

Gastrodin Improves Cognitive Dysfunction in REM Sleep-Deprived Rats by Regulating TLR4/NF-κB and Wnt/β-Catenin Signaling Pathways
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs
2.2. Animals
2.3. REM Sleep Deprivation
2.4. Sodium Pentobarbital-Induced Sleeping
2.5. Morris Water Maze
2.6. HE Staining
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. GAS Improved Sleep Latency and Sleep Duration in REM Sleep-Deprived Rats
3.2. GAS Prevents Cognitive Impairment in REM Sleep-Deprived Rats
3.3. GAS Attenuates Pathological Damage in the Hippocampus and Cortex of REM Sleep-Deprived Rats
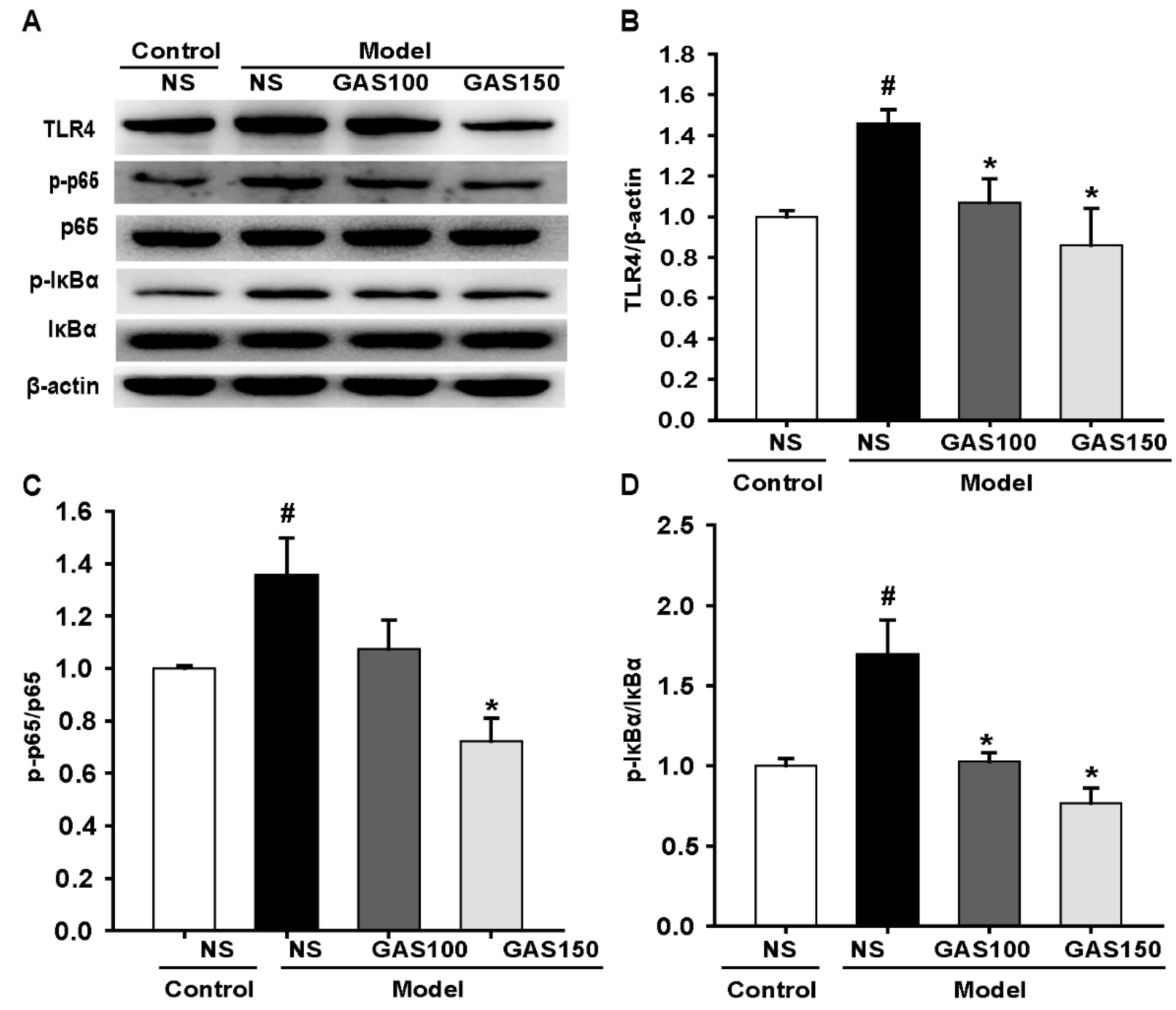
3.4. GAS Suppressed the TLR4/NF-κB Signaling Pathway in REM Sleep-Deprived Rats
3.5. GAS Activated the Wnt/β-Catenin Signaling Pathway in REM Sleep-Deprived Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Groeger, J.A.; Lo, J.C.; Santhi, N.; Lazar, A.S.; Dijk, D.J. Contrasting Effects of Sleep Restriction, Total Sleep Deprivation, and Sleep Timing on Positive and Negative Affect. Front. Behav. Neurosci. 2022, 16, 911994. [Google Scholar] [CrossRef] [PubMed]
- Pires, G.N.; Tufik, S.; Andersen, M.L. Relationship between sleep deprivation and anxiety—Experimental research perspective. Einstein 2012, 10, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.N.; Cui, Y.; Zhang, B.; Huang, S.M. Sleep deficiency promotes Alzheimer’s disease development and progression. Front. Neurol. 2022, 13, 1053942. [Google Scholar] [CrossRef] [PubMed]
- Kupeli Akkol, E.; Bardakci, H.; Yucel, C.; Seker Karatoprak, G.; Karpuz, B.; Khan, H. A New Perspective on the Treatment of Alzheimer’s Disease and Sleep Deprivation-Related Consequences: Can Curcumin Help? Oxidative Med. Cell. Longev. 2022, 2022, 6168199. [Google Scholar] [CrossRef]
- Wu, H.; Dunnett, S.; Ho, Y.S.; Chang, R.C. The role of sleep deprivation and circadian rhythm disruption as risk factors of Alzheimer’s disease. Front. Neuroendocrinol. 2019, 54, 100764. [Google Scholar] [CrossRef]
- Aurich, M.F.; Rodrigues, L.S.; Targa, A.D.S.; Noseda, A.C.D.; Cunha, F.D.W.; Lima, M.M.S. Olfactory impairment is related to REM sleep deprivation in rotenone model of Parkinson’s disease. Sleep Sci. 2017, 10, 47–54. [Google Scholar] [CrossRef]
- Hu, B.; Liu, C.; Lv, T.; Luo, F.; Qian, C.; Zhang, J.; Ye, M.; Liu, Z. Meta-analysis of sleep deprivation effects on depression in rodents. Brain Res. 2022, 1782, 147841. [Google Scholar] [CrossRef]
- Boyce, R.; Williams, S.; Adamantidis, A. REM sleep and memory. Curr. Opin. Neurobiol. 2017, 44, 167–177. [Google Scholar] [CrossRef]
