CrossRef]
- Ferrari, F.; Villa, R.F. The Neurobiology of Depression: An Integrated Overview from Biological Theories to Clinical Evidence. Mol. Neurobiol. 2017, 54, 4847–4865. [Google Scholar] [CrossRef]
- Leonard, B.E. Inflammation and depression: A causal or coincidental link to the pathophysiology? Acta Neuropsychiatr. 2018, 30, 1–16. [Google Scholar] [CrossRef]
- Harmer, B.; Lee, S.; Duong, T.H.; Saadabadi, A. Suicidal Ideation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: http://www.ncbi.nlm.nih.gov/books/NBK565877/ (accessed on 24 October 2022).
- Stanley, B.; Siever, L.J. The interpersonal dimension of borderline personality disorder: Toward a neuropeptide model. Am. J. Psychiatry 2010, 167, 24–39. [Google Scholar] [CrossRef]
- Bartlett, E.A.; Zanderigo, F.; Stanley, B.; Choo, T.H.; Galfalvy, H.C.; Pantazatos, S.P.; Sublette, M.E.; Miller, J.M.; Oquendo, M.A.; Mann, J.J. In vivo serotonin transporter and 1A receptor binding potential and ecological momentary assessment (EMA) of stress in major depression and suicidal behavior. Eur. Neuropsychopharmacol. 2023, 70, 1–13. [Google Scholar] [CrossRef]
- Brisch, R.; Steiner, J.; Mawrin, C.; Krzyżanowska, M.; Jankowski, Z.; Gos, T. Microglia in the dorsal raphe nucleus plays a potential role in both suicide facilitation and prevention in affective disorders. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Valin, K.L.; Dixon, M.L.; Leavenworth, J.W. The Role of Microglia and Macrophages in CNS Homeostasis, Autoimmunity, and Cancer. J. Immunol. Res. 2017, 2017, 5150678. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, A.; Szebeni, K.; DiPeri, T.P.; Johnson, L.A.; Stockmeier, C.A.; Crawford, J.D.; Chandley, M.J.; Hernandez, L.J.; Burgess, K.C.; Brown, R.W.; et al. Elevated DNA Oxidation and DNA Repair Enzyme Expression in Brain White Matter in Major Depressive Disorder. Int. J. Neuropsychopharmacol. 2017, 20, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Gagne, C.; Piot, A.; Brake, W.G. Depression, Estrogens, and Neuroinflammation: A Preclinical Review of Ketamine Treatment for Mood Disorders in Women. Front. Psychiatry 2021, 12, 797577. [Google Scholar] [CrossRef] [PubMed]
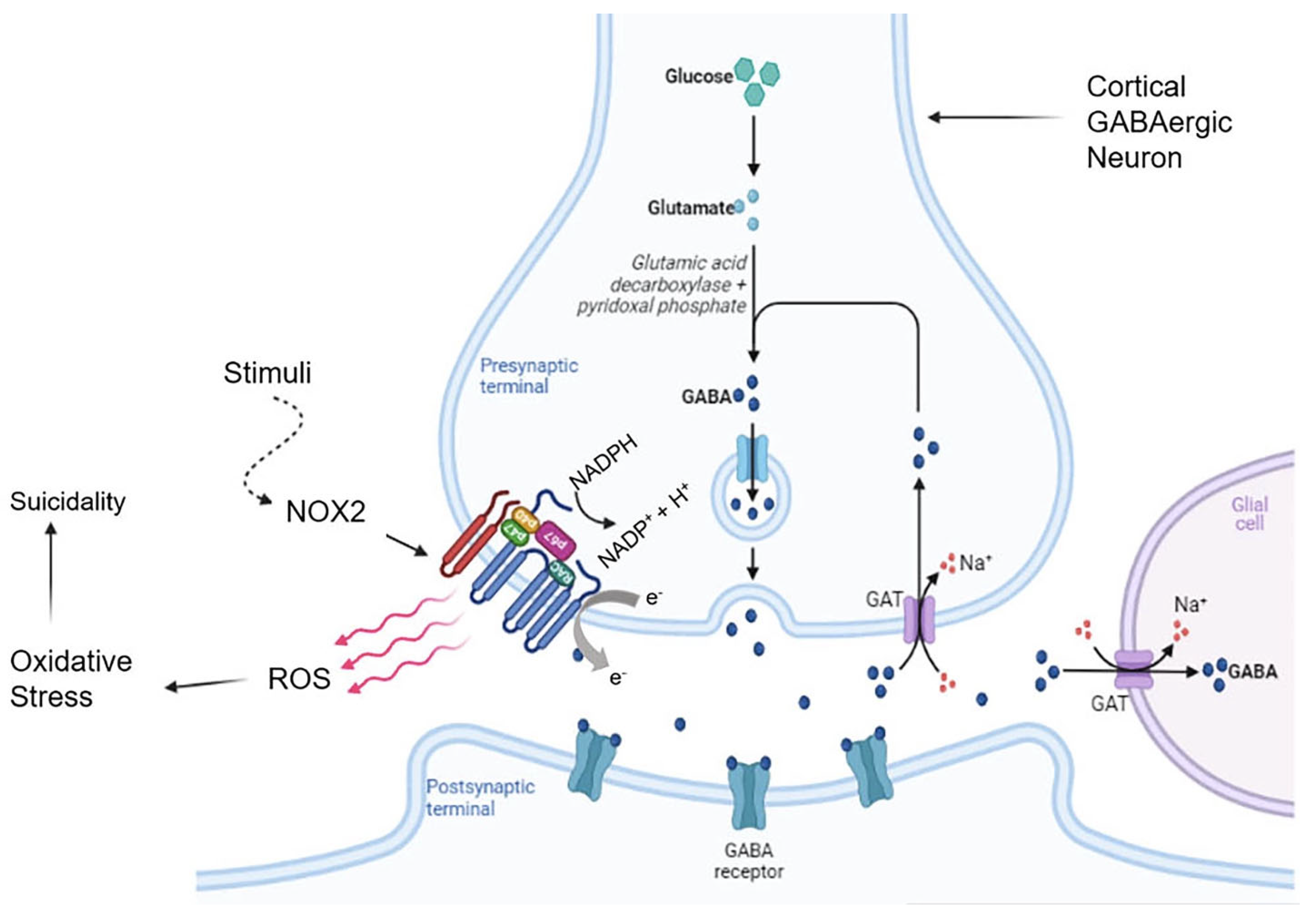
- Schiavone, S.; Neri, M.; Mhillaj, E.; Morgese, M.G.; Cantatore, S.; Bove, M.; Riezzo, I.; Tucci, P.; Pomara, C.; Turillazzi, E.; et al. The NADPH oxidase NOX2 as a novel biomarker for suicidality: Evidence from human post mortem brain samples. Transl. Psychiatry 2016, 6, e813. [Google Scholar] [CrossRef]
