
Tics and Emotions



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Anatomical Circuitry and Tic Behavior
2.1. Motor Aspects of Tics
2.2. Non-Motor Aspects of Tics
2.3. Expression Networks
3. Behavioral Aspects of Tics
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruhrman, D.; Gev, E.; Benaroya-Milshtein, N.; Fennig, S.; Krispin, O.; Apter, A.; Steinberg, T. Non-Motor Aspects of Tic Disorders—New Developments. Front. Psychiatry 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Cavanna, A.E.; Black, K.; Hallett, M.; Voon, V. Neurobiology of the Premonitory Urge in Tourette’s Syndrome: Pathophysiology and Treatment Implications. J. Neuropsychiatry Clin. Neurosci. 2017, 29, 95–104. [Google Scholar] [CrossRef]
- Alm, P.A. Stuttering and the basal ganglia circuits: A critical review of possible relations. J. Commun. Disord. 2004, 37, 325–369. [Google Scholar] [CrossRef]
- Martino, D.; Hedderly, T. Tics and stereotypies: A comparative clinical review. Park. Relat. Disord. 2019, 59, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.; Mahone, E.M.; Singer, H.S. Nonautistic Motor Stereotypies: Clinical Features and Longitudinal Follow-Up. Pediatr. Neurol. 2008, 38, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Bostan, A.C.; Strick, P.L. The basal ganglia and the cerebellum: Nodes in an integrated network. Nat. Rev. Neurosci. 2018, 19, 338–350. [Google Scholar] [CrossRef]
- Hoshi, E.; Tremblay, L.; Féger, J.; Carras, P.L.; Strick, P.L. The cerebellum communicates with the basal ganglia. Nat. Neurosci. 2005, 8, 1491–1493. [Google Scholar] [CrossRef]
- Bornstein, R.A.; Stefl, M.E.; Hammond, L. A survey of Tourette syndrome patients and their families: The 1987 Ohio Tourette Survey. J. Neuropsychiatry Clin. Neurosci. 1990, 2, 275–281. [Google Scholar] [CrossRef]
- Eapen, V.; Fox-Hiley, P.; Banerjee, S.; Robertson, M. Clinical features and associated psychopathology in a Tourette syndrome cohort. Acta Neurol. Scand. 2004, 109, 255–260. [Google Scholar] [CrossRef]
- Silva, R.R.; Munoz, D.M.; Barickman, J.; Friedhoff, A.J. Environmental Factors and Related Fluctuation of Symptoms in Children and Adolescents with Tourette’s Disorder. J. Child Psychol. Psychiatry 1995, 36, 305–312. [Google Scholar] [CrossRef]
- Findley, D.B.; Leckman, J.F.; Katsovich, L.; Lin, H.; Zhang, H.; Grantz, H.; Otka, J.; Lombroso, P.J.; King, R.A. Development of the Yale Children’s Global Stress Index (YCGSI) and Its Application in Children and Adolescents with Tourette’s Syndrome and Obsessive-Compulsive Disorder. J. Am. Acad. Child Adolesc. Psychiatry 2003, 42, 450–457. [Google Scholar] [CrossRef]
- Hoekstra, P.J.; Steenhuis, M.-P.; Kallenberg, C.G.M.; Minderaa, R.B. Association of Small Life Events with Self Reports of Tic Severity in Pediatric and Adult Tic Disorder Patients. J. Clin. Psychiatry 2004, 65, 426–431. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, K. A cognitive-behavioral/psychophysiological model of tic disorders. Behav. Res. Ther. 2002, 40, 1113–1142. [Google Scholar] [CrossRef]
