Intentional Binding Effects in the Experience of Noticing the Regularity of a Perceptual-Motor Task


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
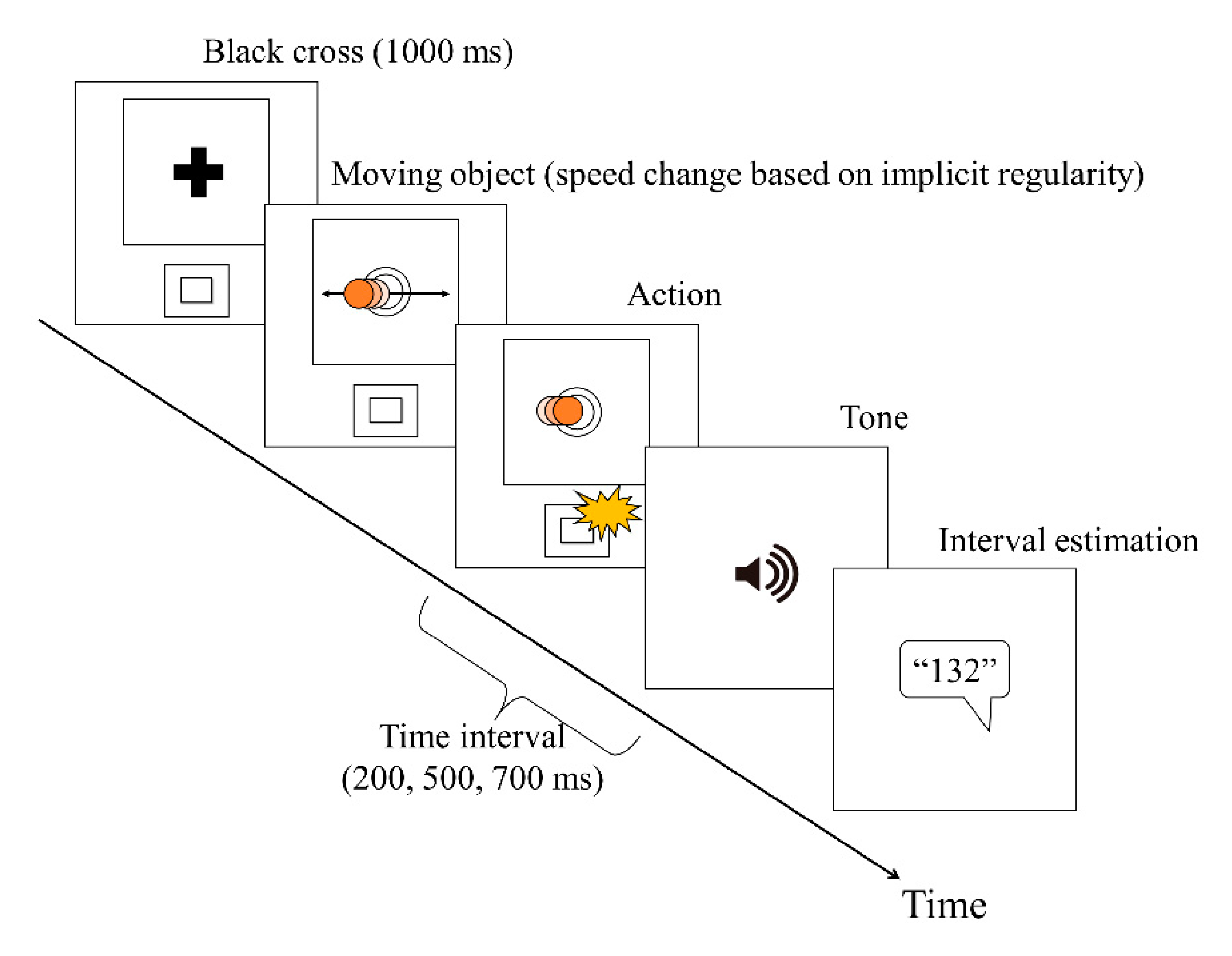
2.3. Procedure
2.3.1. Practice Task
2.3.2. Time Baseline Task
2.3.3. Experimental Motor Task
2.4. Data Analysis
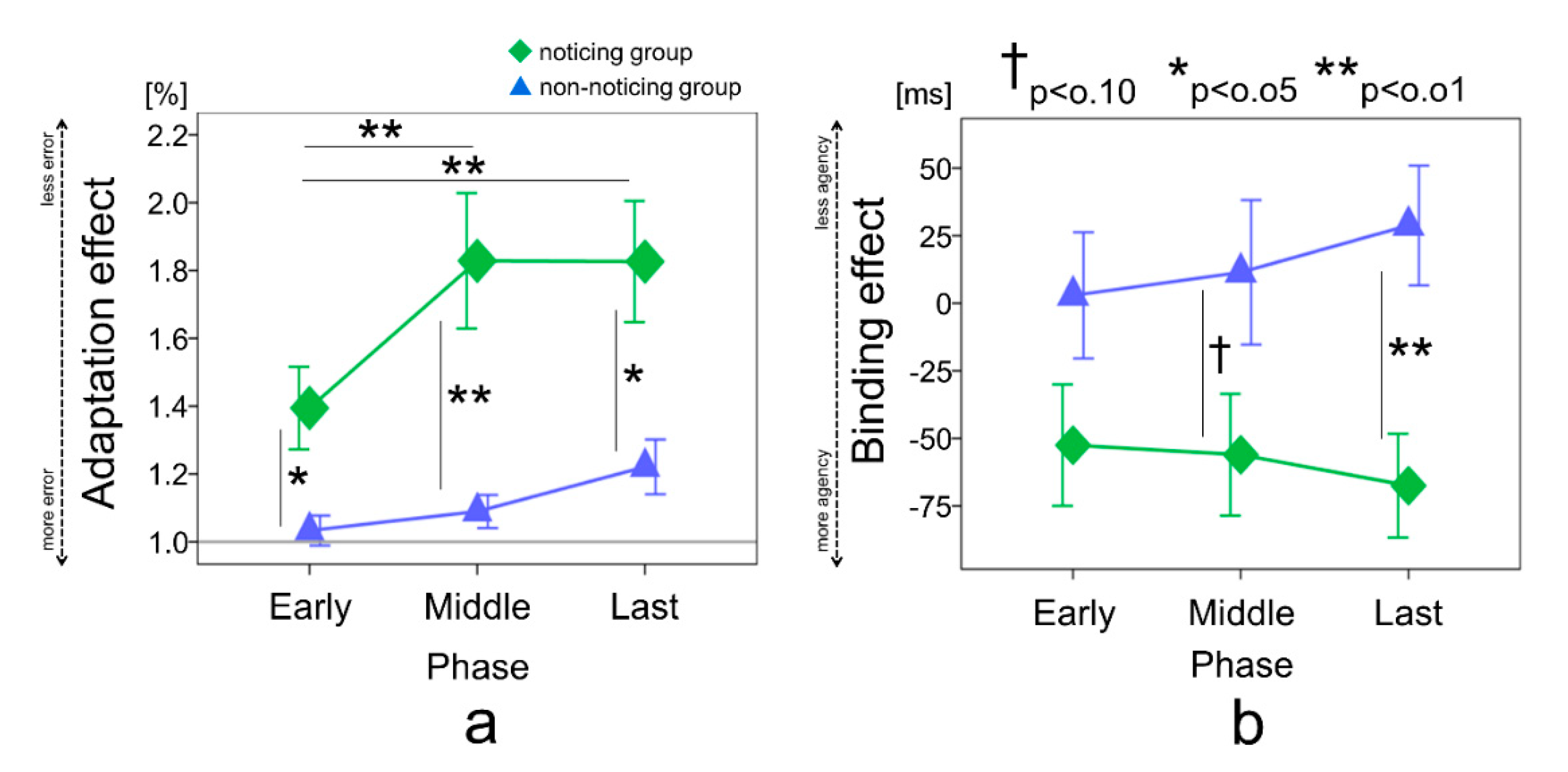
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Doya, K. Complementary roles of basal ganglia and cerebellum in learning and motor control. Curr. Opin. Neurobiol. 2000, 10, 732–739. [Google Scholar] [CrossRef]
- Kim, S.I. Neuroscientific model of motivational process. Front. Psychol. 2013, 4, 98. [Google Scholar] [CrossRef]
- Schultz, W. Behavioral theories and the neurophysiology of reward. Annu. Rev. Psychol. 2006, 57, 87–115. [Google Scholar] [CrossRef]
- Imamizu, H.; Miyauchi, S.; Tamada, T.; Sasaki, Y.; Takino, R.; Pütz, B.; Yoshioka, T.; Kawato, M. Human cerebellar activity reflecting an acquired internal model of a new tool. Nature 2000, 403, 192–195. [Google Scholar] [CrossRef]
- Wolpert, D.M. Computational approaches to motor control. Trends Cogn. Sci. 1997, 1, 209–216. [Google Scholar] [CrossRef]
- Blakemore, S.J.; Frith, C.D.; Wolpert, D.M. Spatio-temporal prediction modulates the perception of self-produced stimuli. J. Cogn. Neurosci. 1999, 11, 551–559. [Google Scholar] [CrossRef]
- Blakemore, S.J.; Wolpert, D.M.; Frith, C.D. Central cancellation of self-produced tickle sensation. Nat. Neurosci. 1998, 1, 635–640. [Google Scholar] [CrossRef]
- Frith, C.D.; Blakemore, S.J.; Wolpert, D.M. Abnormalities in the awareness and control of action. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1771–1788. [Google Scholar] [CrossRef]
- Haggard, P. Sense of agency in the human brain. Nat. Rev. Neurosci. 2017, 18, 196–207. [Google Scholar] [CrossRef]
