

Prefrontal Cortex Hemodynamics and Functional Connectivity Changes during Performance Working Memory Tasks in Older Adults with Sleep Disorders
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Trial Design and Participants
2.2. Diagnostic Criteria
2.2.1. PSQI
2.2.2. MoCA
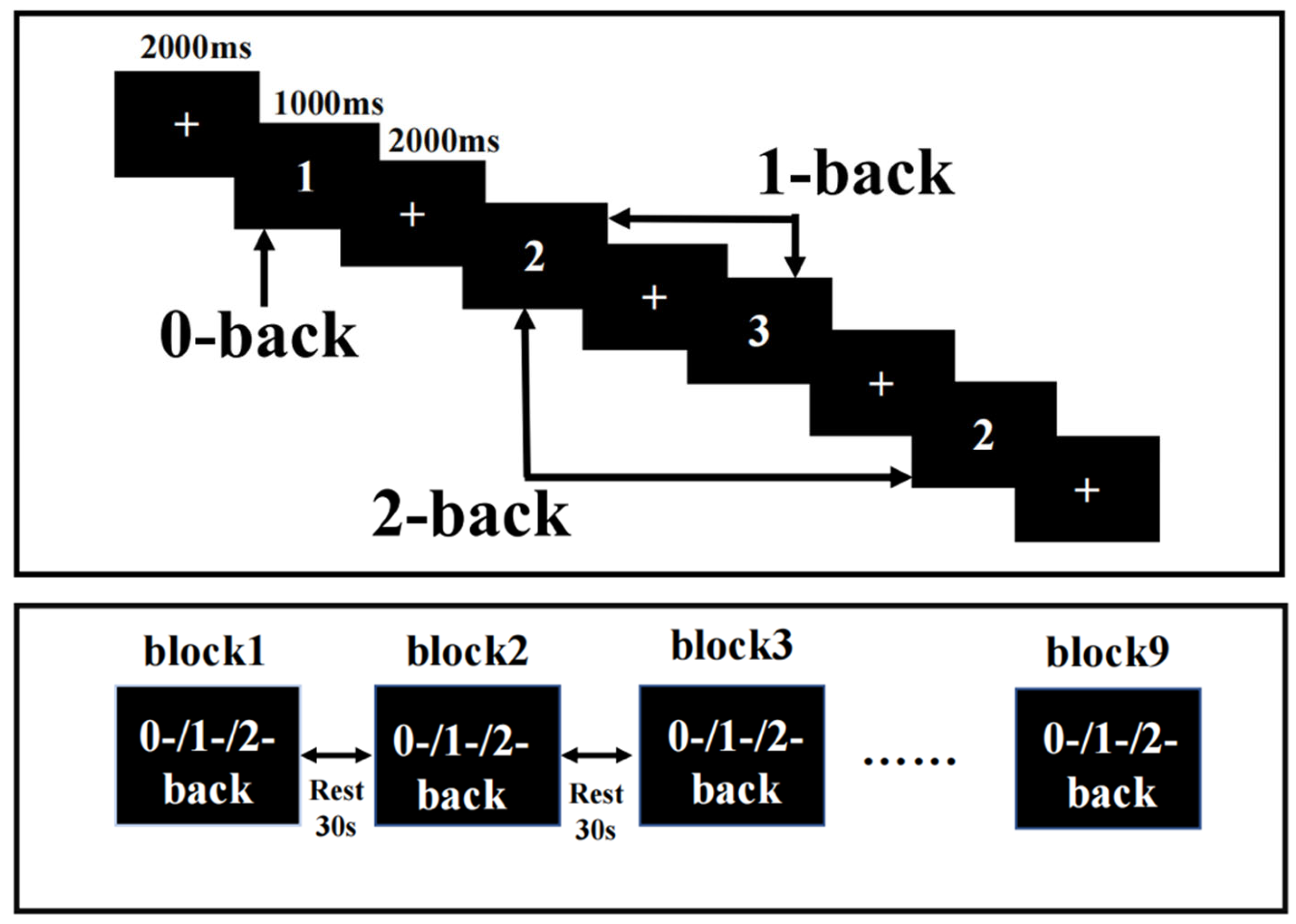
2.3. N-Back Task Design
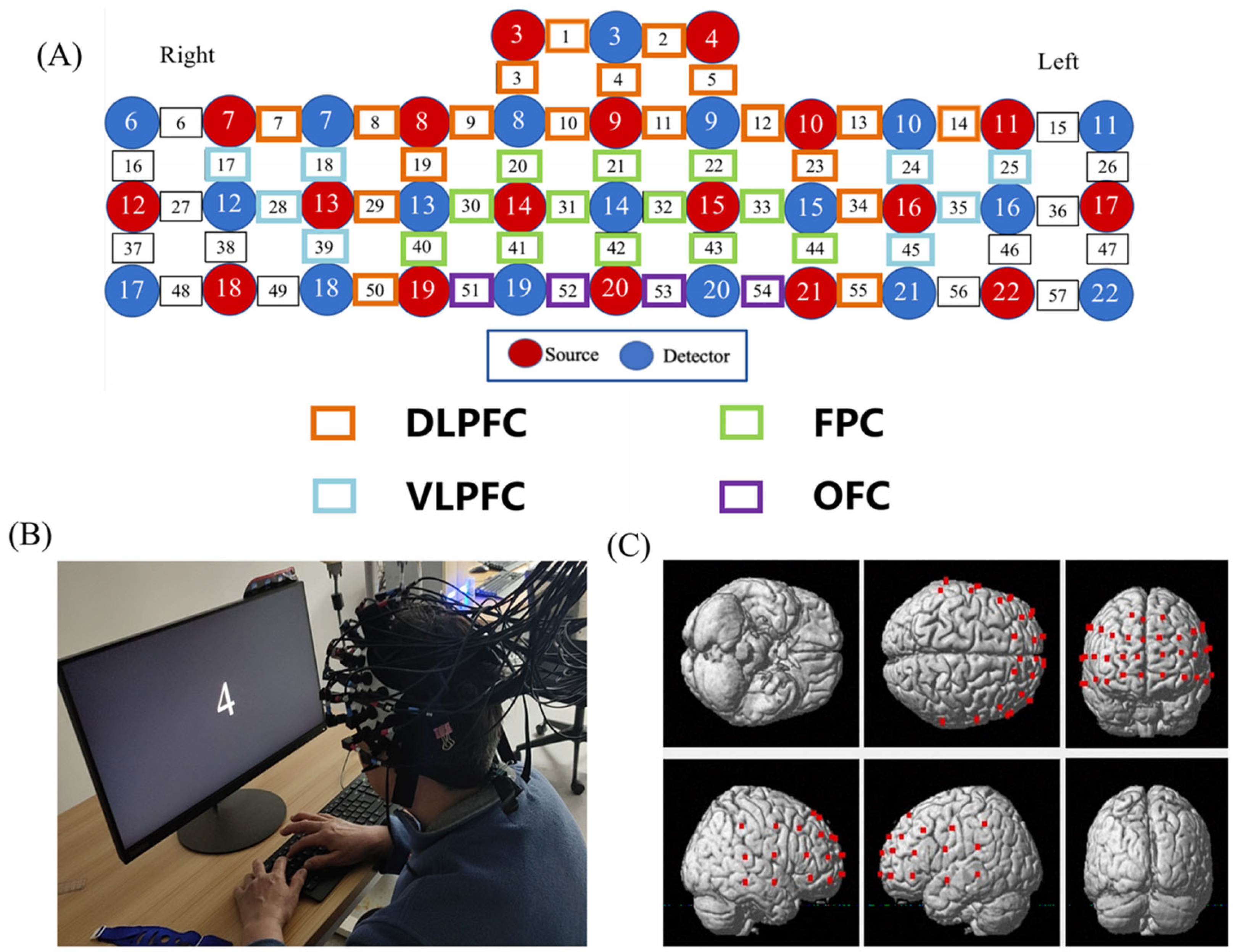
2.4. FNIRS Data Acquisition
2.5. fNIRS Data Analysis
2.6. Sample Sizes
2.7. Statistical Analysis
3. Results
3.1. Demographics
3.2. N-Back Performance
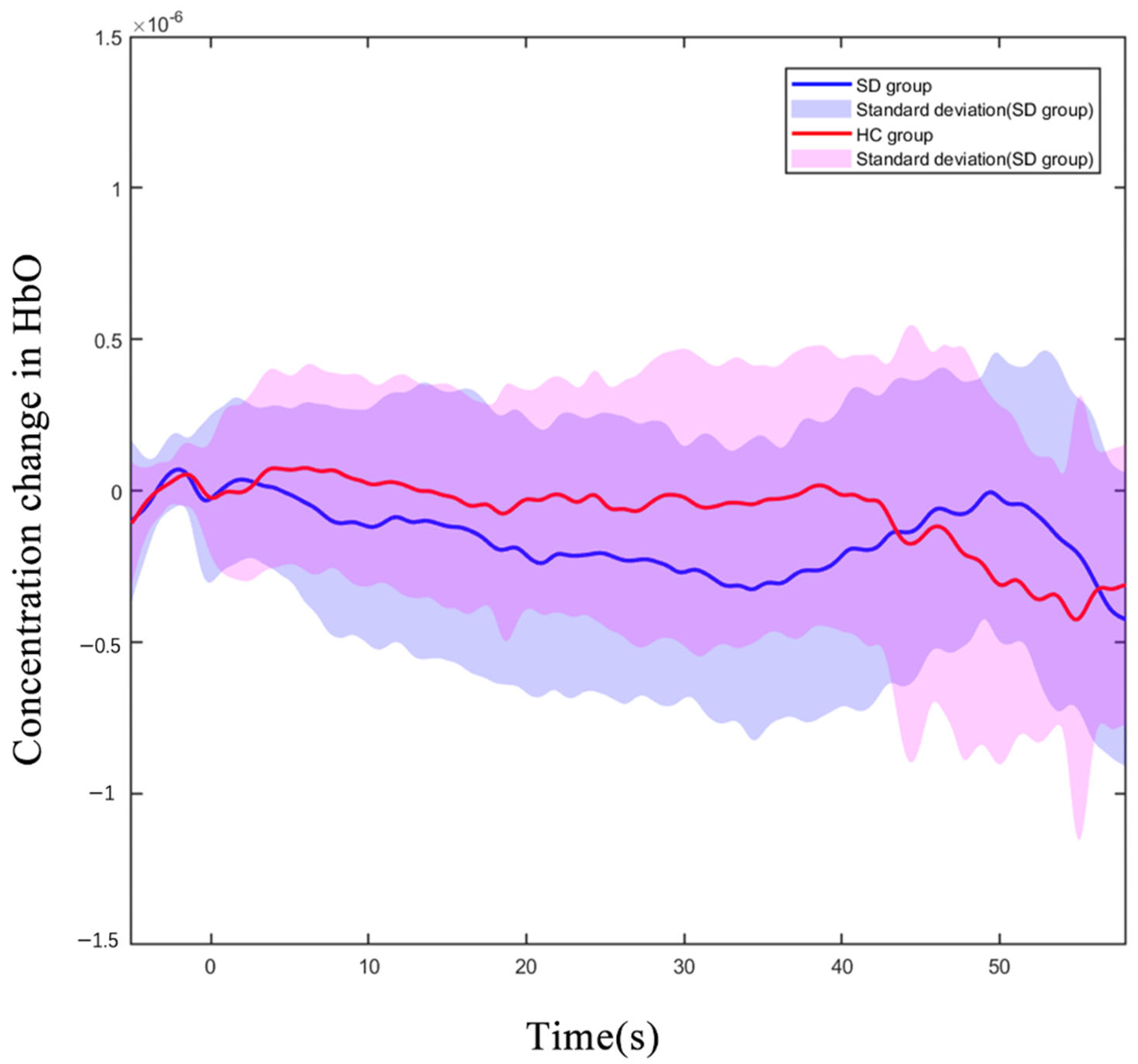
3.3. The PFC’s Hemodynamic Response during N-Back Task
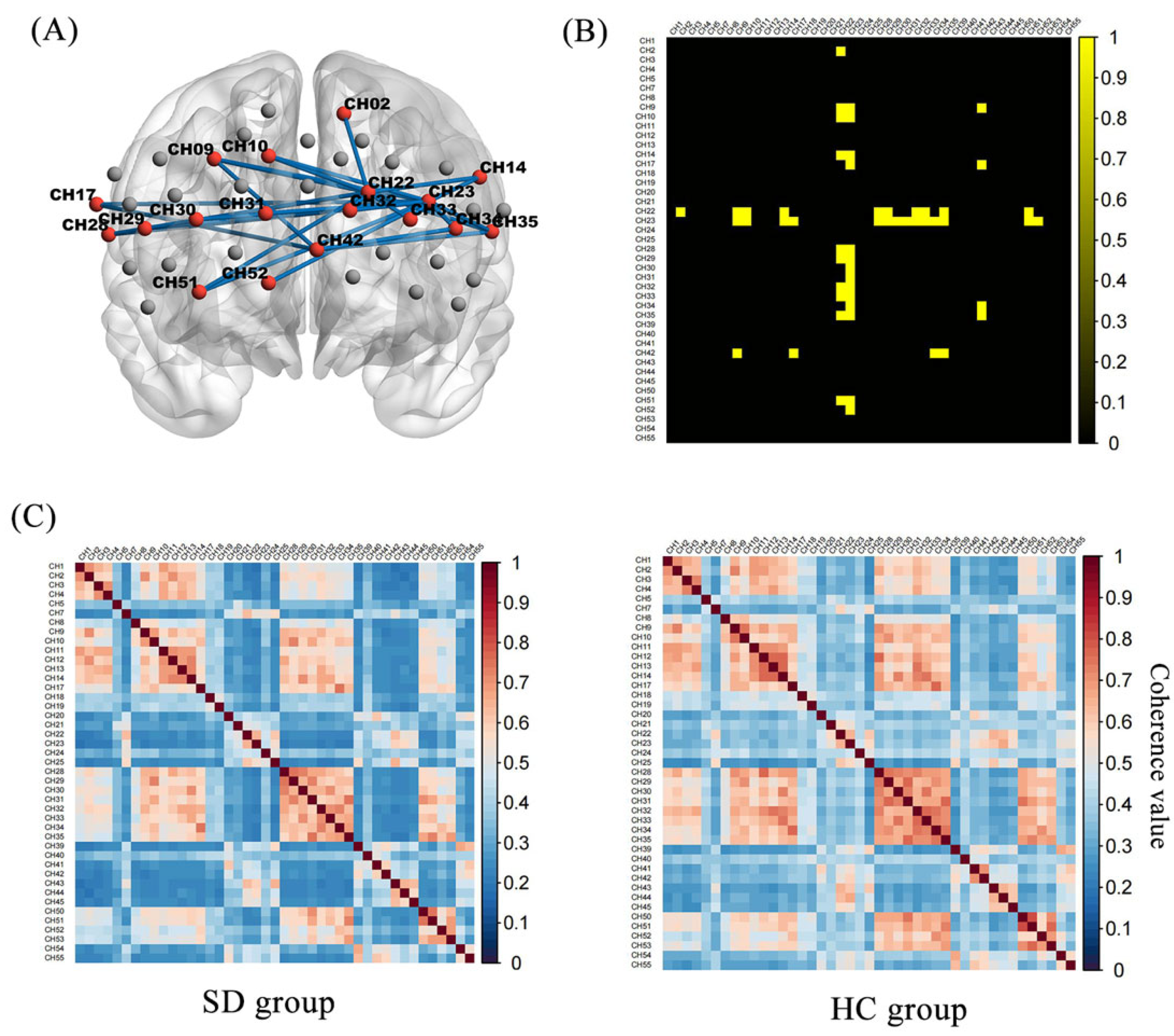
3.4. Functional Connectivity of Prefrontal Areas
4. Discussion
Limitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yaremchuk, K. Sleep Disorders in the Elderly. Clin. Geriatr. Med. 2018, 34, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.E.; Shema, S.J.; Kaplan, G.A.; Strawbridge, W.J. Sleep complaints and depression in an aging cohort: A prospective perspective. Am. J. Psychiatry 2000, 157, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Tan, C.C.; Zou, J.J.; Cao, X.P.; Tan, L. Sleep problems and risk of all-cause cognitive decline or dementia: An updated systematic review and meta-analysis. J.Neurol. Neurosurg. Psychiatry 2020, 91, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Wardle-Pinkston, S.; Slavish, D.C.; Taylor, D.J. Insomnia and cognitive performance: A systematic review and meta-analysis. Sleep Med. Rev. 2019, 48, 101205. [Google Scholar] [CrossRef] [PubMed]
- Fortier-Brochu, E.; Beaulieu-Bonneau, S.; Ivers, H.; Morin, C.M. Insomnia and daytime cognitive performance: A meta-analysis. Sleep Med. Rev. 2012, 16, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Baddeley, A. Working memory: Theories, models, and controversies. Annu. Rev. Psychol. 2012, 63, 1–29. [Google Scholar] [CrossRef]
- Naëgelé, B.; Launois, S.H.; Mazza, S.; Feuerstein, C.; Pépin, J.L.; Lévy, P. Which memory processes are affected in patients with obstructive sleep apnea? An evaluation of 3 types of memory. Sleep 2006, 29, 533–544. [Google Scholar] [CrossRef]