- O’Connor, K.; Brisebois, H.; Brault, M.; Robillard, S.; Loiselle, J. Behavioral activity associated with onset in chronic tic and habit disorder. Behav. Res. Ther. 2003, 41, 241–249. [Google Scholar] [CrossRef]
- Conelea, C.; Woods, D.W. The influence of contextual factors on tic expression in Tourette’s syndrome: A review. J. Psychosom. Res. 2008, 65, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Mink, J.W. Basal ganglia dysfunction in Tourette’s syndrome: A new hypothesis. Pediatr. Neurol. 2001, 25, 190–198. [Google Scholar] [CrossRef]
- Caligiore, D.; Mannella, F.; Arbib, M.A.; Baldassarre, G. Dysfunctions of the basal ganglia-cerebellar-thalamo-cortical system produce motor tics in Tourette syndrome. PLoS Comput. Biol. 2017, 13, e1005395. [Google Scholar] [CrossRef]
- Godar, S.C.; Bortolato, M. What makes you tic? Translational approaches to study the role of stress and contextual triggers in Tourette syndrome. Neurosci. Biobehav. Rev. 2017, 76, 123–133. [Google Scholar] [CrossRef]
- Buse, J.; Kirschbaum, C.; Leckman, J.F.; Münchau, A.; Roessner, V. The Modulating Role of Stress in the Onset and Course of Tourette’s Syndrome. Behav. Modif. 2014, 38, 184–216. [Google Scholar] [CrossRef]
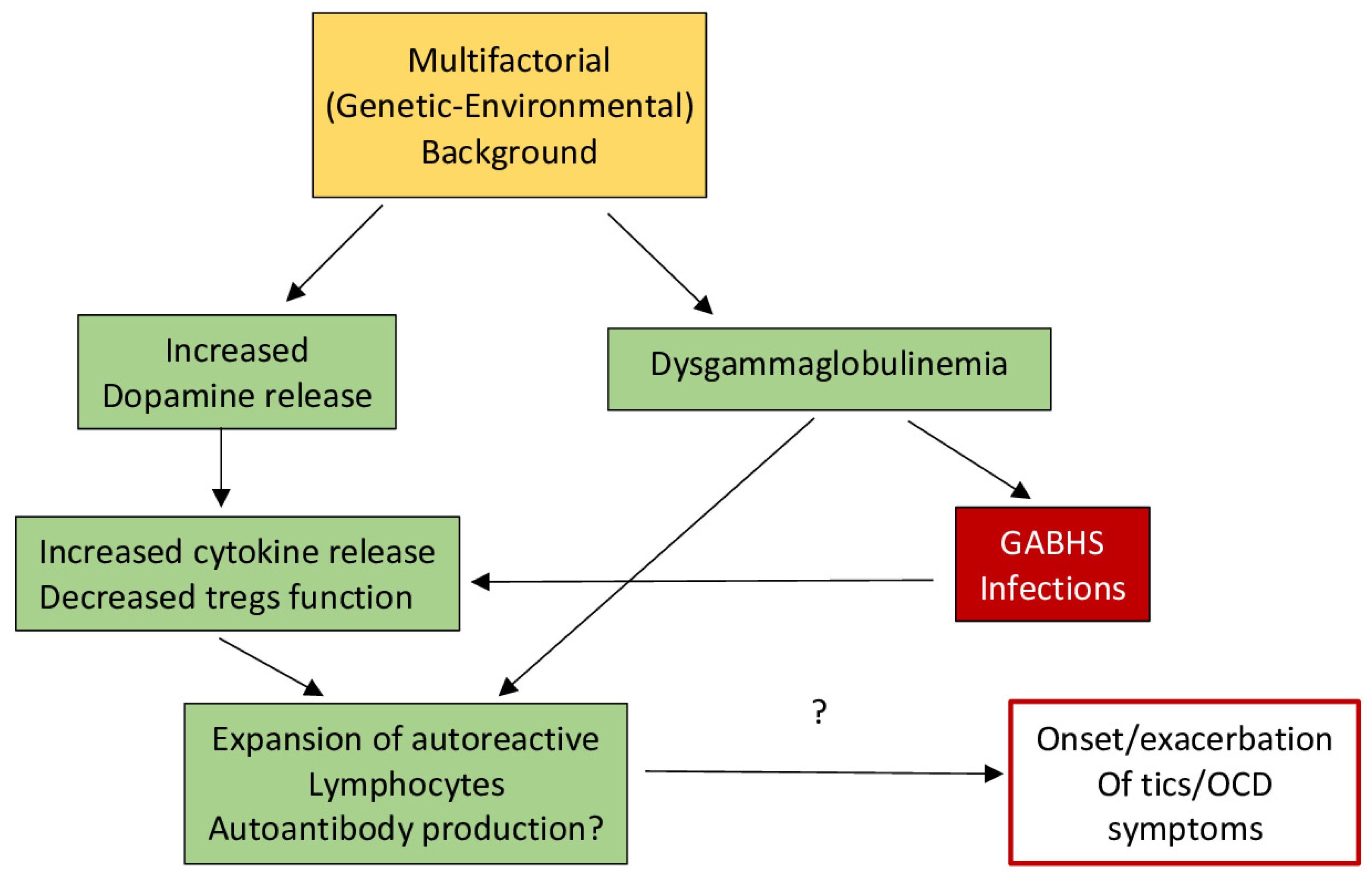
- Martino, D.; Dale, R.C.; Gilbert, D.L.; Giovannoni, G.; Leckman, J.F. Immunopathogenic mechanisms in tourette syndrome: A critical review. Mov. Disord. 2009, 24, 1267–1279. [Google Scholar] [CrossRef]
- Pacheco, R.; Prado, C.E.; Barrientos, M.J.; Bernales, S. Role of dopamine in the physiology of T-cells and dendritic cells. J. Neuroimmunol. 2009, 216, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Leckman, J.F.; Katsovich, L.; Kawikova, I.; Lin, H.; Zhang, H.; Krönig, H.; Morshed, S.; Parveen, S.; Grantz, H.; Lombroso, P.J.; et al. Increased serum levels of interleukin-12 and tumor necrosis factor-alpha in Tourette’s syndrome. Biol. Psychiatry 2005, 57, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Leslie, D.L.; Kozma, L.; Martin, A.; Landeros, A.; Katsovich, L.; King, R.A.; Leckman, J.F. Neuropsychiatric Disorders Associated With Streptococcal Infection: A Case-Control Study Among Privately Insured Children. J. Am. Acad. Child Adolesc. Psychiatry 2008, 47, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.W. Pathogenesis of Group A Streptococcal Infections. Clin. Microbiol. Rev. 2000, 13, 470–511. [Google Scholar] [CrossRef]
- Husby, G.; Van De Rijn, I.; Zabriskie, J.B.; Abdin, Z.H.; Williams, R.C. Antibodies reacting with cytoplasm of subthalamic and caudate nuclei neurons in chorea and acute rheumatic fever. J. Exp. Med. 1976, 144, 1094–1110. [Google Scholar] [CrossRef]
- Church, A.J.; Cardoso, F.; Dale, R.C.; Lees, A.J.; Thompson, E.J.; Giovannoni, G. Anti–basal ganglia antibodies in acute and persistent Sydenham’s chorea. Neurology 2002, 59, 227–231. [Google Scholar] [CrossRef]