- Paribello, P.; Manchia, M.; Bosia, M.; Pinna, F.; Carpiniello, B.; Comai, S. Melatonin and aggressive behavior: A systematic review of the literature on preclinical and clinical evidence. J. Pineal Res. 2022, 72, e12794. [Google Scholar] [CrossRef]
- Manchia, M.; Carpiniello, B.; Valtorta, F.; Comai, S. Serotonin Dysfunction, Aggressive Behavior, and Mental Illness: Exploring the Link Using a Dimensional Approach. ACS Chem. Neurosci. 2017, 8, 961–972. [Google Scholar] [CrossRef]
- Comai, S.; Bertazzo, A.; Brughera, M.; Crotti, S. Tryptophan in health and disease. Adv. Clin. Chem. 2020, 95, 165–218. [Google Scholar] [CrossRef]
- Mándi, Y.; Stone, T.W.; Guillemin, G.J.; Vécsei, L.; Williams, R.O. Editorial: Multiple Implications of the Kynurenine Pathway in Inflammatory Diseases: Diagnostic and Therapeutic Applications. Front. Immunol. 2022, 13, 860867. [Google Scholar] [CrossRef]
- Chen, Y.; Guillemin, G.J. Kynurenine pathway metabolites in humans: Disease and healthy States. Int. J. Tryptophan Res. IJTR 2009, 2, 1–19. [Google Scholar] [CrossRef]
- Schwarcz, R.; Stone, T.W. The kynurenine pathway and the brain: Challenges, controversies and promises. Neuropharmacology 2017, 112, 237–247. [Google Scholar] [CrossRef]
- Postolache, T.T.; Brenner, L.A.; Duncan, E.; Federline, A.; Massa, N.; Barnhart, M.; Deriggs, D.; Yen, P.; Plater, L.Y.; Potocki, E.; et al. The Kynurenine Pathway, Suicide Attempts and Suicide Risk Factors in US Veterans. Biol. Psychiatry 2022, 91, S74. [Google Scholar] [CrossRef]
- Achtyes, E.; Keaton, S.A.; Smart, L.; Burmeister, A.R.; Heilman, P.L.; Krzyzanowski, S.; Nagalla, M.; Guillemin, G.J.; Galvis, M.L.E.; Lim, C.K.; et al. Inflammation and kynurenine pathway dysregulation in post-partum women with severe and suicidal depression. Brain. Behav. Immun. 2020, 83, 239–247. [Google Scholar] [CrossRef]
- Messaoud, A.; Rym, M.; Wahiba, D.; Neffati, F.; Najjar, M.F.; Gobbi, G.; Manchia, M.; Valtorta, F.; Lotfi, G.; Comai, S.; et al. Investigation of the relationship among cortisol, pro-inflammatory cytokines, and the degradation of tryptophan into kynurenine in patients with major depression and suicidal behavior. Curr. Top. Med. Chem. 2022, 22, 2119–2125. [Google Scholar] [CrossRef]