- Turner, T.H.; Drummond, S.P.; Salamat, J.S.; Brown, G.G. Effects of 42 h of total sleep deprivation on component processes of verbal working memory. Neuropsychology 2007, 21, 787–795. [Google Scholar] [CrossRef]
- Okuda, M.; Noda, A.; Iwamoto, K.; Nakashima, H.; Takeda, K.; Miyata, S.; Yasuma, F.; Ozaki, N.; Shimouchi, A. Effects of long sleep time and irregular sleep-wake rhythm on cognitive function in older people. Sci. Rep. 2021, 11, 7039. [Google Scholar] [CrossRef] [PubMed]
- Shekleton, J.A.; Flynn-Evans, E.E.; Miller, B.; Epstein, L.J.; Kirsch, D.; Brogna, L.A.; Burke, L.M.; Bremer, E.; Murray, J.M.; Gehrman, P.; et al. Neurobehavioral performance impairment in insomnia: Relationships with self-reported sleep and daytime functioning. Sleep 2014, 37, 107–116. [Google Scholar] [CrossRef] [PubMed]
- D’Esposito, M.; Postle, B.R.; Ballard, D.; Lease, J. Maintenance versus manipulation of information held in working memory: An event-related fMRI study. Brain Cogn. 1999, 41, 66–86. [Google Scholar] [CrossRef]
- Wu, G.; Wang, Y.; Mwansisya, T.E.; Pu, W.; Zhang, H.; Liu, C.; Yang, Q.; Chen, E.Y.; Xue, Z.; Liu, Z.; et al. Effective connectivity of the posterior cingulate and medial prefrontal cortices relates to working memory impairment in schizophrenic and bipolar patients. Schizophr. Res. 2014, 158, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Grandelis, A.; Maddock, R.J. Dorsolateral Prefrontal Cortex GABA Concentration in Humans Predicts Working Memory Load Processing Capacity. J. Neurosci. 2016, 36, 11788–11794. [Google Scholar] [CrossRef] [PubMed]
- Owen, A.M.; McMillan, K.M.; Laird, A.R.; Bullmore, E. N-back working memory paradigm: A meta-analysis of normative functional neuroimaging studies. Hum. Brain Mapp. 2005, 25, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Barbey, A.K.; Koenigs, M.; Grafman, J. Orbitofrontal contributions to human working memory. Cereb. Cortex 2011, 21, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Hampson, M.; Driesen, N.R.; Skudlarski, P.; Gore, J.C.; Constable, R.T. Brain connectivity related to working memory performance. J. Neurosci. 