- Ferrari, M.; Termine, C.; Franciotta, D.; Castiglioni, E.; Pagani, A.; Lanzi, G.; Marino, F.; Lecchini, S.; Cosentino, M.; Balottin, U. Dopaminergic receptor D5 mRNA expression is increased in circulating lymphocytes of Tourette syndrome patients. J. Psychiatr. Res. 2008, 43, 24–29. [Google Scholar] [CrossRef]
- Nagai, Y.; Ueno, S.; Saeki, Y.; Soga, F.; Hirano, M.; Yanagihara, T. Decrease of the D3 dopamine receptor mRNA expression in lymphocytes from patients with Parkinson’s disease. Neurology 1996, 46, 791–795. [Google Scholar] [CrossRef]
- Barbanti, P.; Fabbrini, G.; Ricci, A.; Bruno, G.; Cerbo, R.; Bronzetti, E.; Amenta, F.; Lenzi, G.L. Reduced density of dopamine D2-like receptors on peripheral blood lymphocytes in Alzheimer’s disease. Mech. Ageing Dev. 2000, 120, 65–75. [Google Scholar] [CrossRef]
- Ilani, T.; Ben-Shachar, D.; Strous, R.D.; Mazor, M.; Sheinkman, A.; Kotler, M.; Fuchs, S. A peripheral marker for schizophrenia: Increased levels of D3 dopamine receptor mRNA in blood lymphocytes. Proc. Natl. Acad. Sci. USA 2001, 98, 625–628. [Google Scholar] [CrossRef]
- Rocca, P.; De Leo, C.; Eva, C.; Marchiaro, L.; Milani, A.M.; Musso, R.; Ravizza, L.; Zanalda, E.; Bogetto, F. Decrease of the D4 dopamine receptor messenger RNA expression in lymphocytes from patients with major depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2002, 26, 1155–1160. [Google Scholar] [CrossRef]
- Yang, Z.; Asico, L.D.; Yu, P.; Wang, Z.; Jones, J.E.; Escano, C.S.; Wang, X.; Quinn, M.T.; Sibley, D.R.; Romero, G.G.; et al. D5dopamine receptor regulation of reactive oxygen species production, NADPH oxidase, and blood pressure. Am. J. Physiol. Integr. Comp. Physiol. 2006, 290, R96–R104. [Google Scholar] [CrossRef] [PubMed]
- Levings, M.K.; Allan, S.; D’Hennezel, E.; Piccirillo, C.A. Functional Dynamics of Naturally Occurring Regulatory T Cells in Health and Autoimmunity. Synthetic Vaccines 2006, 92, 119–155. [Google Scholar] [CrossRef]
- Bronfeld, M.; Bar-Gad, I. Tic Disorders. Neuroscience 2013, 19, 101–108. [Google Scholar] [CrossRef]
- McCairn, K.W.; Iriki, A.; Isoda, M. Global Dysrhythmia of Cerebro-Basal Ganglia–Cerebellar Networks Underlies Motor Tics following Striatal Disinhibition. J. Neurosci. 2013, 33, 697–708. [Google Scholar] [CrossRef]
- Yelnik, J. Functional anatomy of the basal ganglia. Mov. Disord. 2002, 17, S15–S21. [Google Scholar] [CrossRef] [PubMed]
- McCairn, K.W.; Nagai, Y.; Hori, Y.; Ninomiya, T.; Kikuchi, E.; Lee, J.-Y.; Suhara, T.; Iriki, A.; Minamimoto, T.; Takada, M.; et al. A Primary Role for Nucleus Accumbens and Related Limbic Network in Vocal Tics. Neuron 2016, 89, 300–307. [Google Scholar] [CrossRef]
- Gemba, H.; Miki, N.; Kazuo, S. Cortical field potentials preceding vocalization and influences of cerebellar hemispherectomy upon them in monkeys. Brain Res. 1995, 697, 143–151. [Google Scholar] [CrossRef]
- Marchand, W.R. Cortico-basal ganglia circuitry: A review of key research and implications for functional connectivity studies of mood and anxiety disorders. Anat. Embryol. 2010, 215, 73–96. [Google Scholar] [CrossRef]