- Messaoud, A.; Mensi, R.; Douki, W.; Naffati, F.; Najjar, M.F.; Gobbi, G.; Valtorta, F.; Gaha, L.; Coami, S. Reduced peripheral availability of tryptophan and increased activation of the kynurenine pathway and cortisol correlate with major depression and suicide. World J. Biol. Psychiatry 2019, 20, 703–711. [Google Scholar] [CrossRef]
- Erhardt, S.; Lim, C.; Linderholm, K.; Janelidze, S.; Lindqvist, D.; Samuelsson, M.; Lundberg, K.; Postolache, T.; Träskman-Bendz, L.; Guillemin, G.; et al. Connecting inflammation with glutamate agonism in suicidality. Neuropsychopharmacology 2013, 38, 743–752. [Google Scholar] [CrossRef]
- Wisłowska-Stanek, A.; Kołosowska, K.; Maciejak, P. Neurobiological Basis of Increased Risk for Suicidal Behaviour. Cells 2021, 10, 2519. [Google Scholar] [CrossRef]
- Bryleva, E.Y.; Brundin, L. Kynurenine pathway metabolites and suicidality. Neuropharmacology 2017, 112, 324–330. [Google Scholar] [CrossRef]
- Serafini, G.; Adavastro, G.; Canepa, G.; Capobianco, L.; Conigliaro, C.; Pittaluga, F.; Belvederi Murri, M.; Valchera, A.; De Berardis, D.; Pompili, M.; et al. Abnormalities in Kynurenine Pathway Metabolism in Treatment-Resistant Depression and Suicidality: A Systematic Review. CNS Neurol. Disord. Drug Targets 2017, 16, 440–453. [Google Scholar] [CrossRef]
- Savitz, J. The kynurenine pathway: A finger in every pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef]
- Pandey, G.N.; Ren, X.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Dwivedi, Y. Brain-derived neurotrophic factor and tyrosine kinase B receptor signalling in post-mortem brain of teenage suicide victims. Int. J. Neuropsychopharmacol. 2008, 11, 1047–1061. [Google Scholar] [CrossRef] [PubMed]
- Juengst, S.B.; Kumar, R.G.; Arenth, P.M.; Wagner, A.K. Exploratory associations with tumor necrosis factor-α, disinhibition and suicidal endorsement after traumatic brain injury. Brain. Behav. Immun. 2014, 41, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Duffy, A.R.; O’Connell, J.R.; Pavlovich, M.; Ryan, K.A.; Lowry, C.A.; Daue, M.; Raheja, U.K.; Brenner, L.A.; Markon, A.O.; Punzalan, C.M.; et al. Toxoplasma gondii Serointensity and Seropositivity: Heritability and Household-Related Associations in the Old Order Amish. Int. J. Environ. Res. Public Health 2019, 16, 3732. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Xu, C.; Zhao, G.; Xie, H. Epigenetic Manipulation of Psychiatric Behavioral Disorders Induced by Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2022, 12, 803502. [Google Scholar] [CrossRef]