2006, 26, 13338–13343. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Jiang, Z.; Jiang, B.; McClure, M.A.; Mu, Q. High-Frequency Repetitive Transcranial Magnetic Stimulation Could Improve Impaired Working Memory Induced by Sleep Deprivation. Neural Plast. 2019, 2019, 7030286. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, E.; Zhang, H.; Dou, S.; Liu, L.; Tong, L.; Lei, Y.; Wang, M.; Xu, J.; Shi, D.; et al. Functional connectivity changes between parietal and prefrontal cortices in primary insomnia patients: Evidence from resting-state fMRI. Eur. J. Med. Res. 2014, 19, 32. [Google Scholar] [CrossRef]
- Drummond, S.P.; Walker, M.; Almklov, E.; Campos, M.; Anderson, D.E.; Straus, L.D. Neural correlates of working memory performance in primary insomnia. Sleep 2013, 36, 1307–1316. [Google Scholar] [CrossRef]
- Blum, L.; Hofmann, A.; Rosenbaum, D.; Elshehabi, M.; Suenkel, U.; Fallgatter, A.J.; Ehlis, A.C.; Metzger, F.G. Effects of aging on functional connectivity in a neurodegenerative risk cohort: Resting state versus task measurement using near-infrared spectroscopy. Sci. Rep. 2022, 12, 11262. [Google Scholar] [CrossRef]
- Canessa, N.; Castronovo, V.; Cappa, S.F.; Marelli, S.; Iadanza, A.; Falini, A.; Ferini-Strambi, L. Sleep apnea: Altered brain connectivity underlying a working-memory challenge. NeuroImage Clin. 2018, 19, 56–65. [Google Scholar] [CrossRef]
- Chen, W.L.; Wagner, J.; Heugel, N.; Sugar, J.; Lee, Y.W.; Conant, L.; Malloy, M.; Heffernan, J.; Quirk, B.; Zinos, A.; et al. Functional Near-Infrared Spectroscopy and Its Clinical Application in the Field of Neuroscience: Advances and Future Directions. Front. Neurosci. 2020, 14, 724. [Google Scholar] [CrossRef]
- Buysse, D.J.; Reynolds, C.F., 3rd; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.C.; Tak, S.; Jang, K.E.; Jung, J.; Jang, J. NIRS-SPM: Statistical parametric mapping for near-infrared spectroscopy. Neuroimage 2009, 44, 428–447. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhu, W.; Wen, X.; Zang, Z.; Da, Y.; Lu, J. Different sensorimotor mechanism in fast and slow progression amyotrophic lateral sclerosis. Hum. Brain Mapp. 2022, 43, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, Y. Functional near-infrared optical imaging: Utility and limitations in human brain mapping. Psychophysiology 2003, 40, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Olaithe, M.; Ree, M.; McArdle, N.; Donaldson, S.; Pushpanathan, M.; Eastwood, P.R.; Bucks, R.S. Cognitive Dysfunction in Insomnia Phenotypes: Further Evidence for Different Disorders. Front. Psychiatry 2021, 12, 688672. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Earlbaum Associates: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Kerby, D.S. The Simple Difference Formula: An Approach to Teaching Nonparametric Correlation. Compr. Psychol. 2014, 3, 1078–1092. [Google Scholar] [CrossRef]
- Callicott, J.H.; Bertolino, A.; Mattay, V.S.; Langheim, F.J.; Duyn, J.; Coppola, R.; Goldberg, T.E.; Weinberger, D.R. Physiological dysfunction of the dorsolateral prefrontal cortex in schizophrenia revisited. Cereb. Cortex 2000, 10, 1078–1092. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Iwamoto, K.; Kawano, N.; Noda, Y.; Ozaki, N.; Noda, A. Differential effects of physical activity and sleep duration on cognitive function in young adults. J. Sport Health Sci. 2018, 7, 227–236. [Google Scholar] [CrossRef]
- Gelir, E.; Başaran, C.; Bayrak, S.; Yağcıoğlu, S.; Budak, M.T.; Fırat, H.; Ungan, P. Electrophysiological assessment of the effects of obstructive sleep apnea on cognition. PLoS ONE 2014, 9, e90647. [Google Scholar] [CrossRef] [PubMed]
- Porkka-Heiskanen, T.; Zitting, K.M.; Wigren, H.K. Sleep, its regulation and possible mechanisms of sleep disturbances. Acta Physiol. 2013, 208, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Nikolin, S.; Tan, Y.Y.; Schwaab, A.; Moffa, A.; Loo, C.K.; Martin, D. An investigation of working memory deficits in depression using the n-back task: A systematic review and meta-analysis. J. Affect. Disord. 