- LeDoux, J.E.; Ruggiero, D.A.; Reis, D.J. Projections to the subcortical forebrain from anatomically defined regions of the medial geniculate body in the rat. J. Comp. Neurol. 1985, 242, 182–213. [Google Scholar] [CrossRef]
- Butler, T.; Pan, H.; Tuescher, O.; Engelien, A.; Goldstein, M.; Epstein, J.; Weisholtz, D.; Root, J.; Protopopescu, X.; Cunningham-Bussel, A.; et al. Human fear-related motor neurocircuitry. Neuroscience 2007, 150, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.E.; Moghaddam, B. Amygdala Regulation of Nucleus Accumbens Dopamine Output is Governed by the Prefrontal Cortex. J. Neurosci. 2001, 21, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Mogenson, G.J.; Jones, D.L.; Yim, C.Y. From motivation to action: Functional interface between the limbic system and the motor system. Prog. Neurobiol. 1980, 14, 69–97. [Google Scholar] [CrossRef]
- Pogorelov, V.; Xu, M.; Smith, H.R.; Buchanan, G.; Pittenger, C. Corticostriatal interactions in the generation of tic-like behaviors after local striatal disinhibition. Exp. Neurol. 2015, 265, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Maia, T.; Marsh, R.; Colibazzi, T.; Gerber, A.; Peterson, B.S. The Neural Circuits That Generate Tics in Tourette’s Syndrome. Am. J. Psychiatry 2011, 168, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Yael, D.; Vinner, E.; Bar-Gad, I. Pathophysiology of tic disorders. Mov. Disord. 2015, 30, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Neuner, I.; Werner, C.J.; Arrubla, J.; Stã¶cker, T.; Ehlen, C.; Wegener, H.P.; Schneider, F.; Shah, N.J. Imaging the where and when of tic generation and resting state networks in adult Tourette patients. Front. Hum. Neurosci. 2014, 8, 362. [Google Scholar] [CrossRef]
- Aylward, E.H.; Reiss, A.L.; Reader, M.J.; Singer, H.S.; Brown, J.E.; Denckla, M.B. Basal Ganglia Volumes in Children With Attention-Deficit Hyperactivity Disorder. J. Child Neurol. 1996, 11, 112–115. [Google Scholar] [CrossRef]
- Nakao, T.; Radua, J.; Rubia, K.; Mataix-Cols, D. Gray Matter Volume Abnormalities in ADHD: Voxel-Based Meta-Analysis Exploring the Effects of Age and Stimulant Medication. Am. J. Psychiatry 2011, 168, 1154–1163. [Google Scholar] [CrossRef]
- Bloch, M.H.; Leckman, J.F.; Zhu, H.; Peterson, B.S. Caudate volumes in childhood predict symptom severity in adults with Tourette syndrome. Neurology 2005, 65, 1253–1258. [Google Scholar] [CrossRef]
- Peterson, B.S.; Thomas, P.; Kane, M.J.; Scahill, L.; Zhang, H.; Bronen, R.; King, R.A.; Leckman, J.F.; Staib, L. Basal Ganglia Volumes in Patients with Gilles de la Tourette Syndrome. Arch. Gen. Psychiatry 2003, 60, 415–424. [Google Scholar] [CrossRef]