- Gallagher, S. Philosophical conceptions of the self: Implications for cognitive science. Trends Cogn. Sci. 2000, 4, 14–21. [Google Scholar] [CrossRef]
- Sato, A.; Yasuda, A. Illusion of sense of self-agency: Discrepancy between the predicted and actual sensory consequences of actions modulates the sense of self-agency, but not the sense of self-ownership. Cognition 2005, 94, 241–255. [Google Scholar] [CrossRef]
- Shimada, S.; Qi, Y.; Hiraki, K. Detection of visual feedback delay in active and passive self-body movements. Exp. Brain Res. 2010, 201, 359–364. [Google Scholar] [CrossRef]
- Castiello, U.; Paulignan, Y.; Jeannerod, M. Temporal dissociation of motor responses and subjective awareness: A study in normal subjects. Brain 1991, 114, 2639–2655. [Google Scholar] [CrossRef]
- Fourneret, P.; Jeannerod, M. Limited conscious monitoring of motor performance in normal subjects. Neuropsychologia 1998, 36, 1133–1140. [Google Scholar] [CrossRef]
- Wolpe, N.; Rowe, J.B. Beyond the “urge to move”: Objective measures for the study of agency in the post-Libet era. Front. Hum. Neurosci. 2014, 20, 450. [Google Scholar] [CrossRef]
- Kelso, J.A.S. On the self-organizing origins of agency. Trends Cogn. Sci. 2016, 20, 490–499. [Google Scholar] [CrossRef]
- Bandura, A. Social cognitive theory: An agentic perspective. Annu. Rev. Psychol. 2001, 52, 1–26. [Google Scholar] [CrossRef]
- Wen, W.; Yamashita, A.; Asama, H. The influence of goals on sense of control. Conscious. Cogn. 2015, 37, 83–90. [Google Scholar] [CrossRef]
- Haggard, P.; Clark, S.; Kalogeras, J. Voluntary action and conscious awareness. Nat. Neurosci. 2002, 5, 382–385. [Google Scholar] [CrossRef]
- Moore, J.W.; Obhi, S.S. Intentional binding and the sense of agency: A review. Conscious. Cogn. 2012, 21, 546–561. [Google Scholar] [CrossRef]