- Xiong, W.; Cao, X.; Zeng, Y.; Qin, X.; Zhu, M.; Ren, J.; Wu, Z.; Huang, Q.; Zhang, Y.; Wang, M.; et al. Astrocytic Epoxyeicosatrienoic Acid Signaling in the Medial Prefrontal Cortex Modulates Depressive-like Behaviors. J. Neurosci. 2019, 39, 4606–4623. [Google Scholar] [CrossRef]
- Fond, G.; Loundou, A.; Rabu, C.; Macgregor, A.; Lançon, C.; Brittner, M.; Micoulaud-Franchi, J.-A.; Richieri, R.; Courtet, P.; Abbar, M.; et al. Ketamine administration in depressive disorders: A systematic review and meta-analysis. Psychopharmacology 2014, 231, 3663–3676. [Google Scholar] [CrossRef]
- Witt, K.; Potts, J.; Hubers, A.; Grunebaum, M.F.; Murrough, J.; Loo, C.; Cipriani, A.; Hawton, K. Ketamine for suicidal ideation in adults with psychiatric disorders: A systematic review and meta-analysis of treatment trials. Aust. N. Z. J. Psychiatry 2020, 54, 29–45. [Google Scholar] [CrossRef]
- Gould, T.D.; Zarate, C.A.; Thompson, S.M. Molecular Pharmacology and Neurobiology of Rapid-Acting Antidepressants. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 213–236. [Google Scholar] [CrossRef]
- Bellairs-Walsh, I.; Byrne, S.J.; Bendall, S.; Perry, Y.; Krysinska, K.; Lin, A.; Michail, M.; Lamblin, M.; Li, T.Y.; Hetrick, S.; et al. Working with Young People at Risk of Suicidal Behaviour and Self-Harm: A Qualitative Study of Australian General Practitioners’ Perspectives. Int. J. Environ. Res. Public Health 2021, 18, 12926. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou Chahla, M.N.; Khalil, M.I.; Comai, S.; Brundin, L.; Erhardt, S.; Guillemin, G.J. Biological Factors Underpinning Suicidal Behaviour: An Update. Brain Sci. 2023, 13, 505. https://doi.org/10.3390/brainsci13030505
Abou Chahla MN, Khalil MI, Comai S, Brundin L, Erhardt S, Guillemin GJ. Biological Factors Underpinning Suicidal Behaviour: An Update. Brain Sciences. 2023; 13(3):505. https://doi.org/10.3390/brainsci13030505
Chicago/Turabian StyleAbou Chahla, Maya N., Mahmoud I. Khalil, Stefano Comai, Lena Brundin, Sophie Erhardt, and Gilles J. Guillemin. 2023. "Biological Factors Underpinning Suicidal Behaviour: An Update" Brain Sciences 13, no. 3: 505. https://doi.org/10.3390/brainsci13030505
APA StyleAbou Chahla, M. N., Khalil, M. I., Comai, S., Brundin, L., Erhardt, S., & Guillemin, G. J. (2023). Biological Factors Underpinning Suicidal Behaviour: An Update. Brain Sciences, 13(3), 505. https://doi.org/10.3390/brainsci13030505