- Eddy, C.M.; Rickards, H.E.; Cavanna, A.E. Physiological Awareness Is Negatively Related to Inhibitory Functioning in Tourette Syndrome. Behav. Modif. 2014, 38, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Critchley, H.D.; Wiens, S.; Rotshtein, P.; Öhman, A.; Dolan, R. Neural systems supporting interoceptive awareness. Nat. Neurosci. 2004, 7, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Sutherland, A.N. Using animal models to develop therapeutics for Tourette Syndrome. Pharmacol. Ther. 2005, 108, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Zebardast, N.; Crowley, M.J.; Bloch, M.H.; Mayes, L.C.; Wyk, B.V.; Leckman, J.F.; Pelphrey, K.A.; Swain, J.E. Brain mechanisms for prepulse inhibition in adults with Tourette syndrome: Initial findings. Psychiatry Res. Neuroimaging 2013, 214, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Conelea, C.; Woods, D.W.; Brandt, B.C. The impact of a stress induction task on tic frequencies in youth with Tourette Syndrome. Behav. Res. Ther. 2011, 49, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Baldan, L.C.; Williams, K.A.; Gallezot, J.-D.; Pogorelov, V.; Rapanelli, M.; Crowley, M.; Anderson, G.M.; Loring, E.; Gorczyca, R.; Billingslea, E.; et al. Histidine Decarboxylase Deficiency Causes Tourette Syndrome: Parallel Findings in Humans and Mice. Neuron 2014, 82, 1186–1187. [Google Scholar] [CrossRef]
- Melillo, R.; Leisman, G. Neurobehavioral Disorders of Childhood: An Evolutionary Perspective; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Leisman, G.; Braun-Benjamin, O.; Melillo, R. Cognitive-motor interactions of the basal ganglia in development. Front. Syst. Neurosci. 2014, 8, 16. [Google Scholar] [CrossRef]
- Hoogduin, K.; Verdellen, C.; Cath, D. Exposure and response prevention in the treatment of Gilles de la Tourette’s syndrome: Four case studies. Clin. Psychol. Psychother. Int. J. Theory Practice 1997, 4, 125–135. [Google Scholar] [CrossRef]
- O’Connor, K.; Lavoie, M.; Blanchet, P.; St-Pierre-Delorme, M.E. Evaluation of a cognitive psychophysiological model for management of tic disorders: An open trial. Br. J. Psychiatry 2016, 209, 76–83. [Google Scholar] [CrossRef]
- Cavanna, A.E.; Selvini, C.; Luoni, C.; Eddy, C.M.; Ali, F.; Blangiardo, R.; Gagliardi, E.; Balottin, U.; Termine, C. Measuring Anger Expression in Young Patients with Tourette Syndrome. Child. Health Care 2014, 44, 264–276. [Google Scholar] [CrossRef]
- Freeman, R.D. Tourette Tourette Syndrome International Database Consortium Tic disorders and ADHD: Answers from a world-wide clinical dataset on Tourette syndrome. Eur. Child Adolesc. Psychiatry 2007, 16, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Coffey, B.J.; Biederman, J.; Smoller, J.W.; Geller, D.A.; Sarin, P.; Schwartz, S.; Kim, G.S. Anxiety Disorders and Tic Severity in Juveniles with Tourette’s Disorder. J. Am. Acad. Child Adolesc. Psychiatry 2000, 39, 562–568. [Google Scholar] [CrossRef]
- Dehning, S.; Burger, M.B.; Krause, D.; Jobst, A.; Yundina, E.; Müller, N.; Meyer, S.; Zill, P.; Buchheim, A. Tourette syndrome is associated with insecure attachment and higher aggression. Int. J. Neurosci. 2014, 125, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.; Renk, K. I’m Uncomfortable, You’re Uncomfortable, We’re Uncomfortable: An Integrative Family Approach to the Treatment of Tourette’s Disorder and Separation Anxiety Disorder in a Young Child. Clin. Case Stud. 2017, 16, 446–463. [Google Scholar] [CrossRef]