2021, 284, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hørlyck, L.D.; Bisby, J.A.; King, J.A.; Burgess, N. Wakeful rest compared to vigilance reduces intrusive but not deliberate memory for traumatic videos. Sci. Rep. 2019, 9, 13403. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Noda, A.; Yasuma, F.; Matsubara, Y.; Miyata, S.; Iwamoto, K.; Miyazaki, S.; Ozaki, N. Effects of sleep-disordered breathing and hypertension on cognitive function in elderly adults. Clin. Exp. Hypertens 2020, 42, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Fishburn, F.A.; Norr, M.E.; Medvedev, A.V.; Vaidya, C.J. Sensitivity of fNIRS to cognitive state and load. Front. Hum. Neurosci. 2014, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Jacola, L.M.; Willard, V.W.; Ashford, J.M.; Ogg, R.J.; Scoggins, M.A.; Jones, M.M.; Wu, S.; Conklin, H.M. Clinical utility of the N-back task in functional neuroimaging studies of working memory. J. Clin. Exp. Neuropsychol. 2014, 36, 875–886. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, S.; Lu, J.; Lei, Y.; Li, H. Social exclusion increases the executive function of attention networks. Sci. Rep. 2021, 11, 9494. [Google Scholar] [CrossRef]
- Hsiao, K.; Noble, C.; Pitman, W.; Yadav, N.; Kumar, S.; Keele, G.R.; Terceros, A.; Kanke, M.; Conniff, T.; Cheleuitte-Nieves, C.; et al. A Thalamic Orphan Receptor Drives Variability in Short-Term Memory. Cell 2020, 183, 522–536.e19. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, R.; Venkatesan, S.; Binko, M.; Bang, J.Y.; Cajanding, J.D.; Briggs, C.; Sargin, D.; Imayoshi, I.; Lambe, E.K.; Kim, J.C. Cholecystokinin-Expressing Interneurons of the Medial Prefrontal Cortex Mediate Working Memory Retrieval. J. Neurosci. 2020, 40, 2314–2331. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.A.; Parsa, K.; Geiger, S.; Zaragoza, R.; Kermanian, R.; Miguel, H.; Dashtestani, H.; Chowdhry, F.A.; Smith, E.; Aram, S.; et al. Exploring the role of task performance and learning style on prefrontal hemodynamics during a working memory task. PLoS ONE 2018, 13, e0198257. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, L.; Wang, E.; Zhang, H.; Dou, S.; Tong, L.; Cheng, J.; Chen, C.; Shi, D. Abnormal Neural Network of Primary Insomnia: Evidence from Spatial Working Memory Task fMRI. Eur. Neurol. 