- Engbert, K.; Wohlschläger, A.; Thomas, R.; Haggard, P. Agency, subjective time, and other minds. J. Exp. Psychol. Hum. Percept. Perform. 2007, 33, 1261–1268. [Google Scholar] [CrossRef]
- Moore, J.W.; Wegner, D.M.; Haggard, P. Modulating the sense of agency with external cues. Conscious. Cogn. 2009, 18, 1056–1064. [Google Scholar] [CrossRef]
- Takahata, K.; Takahashi, H.; Maeda, T.; Umeda, S.; Suhara, T.; Mimura, M.; Kato, M. It’s not my fault: Postdictive modulation of intentional binding by monetary gains and losses. PLoS ONE 2012, 7, e53421. [Google Scholar] [CrossRef]
- Moore, J.W.; Schneider, S.A.; Schwingenschuh, P.; Moretto, G.; Bhatia, K.P.; Haggard, P. Dopaminergic medication boosts action-effect binding in Parkinson’s disease. Neuropsychologia. 2010, 48, 1125–1132. [Google Scholar] [CrossRef]
- Morioka, S.; Hayashida, K.; Nishi, Y.; Negi, S.; Nishi, Y.; Osumi, M.; Nobusako, S. Changes in intentional binding effect during a novel perceptual-motor task. PeerJ 2018, 6, e6066. [Google Scholar] [CrossRef]
- Poonian, S.K.; Cunnington, R. Intentional binding in self-made and observed actions. Exp. Brain Res. 2013, 229, 419–427. [Google Scholar] [CrossRef]
- Poonian, S.K.; McFadyen, J.; Ogden, J.; Cunnington, R. Implicit agency in observed actions: Evidence for N1 suppression of tones caused by self-made and observed actions. J. Cogn. Neurosci. 2015, 27, 752–764. [Google Scholar] [CrossRef]
- Berberian, B.; Sarrazin, J.C.; Blaye, P.L.; Haggard, P. Automation technology and sense of control: A window on human agency. PLoS ONE 2012, 7, e34075. [Google Scholar] [CrossRef]
- Humphreys, G.R.; Buehner, M.J. Magnitude estimation reveals temporal binding at super-second intervals. J. Exp. Psychol. Hum. Percept. Perform. 2009, 35, 1542–1549. [Google Scholar] [CrossRef]