- Corbett, B.; Mendoza, S.; Baym, C.; Bunge, S.; Levine, S. Examining cortisol rhythmicity and responsivity to stress in children with Tourette syndrome. Psychoneuroendocrinology 2008, 33, 810–820. [Google Scholar] [CrossRef][Green Version]
- Shapiro, A.K.; Shapiro, E.S.; Young, J.G.; Feinberg, T.E. Gilles De La Tourette Syndrome, 2nd ed.; Raven Press: New York, NY, USA, 1988. [Google Scholar]
- Nickel, C.; Tanca, S.; Kolowos, S.; Pedrosa-Gil, F.; Bachler, E.; Loew, T.; Gross, M.; Rother, W.K.; Nickel, M.K. Men with chronic occupational stress benefit from behavioural/psycho-educational group training: A randomized, prospective, controlled trial. Psychol. Med. 2006, 37, 1141–1149. [Google Scholar] [CrossRef]
- A Nicolson, N.; van Diest, R. Salivary cortisol patterns in vital exhaustion. J. Psychosom. Res. 2000, 49, 335–342. [Google Scholar] [CrossRef]
- Ockenfels, M.C.; Porter, L.; Smyth, J.; Kirschbaum, C.; Hellhammer, D.H.; Stone, A.A. Effect of Chronic Stress Associated With Unemployment on Salivary Cortisol. Psychosom. Med. 1995, 57, 460–467. [Google Scholar] [CrossRef]
- Ehawksley, J.; Cavanna, A.E.; Enagai, Y. The role of the autonomic nervous system in Tourette Syndrome. Front. Neurosci. 2015, 9, 117. [Google Scholar] [CrossRef]
- Van Dijk, J.; Koenderink, M.; Kramer, C.; Heijer, J.D.; Roos, R. Non-invasive assessment of autonomic nervous function in Gilles de la Tourette syndrome. Clin. Neurol. Neurosurg. 1992, 94, 157–159. [Google Scholar] [CrossRef]
- Tulen, J.H.M.; Van De Wetering, B.J.M.; Boomsma, F. Autonomic Regulation during Rest and Mental Load in Gilles de la Tourette Syndrome. Psychol. Rep. 1998, 83, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y.; Cavanna, A.; Critchley, H.D. Influence of sympathetic autonomic arousal on tics: Implications for a therapeutic behavioral intervention for Tourette syndrome. J. Psychosom. Res. 2009, 67, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Katsovich, L.; Ghebremichael, M.; Findley, D.B.; Grantz, H.; Lombroso, P.J.; King, R.A.; Zhang, H.; Leckman, J.F. Psychosocial stress predicts future symptom severities in children and adolescents with Tourette syndrome and/or obsessive-compulsive disorder. J. Child Psychol. Psychiatry 2007, 48, 157–166. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leisman, G.; Sheldon, D. Tics and Emotions. Brain Sci. 2022, 12, 242. https://doi.org/10.3390/brainsci12020242
Leisman G, Sheldon D. Tics and Emotions. Brain Sciences. 2022; 12(2):242. https://doi.org/10.3390/brainsci12020242
Chicago/Turabian StyleLeisman, Gerry, and Dana Sheldon. 2022. "Tics and Emotions" Brain Sciences 12, no. 2: 242. https://doi.org/10.3390/brainsci12020242
APA StyleLeisman, G., & Sheldon, D. (2022). Tics and Emotions. Brain Sciences, 12(2), 242. https://doi.org/10.3390/brainsci12020242