2016, 75, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.K.; Wu, J.M.; Zhou, G.; Zhu, F.; Zhang, Q.; Yang, X.J.; Qin, Z.; Zhao, N.; Chen, H.; Zhang, Z.J. Antidepressant Monotherapy and Combination Therapy with Acupuncture in Depressed Patients: A Resting-State Functional Near-Infrared Spectroscopy (fNIRS) Study. Neurotherapeutics 2021, 18, 2651–2663. [Google Scholar] [CrossRef]
- Thomas, R.J.; Rosen, B.R.; Stern, C.E.; Weiss, J.W.; Kwong, K.K. Functional imaging of working memory in obstructive sleep-disordered breathing. J. Appl. Physiol. 2005, 98, 2226–2234. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Nahas, Z.; Johnson, K.A.; Yamanaka, K.; Mishory, A.; Koola, J.; Hill, S.; Horner, M.D.; Bohning, D.E.; George, M.S. Decreased cortical response to verbal working memory following sleep deprivation. Sleep 2005, 28, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Shigaeff, N. Sleep disorders and attention: A systematic review. Arq. Neuro-Psiquiatr. 2022, 80, 530–538. [Google Scholar] [CrossRef]
- Konishi, M.; McLaren, D.G.; Engen, H.; Smallwood, J. Shaped by the Past: The Default Mode Network Supports Cognition that Is Independent of Immediate Perceptual Input. PLoS ONE 2015, 10, e0132209. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, F.; Wang, H.; Yang, A.; Gao, C.; Li, Z.; Li, X. Connectivity properties in the prefrontal cortex during working memory: A near-infrared spectroscopy study. J. Biomed. Opt. 2019, 24, 1–7. [Google Scholar] [CrossRef]
- Friston, K.J. Functional and effective connectivity in neuroimaging: A synthesis. Hum. Brain Mapp. 1994, 2, 56–78. [Google Scholar] [CrossRef]
- Otsuka, Y.; Osaka, N. An fMRI study of verbal working memory-Effects of aging on ACC-PFC network connectivity. Jpn. J. Physiol. Psychol. Psychophysiol. 2005, 23, 11–18. [Google Scholar] [CrossRef]
- Hocke, L.M.; Duszynski, C.C.; Debert, C.T.; Dleikan, D.; Dunn, J.F. Reduced Functional Connectivity in Adults with Persistent Post-Concussion Symptoms: A Functional Near-Infrared Spectroscopy Study. J. Neurotrauma 2018, 35, 1224–1232. [Google Scholar] [CrossRef]