- Kristjansson, A.; Campana, G. Where perception meets memory: A review of repetition priming in visual search tasks. Atten. Percept. Psychophys. 2010, 72, 5–18. [Google Scholar] [CrossRef]
- Fuggetta, G.; Lanfranchi, S.; Campana, G. Attention has memory: Priming for the size of the attentional focus. Spat Vis. 2009, 22, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Di Costa, S.; Théro, H.; Chambon, V.; Haggard, P. Try and try again: Post-error boost of an implicit measure of agency. Q. J. Exp. Psychol. 2017, 71, 1584–1595. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, Y.; Otani, T.; Morioka, S. Dynamic relationship between sense of agency and post-stroke sensorimotor deficits: A longitudinal case study. Brain Sci. 2020, 10, 294. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashida, K.; Nishi, Y.; Masuike, A.; Morioka, S. Intentional Binding Effects in the Experience of Noticing the Regularity of a Perceptual-Motor Task. Brain Sci. 2020, 10, 659. https://doi.org/10.3390/brainsci10090659
Hayashida K, Nishi Y, Masuike A, Morioka S. Intentional Binding Effects in the Experience of Noticing the Regularity of a Perceptual-Motor Task. Brain Sciences. 2020; 10(9):659. https://doi.org/10.3390/brainsci10090659
Chicago/Turabian StyleHayashida, Kazuki, Yuki Nishi, Akihiro Masuike, and Shu Morioka. 2020. "Intentional Binding Effects in the Experience of Noticing the Regularity of a Perceptual-Motor Task" Brain Sciences 10, no. 9: 659. https://doi.org/10.3390/brainsci10090659
APA StyleHayashida, K., Nishi, Y., Masuike, A., & Morioka, S. (2020). Intentional Binding Effects in the Experience of Noticing the Regularity of a Perceptual-Motor Task. Brain Sciences, 10(9), 659. https://doi.org/10.3390/brainsci10090659