- Milman, N.E.P.; Tinsley, C.E.; Raju, R.M.; Lim, M.M. Loss of sleep when it is needed most—Consequences of persistent developmental sleep disruption: A scoping review of rodent models. Neurobiol. Sleep Circadian Rhythm. 2023, 14, 100085. [Google Scholar] [CrossRef]
- Agrawal, S.; Singh, V.; Singh, C.; Singh, A. A review on pathophysiological aspects of Sleep Deprivation. CNS Neurol. Disord. Drug Targets 2022. [Google Scholar] [CrossRef]
- de Mendonca, F.M.R.; de Mendonca, G.; Souza, L.C.; Galvao, L.P.; Paiva, H.S.; de Azevedo Marques Perico, C.; Torales, J.; Ventriglio, A.; Castaldelli-Maia, J.M.; Silva, A.S.M. Benzodiazepines and Sleep Architecture: A Systematic Review. CNS Neurol. Disord. Drug Targets 2023, 22, 172–179. [Google Scholar] [PubMed]
- Simon, K.C.; Whitehurst, L.N.; Zhang, J.; Mednick, S.C. Zolpidem Maintains Memories for Negative Emotions Across a Night of Sleep. Affect. Sci. 2022, 3, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Wichniak, A.; Wierzbicka, A. The effects of antidepressants on sleep in depressed patients with particular reference to trazodone in comparison to agomelatine, amitriptyline, doxepin, mianserine and mirtazapine. Pol. Merkur. Lek. Organ Pol. Tow. Lek. 2011, 31, 65–70. [Google Scholar]
- de Oliveira, P.; Cella, C.; Locker, N.; Ravindran, K.K.G.; Mendis, A.; Wafford, K.; Gilmour, G.; Dijk, D.J.; Winsky-Sommerer, R. Improved Sleep, Memory, and Cellular Pathological Features of Tauopathy, Including the NLRP3 Inflammasome, after Chronic Administration of Trazodone in rTg4510 Mice. J. Neurosci. Off. J. Soc. Neurosci. 2022, 42, 3494–3509. [Google Scholar] [CrossRef]
- Heese, K. Gastrodia elata Blume (Tianma): Hope for Brain Aging and Dementia. Evid.-Based Complement. Altern. Med. eCAM 2020, 2020, 8870148. [Google Scholar] [CrossRef]
- Choi, J.J.; Oh, E.H.; Lee, M.K.; Chung, Y.B.; Hong, J.T.; Oh, K.W. Gastrodiae Rhizoma Ethanol Extract Enhances Pentobarbital-Induced Sleeping Behaviors and Rapid Eye Movement Sleep via the Activation of GABAA-ergic Transmission in Rodents. Evid.-Based Complement. Altern. Med. eCAM 2014, 2014, 426843. [Google Scholar] [CrossRef]
- Xu, C.B.; Guo, Q.L.; Wang, Y.N.; Lin, S.; Zhu, C.G.; Shi, J.G. Gastrodin Derivatives from Gastrodia elata. Nat. Prod. Bioprospect. 2019, 9, 393–404. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, J.; Peng, M.; Meng, H.; Ma, H.; Cai, P.; Xu, Y.; Zhao, Q.; Si, G. A Review on Central Nervous System Effects of Gastrodin. Front. Pharmacol. 2018, 9, 24. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Gao, Y.G.; Zang, P.; Gu, P.P.; Zhao, Y.; He, Z.M.; Zhu, H.Y. Research progress on mechanism of gastrodin and p-hydroxybenzyl alcohol on central nervous system. China J. Chin. Mater. Med. 2020, 45, 312–320. [Google Scholar] [CrossRef]