- Schmidt, C.C.; Timpert, D.C.; Arend, I.; Vossel, S.; Fink, G.R.; Henik, A.; Weiss, P.H. Control of response interference: Caudate nucleus contributes to selective inhibition. Sci. Rep. 2020, 10, 20977. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Dong, M.; Yin, Y.; Hua, K.; Fu, S.; Jiang, G. Abnormal whole-brain functional connectivity in patients with primary insomnia. Neuropsychiatr. Dis. Treat. 2017, 13, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, J.; Liu, B.X.; Dai, X.J. Altered connection properties of important network hubs may be neural risk factors for individuals with primary insomnia. Sci. Rep. 2018, 8, 5891. [Google Scholar] [CrossRef] [PubMed]
- Verweij, I.M.; Romeijn, N.; Smit, D.J.; Piantoni, G.; Van Someren, E.J.; van der Werf, Y.D. Sleep deprivation leads to a loss of functional connectivity in frontal brain regions. BMC Neurosci. 2014, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Corbo, I.; Casagrande, M. Higher-Level Executive Functions in Healthy Elderly and Mild Cognitive Impairment: A Systematic Review. J. Clin. Med. 2022, 11, 1204. [Google Scholar] [CrossRef] [PubMed]
- Payton, N.M.; Marseglia, A.; Grande, G.; Fratiglioni, L.; Kivipelto, M.; Bäckman, L.; Laukka, E.J. Trajectories of cognitive decline and dementia development: A 12-year longitudinal study. Alzheimer Dement. 2022, 1–11. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Excluded Channels | Excluded Channels Number/All |
|---|---|---|
| 10 | 13 | 1/43 |
| 27 | 7, 17 | 2/43 |
| 31 | 2 | 1/43 |
| 56 | 7 | 1/43 |
| 58 | 1, 8 | 2/43 |
| SD Group (n = 37) | HC Group (n = 37) | t/Z/χ2 | p-Value | |
|---|---|---|---|---|
| Age (years) | 66.92 ± 3.59 | 66.86 ± 3.57 | 0.07 | 0.95 |
| Education (year) | 11.35 ± 1.96 | 11.41 ± 2.53 | −0.10 | 0.92 |
| Gender (male/female) | 11/26 | 10/27 | 0.07 | 0.80 |
| MoCA | 27.00 (26.00–28.00) | 27.00 (26.00–27.00) | −1.29 | 0.20 |
| GDS | 5.00 (4.00–5.00) | 4.00 (4.00–5.00) | −1.16 | 0.25 |
| Tea drinking situation | ||||
| Never | 18 | 18 | −0.112 | 0.911 |