- Liu, B.; Li, F.; Shi, J.; Yang, D.; Deng, Y.; Gong, Q. Gastrodin ameliorates subacute phase cerebral ischemia-reperfusion injury by inhibiting inflammation and apoptosis in rats. Mol. Med. Rep. 2016, 14, 4144–4152. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, P.; Yang, L.; Duan, X. Network pharmacology and molecular docking analysis on molecular targets and mechanisms of Gastrodia elata Blume in the treatment of ischemic stroke. Exp. Ther. Med. 2022, 24, 742. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Gao, J.M.; Li, F.; Gong, Q.H.; Shi, J.S. Gastrodin Attenuates Bilateral Common Carotid Artery Occlusion-Induced Cognitive Deficits via Regulating Abeta-Related Proteins and Reducing Autophagy and Apoptosis in Rats. Front. Pharmacol. 2018, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Chen, H.; Meng, Z.; Meng, S. Gastrodin and Vascular Dementia: Advances and Current Perspectives. Evid.-Based Complement. Altern. Med. eCAM 2022, 2022, 2563934. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, E.; Yang, H.; Chen, Y.; Tao, L.; Xu, Y.; Chen, T.; Shen, X. Gastrodin Ameliorates Cognitive Dysfunction in Vascular Dementia Rats by Suppressing Ferroptosis via the Regulation of the Nrf2/Keap1-GPx4 Signaling Pathway. Molecules 2022, 27, 6311. [Google Scholar] [CrossRef] [PubMed]
- Ugalde-Muniz, P.; Hernandez-Luna, M.G.; Garcia-Velasco, S.; Lugo-Huitron, R.; Murcia-Ramirez, J.; Martinez-Tapia, R.J.; Noriega-Navarro, R.; Navarro, L. Activation of dopamine D2 receptors attenuates neuroinflammation and ameliorates the memory impairment induced by rapid eye movement sleep deprivation in a murine model. Front. Neurosci. 2022, 16, 988167. [Google Scholar] [CrossRef]
- Zhu, Y.; Gao, M.; Huang, H.; Gao, S.H.; Liao, L.Y.; Tao, Y.; Cheng, H.; Gao, C.Y. p75NTR Ectodomain Ameliorates Cognitive Deficits and Pathologies in a Rapid Eye Movement Sleep Deprivation Mice Model. Neuroscience 2022, 496, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Yu, L.H.; Zhang, J.; Xie, D.J.; Zhang, X.X.; Yu, J.M. Network Pharmacology-Based and Molecular Docking-Based Analysis of Suanzaoren Decoction for the Treatment of Parkinson’s Disease with Sleep Disorder. BioMed Res. Int. 2021, 2021, 1752570. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, Y.; Luo, Y.; Du, Y.; Zhang, X.; Fu, J. Curcumin inhibits LPS-induced neuroinflammation by promoting microglial M2 polarization via TREM2/ TLR4/ NF-kappaB pathways in BV2 cells. Mol. Immunol. 2019, 116, 29–37. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Ling, E.A.; Lu, D. Microglia mediated neuroinflammation—Signaling regulation and therapeutic considerations with special reference to some natural compounds. Histol. Histopathol. 2020, 35, 1229–1250. [Google Scholar] [CrossRef]
- Marchetti, B.; Tirolo, C.; L’Episcopo, F.; Caniglia, S.; Testa, N.; Smith, J.A.; Pluchino, S.; Serapide, M.F. Parkinson’s disease, aging and adult neurogenesis: Wnt/beta-catenin signalling as the key to unlock the mystery of endogenous brain repair. Aging Cell 2020, 19, e13101. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Bian, L.G.; Yang, P.; Sui, Y.; Li, R.; Chen, Y.L.; Sun, L.; Ai, Q.L.; Zhong, L.M.; Lu, D. Gastrodin attenuates proliferation and inflammatory responses in activated microglia through Wnt/beta-catenin signaling pathway. Brain Res. 2019, 1717, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Huang, B.; Qian, X.; Zhang, Y.; Teng, B. Gastrodin improves preeclampsia-induced cell apoptosis by regulation of TLR4/NF-kappaB in rats. Food Sci. Nutr. 2020, 8, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin Alleviates Acute Sleep Deprivation-Induced Memory Loss in Mice by Suppressing Hippocampal Ferroptosis. Front. Pharmacol. 2021, 12, 708645. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.J.; Wang, Q.Q.; Zhang, X.P.; Wang, G.Y.; Liu, T.; Deng, N.; Yan, D.Q. Establishment of a rat model with ageing insomnia induced by D-galactosef and para-chlorophenylalanine. Exp. Ther. Med. 2020, 20, 3228–3236. [Google Scholar] [CrossRef] [PubMed]
- Zamore, Z.; Veasey, S.C. Neural consequences of chronic sleep disruption. Trends Neurosci. 2022, 45, 678–691. [Google Scholar] [CrossRef]
- Villafuerte, G.; Miguel-Puga, A.; Rodriguez, E.M.; Machado, S.; Manjarrez, E.; Arias-Carrion, O. Sleep deprivation and oxidative stress in animal models: A systematic review. Oxidative Med. Cell Longev. 2015, 2015, 234952. [Google Scholar] [CrossRef]
- Alkadhi, K.A.; Alhaider, I.A. Caffeine and REM sleep deprivation: Effect on basal levels of signaling molecules in area CA1. Mol. Cell Neurosci. 2016, 71, 125–131. [Google Scholar] [CrossRef]
- Turan, I.; Sayan Ozacmak, H.; Ozacmak, V.H.; Ergenc, M.; Bayraktaroglu, T. The effects of glucagon-like peptide 1 receptor agonist (exenatide) on memory impairment, and anxiety- and depression-like behavior induced by REM sleep deprivation. Brain Res. Bull. 2021, 174, 194–202. [Google Scholar] [CrossRef]
- Stokholm, M.G.; Iranzo, A.; Ostergaard, K.; Serradell, M.; Otto, M.; Svendsen, K.B.; Garrido, A.; Vilas, D.; Borghammer, P.; Santamaria, J.; et al. Assessment of neuroinflammation in patients with idiopathic rapid-eye-movement sleep behaviour disorder: A case-control study. Lancet Neurol. 2017, 16, 789–796. [Google Scholar] [CrossRef]