| Rarely | 6 | 7 | ||
| Regular | 13 | 12 | ||
| Coffee drinking situation | ||||
| Never | 27 | 32 | −1.427 | 0.154 |
| Rarely | 6 | 3 | ||
| Regular | 4 | 2 |
| SD Group (n = 37) | HC Group (n = 37) | t/Z | p-Value | Effect Size | |
|---|---|---|---|---|---|
| 0-back Accuracy | 97.62 (92.86–100.00) | 100.00 (97.62–100.00) | 2.15 | 0.03 * | 0.26 |
| 0-back RT (ms) | 571.49 (504.40–616.06) | 567.60 (505.93–607.84) | 0.06 | 0.95 | −0.008 |
| 1-back Accuracy | 90.48 (85.71–94.05) | 90.48 (88.10–92.86) | 0.43 | 0.67 | −0.057 |
| 1-back RT (ms) | 691.82 ± 110.67 | 685.06 ± 116.35 | 0.26 | 0.80 | 0.059 |
| 2-back Accuracy | 83.59 ± 7.87 | 86.81 ± 6.14 | −1.96 | 0.05 * | −0.456 |
| 2-back RT (ms) | 798.29 (719.48–934.14) | 833.90 (737.95–921.62) | 0.33 | 0.74 | −0.045 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Zhang, L.; Zhu, J.; Guo, Z.; Lin, M.; Bai, L.; Zheng, P.; Liu, W.; Huang, J.; Liu, Z. Prefrontal Cortex Hemodynamics and Functional Connectivity Changes during Performance Working Memory Tasks in Older Adults with Sleep Disorders. Brain Sci. 2023, 13, 497. https://doi.org/10.3390/brainsci13030497
Gao J, Zhang L, Zhu J, Guo Z, Lin M, Bai L, Zheng P, Liu W, Huang J, Liu Z. Prefrontal Cortex Hemodynamics and Functional Connectivity Changes during Performance Working Memory Tasks in Older Adults with Sleep Disorders. Brain Sciences. 2023; 13(3):497. https://doi.org/10.3390/brainsci13030497
Chicago/Turabian StyleGao, Jiahui, Lin Zhang, Jingfang Zhu, Zhenxing Guo, Miaoran Lin, Linxin Bai, Peiyun Zheng, Weilin Liu, Jia Huang, and Zhizhen Liu. 2023. "Prefrontal Cortex Hemodynamics and Functional Connectivity Changes during Performance Working Memory Tasks in Older Adults with Sleep Disorders" Brain Sciences 13, no. 3: 497. https://doi.org/10.3390/brainsci13030497
APA StyleGao, J., Zhang, L., Zhu, J., Guo, Z., Lin, M., Bai, L., Zheng, P., Liu, W., Huang, J., & Liu, Z. (2023). Prefrontal Cortex Hemodynamics and Functional Connectivity Changes during Performance Working Memory Tasks in Older Adults with Sleep Disorders. Brain Sciences, 13(3), 497. https://doi.org/10.3390/brainsci13030497