- Ye, T.; Meng, X.; Wang, R.; Zhang, C.; He, S.; Sun, G.; Sun, X. Gastrodin Alleviates Cognitive Dysfunction and Depressive-Like Behaviors by Inhibiting ER Stress and NLRP3 Inflammasome Activation in db/db Mice. Int. J. Mol. Sci. 2018, 19, 3977. [Google Scholar] [CrossRef]
- Wang, S.; Nan, Y.; Zhu, W.; Yang, T.; Tong, Y.; Fan, Y. Gastrodin improves the neurological score in MCAO rats by inhibiting inflammation and apoptosis, promoting revascularization. Int. J. Clin. Exp. Pathol. 2018, 11, 5343–5350. [Google Scholar] [PubMed]
- Zusso, M.; Lunardi, V.; Franceschini, D.; Pagetta, A.; Lo, R.; Stifani, S.; Frigo, A.C.; Giusti, P.; Moro, S. Ciprofloxacin and levofloxacin attenuate microglia inflammatory response via TLR4/NF-kB pathway. J. Neuroinflamm. 2019, 16, 148. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, L.; Liu, T.; Wang, J.; Wen, A.; Ding, Y. Ellagic acid protects mice against sleep deprivation-induced memory impairment and anxiety by inhibiting TLR4 and activating Nrf2. Aging 2020, 12, 10457–10472. [Google Scholar] [CrossRef]
- Gao, T.; Wang, Z.; Dong, Y.; Cao, J.; Chen, Y. Melatonin-Mediated Colonic Microbiota Metabolite Butyrate Prevents Acute Sleep Deprivation-Induced Colitis in Mice. Int. J. Mol. Sci. 2021, 22, 11894. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, B. Wnt/beta-Catenin Signaling Pathway Governs a Full Program for Dopaminergic Neuron Survival, Neurorescue and Regeneration in the MPTP Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 3743. [Google Scholar] [CrossRef]
- Anthony, C.C.; Robbins, D.J.; Ahmed, Y.; Lee, E. Nuclear Regulation of Wnt/beta-Catenin Signaling: It’s a Complex Situation. Genes 2020, 11, 886. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Li, F.; Xu, Y.; Wu, Q.; Shi, J. Gastrodin Improves Cognitive Dysfunction in REM Sleep-Deprived Rats by Regulating TLR4/NF-κB and Wnt/β-Catenin Signaling Pathways. Brain Sci. 2023, 13, 179. https://doi.org/10.3390/brainsci13020179
Liu B, Li F, Xu Y, Wu Q, Shi J. Gastrodin Improves Cognitive Dysfunction in REM Sleep-Deprived Rats by Regulating TLR4/NF-κB and Wnt/β-Catenin Signaling Pathways. Brain Sciences. 2023; 13(2):179. https://doi.org/10.3390/brainsci13020179
Chicago/Turabian StyleLiu, Bo, Fei Li, Yunyan Xu, Qin Wu, and Jingshan Shi. 2023. "Gastrodin Improves Cognitive Dysfunction in REM Sleep-Deprived Rats by Regulating TLR4/NF-κB and Wnt/β-Catenin Signaling Pathways" Brain Sciences 13, no. 2: 179. https://doi.org/10.3390/brainsci13020179
APA StyleLiu, B., Li, F., Xu, Y., Wu, Q., & Shi, J. (2023). Gastrodin Improves Cognitive Dysfunction in REM Sleep-Deprived Rats by Regulating TLR4/NF-κB and Wnt/β-Catenin Signaling Pathways. Brain Sciences, 13(2), 179. https://doi.org/10.3390/brainsci13020179